Abiotic Degradation of the Toxin Simplexin by Soil Collected from a Pimelea-Infested Paddock

 , , ,
, , , 
Abstract

1. Introduction
2. Results and Discussion
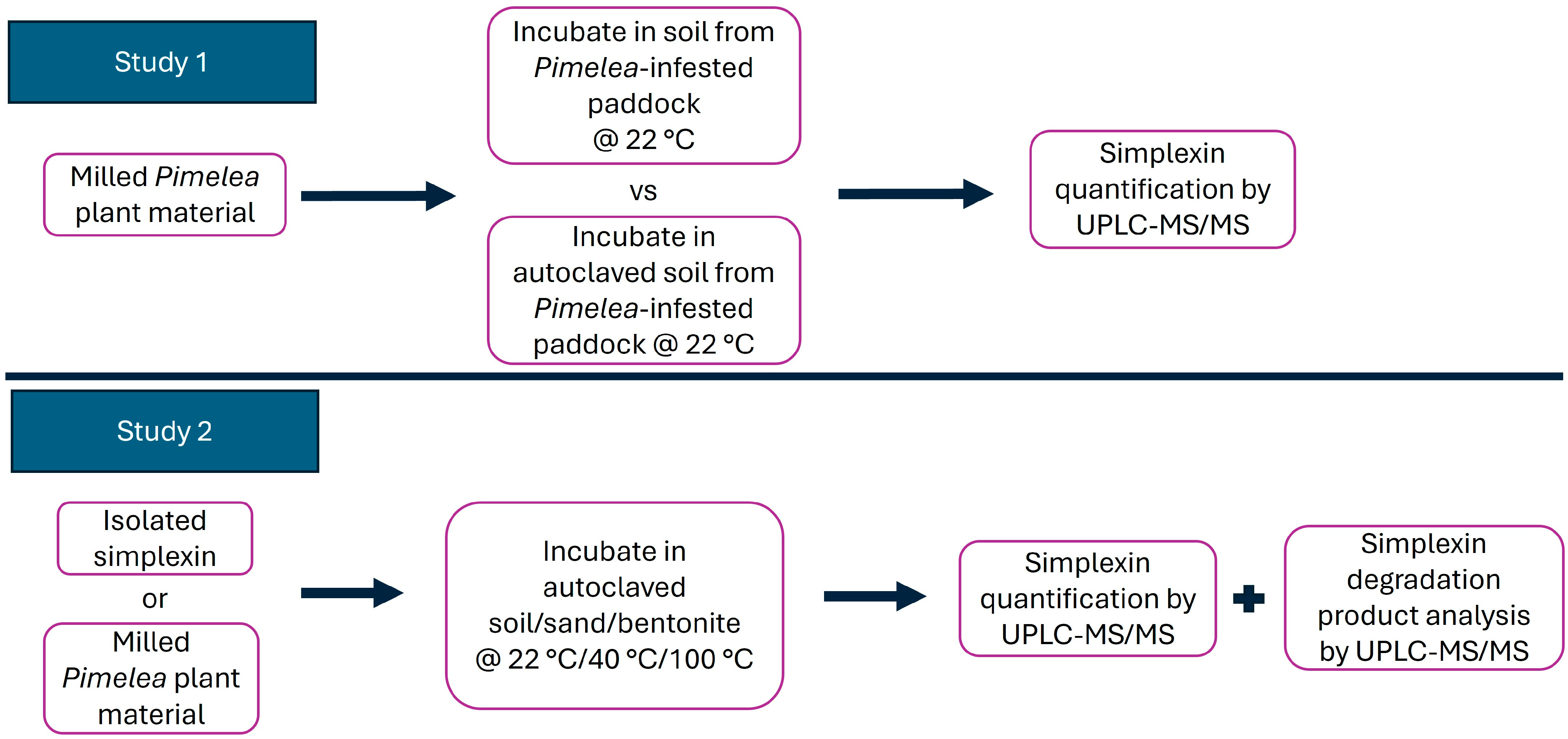
2.1. Incubation of Milled P. trichostachya with Field-Collected Soil
2.2. Simplexin Degradation by Heat
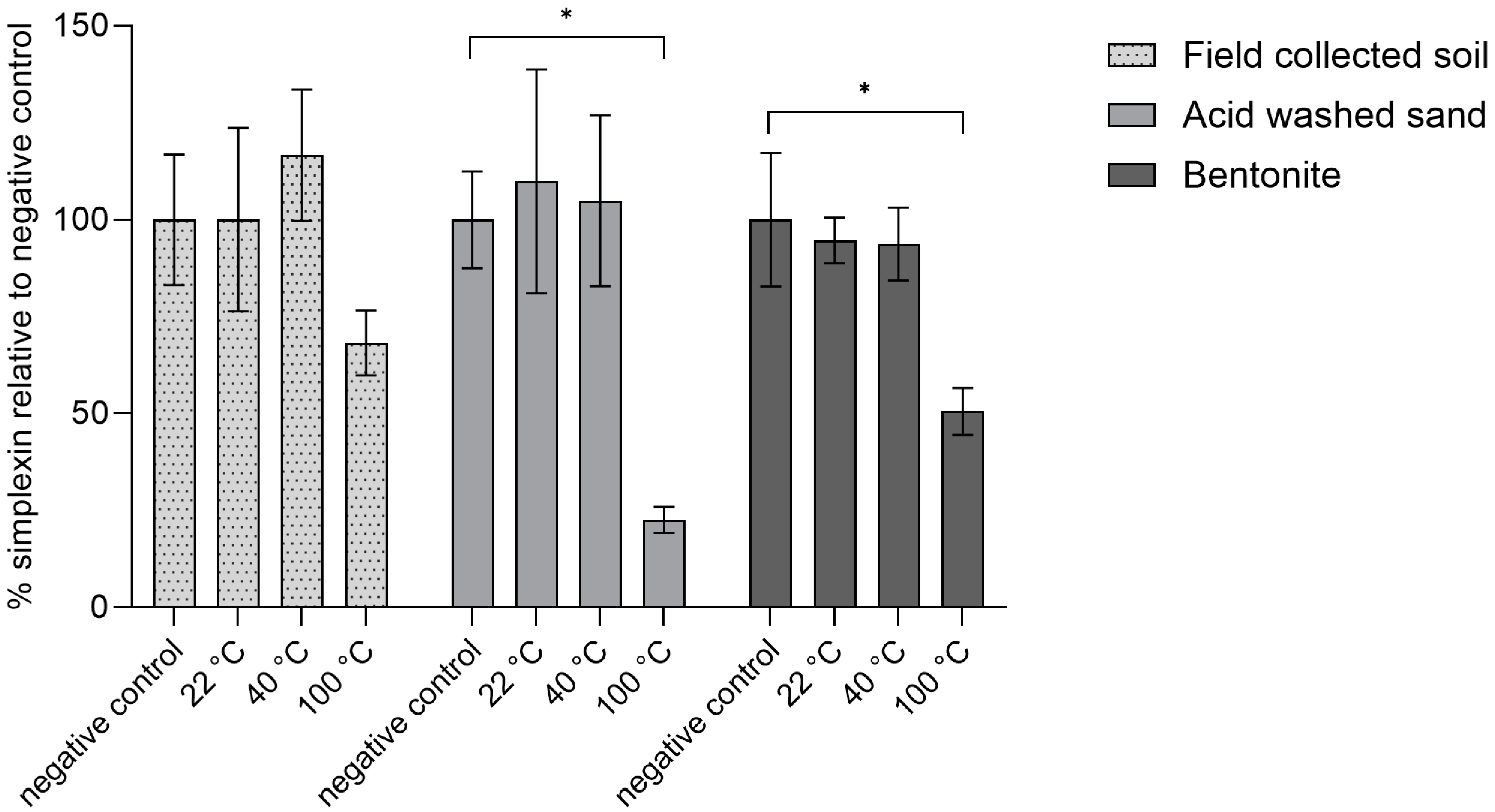
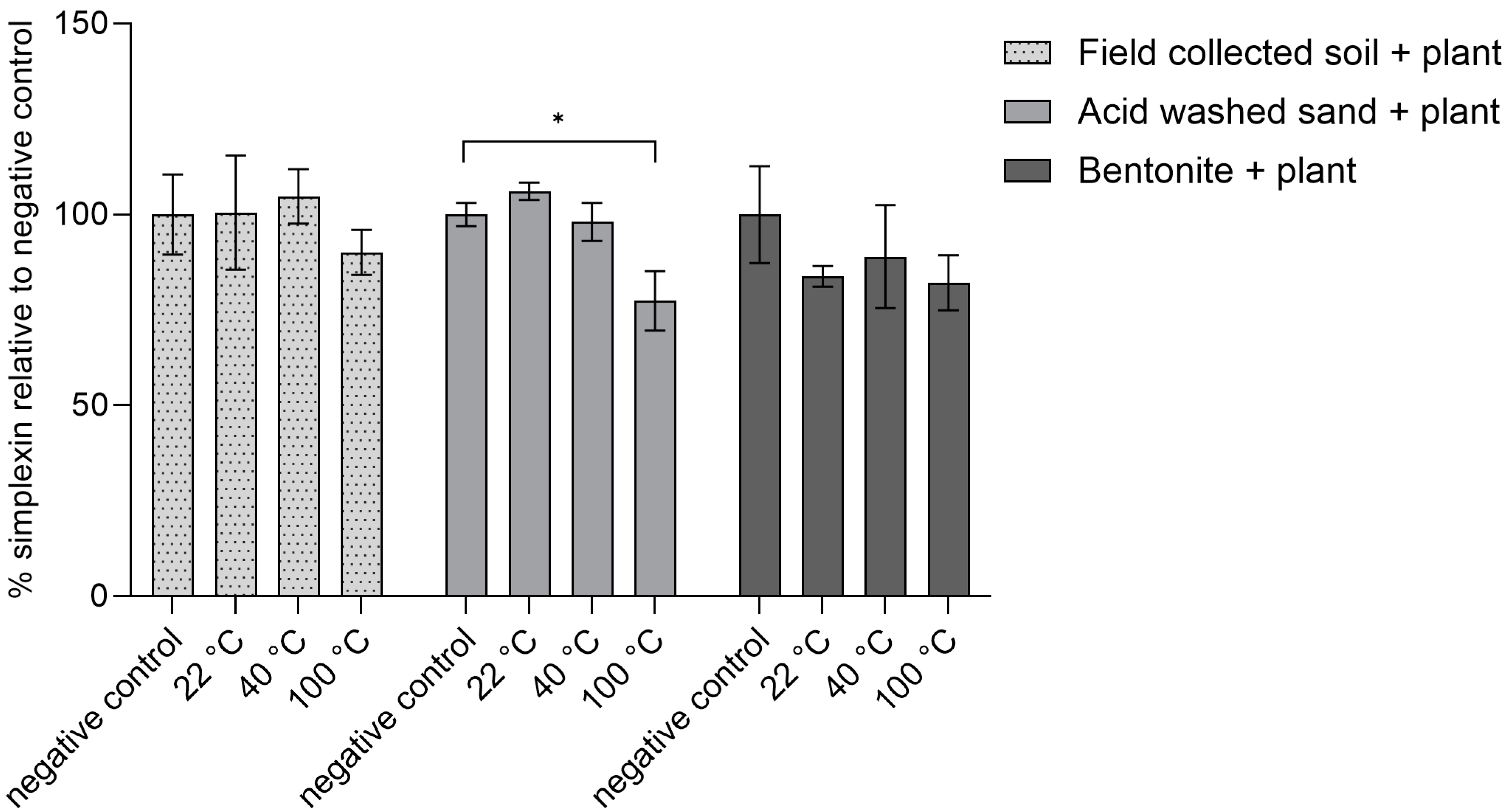
2.2.1. Heating Effect on Isolated Simplexin Preadsorbed to Sand, Bentonite and Field-Collected Soil
2.2.2. Heating Effect on Milled P. trichostachya Mixed with Field-Collected Soil, Sand and Bentonite
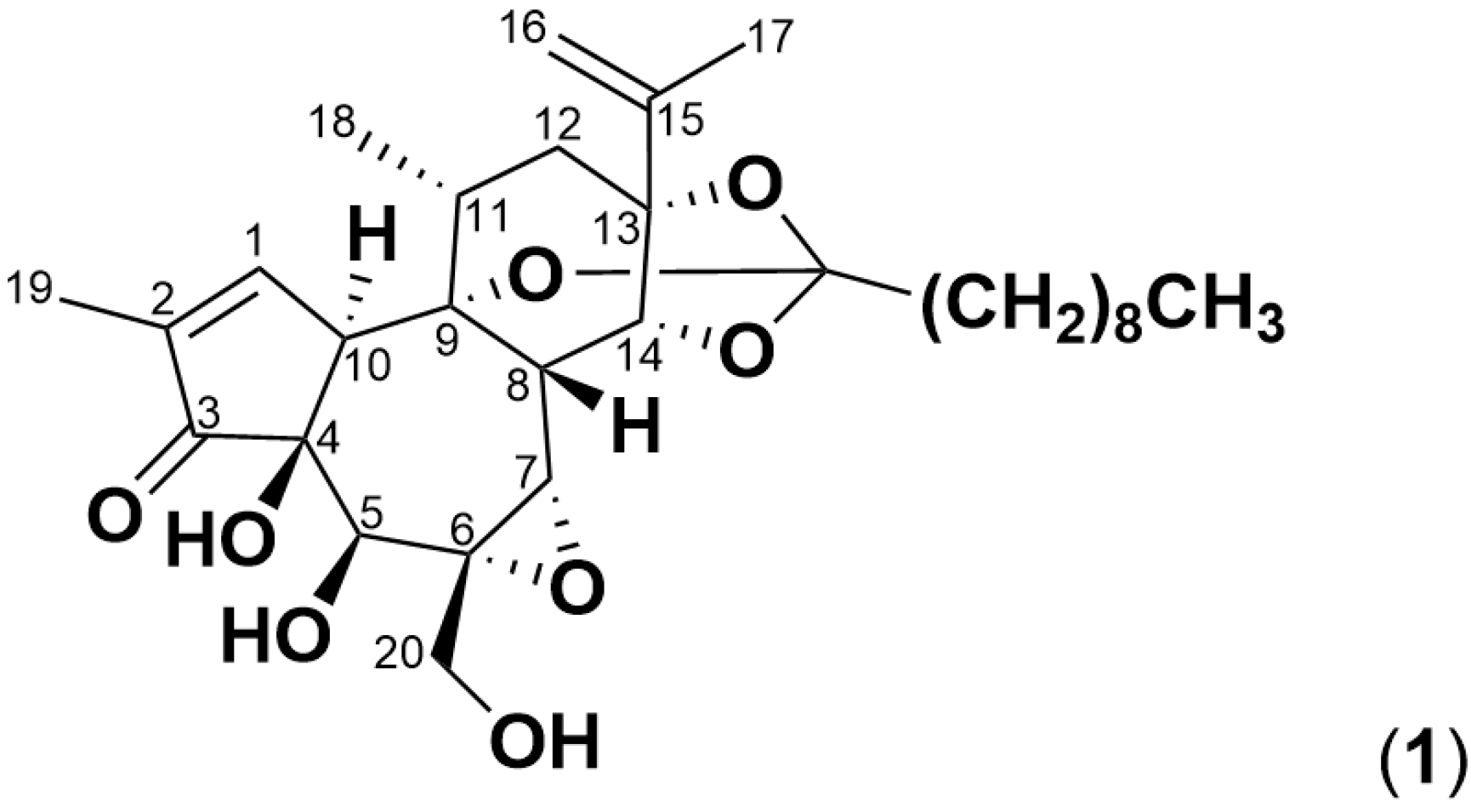
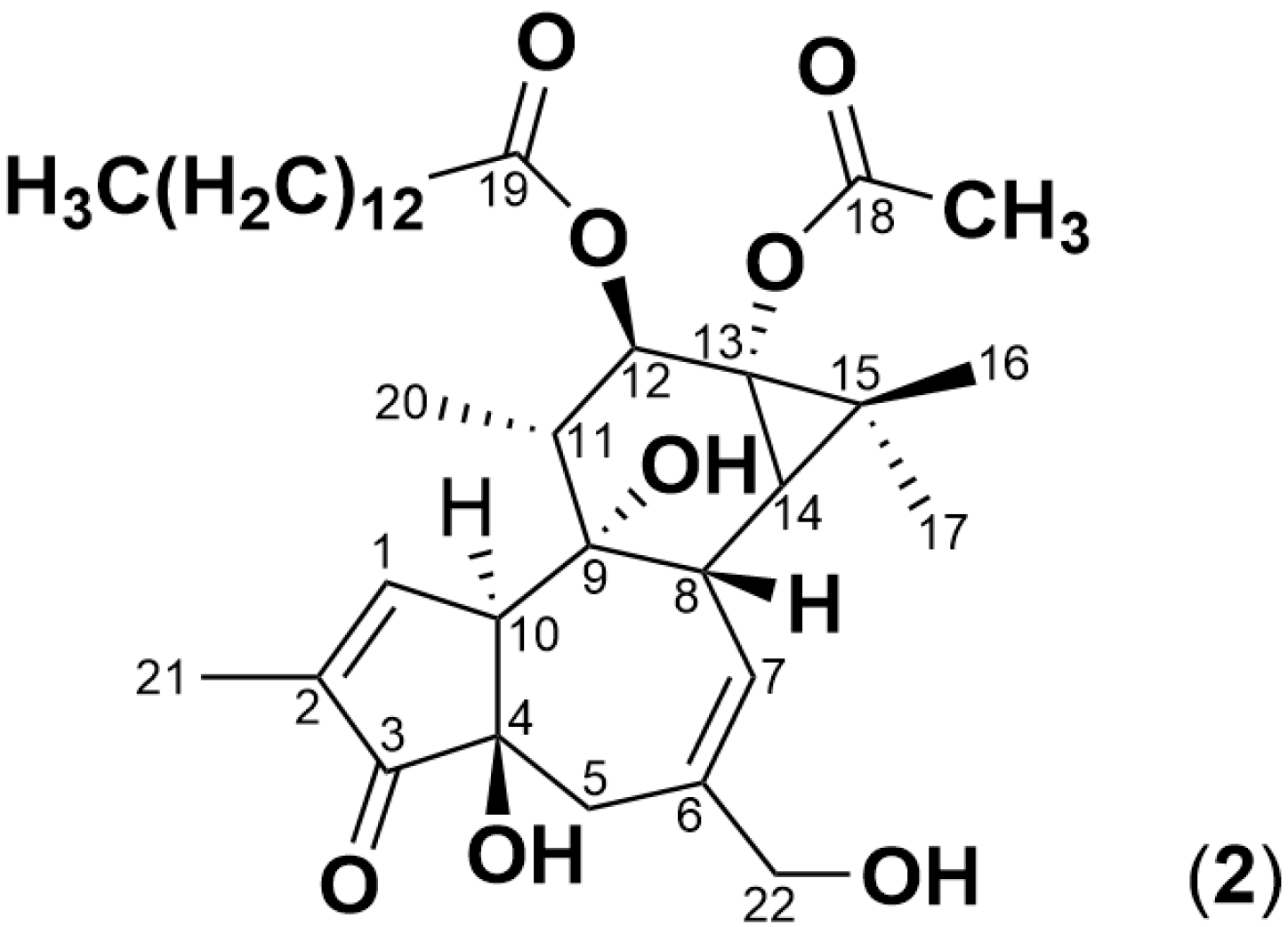
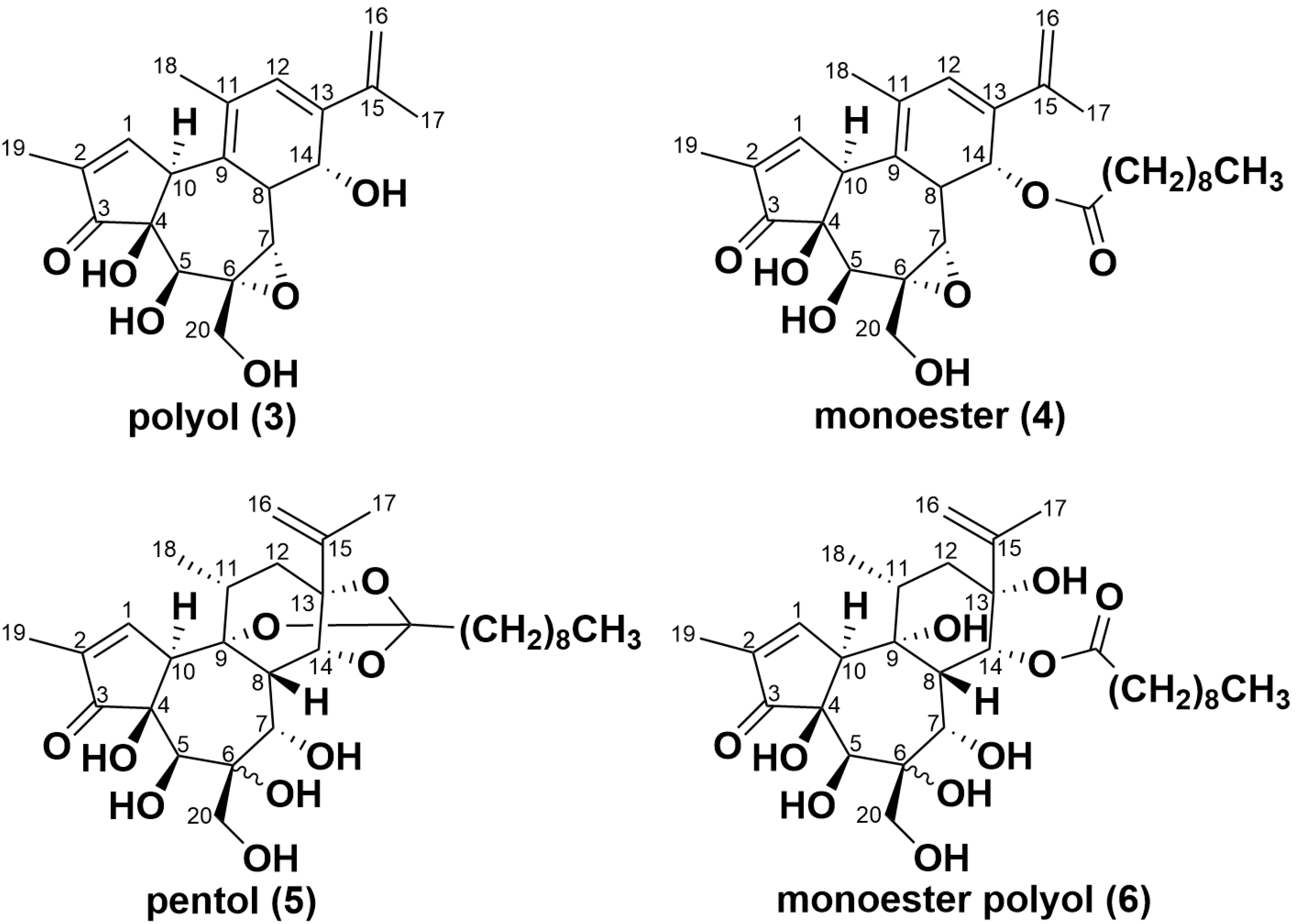
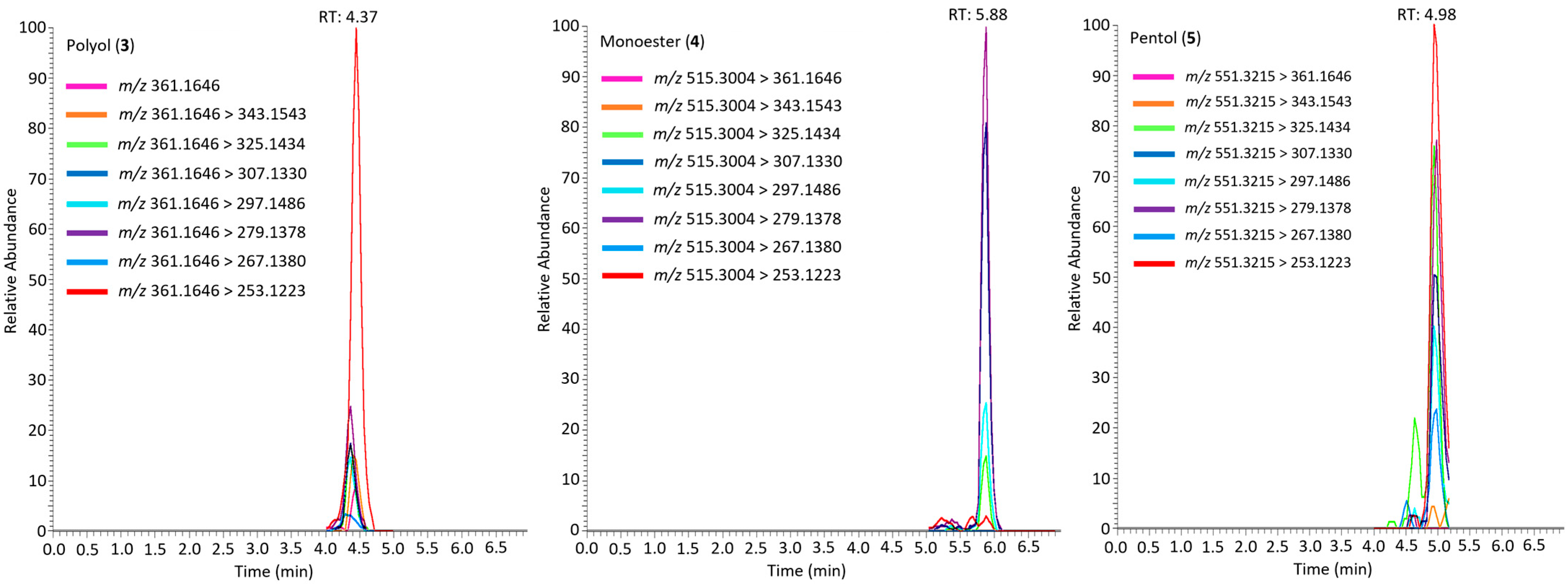
2.3. Simplexin Breakdown Products
3. Conclusions
4. Materials and Methods
4.1. General Experimental Materials
4.2. Sample Material Collection for the Degradation Study
4.2.1. Soil
4.2.2. Plant Material
4.3. P. trichostachya Seed Collection and Simplexin Isolation from Seeds
4.4. Incubation of Milled P. trichostachya on Field-Collected Soil
4.5. Simplexin Degradation by Heat
4.5.1. Heating Effect of Isolated Simplexin Preadsorbed to Sand, Bentonite and Field-Collected Soil
4.5.2. Heating Effect on Milled P. trichostachya Mixed with Field-Collected Soil, Sand and Bentonite
4.6. Simplexin Quantification in Plant and Preadsorbed Material Samples
