Metal Ions and Chemical Modification Reagents Inhibit the Enzymatic Activity of Lecithin-Dependent Hemolysin from Vibrio parahaemolyticus

 ,
, 
 , and
, and 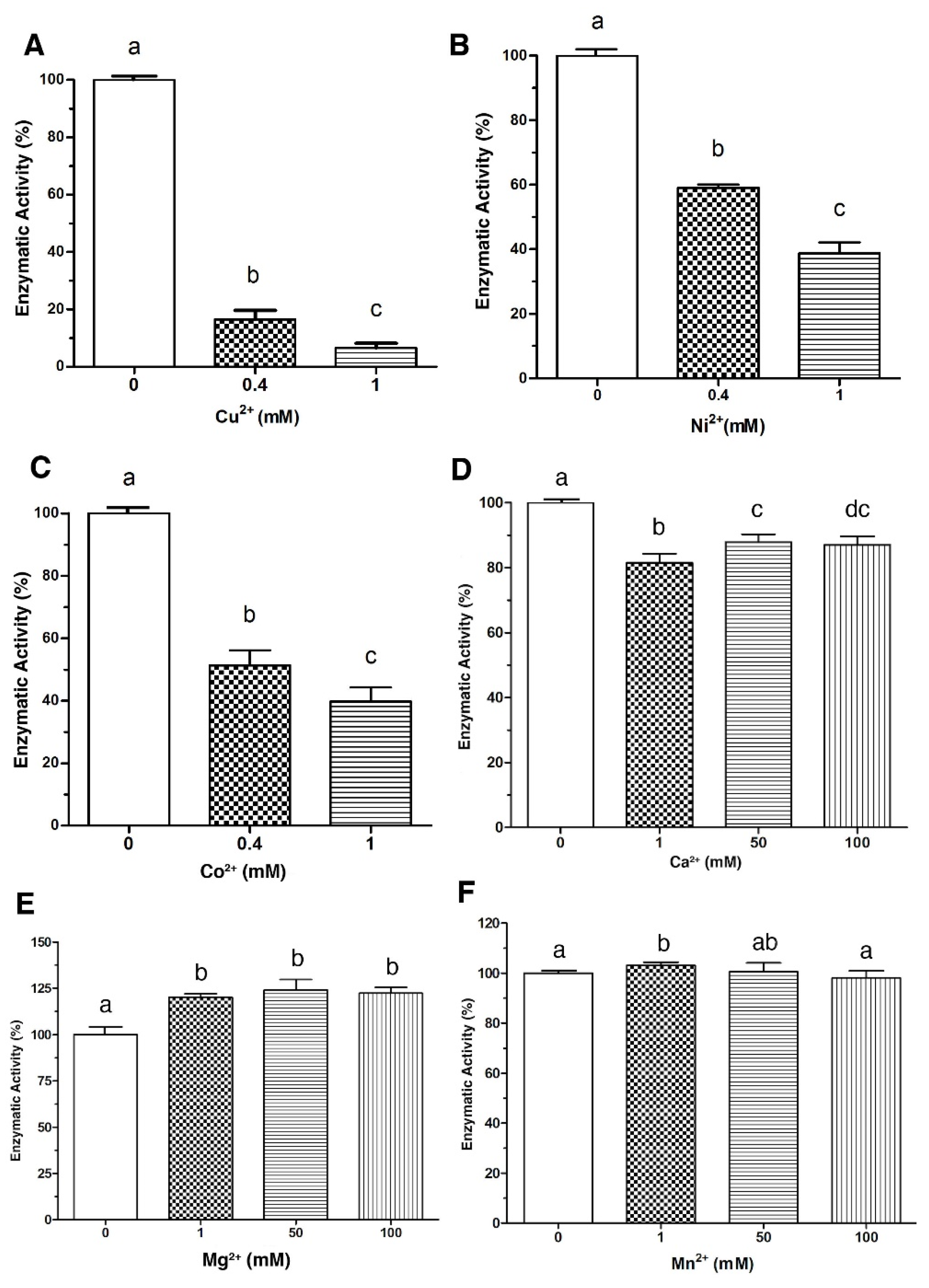{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
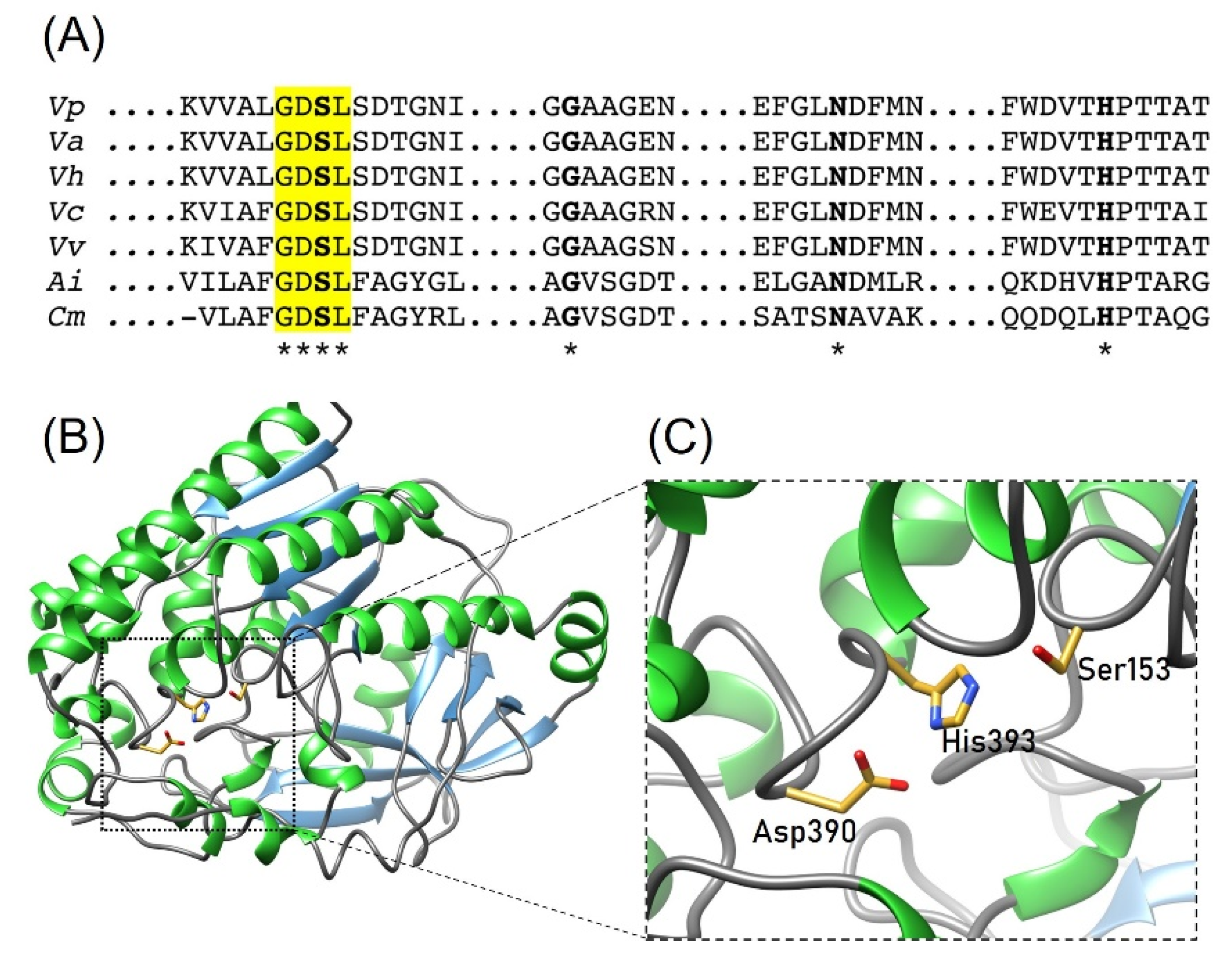{kind=link}
Abstract
1. Introduction
2. Results
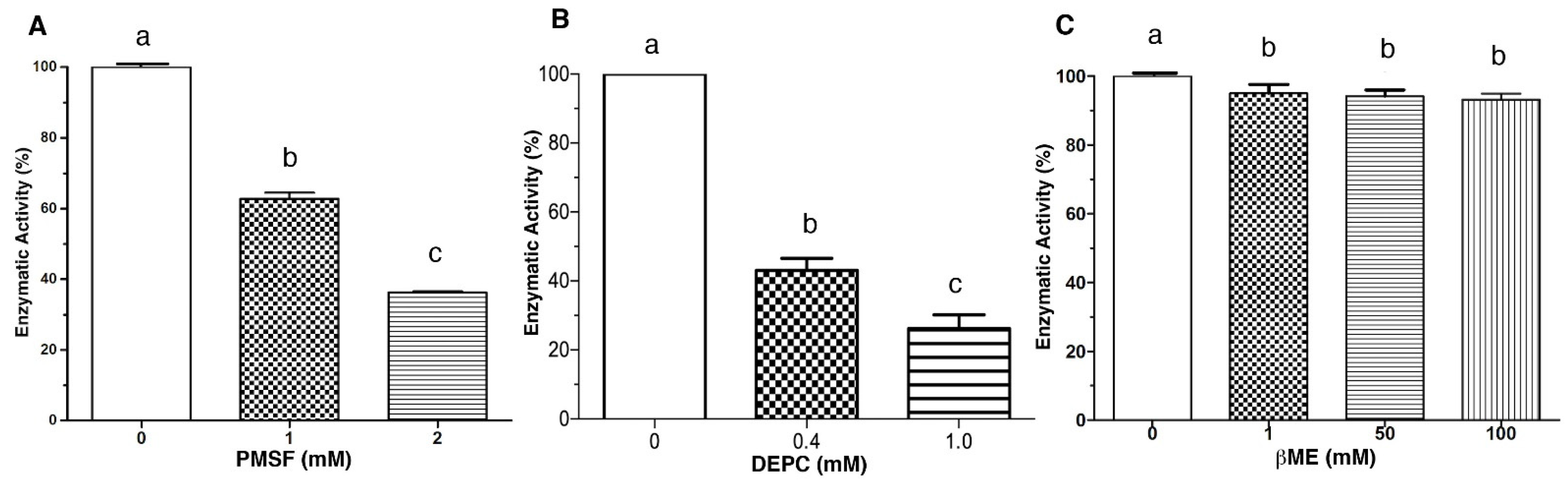
2.1. Effect of Metal Ions and Chemical Reagents on the Enzymatic Activity of LDH
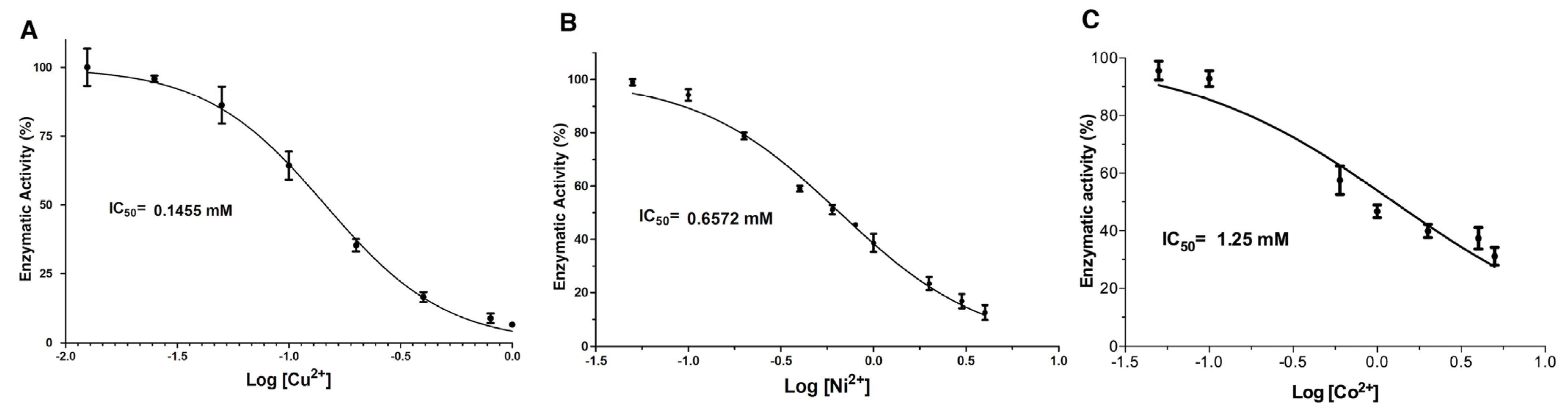
2.2. Dose-Response Analysis of the Inhibition Effect Caused by Metal Ions and Chemical Reagents
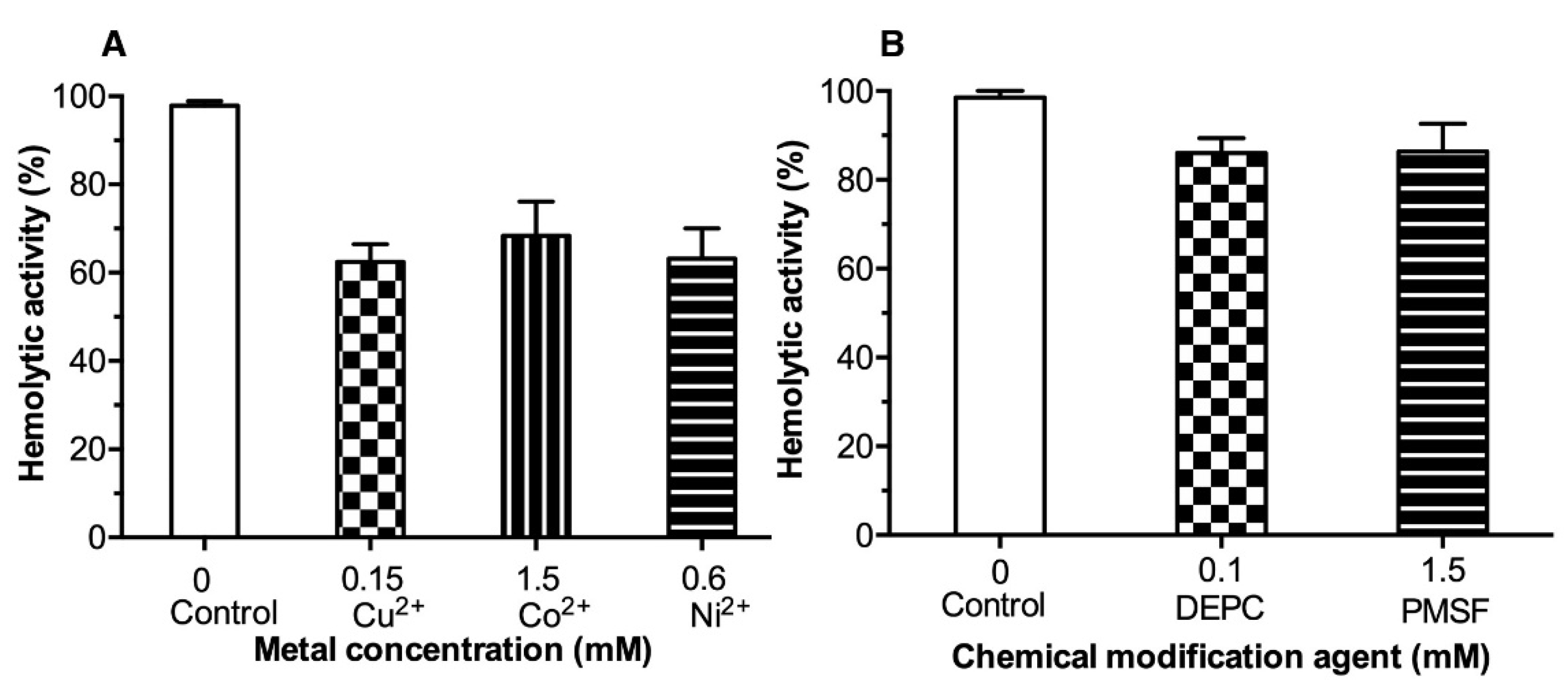
2.3. Effect of Metal Ions and Chemical Reagents on the LDH Hemolytic Activity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. LDH Protein Expression and Purification
5.2. Enzymatic Activity and Hemolytic Assay
5.3. Inhibition of Enzymatic and Hemolytic Activity by Metal Ions and Chemical Modification Reagents
5.4. Dose-Response Analysis
5.5. Structural Modeling
5.6. Statistical Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Rajeev, R.; Adithya, K.K.; Kiran, G.S.; Selvin, J. Healthy microbiome: A key to successful and sustainable shrimp aquaculture. Rev. Aquac. 2021, 13, 238–258. [Google Scholar] [CrossRef]
- Flegel, T.W. A future vision for disease control in shrimp aquaculture. J. World Aquac. Soc. 2019, 50, 249–266. [Google Scholar] [CrossRef]
- Lai, H.-C.; Ng, T.H.; Ando, M.; Lee, C.-T.; Chen, I.T.; Chuang, J.-C.; Mavichak, R.; Chang, S.-H.; Yeh, M.-D.; Chiang, Y.-A.; et al. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp. Fish Shellfish. Immunol. 2015, 47, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, X.; Fan, H.; Li, Y.; Hu, Q.; Yang, R.; Cui, Y. Genetic diversity, virulence factors and farm-to-table spread pattern of Vibrio parahaemolyticus food-associated isolates. Food Microbiol. 2019, 84, 103270. [Google Scholar] [CrossRef]
