Co-Occurrence of Hypoglycin A and Hypoglycin B in Sycamore and Box Elder Maple Proved by LC-MS/MS and LC-HR-MS


Abstract
:1. Introduction
2. Results and Discussion
2.1. Sample Selection
2.2. Extraction Optimization
2.3. Method Performance and Validation
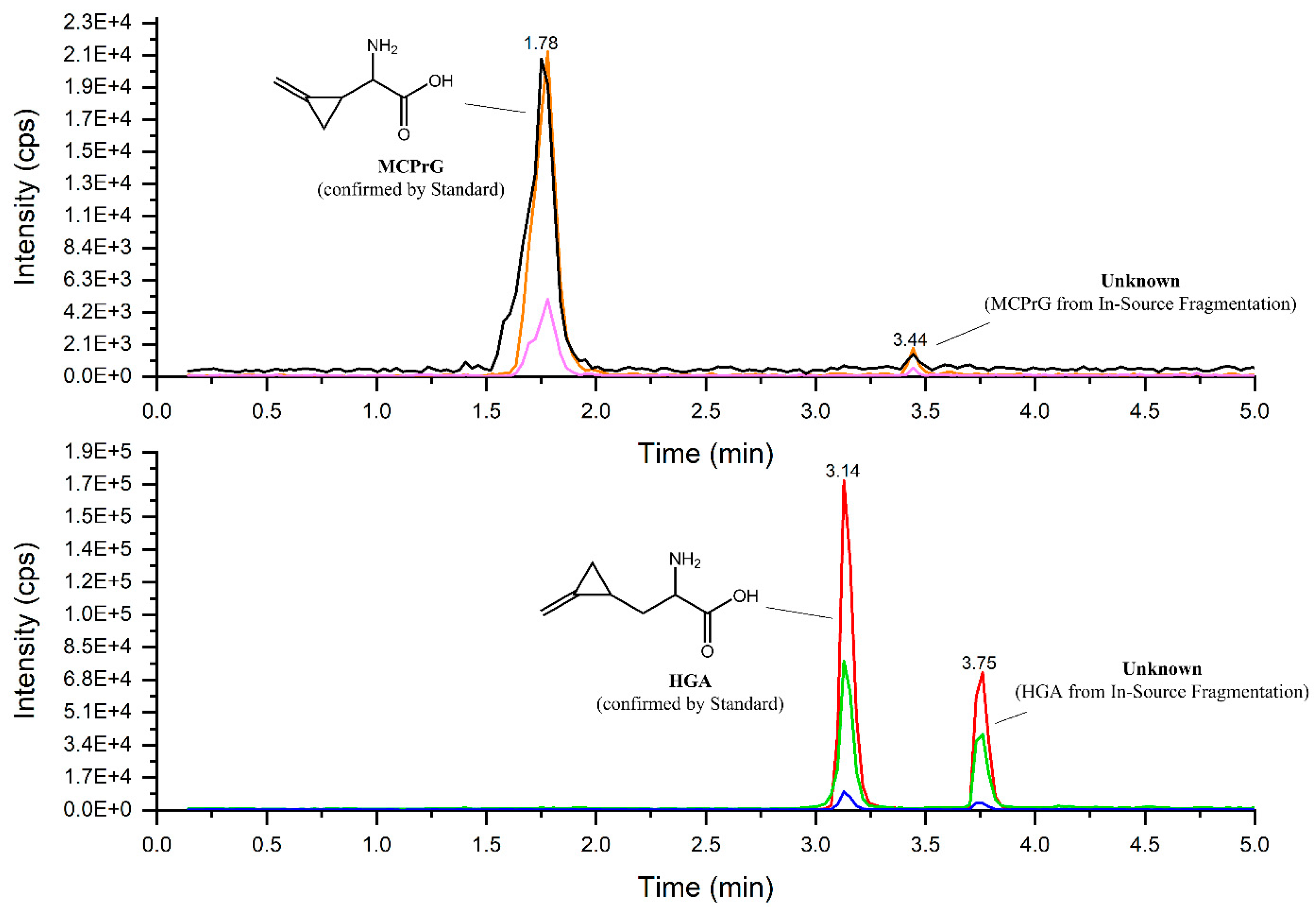
2.4. HGA and MCPrG in Field Samples
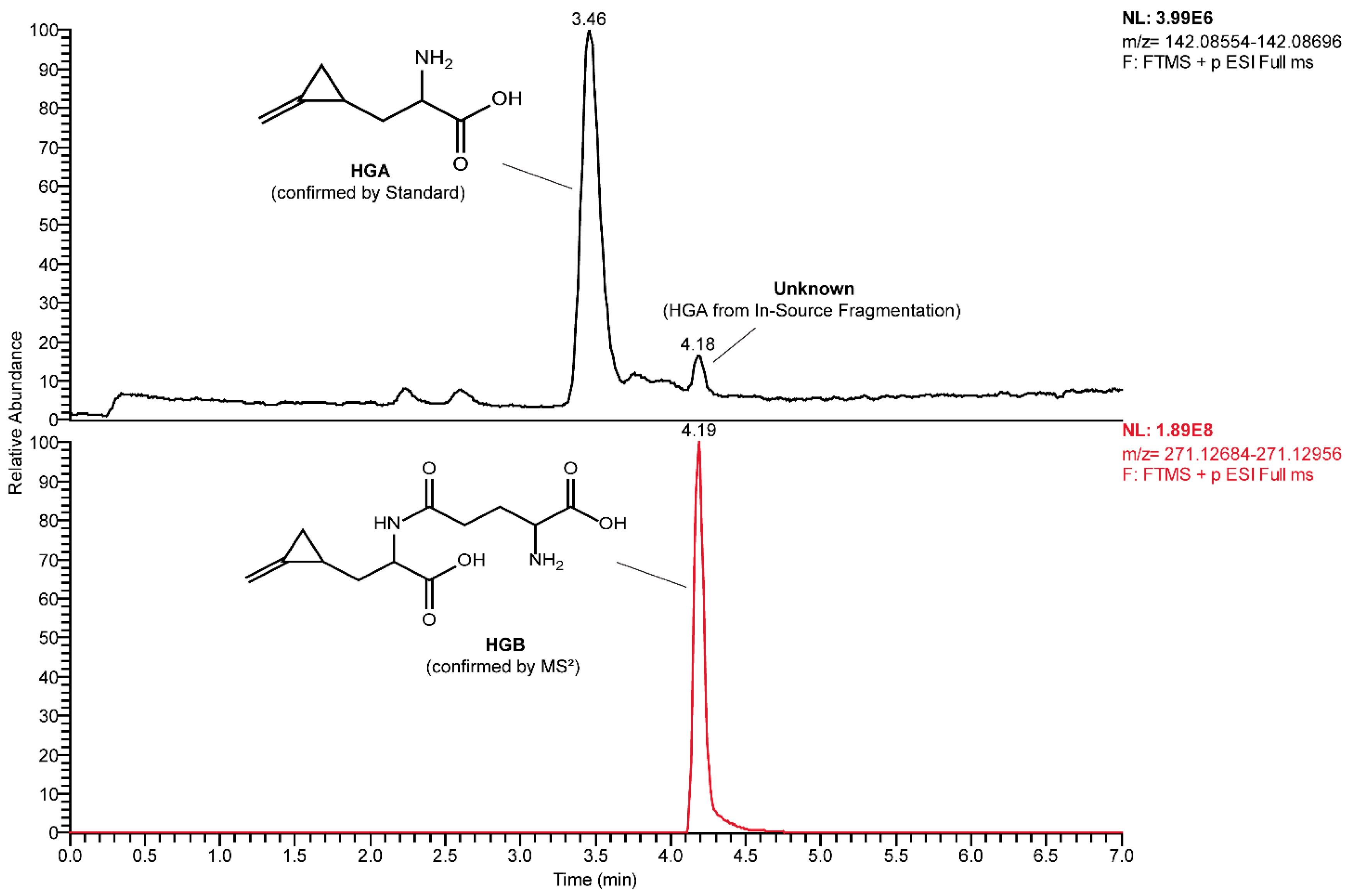
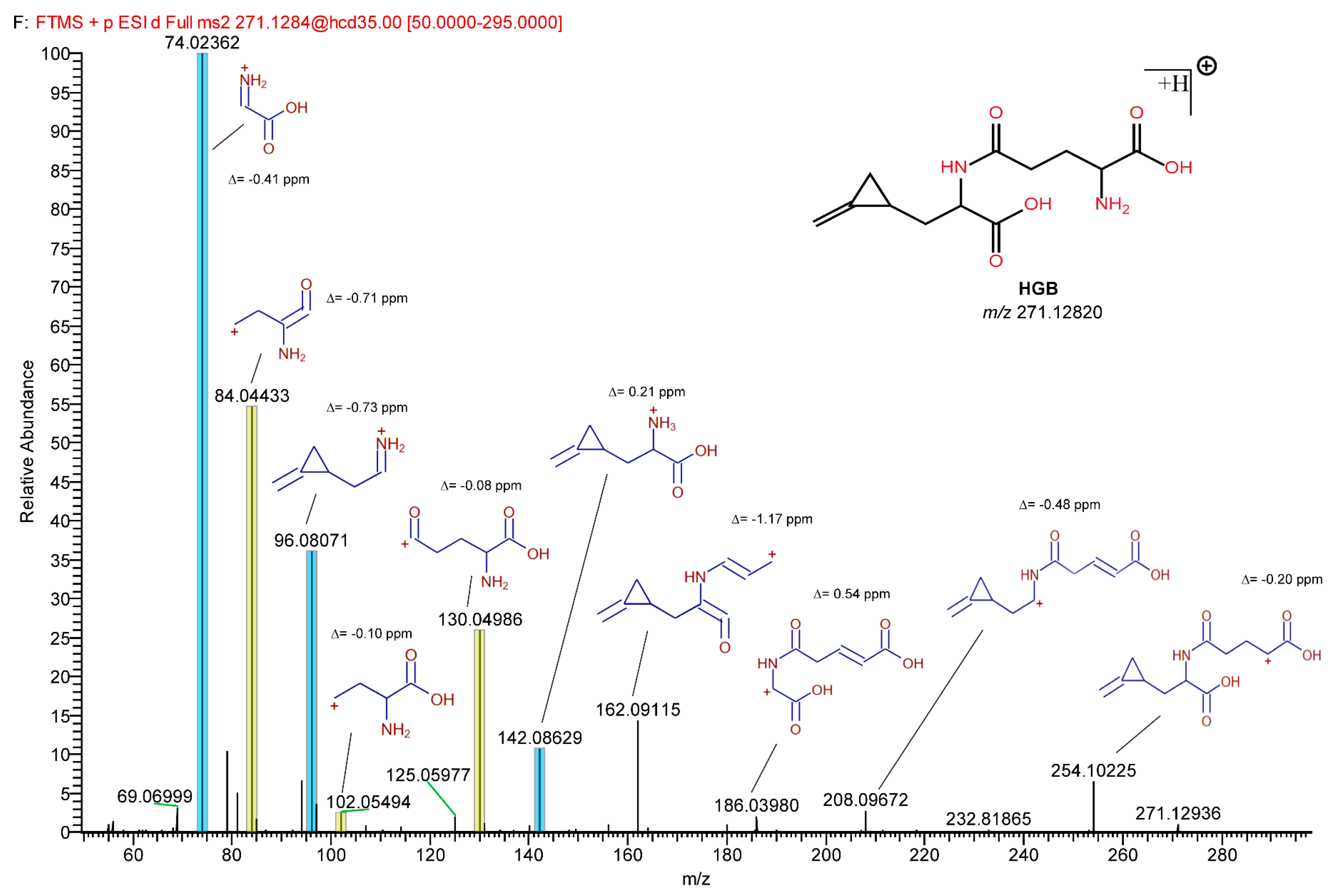
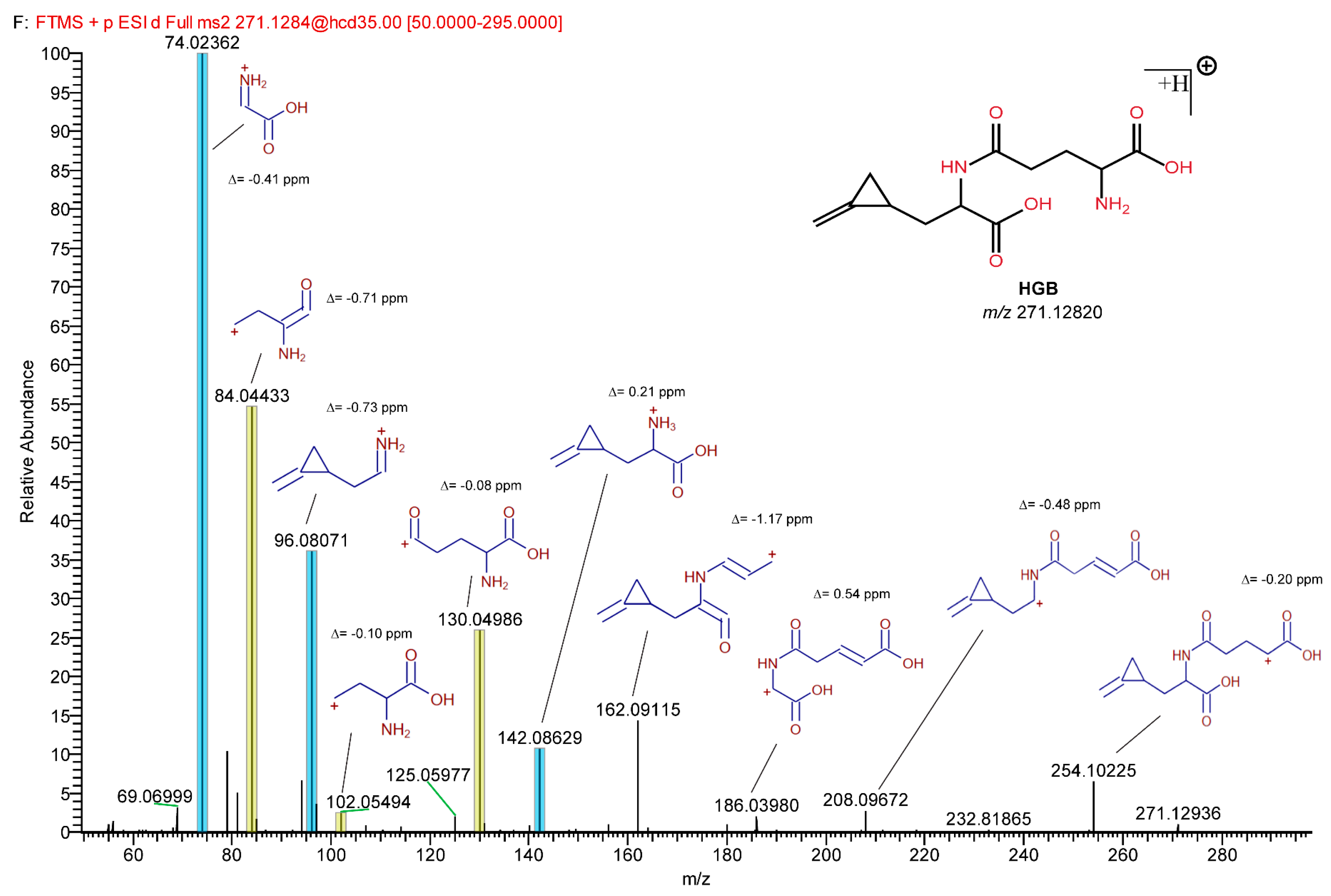
2.5. Detection of HGB and γ-Glutamyl-MCPrG in Sycamore and Box Elder Maple Samples
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Standards
4.2. Plant Materials
4.3. Sample Preparation
4.4. Stock and Working Standard Preparation
4.5. LC-MS/MS Instrumentation and Measurements
4.6. LC-HRMS/MS Instrumentation and Measurements
4.7. Method Validation
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manchester, K. Biochemistry of hypoglycin. FEBS Lett. 1974, 40, S128–S132. [Google Scholar] [CrossRef]
- Gaillard, Y.; Carlier, J.; Berscht, M.; Mazoyer, C.; Bevalot, F.; Guitton, J.; Fanton, L. Fatal intoxication due to ackee (Blighia sapida) in Suriname and French Guyana. GC–MS detection and quantification of hypoglycin-A. Forensic Sci. Int. 2011, 206, e103–e107. [Google Scholar] [PubMed]
- Bowen-Forbes, C.S.; Minott, D.A. Tracking hypoglycins A and B over different maturity stages: Implications for detoxification of ackee (Blighia sapida K.D. Koenig) fruits. J. Agric. Food Chem. 2011, 59, 3869–3875. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, S.L.; Carter, M.D.; Hayes, S.R.; Graham, L.A.; Johnson, D.; Mathews, T.P.; Harden, L.A.; Takeoka, G.R.; Thomas, J.D.; Pirkle, J.L.; et al. Quantification of toxins in soapberry (Sapindaceae) arils: Hypoglycin A and methylenecyclopropylglycine. J. Agric. Food Chem. 2016, 64, 5607–5613. [Google Scholar] [CrossRef]
