Spaceflight Changes the Production and Bioactivity of Secondary Metabolites in Beauveria bassiana


Abstract
:1. Introduction
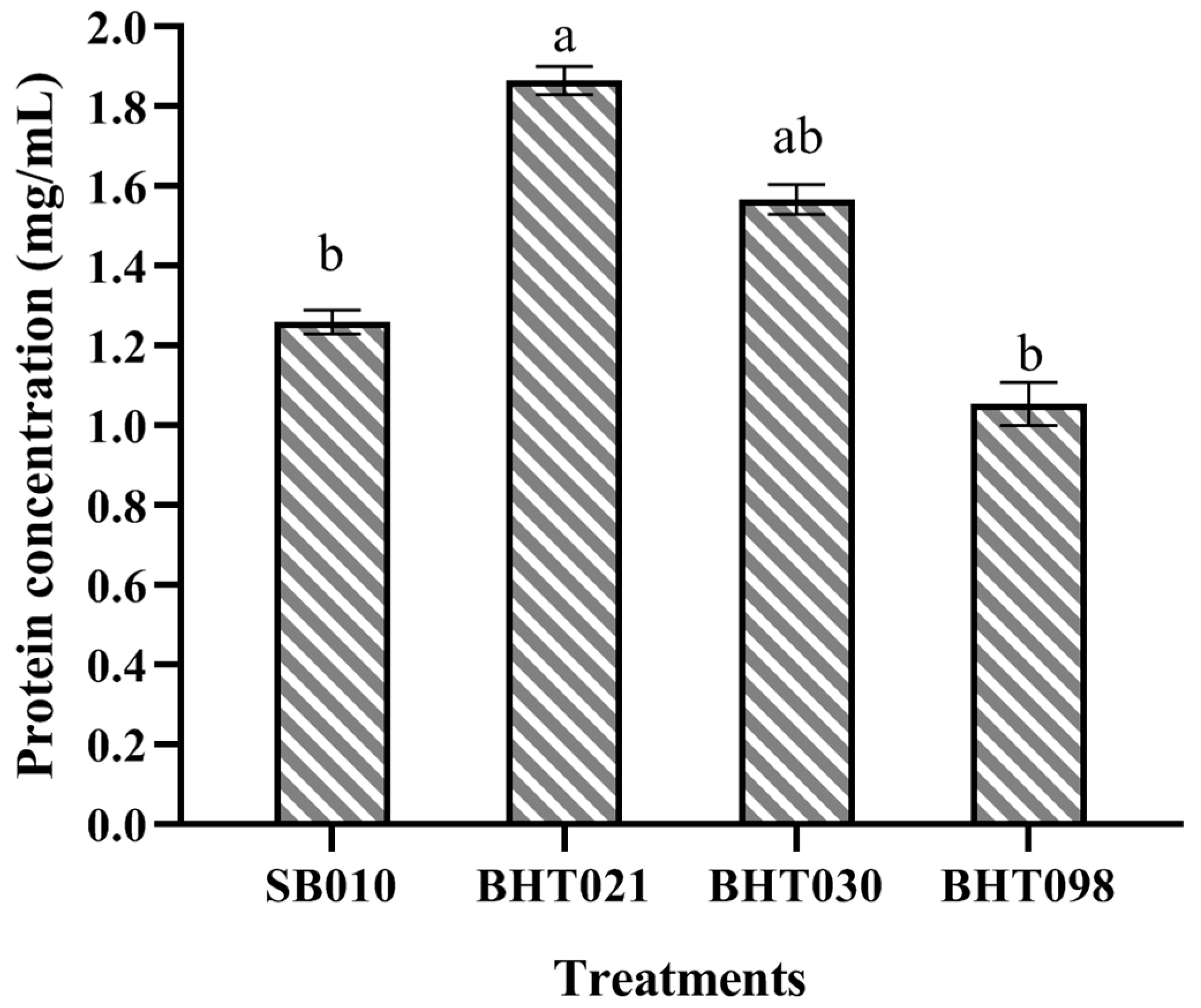
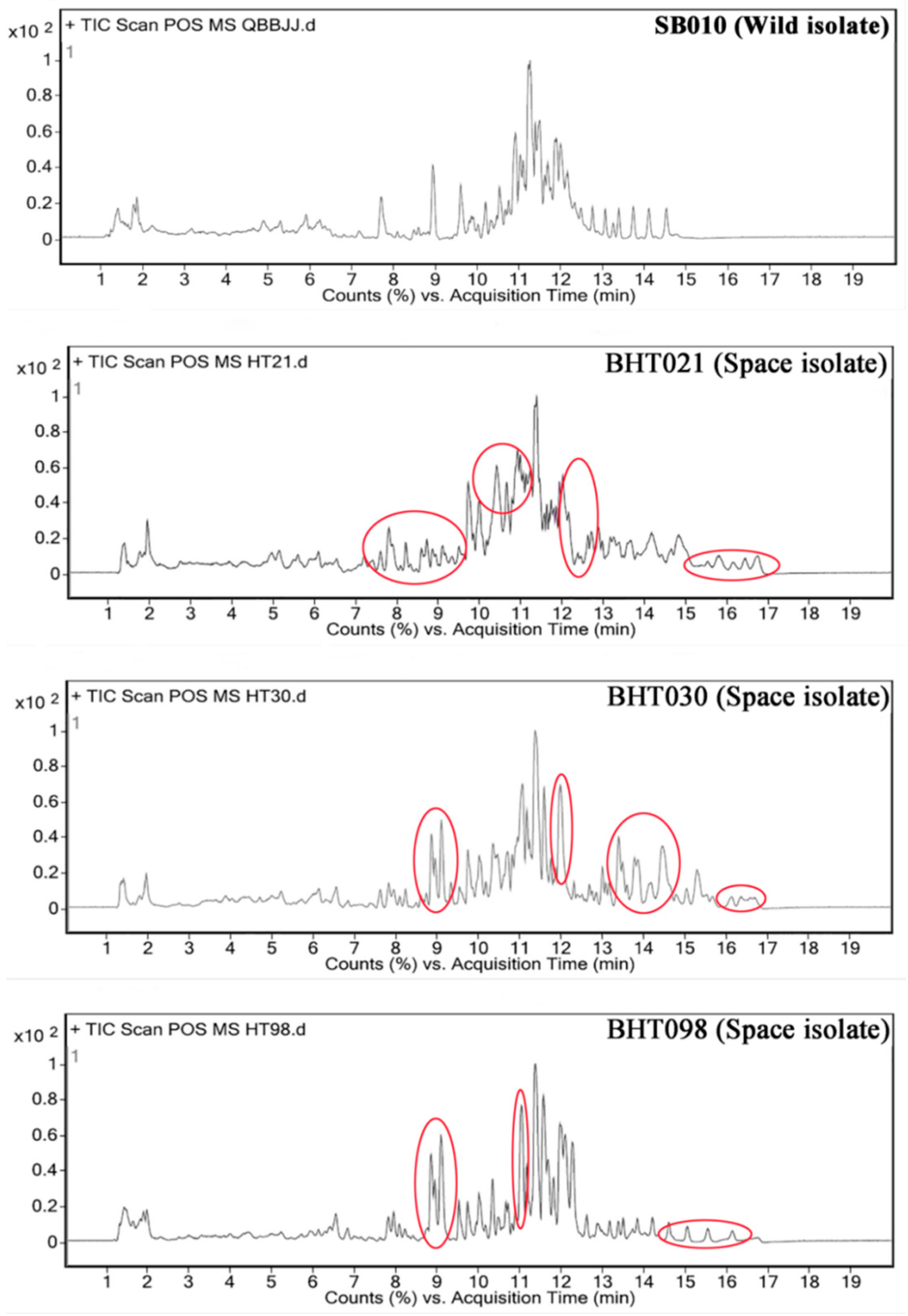
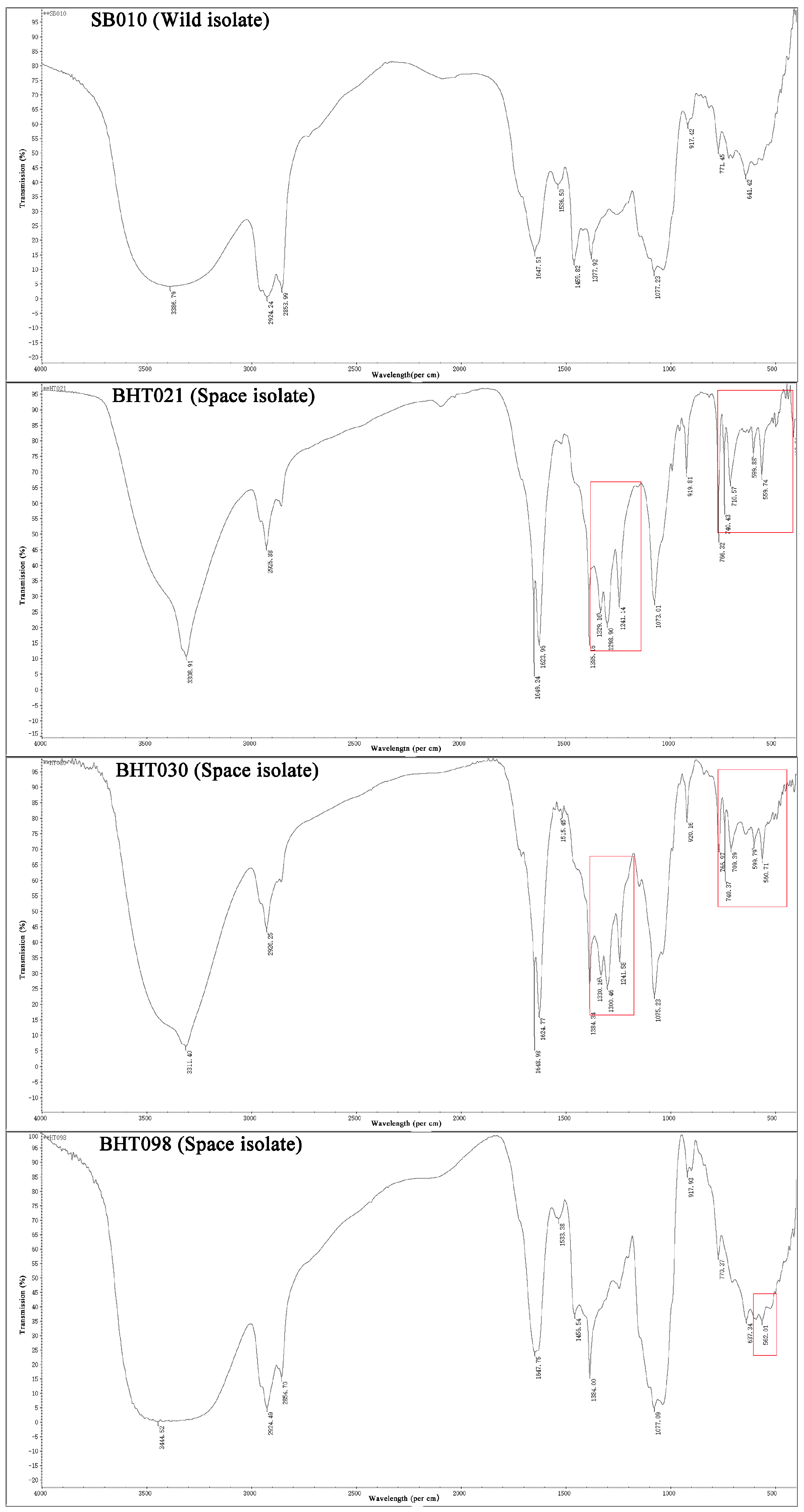
2. Results
2.1. Nuclear Magnetic Resonance (NMR)
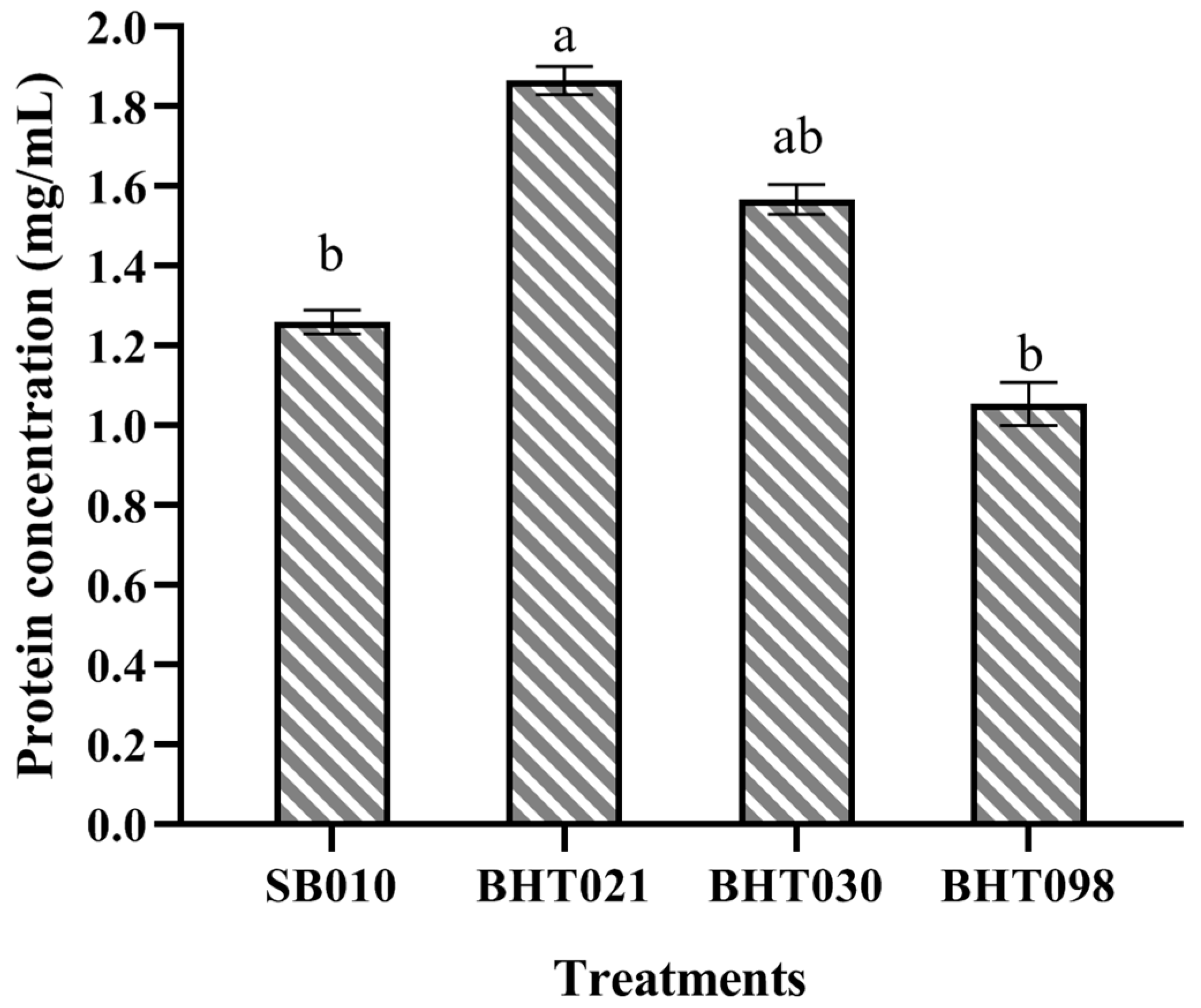
2.2. Toxicity of Metabolites against M. usitatus
2.3. Transmission Electron Microscopic Examination of M. usitatus Midgut following Treatment with Mycelial Extracts from B. bassina Wild Isolate and Space Mutants
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Cultures
5.2. Fungal Preparations
5.3. Exposure of Beauveria bassiana to Spaceflight Conditions
5.4. Production and Characterization of Crude Protein Extracts of Wild Isolate (SB010) and Spaceflight Mutants (BHT021, BHT030, and BHT098) of Beauveria bassiana
5.5. Toxicity of Crude Protein Extracts of Wild Isolate (SB010, and Spaceflight Mutants (BHT021, BHT030, and BHT098) of Beauveria bassiana against Megalurothrips usitatus
5.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, R.; Torres, M.; Clemens, R.; Hateley, S.; Hosmani, R.; Wade, W.; Bhattacharaya, S. Spaceflight and simulated microgravity conditions increase virulence of Serratia marcescens in the Drosophila melanogaster infection model. Npj Microgravity 2020, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Li, D.G.; Huang, Y.; Liu, C.T. Effects of spaceflight and simulated microgravity on microbial growth and secondary metabolism. Milit. Med. Res. 2018, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Barrila, J.; Radtke, L.A.; Crabbe, A.; Saker, S.F.; Herbst-Kralovetz, M.M.; Ott, C.M.; Nickerson, C.A. Organotypic 3D cell culture models: Using the rotating wall vessel to study host–pathogen interactions. Nat. Rev. Microbiol. 