- Wang, R.; Zhong, Y.; Gu, X.; Yuan, J.; Saeed, A.F.; Wang, S. The pathogenesis, detection, and prevention of Vibrio parahaemolyticus. Front. Microbiol. 2015, 6, 144. [Google Scholar] [CrossRef]
- Ghenem, L.; Elhadi, N.; Alzahrani, F.; Nishibuchi, M. Vibrio parahaemolyticus: A review on distribution, pathogenesis, virulence determinants and epidemiology. Saudi J. Med. Med Sci. 2017, 5, 93–103. [Google Scholar] [CrossRef]
- Gotoh, K.; Kodama, T.; Hiyoshi, H.; Izutsu, K.; Park, K.-S.; Dryselius, R.; Akeda, Y.; Honda, T.; Iida, T. Bile acid-induced virulence gene expression of Vibrio parahaemolyticus reveals a novel therapeutic potential for bile acid sequestrants. PLoS ONE 2010, 5, e13365. [Google Scholar] [CrossRef]
- Joshi, J.; Srisala, J.; Truong, V.H.; Chen, I.T.; Nuangsaeng, B.; Suthienkul, O.; Lo, C.F.; Flegel, T.W.; Sritunyalucksana, K.; Thitamadee, S. Variation in Vibrio parahaemolyticus isolates from a single Thai shrimp farm experiencing an outbreak of acute hepatopancreatic necrosis disease (AHPND). Aquaculture 2014, 428–429, 297–302. [Google Scholar] [CrossRef]
- Jia, A.; Woo, N.Y.; Zhang, X.-H. Expression, purification, and characterization of thermolabile hemolysin (TLH) from Vibrio alginolyticus. Dis. Aquat. Org. 2010, 90, 121–127. [Google Scholar] [CrossRef]
- Chen, A.J.; Hasan, N.A.; Haley, B.J.; Taviani, E.; Tarnowski, M.; Brohawn, K.; Johnson, C.N.; Colwell, R.R.; Huq, A. Characterization of pathogenic Vibrio parahaemolyticus from the Chesapeake bay, Maryland. Front. Microbiol. 2017, 8, 2460. [Google Scholar] [CrossRef]