- Sanford, A.; Isenberg, S.L.; Carter, M.D.; Mojica, M.A.; Mathews, T.P.; Harden, L.A.; Takeoka, G.R.; Thomas, J.D.; Pirkle, J.L.; Johnson, R.C. Quantitative HPLC–MS/MS analysis of toxins in soapberry seeds: Methylenecyclopropylglycine and hypoglycin A. Food Chem. 2018, 264, 449–454. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, X.; Zhang, P.; Wang, Y.; Xiao, Y.; Yang, B.; Qu, H.; Jiang, Y. Detection of toxic methylenecyclopropylglycine and hypoglycin A in litchi aril of three Chinese cultivars. Food Chem. 2020, 327, 127013. [Google Scholar] [CrossRef]
- Unger, L.; Nicholson, A.; Jewitt, E.; Gerber, V.; Hegeman, A.; Sweetman, L.; Valberg, S. Hypoglycin A Concentrations in Seeds of Acer pseudoplatanus Trees Growing on Atypical Myopathy-Affected and Control Pastures. J. Vet. Intern. Med. 2014, 28, 1289–1293. [Google Scholar] [CrossRef]
- Baise, E.; Habyarimana, J.A.; Amory, H.; Boemer, F.; Douny, C.; Gustin, P.; Marcillaud-Pitel, C.; Patarin, F.; Weber, M.; Votion, D.M. Samaras and seedlings of Acer pseudoplatanus are potential sources of hypoglycin A intoxication in atypical myopathy without necessarily inducing clinical signs. Equine Vet. J. 2016, 48, 414–417. [Google Scholar] [CrossRef]
- Westermann, C.; Van Leeuwen, R.; van Raamsdonk, L.; Mol, H. Hypoglycin A Concentrations in Maple Tree Species in the Netherlands and the Occurrence of Atypical Myopathy in Horses. J. Vet. Intern. Med. 2016, 30, 880–884. [Google Scholar] [CrossRef]
- Medina, S.G.; Hyde, C.; Lovera, I.; Piercy, R.J. Detection of equine atypical myopathy-associated hypoglycin a in plant material: Optimisation and validation of a novel LC-MS based method without derivatisation. PLoS ONE 2018, 13, e0199521. [Google Scholar]