2010, 8, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.W. Impact of space flight on bacterial virulence and antibiotic susceptibility. Infect. Drug Resist. 2015, 8, 249–262. [Google Scholar] [CrossRef]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Effect of microgravity and space radiation on microbes. Future Micobiol. 2018, 13, 831–847. [Google Scholar] [CrossRef]
- Horneck, G.; Kluas, D.M.; Mancinelli, R. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef]
- Benoit, M.R.; Li, W.; Stodieck, L.S.; Lam, K.S.; Winther, C.L.; Roane, T.M.; Klaus, D.M. Microbial antibiotic production aboard the international Space Station. Appl. Microbiol. Biotechnol. 2006, 70, 403–411. [Google Scholar] [CrossRef]
- Rosenzweig, J.A.; Abogunde, O.; Thomas, K.; Lawal, A.; Nguyen, Y.U.; Sodipe, A.; Jejelowo, A. Spaceflight and modelled microgravity effects on microbial growth and virulence. Appl. Microbiol. Biotechnol. 2010, 85, 885–891. [Google Scholar] [CrossRef]
- Abshire, C.F.; Prasai, K.; Soto, I.; Shi, R.; Concha, M.; Baddoo, M.; Flemington, E.K.; Ennis, D.G.; Scott, R.S.; Harrison, L. Exposure of Mycobacterium marinum to low-shear modelled microgravity: Effect on growth, the transcriptome and survival under stress. Npj Microgravity 2016, 2, 16038. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R. Toward a global picture of bacterial secondary metabolism. J. Ind. Microbiol. Biotechnol. 2019, 46, 301–311. [Google Scholar] [CrossRef]