- Taniguchi, H.; Hirano, H.; Kubomura, S.; Higashi, K.; Mizuguchi, Y. Comparison of the nucleotide sequences of the genes for the thermostable direct hemolysin and the thermolabile hemolysin from Vibrio parahaemolyticus. Microb. Pathog. 1986, 1, 425–432. [Google Scholar] [CrossRef]
- Flores-Díaz, M.; Monturiol-Gross, L.; Naylor, C.; Alape-Girón, A.; Flieger, A. Bacterial Sphingomyelinases and Phospholipases as Virulence Factors. Microbiol. Mol. Biol. Rev. 2016, 80, 597–628. [Google Scholar] [CrossRef]
- Vazquez-Morado, L.E.; Robles-Zepeda, R.E.; Ochoa-Leyva, A.; Arvizu-Flores, A.A.; Garibay-Escobar, A.; Castillo-Yañez, F.; Lopez-zavala, A.A. Biochemical characterization and inhibition of thermolabile hemolysin from Vibrio parahaemolyticus by phenolic compounds. PeerJ 2021, 9, e10506. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.C.; Lee, G.-C.; Liaw, Y.-C.; Huang, T.-H.; Shaw, J.-F. GDSL family of serine esterases/lipases. Prog. Lipid Eesearch 2004, 43, 534–552. [Google Scholar] [CrossRef]
- Chakrabarti, P. Geometry of interaction of metal ions with histidine residues in protein structures. Protein Eng. Des. Sel. 1990, 4, 57–63. [Google Scholar] [CrossRef]
- Rulíšek, L.R.; Vondrášek, J. Coordination geometries of selected transition metal ions (Co2+, Ni2+, Cu2+, Zn2+, Cd2+, and Hg2+) in metalloproteins. J. Inorg. Biochem. 1998, 71, 115–127. [Google Scholar] [CrossRef]
- Migliorini, C.; Porciatti, E.; Luczkowski, M.; Valensin, D. Structural characterization of Cu2+, Ni2+ and Zn2+ binding sites of model peptides associated with neurodegenerative diseases. Coord. Chem. Rev. 2012, 256, 352–368. [Google Scholar] [CrossRef]