4.7. Simplexin Degradation Product HRAMS Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, M.; Silcock, R.; Ossedryver, S.; Milson, J.; Chow, S. Understanding Pimelea Poisoning of Cattle; Department of Employment, Economic Development and Innovation: Brisbane, QLD, Australia, 2009. [Google Scholar]
- Chow, S.; Fletcher, M.T.; McKenzie, R.A. Analysis of daphnane orthoesters in poisonous Australian Pimelea species by liquid chromatography−tandem mass spectrometry. J. Agric. Food. Chem. 2010, 58, 7482–7487. [Google Scholar] [CrossRef] [PubMed]
- Roberts, H.B.; McClure, T.J.; Ritchie, E.; Taylor, W.C.; Freeman, P.W. The isolation and structure of the toxin of Pimelea simplex responsible for St. George disease of cattle. Aust. Vet. J. 1975, 51, 325–326. [Google Scholar] [CrossRef] [PubMed]
- Saleem, R.; Bajwa, A.A.; Campbell, S.; Fletcher, M.T.; Kalaipandian, S.; Adkins, S.W. Poisonous Plants of the Genus Pimelea: A Menace for the Australian Livestock Industry. Toxins 2023, 15, 374. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Chow, S.; Ossedryver, S.M. Effect of increasing low-dose simplexin exposure in cattle consuming Pimelea trichostachya. J. Agric. Food. Chem. 2014, 62, 7402–7406. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, M.; Chow, K.; Silcock, R.; Milson, J. LC/MS/MS analysis of the daphnane orthoester simplexin in poisonous Pimelea species of Australian rangelands. In Poisoning by Plants, Mycotoxins and Related Toxins; Riet-Correea, F., Pfister, J., Schild, A., Wierenga, T., Eds.; CABI: Wallingford, UK, 2011; pp. 550–556. [Google Scholar]