- Valberg, S.J.; Sponseller, B.T.; Hegeman, A.D.; Earing, J.; Bender, J.B.; Martinson, K.L.; Patterson, S.E.; Sweetman, L. Seasonal pasture myopathy/atypical myopathy in North America associated with ingestion of hypoglycin A within seeds of the box elder tree. Equine Vet. J. 2012, 45, 419–426. [Google Scholar] [CrossRef]
- Vashishtha, V.M. Outbreaks of hypoglycemic encephalopathy in Muzzaffarpur, India: Are these caused by toxins in litchi fruit? Indian Pediatr. 2016, 53, 399–402. [Google Scholar]
- Shrivastava, A.; Kumar, A.; Thomas, J.D.; Laserson, K.F.; Bhushan, G.; Carter, M.D.; Chhabra, M.; Mittal, V.; Khare, S.; Sejvar, J.J.; et al. Association of acute toxic encephalopathy with litchi consumption in an outbreak in Muzaffarpur, India, 2014: A case-control study. Lancet Glob. Health 2017, 5, e458–e466. [Google Scholar] [CrossRef]
- Scott, H.H. On the ‘Vomiting Sickness’ of Jamaica. Ann. Trop. Med. Parasitol. 1916, 10, 1–78. [Google Scholar] [CrossRef]
- Trauner, D.A.; Nyhan, W.L.; Sweetman, L.; Tanaka, K.A.Y. Jamaican Vomiting Sickness and Reye’s Syndrome. N. Engl. J. Med. 1976, 295, 1481–1482. [Google Scholar] [PubMed]
- Golden, K.D.; Kean, E.A.; Terry, S.I. Jamaican vomiting sickness: A study of two adult cases. Clin. Chim. Acta 1984, 142, 293–298. [Google Scholar] [CrossRef]
- Joskow, R.; Belson, M.; Vesper, H.; Backer, L.; Rubin, C. Ackee fruit poisoning: An outbreak investigation in Haiti 2000–2001, and review of the literature. Clin. Toxicol. 2006, 44, 267–273. [Google Scholar] [CrossRef]