- Horak, I.; Engelbrecht, G.; van Rensburg, P.J.J.; Claassens, S. Microbial metabolomics: Essential definitions and the importance of cultivation conditions for utilizing Bacillus Species as bionematicides. J. Appl. Microbiol. 2019, 127, 326–343. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Curtis, P.D. The impacts of microgravity on bacterial metabolism. Life 2022, 12, 774. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Demain, A.L. Metabolic regulation and overproduction of primary metabolites. Microb. Biotechnol. 2008, 1, 283–319. [Google Scholar] [CrossRef] [PubMed]
- Milojevic, T.; Weckwerth, W. Molecular mechanisms of microbial survivability in outer space: A systems biology approach. Front. Microbiol. 2020, 11, 923. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Kumar, P. Microbial metabolites in nutrition, healthcare and agriculture. 3 Biotech. 2017, 7, 15. [Google Scholar] [CrossRef]
- Craney, A.; Ozimok, C.; Pimentel-Elardo, S.M.; Capretta, A.; Nodwell, J.R. Chemical perturbation of secondary metabolism demonstrates important links to primary metabolism. Chem. Biol. 2012, 19, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chang, D.; Xu, H.; Chen, J.; Su, L.; Guo, Y.; Chen, Z.; Wang, Y.; Wang, L.; Wang, J. Impact of a short-term exposure to spaceflight on the phenotype, genome, transcriptome and proteome of Escherichia coli. Int. J. Astrobiol. 2015, 14, 435–444. [Google Scholar] [CrossRef]
- Komatsu, M.; Uchiyama, T.; Omura, S.; Cane, D.E.; Ikeda, H. Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 2646–2651. [Google Scholar] [CrossRef]
- Su, X.Y.; Schmitz, G.; Zhang, M.L.; Mackie, R.I.; Cann, I.K.O. Heterologous gene expression in filamentous fungi. Adv. Appl. Microbiol. 2012, 81, 1–61. [Google Scholar]