- Hormisch, D.; Brost, I.; Kohring, G.-W.; Giffhorn, F.; Kroppenstedt, R.; Stackebradt, E.; Färber, P.; Holzapfel, W. Mycobacterium fluoranthenivorans sp. nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. Syst. Appl. Microbiol. 2004, 27, 653–660. [Google Scholar] [CrossRef]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [CrossRef]
- Megharaj, M.; Garthwaite, I.; Thiele, J. Total biodegradation of the oestrogenic mycotoxin zearalenone by a bacterial culture. Lett. Appl. Microbiol. 1997, 24, 329–333. [Google Scholar] [CrossRef]
- Abid, M.; Fayolle, L.; Edel-Hermann, V.; Gautheron, N.; Héraud, C.; Leplat, J.; Steinberg, C. Fate of deoxynivalenol (DON) and impact on the soil microflora and soil fauna. Appl. Soil Ecol. 2021, 162, 103898. [Google Scholar] [CrossRef]
- He, W.-J.; Shi, M.-M.; Yang, P.; Huang, T.; Yuan, Q.-S.; Yi, S.-Y.; Wu, A.-B.; Li, H.-P.; Gao, C.-B.; Zhang, J.-B. Novel soil bacterium strain Desulfitobacterium sp. PGC-3-9 detoxifies trichothecene mycotoxins in wheat via de-epoxidation under aerobic and anaerobic conditions. Toxins 2020, 12, 363. [Google Scholar] [CrossRef]
- Mahajan, R.; Verma, S.; Chatterjee, S. Biodegradation of organophosphorus pesticide profenofos by the bacterium Bacillus sp. PF1 and elucidation of initial degradation pathway. Environ. Technol. 2021, 44, 492–500. [Google Scholar] [CrossRef]
- Wang, G.; Chen, X.; Yue, W.; Zhang, H.; Li, F.; Xiong, M. Microbial degradation of acetamiprid by Ochrobactrum sp. D-12 isolated from contaminated soil. PLoS ONE 2013, 8, e82603. [Google Scholar] [CrossRef] [PubMed]