- Gordon, A. Biochemistry of Hypoglycin and Toxic Hypoglycemic Syndrome. In Food Safety and Quality Systems in Developing Countries: Volume One: Export Challenges and Implementation Strategies; Elsevier: Amsterdam, The Netherlands, 2015; pp. 47–61. [Google Scholar]
- Hörügel, U.; Simon, H.; Dressel, A.; Birke, H.; Matz, K.; Klunker, M. Atypical myopathy in horses: Review and investigation results from Saxony and Thuringia. Prakt. Tierarzt 2014, 95, 444–457. [Google Scholar]
- Aboling, S.; Schliephake, A.; Cavalleri, J.-M.V.; Kamphues, J. Proof of sycamore maple in the content of the digestive tract in a horse on suspect of atypical myopathy. Pferdeheilkunde Equine Med. 2015, 31, 135–139. [Google Scholar] [CrossRef]
- Bochnia, M.; Ziegler, J.; Sander, J.; Uhlig, A.; Schaefer, S.; Vollstedt, S.; Glatter, M.; Abel, S.; Recknagel, S.; Schusser, G.F.; et al. Hypoglycin A Content in Blood and Urine Discriminates Horses with Atypical Myopathy from Clinically Normal Horses Grazing on the Same Pasture. PLoS ONE 2015, 10, e0136785. [Google Scholar] [CrossRef] [PubMed]
- González-Medina, S.; Ireland, J.; Piercy, R.J.; Newton, J.R.; Votion, D.M. Equine atypical myopathy in the UK: Epidemiological characteristics of cases reported from 2011 to 2015 and factors associated with survival. Equine Vet. J. 2017, 49, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Bochnia, M.; Sander, J.; Ziegler, J.; Terhardt, M.; Sander, S.; Janzen, N.; Cavalleri, J.; Zuraw, A.; Wensch-Dorendorf, M.; Zeyner, A. Detection of MCPG metabolites in horses with atypical myopathy. PLoS ONE 2019, 14, e0211698. [Google Scholar] [CrossRef] [PubMed]
- Bunert, C.; Langer, S.; Votion, D.M.; Boemer, F.; Müller, A.; Ternes, K.; Liesegang, A. Atypical myopathy in Père David’s deer (Elaphurus davidianus) associated with ingestion of hypoglycin A. J. Anim. Sci. 2018, 96, 3537–3547. [Google Scholar] [CrossRef] [PubMed]