- Han, J.H.; Lee, J.H.; Choi, Y.H.; Park, J.H.; Choi, T.J.; Kong, I.S. Purification, characterization and molecular cloning of Vibrio fluvialis hemolysin. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2002, 1599, 106–114. [Google Scholar] [CrossRef]
- Miyoshi, S.; Sasahara, K.; Akamatsu, S.; Rahman, M.M.; Katsu, T.; Tomochika, K.; Shinoda, S. Purification and characterization of a hemolysin produced by Vibrio mimicus. Infect. Immun. 1997, 65, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Koffman, B.; Modarress, K.J.; Bashirelahi, N. The effects of various serine protease inhibitors on estrogen receptor steroid binding. J. Steroid Biochem. Mol. Biol. 1991, 38, 569–574. [Google Scholar] [CrossRef]
- Narumi, R.; Yamamoto, T.; Inoue, A.; Arata, T. Substrate-induced conformational changes in sarcoplasmic reticulum Ca2+-ATPase probed by surface modification using diethylpyrocarbonate with mass spectrometry. FEBS Lett. 2012, 586, 3172–3178. [Google Scholar] [CrossRef]
- Shinoda, S.; Matsuoka, H.; Tsuchie, T.; Miyoshi, S.; Yamamoto, S.; Taniguchi, H.; Mizuguchi, Y. Purification and characterization of a lecithin-dependent haemolysin from Escherichia coli transformed by a Vibrio parahaemolyticus gene. J. Gen. Microbiol. 1991, 137, 2705–2711. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pérez-Legaspi, I.A.; Rico-Martínez, R. Phospholipase A2 activity in three species of littoral freshwater rotifers exposed to several toxicants. Environ. Toxicol. Chem. Int. J. 2003, 22, 2349–2353. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, R.; Dubey, R.S. Nickel toxicity inhibits ribonuclease and protease activities in rice seedlings: Protective effects of proline. Plant Growth Regul. 2007, 51, 231–243. [Google Scholar] [CrossRef]