- Devappa, R.K.; Makkar, H.P.; Becker, K. Biodegradation of Jatropha curcas phorbol esters in soil. J. Sci. Food Agric. 2010, 90, 2090–2097. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S.; Francis, G.; Becker, K. Phorbol esters: Structure, biological activity, and toxicity in animals. Int. J. Toxicol. 2007, 26, 279–288. [Google Scholar] [CrossRef]
- Hungerford, N.L.; Ouwerkerk, D.; Gilbert, R.A.; Loh, Z.H.; Gordon, R.J.; Silva, L.F.P.; Fletcher, M.T. A feeding trial to investigate strategies to mitigate the impacts of Pimelea poisoning in Australian cattle. J. Agric. Food. Chem. 2024, 72, 15572–15585. [Google Scholar] [CrossRef] [PubMed]
- Loh, Z.H.; Ouwerkerk, D.; Klieve, A.V.; Hungerford, N.; Fletcher, M.T. Toxin degradation by rumen microorganisms: A review. Toxins 2020, 12, 664. [Google Scholar] [CrossRef]
- Atlas of Living Australia. Pimelea trichostachya Lindl. Available online: https://bie.ala.org.au/species/https://id.biodiversity.org.au/node/apni/2891911# (accessed on 18 February 2025).
- Ndiaye, S.; Zhang, M.; Fall, M.; Ayessou, N.M.; Zhang, Q.; Li, P. Current review of mycotoxin biodegradation and bioadsorption: Microorganisms, mechanisms, and main important applications. Toxins 2022, 14, 729. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J. Soil moisture as a factor affecting the microbiological and biochemical activity of soil. Plant Soil Environ. 2016, 62, 250–255. [Google Scholar] [CrossRef]