- Bochnia, M.; Ziemssen, E.; Sander, J.; Stief, B.; Zeyner, A. Methylenecyclopropylglycine and hypoglycin A intoxication in three Pére David’s Deers (Elaphurus davidianus) with atypical myopathy. Vet. Med. Sci. 2021, 7, 998–1005. [Google Scholar] [CrossRef]
- Hirz, M.; Gregersen, H.A.; Sander, J.; Votion, D.M.; Schänzer, A.; Köhler, K.; Herden, C. Atypical myopathy in 2 Bactrian camels. J. Veter. Diagn. Investig. 2021, 33, 961–965. [Google Scholar] [CrossRef]
- Renaud, B.; François, A.-C.; Boemer, F.; Kruse, C.; Stern, D.; Piot, A.; Petitjean, T.; Gustin, P.; Votion, D.-M. Grazing Mares on Pasture with Sycamore Maples: A Potential Threat to Suckling Foals and Food Safety through Milk Contamination. Animals 2021, 11, 87. [Google Scholar] [CrossRef]
- Sander, J.; Terhardt, M.; Janzen, N. Detection of maple toxins in mare’s milk. J. Vet. Intern. Med. 2021, 35, 606–609. [Google Scholar] [CrossRef]
- Bochnia, M.; Ziegler, J.; Glatter, M.; Zeyner, A. Hypoglycin A in Cow’s Milk—A Pilot Study. Toxins 2021, 13, 381. [Google Scholar] [CrossRef]
- Westermann, C.M.; Dorland, L.; Votion, D.M.; De Sain-van der Velden, M.G.M.; Wijnberg, I.D.; Wanders, R.J.A.; Spliet, W.G.M.; Testerink, N.; Berger, R.; Ruiter, J.P.N.; et al. Acquired multiple Acyl-CoA dehydrogenase deficiency in 10 horses with atypical myopathy. Neuromuscul. Disord. 2008, 18, 355–364. [Google Scholar] [CrossRef]
- Sander, J.; Terhardt, M.; Sander, S.; Janzen, N. Quantification of Methylenecyclopropyl Compounds and Acyl Conjugates by UPLC-MS/MS in the Study of the Biochemical Effects of the Ingestion of Canned Ackee (Blighia sapida) and Lychee (Litchi chinensis). J. Agric. Food Chem. 2017, 65, 2603–2608. [Google Scholar] [CrossRef]