- Maheshwari, R.; Dubey, R.S. Inhibition of ribonuclease and protease activities in germinating rice seeds exposed to nickel. Acta Physiol. Plant. 2008, 30, 863–872. [Google Scholar] [CrossRef]
- Li, Z.; Li, L.; Huo, Y.; Chen, Z.; Zhao, Y.; Huang, J.; Jian, S.; Rong, Z.; Wu, D.; Gan, J. Structure-guided protein engineering increases enzymatic activities of the SGNH family Esterases. Biotechnol. Biofuels 2020, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Baran, E.J. Cobalto: Un elemento crítico y estratégico. An. Acad. Nac. Cienc. Exactas Físicas Nat. 2018, 70, 77–106. [Google Scholar]
- Martins, L.S.; Lameira, J.; Kruger, H.G.; Alves, C.N.; Silva, J.R.A. Evaluating the performance of a non-bonded Cu2+ model including jahn—Teller effect into the binding of tyrosinase inhibitors. Int. J. Mol. Sci. 2020, 21, 4783. [Google Scholar] [CrossRef]
- Rettberg, L.A.; Wilcoxen, J.; Jasniewski, A.J.; Lee, C.C.; Tanifuji, K.; Hu, Y.; Britt, R.D.; Ribbe, M.W. Identity and function of an essential nitrogen ligand of the nitrogenase cofactor biosynthesis protein NifB. Nat. Commun. 2020, 11, 1757. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Moreira, D.; de Moraes, F.R.; Caruso, Í.P.; Chaim, O.M.; Senff-Ribeiro, A.; Ullah, A.; da Silva, L.S.; Chahine, J.; Arni, R.K.; Veiga, S.S. Potential implications for designing drugs against the brown spider venom phospholipase-d. J. Cell. Biochem. 2017, 118, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Tingting, Y.; Junmei, D.; Qingxia, Z.; Nanyu, H.; Jialin, Y.; Yunjuan, Y.; Junjun, L.; Yuelin, M.; Qian, W. Identification and characterization of a new alkaline sgnh hydrolase from a thermophilic Bacterium Bacillus sp K91. J. Microbiol. Biotechnol. 2016, 26, 730–738. [Google Scholar] [CrossRef]