- Salazar-Villegas, A.; Blagodatskaya, E.; Dukes, J.S. Changes in the size of the active microbial pool explain short-term soil respiratory responses to temperature and moisture. Front. Microbiol. 2016, 7, 524. [Google Scholar] [CrossRef]
- Vischetti, C.; Esposito, A. Degradation and transformation of a potential natural herbicide in three soils. J. Agric. Food. Chem. 1999, 47, 3901–3904. [Google Scholar] [CrossRef]
- Moran, J. How the rumen works. In Tropical Dairy Farming: Feeding Managment for Small Holder Dairy Farmers in the Humid Tropics; Landlinks Press: Collingwood, Australia, 2005; pp. 41–49. [Google Scholar]
- Hennessy, R.C.; Jørgensen, N.O.; Scavenius, C.; Enghild, J.J.; Greve-Poulsen, M.; Sørensen, O.B.; Stougaard, P. A screening method for the isolation of bacteria capable of degrading toxic steroidal glycoalkaloids present in potato. Front. Microbiol. 2018, 9, 2648. [Google Scholar] [CrossRef]
- Hennessy, R.C.; Nielsen, S.D.; Greve-Poulsen, M.; Larsen, L.B.; Sørensen, O.B.; Stougaard, P. Discovery of a bacterial gene cluster for deglycosylation of toxic potato steroidal glycoalkaloids α-chaconine and α-solanine. J. Agric. Food Chem. 2020, 68, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Loh, Z.H. Mitigating the Effects of the Toxin Simplexin in Pimelea Poisoning of Cattle by Development of a Microbial Probiotic. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 2022. [Google Scholar]
- Peivasteh-Roudsari, L.; Pirhadi, M.; Shahbazi, R.; Eghbaljoo-Gharehgheshlaghi, H.; Sepahi, M.; Mirza Alizadeh, A.; Tajdar-oranj, B.; Jazaeri, S. Mycotoxins: Impact on health and strategies for prevention and detoxification in the food chain. Food Rev. Int. 2021, 38, 193–224. [Google Scholar] [CrossRef]