- Wang, Q.; Zhong, C.; Xiao, H. Genetic engineering of filamentous fungi for efficient protein expression and secretion. Front. Bioeng. Biotechnol. 2020, 8, 293. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C.; Zied, D.C. Filamentous fungi in biological control: Current status and future perspectives. Chil. J. Agri. Res. 2019, 79, 307–315. [Google Scholar] [CrossRef]
- Wu, J.H.; Yu, X.; Wang, X.S.; Tang, L.D.; Ali, S. Matrine enhances the pathogenicity of Beauveria brongniartii against Spodoptera litura (Lepidoptera: Noctuidae). Front. Microbiol. 2019, 10, 1812. [Google Scholar] [CrossRef] [PubMed]
- Leys, N.M.E.J.; Hendrickx, L.; De Boever, P.; Baatout, S.; Mergeay, M. Space flight effects on bacterial physiology. J. Biol. Regul. Homeost. Agents 2004, 18, 193–199. [Google Scholar] [PubMed]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial responses to microgravity and other low-shear environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. Secondary metabolism in simulated microgravity. Chem. Rec. 2001, 1, 333–346. [Google Scholar] [CrossRef]
- Benoit, M.R.; Klaus, D.M. Microgravity, bacteria, and the influence of motility. Adv. Space Res. 2007, 39, 1225–1232. [Google Scholar] [CrossRef]
- Bibb, M.J. Regulation of secondary metabolism in Streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef]
- Viollier, P.H.; Kelemen, G.H.; Dale, G.E.; Nguyen, K.T.; Buttner, M.J.; Thompson, C.J. Specialized osmotic stress response systems involve multiple SigB-like sigma factors in Streptomyces coelicolor. Mol. Microbiol. 2003, 47, 699–714. [Google Scholar] [CrossRef]