- Garba, L.M.; Sallau, A.B.; Abubakar, M.; Hussain, A.M.; Ibrahim, Z.; Muhammad, Z. Biochemical and kinetic properties of crude phospholipase A2 from Naja nigricollis venom. Sci. World J. 2019, 14, 61–64. [Google Scholar]
- Dawson, R.; Irvine, R.; Hemington, N.; Hirasawa, K. The stimulation of the brain alkaline phospholipase A 1 attacking phosphatidylethanolamine by various salts and metal chelators. Neurochem. Res. 1982, 7, 1149–1161. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, C.; Ma, Q. Structural analysis of a Vibrio phospholipase reveals an unusual Ser-His-chloride catalytic triad. J. Biol. Chem. 2019, 294, 11391–11401. [Google Scholar] [CrossRef] [PubMed]
- Kraut, D.; Goff, H.; Pai, R.K.; Hosea, N.A.; Silman, I.; Sussman, J.L.; Taylor, P.; Voet, J.G. Inactivation studies of acetylcholinesterase with phenylmethylsulfonyl fluoride. Mol. Pharmacol. 2000, 57, 1243–1248. [Google Scholar] [PubMed]
- Chakraborti, S.; Michael, J.R. Involvement of a serine esterase in oxidant-mediated activation of phospholipase A2 in pulmonary endothelium. FEBS Lett. 1991, 281, 185–187. [Google Scholar] [CrossRef]
- Kwak, W.J.; Moon, T.C.; Lin, C.X.; Rhyn, H.G.; Jung, H.; Lee, E.; Kwon, D.Y.; Son, K.H.; Kim, H.P.; Kang, S.S. Papyriflavonol A from Broussonetia papyrifera inhibits the passive cutaneous anaphylaxis reaction and has a secretory phospholipase A2-inhibitory activity. Biol. Pharm. Bull. 2003, 26, 299–302. [Google Scholar] [CrossRef]