- Tripathi, S.; Mishra, H. Studies on the efficacy of physical, chemical and biological aflatoxin B1 detoxification approaches in red chilli powder. Int. J. Food Saf. Nutr. Public Health Technol. 2009, 2, 69–77. [Google Scholar] [CrossRef]
- Anandan, S.; Kumar, G.A.; Ghosh, J.; Ramachandra, K. Effect of different physical and chemical treatments on detoxification of ricin in castor cake. Anim. Feed Sci. Technol. 2005, 120, 159–168. [Google Scholar] [CrossRef]
- Mrkajic, N.S.; Hama, J.R.; Strobel, B.W.; Hansen, H.C.B.; Rasmussen, L.H.; Pedersen, A.-K.; Christensen, S.C.; Hedegaard, M.J. Removal of phytotoxins in filter sand used for drinking water treatment. Water Res. 2021, 205, 117610. [Google Scholar] [CrossRef]
- Armor, J.N.; Zambri, P.M. Silica as an oxidation catalyst. J. Catal. 1982, 73, 57–65. [Google Scholar] [CrossRef]
- Rodrigues, M. Physical and catalytic characterization of smectites from Boa-Vista, Paraíba, Brazil. Cerâmica 2003, 49, 146–150. [Google Scholar] [CrossRef]
- Bouguerra Neji, S.; Trabelsi, M.; Frikha, M.H. Esterification of fatty acids with short-chain alcohols over commercial acid clays in a semi-continuous reactor. Energies 2009, 2, 1107–1117. [Google Scholar] [CrossRef]
- Tennant, D.; Scholz, G.; Dixon, J.; Purdie, B. Physical and chemical characteristics of duplex soils and their distribution in the south-west of Western Australia. Aust. J. Exp. Agric. 1992, 32, 827–843. [Google Scholar] [CrossRef]