- Lam, K.; Gustavson, D.R.; Pirnik, D.L.; Pack, E.; Bulanhagui, C.; Mamber, S.W.; Forenza, S.; Stodieck, L.S.; Klaus, D.M. The effect of space flight on the production of actinomycin D by Streptomyces plicatus. J. Ind. Microbiol. Biotechnol. 2002, 29, 299–302. [Google Scholar] [CrossRef]
- Luo, A.; Gao, C.; Song, Y.; Tan, H.; Liu, Z. Biological responses of a Streptomyces strain producing- nikkomycin to space flight. Space Med. Med. Eng. 1998, 11, 411–414. [Google Scholar]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.F.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, P.J.; Contreras, J.; Quinto, V.; Grávalos, C.; Fernández, E.; Bielza, P. Metabolic mechanisms of insecticide resistance in the western flower thrips, Frankliniella occidentalis (Pergande). Pest. Manag. Sci. 2005, 61, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Yang, B.; Wu, J.H.; Ali, S. Identification and virulence characterization of two Akanthomyces attenuatus isolates against Megalurothrips usitatus (Thysanopera: Thripidea). Insects 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Huang, Z.; Ren, S.X. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J. Pest. Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Zhao, J.; Yao, Y.N.; Wei, Y.; Huang, S.; Keyhani, N.O.; Huang, Z. Screening of Metarhizium anisopliae UV-induced mutants for faster growth yields a hyper-virulent isolate with greater UV and thermal tolerances. Appl. Microbiol. Biotechnol. 2016, 100, 9217–9228. [Google Scholar] [CrossRef]