- Martinkova, M.; Igarashi, J.; Shimizu, T. Eukaryotic initiation factor 2α kinase is a nitric oxide-responsive mercury sensor enzyme: Potent inhibition of catalysis by the mercury cation and reversal by nitric oxide. FEBS Lett. 2007, 581, 4109–4114. [Google Scholar] [CrossRef][Green Version]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Tanaka, K.; Itazaki, H.; Yoshida, T. Cinatrins, a novel family of phospholipase A2 inhibitors ii. Biological activities. J. Antibiot. 1992, 45, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vazquez-Armenta, F.J.; Valdez-Olmos, U.F.; Arvizu-Flores, A.A.; Ayala-Zavala, J.F.; Ochoa-Leyva, A.; Lopez-Zavala, A.A. Metal Ions and Chemical Modification Reagents Inhibit the Enzymatic Activity of Lecithin-Dependent Hemolysin from Vibrio parahaemolyticus. Toxins 2022, 14, 609. https://doi.org/10.3390/toxins14090609
Vazquez-Armenta FJ, Valdez-Olmos UF, Arvizu-Flores AA, Ayala-Zavala JF, Ochoa-Leyva A, Lopez-Zavala AA. Metal Ions and Chemical Modification Reagents Inhibit the Enzymatic Activity of Lecithin-Dependent Hemolysin from Vibrio parahaemolyticus. Toxins. 2022; 14(9):609. https://doi.org/10.3390/toxins14090609
Chicago/Turabian StyleVazquez-Armenta, Francisco Javier, Uriel Felipe Valdez-Olmos, Aldo Alejandro Arvizu-Flores, Jesus Fernando Ayala-Zavala, Adrian Ochoa-Leyva, and Alonso Alexis Lopez-Zavala. 2022. "Metal Ions and Chemical Modification Reagents Inhibit the Enzymatic Activity of Lecithin-Dependent Hemolysin from Vibrio parahaemolyticus" Toxins 14, no. 9: 609. https://doi.org/10.3390/toxins14090609
APA StyleVazquez-Armenta, F. J., Valdez-Olmos, U. F., Arvizu-Flores, A. A., Ayala-Zavala, J. F., Ochoa-Leyva, A., & Lopez-Zavala, A. A. (2022). Metal Ions and Chemical Modification Reagents Inhibit the Enzymatic Activity of Lecithin-Dependent Hemolysin from Vibrio parahaemolyticus. Toxins, 14(9), 609. https://doi.org/10.3390/toxins14090609