- Barceloux, D.G. Akee Fruit and Jamaican Vomiting Sickness (Blighia sapida Köenig). Dis. Mon. 2009, 55, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Seeff, L.; Stickel, F.; Navarro, V.J. Chapter 35—Hepatotoxicity of Herbals and Dietary Supplements. In Drug-Induced Liver Disease, 3rd ed.; Kaplowitz, N., DeLeve, L.D., Eds.; Academic Press: Boston, MA, USA, 2013; pp. 631–657. [Google Scholar]
- Hassall, C.H.; Reyle, K.; Feng, P. Hypoglycin A, B: Biologically active polypeptides from Blighia sapida. Nature 1954, 173, 356–357. [Google Scholar] [CrossRef] [PubMed]
- Golden, K.D.; Williams, O.J.; Bailey-Shaw, Y. High-Performance Liquid Chromatographic Analysis of Amino Acids in Ackee Fruit with Emphasis on the Toxic Amino Acid Hypoglycin A. J. Chromatogr. Sci. 2002, 40, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowden, L.; Pratt, H.M. Cyclopropylamino acids of the genus Acer: Distribution and biosynthesis. Phytochemistry 1973, 12, 1677–1681. [Google Scholar] [CrossRef]
- Kean, E.A. Hypoglycin. In Toxicants of Plant Origin: Proteins and Amino Acids; Cheeke, P.R., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1989; Volume III, pp. 229–262. [Google Scholar]
- Brown, M.; Bates, R.; McGowan, C.; Cornell, J. Influence of Fruit Maturity on the Hypoglycin a Level in Ackee (Blighia sapida). J. Food Saf. 1991, 12, 167–177. [Google Scholar] [CrossRef]
- McGowan, C.; Wiley, V.A.; Bates, R.P. Application of methodology for RP-HPLC amino acid analysis to the measurement of hypoglycin A. BioChromatography 1989, 4, 161–164. [Google Scholar]
- Whitaker, T.B.; Saltsman, J.J.; Ware, G.M.; Slate, A.B. Evaluating the Performance of Sampling Plans to Detect Hypoglycin A in Ackee Fruit Shipments Imported into the United States. J. AOAC Int. 2007, 90, 1060–1072. [Google Scholar] [CrossRef]