- Loh, Z.H.; Hungerford, N.L.; Ouwerkerk, D.; Klieve, A.V.; Fletcher, M.T. Identification of acid hydrolysis metabolites of the Pimelea toxin simplexin for targeted UPLC-MS/MS analysis. Toxins 2023, 15, 551. [Google Scholar] [CrossRef]
- Freeman, P.; Ritchie, E.; Taylor, W. The constituents of Australian Pimelea spp. I. The isolation and structure of the toxin of Pimelea simplex and P. trichostachya form B responsible for St. George disease of cattle. Aust. J. Chem. 1979, 32, 2495–2506. [Google Scholar] [CrossRef]
- Smułek, W.; Kaczorek, E. Factors influencing the bioavailability of organic molecules to bacterial cells—A mini-review. Molecules 2022, 27, 6579. [Google Scholar] [CrossRef] [PubMed]
- Australian Bureau of Meteorology. Daily Maximum Temperature. Available online: http://www.bom.gov.au/jsp/ncc/cdio/weatherData/av?p_nccObsCode=36&p_display_type=dataFile&p_stn_num=044010 (accessed on 9 July 2024).
- Sakata, K.; Kawazu, K.; Mitsui, T. Studies on a piscicidal constituent of Hura crepitans. Part II. Chemical structure of huratoxin. Agr. Biol. Chem. 1971, 35, 2113–2126. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simplexin Concentration (mg/kg Plant Material) | ||||||
|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 2 | Day 3 | Day 6 | Day 7 | |
| Sterile field-collected soil + Pimelea | 85.3 ± 19.2 a | 96.4 ± 10.7 a | 101.1 ± 29.3 a | 100.1 ± 25.0 a | 104.2 ± 19.6 a | 117.1 ± 20.1 a |