- Wu, J.; Yang, B.; Xu, J.; Cuthbertson, A.G.S.; Ali, S. Characterization and toxicity of crude toxins produced by cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch). Toxins 2021, 13, 220. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- SAS Institute. SAS User’s Guide; Statistics SAS Institute: Cary, NC, USA, 2000. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Concentration (μg/mL) | Mortality (%) |
|---|---|---|
| SB010 | ddH2O | 5.00 j |
| 100 | 88.33 ab | |
| 75 | 80.00 c | |
| 50 | 73.33 d | |
| 25 | 65.00 ef | |
| 12.5 | 50.00 h | |
| BHT021 | ddH2O | 5.00 j |
| 100 | 100.00 a | |
| 75 | 91.67 ab | |
| 50 | 83.33 bc | |
| 25 | 70.00 e | |
| 12.5 | 63.33 f | |
| BHT030 | ddH2O | 5.00 |
| 100 | 95.00 a | |
| 75 | 83.33 | |
| 50 | 75.00 d | |
| 25 | 63.33 f | |
| 12.5 | 56.67 g | |
| BHT098 | ddH2O | 5.00 j |
| 100 | 81.67 c | |
| 75 | 73.33 d | |
| 50 | 61.67 f | |
| 25 | 55.00 g | |
| 12.5 | 46.67 i |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, X.; Zhang, J.; Ali, S.; Wu, J. Spaceflight Changes the Production and Bioactivity of Secondary Metabolites in Beauveria bassiana. Toxins 2022, 14, 555. https://doi.org/10.3390/toxins14080555
Zhang Y, Zhang X, Zhang J, Ali S, Wu J. Spaceflight Changes the Production and Bioactivity of Secondary Metabolites in Beauveria bassiana. Toxins. 2022; 14(8):555. https://doi.org/10.3390/toxins14080555
Chicago/Turabian StyleZhang, Youdan, Xiaochen Zhang, Jieming Zhang, Shaukat Ali, and Jianhui Wu. 2022. "Spaceflight Changes the Production and Bioactivity of Secondary Metabolites in Beauveria bassiana" Toxins 14, no. 8: 555. https://doi.org/10.3390/toxins14080555
APA StyleZhang, Y., Zhang, X., Zhang, J., Ali, S., & Wu, J. (2022). Spaceflight Changes the Production and Bioactivity of Secondary Metabolites in Beauveria bassiana. Toxins, 14(8), 555. https://doi.org/10.3390/toxins14080555