- Sander, J.; Terhardt, M.; Sander, S.; Janzen, N. Quantification of hypoglycin A as butyl ester. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1029–1030, 169–173. [Google Scholar] [CrossRef]
- Żuraw, A.; Dietert, K.; Kühnel, S.; Sander, J.; Klopfleisch, R. Equine atypical myopathy caused by hypoglycin A intoxication associated with ingestion of sycamore maple tree seeds. Equine Vet. J. 2016, 48, 418–421. [Google Scholar] [CrossRef]
- González-Medina, S.; Bevin, W.; Alzola-Domingo, R.; Chang, Y.; Piercy, R.J. Hypoglycin A absorption in sheep without concurrent clinical or biochemical evidence of disease. J. Vet. Intern. Med. 2021, 35, 1170–1176. [Google Scholar] [CrossRef]
- Kean, E.A.; Hare, E.R. Gamma-Glutamyl-Transferase Transpeptidase of the Ackee Plant. Phytochemistry 1980, 19, 199–203. [Google Scholar] [CrossRef]
- Young, M. Studies on the Growth in Culture of Plant Cells: XVI. Nitrogen Assimilation during Nitrogen-Limited Growth of Acer pseudoplatanus L. Cells in Chemostat Culture. J. Exp. Bot. 1973, 24, 1172–1185. [Google Scholar] [CrossRef]
- Chen, K.K.; Anderson, R.C.; McCowen, M.C.; Harris, P.N. Pharmacologic action of hypoglycin A and B. J. Pharmacol. Exp. Ther. 1957, 121, 272–285. [Google Scholar] [PubMed]
- Persaud, T.V. Effect of intra-amniotic administration of hypoglycin B on foetal development in the rat. Exp. Pathol. 1972, 6, 55–58. [Google Scholar]
- SANTE/2019/12682; Guidance Document on Analytical Quality Control and Method Validation Procedures for Pesticide Residues Analysis in Food and Feed. European-Commission, Directorate-General for Health and Food Safety SANTE: Brussels, Belgium, 2019.