| Field-collected soil + Pimelea | 72.8 ± 8.1 a | 99.4 ± 6.2 a | 98.5 ± 8.8 a | 100.7 ± 13.7 a | 94.6 ± 23.7 a | 103.5 ± 22.2 a |
| Molecular Formula | Species | Calculated Molecular ion (m/z) [M + H]+ | Compound |
|---|---|---|---|
| C30H44O8 | + H+ | 533.3109 | simplexin (1) |
| C36H56O8 | + H+ | 617.4080 | PMA (2) |
| C20H24O6 | + H+ | 361.1646 | polyol (3) |
| C30H42O7 | + H+ | 515.3003 | monoester (4) |
| C30H46O9 | + H+ | 551.3215 | pentol (5) |
| C30H48O10 | + H+ | 569.3320 | monoester polyol (6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loh, Z.H.; Hungerford, N.L.; Ouwerkerk, D.; Klieve, A.V.; Fletcher, M.T. Abiotic Degradation of the Toxin Simplexin by Soil Collected from a Pimelea-Infested Paddock. Toxins 2025, 17, 124. https://doi.org/10.3390/toxins17030124
Loh ZH, Hungerford NL, Ouwerkerk D, Klieve AV, Fletcher MT. Abiotic Degradation of the Toxin Simplexin by Soil Collected from a Pimelea-Infested Paddock. Toxins. 2025; 17(3):124. https://doi.org/10.3390/toxins17030124
Chicago/Turabian StyleLoh, Zhi Hung, Natasha L. Hungerford, Diane Ouwerkerk, Athol V. Klieve, and Mary T. Fletcher. 2025. "Abiotic Degradation of the Toxin Simplexin by Soil Collected from a Pimelea-Infested Paddock" Toxins 17, no. 3: 124. https://doi.org/10.3390/toxins17030124
APA StyleLoh, Z. H., Hungerford, N. L., Ouwerkerk, D., Klieve, A. V., & Fletcher, M. T. (2025). Abiotic Degradation of the Toxin Simplexin by Soil Collected from a Pimelea-Infested Paddock. Toxins, 17(3), 124. https://doi.org/10.3390/toxins17030124