- European-Commission; Wenzl, T.; Haedrich, J.; Schaechtele, A.; Robouch, P.; Stroka, J. Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food; Joint Research Centre (JRC): Geel, Belgium, 2016. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | HGA | MCPrG | |
|---|---|---|---|
| Calibration range (ng/mL) | 0.5–100 | 0.5–100 | |
| (10–2000 µg/kg) | (10–2000 µg/kg) | ||
| Correlation coefficient (r) | 0.9999 | 0.9999 | |
| LOD (µg/kg) | 5.0 | 6.4 | |
| LOQ (µg/kg) | 16.4 | 21.2 | |
| Recovery (%) | 20 µg/kg | 95.5 | 105.1 |
| 50 µg/kg | 104.7 | 99.6 | |
| 500 µg/kg | 92.1 | 98.1 | |
| 1500 µg/kg | 89.7 | 93.5 | |
| Repeatability (RSDr) (%) | 20 µg/kg | 6.5 | 7.8 |
| 50 µg/kg | 12.9 | 6.0 | |
| 500 µg/kg | 2.6 | 4.3 | |
| 1500 µg/kg | 13.2 | 1.0 | |
| Within-laboratory reproducibility (RSDwR) (%) | 20 µg/kg | 10.8 | 10.7 |
| 50 µg/kg | 16.0 | 7.3 | |
| 500 µg/kg | 9.4 | 9.0 | |
| 1500 µg/kg | 9.4 | 8.1 | |
| Matrix effect (%) | 48 | 53 | |
| Species | Sample Type | Concentration (mg/kg) | Peak Area | Peak Area Ratio | |||
|---|---|---|---|---|---|---|---|
| HGA | MCPrG | HGB | γ-glutamyl-MCPrG | HGB/HGA | γ-glutamyl-MCPrG/MCPrG | ||
| A. pseudoplatanus (sycamore maple) | Leaves | 512 | 11 | 1.3E+07 | 2.1E+07 | 0.31 | 9.9 |
| (n = 7) | (120–3202) | (6.3–158) | (1.0E+07–2.2E+08) | (3.4E+06–1.9E+08) | (0.08–1.1) | (0.21–22) | |
| Seeds | 1333 | 146 | 1.2E+08 | 1.4E+08 | 0.90 | 5.4 | |
| (n = 7) | (266–2962) | (35–267) | (8.6E+06–1.3E+09) | (1.1E+06–4.1E+08) | (0.03–29) | (0.08–46) | |
| Seedlings | 3865 | 395 | 6.7E+08 | 3.0E+08 | 1.2 | 7.9 | |
| (n = 2) | (3223–4508) | (290–500) | (5.8E+08–7.6E+08) | (2.4E+08–3.6E+08) | (1.2–1.3) | (7.4–8.5) | |
| A. negundo (box elder maple) | Leaves | 535 | 3.4 | 1.7E+07 | 9.3E+06 | 0.26 | 26 |
| (n = 2) | (24–1047) | (2.7–4.1) | (1.0E+06–3.3E+07) | (3.4E+06–1.5E+07) | (0.24–0.29) | (22–29) | |
| Seeds | 410 | 39 | 1.9E+08 | 1.7E+08 | 3.6 | 36 | |
| (n = 2) | (236–584) | (22–56) | (1.6E+08–2.3E+08) | (7.6E+07–2.6E+08) | (2.9–4.3) | (30–42) | |
| A. platanoides (Norway maple) | Leaves and seeds | N.D. | N.D. | N.D. | N.D. | N.A. | N.A. |
| (n = 3) | |||||||
| A. campestre (field maple) | Leaves and seeds | N.D. | N.D. | N.D. | N.D. | N.A. | N.A. |
| A. tataricum (Tatar maple) | Leaves and seeds | N.D. | N.D. | N.D. | N.D. | N.A. | N.A. |
| A. cappadocicum (Cappadocian maple) | Leaves and seeds | N.D. | N.D. | N.D. | N.D. | N.A. | N.A. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Khatib, A.H.; Engel, A.M.; Weigel, S. Co-Occurrence of Hypoglycin A and Hypoglycin B in Sycamore and Box Elder Maple Proved by LC-MS/MS and LC-HR-MS. Toxins 2022, 14, 608. https://doi.org/10.3390/toxins14090608
El-Khatib AH, Engel AM, Weigel S. Co-Occurrence of Hypoglycin A and Hypoglycin B in Sycamore and Box Elder Maple Proved by LC-MS/MS and LC-HR-MS. Toxins. 2022; 14(9):608. https://doi.org/10.3390/toxins14090608
Chicago/Turabian StyleEl-Khatib, Ahmed H., Anna Maria Engel, and Stefan Weigel. 2022. "Co-Occurrence of Hypoglycin A and Hypoglycin B in Sycamore and Box Elder Maple Proved by LC-MS/MS and LC-HR-MS" Toxins 14, no. 9: 608. https://doi.org/10.3390/toxins14090608
APA StyleEl-Khatib, A. H., Engel, A. M., & Weigel, S. (2022). Co-Occurrence of Hypoglycin A and Hypoglycin B in Sycamore and Box Elder Maple Proved by LC-MS/MS and LC-HR-MS. Toxins, 14(9), 608. https://doi.org/10.3390/toxins14090608