Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance



Abstract
1. Introduction
2. BMAA Producers and Pathways of Biosynthesis
2.1. History of BMAA Discovery
2.2. BMAA Producers
2.2.1. Cyanobacteria
2.2.2. Diatoms
2.2.3. Dinoflagellates
2.2.4. Bacteria
2.2.5. Plants
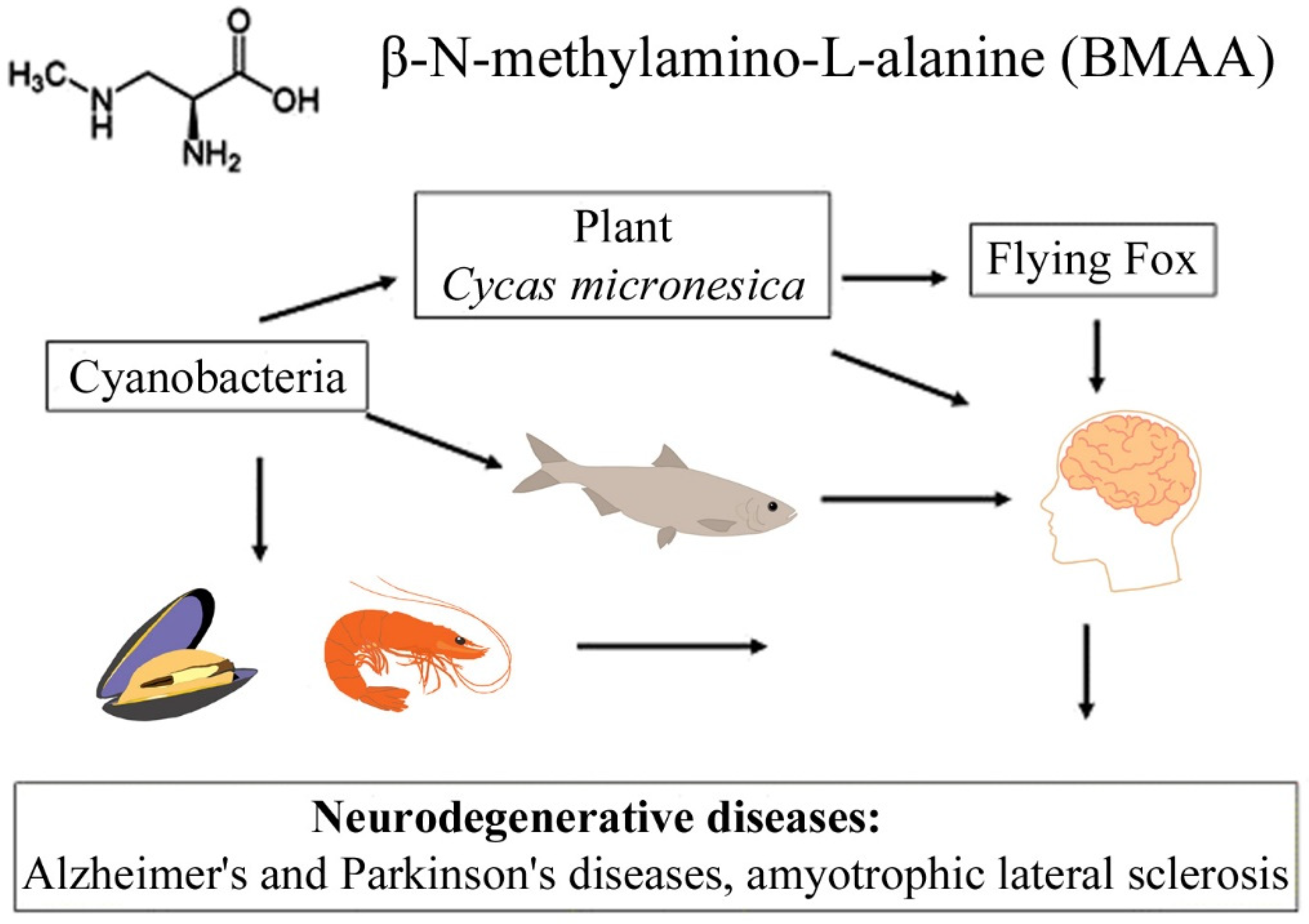
2.3. Bioaccumulation and Biomagnification
2.4. Biosynthesis Pathways of BMAA: Different Hypothesis
3. BMAA Identification in Cells and Natural Environment Samples
3.1. Experimental Approaches and Methods Used for BMAA Identification
3.2. Methodological Difficulties in BMAA Identification
4. Molecular Mechanisms Underlying BMAA Toxicity
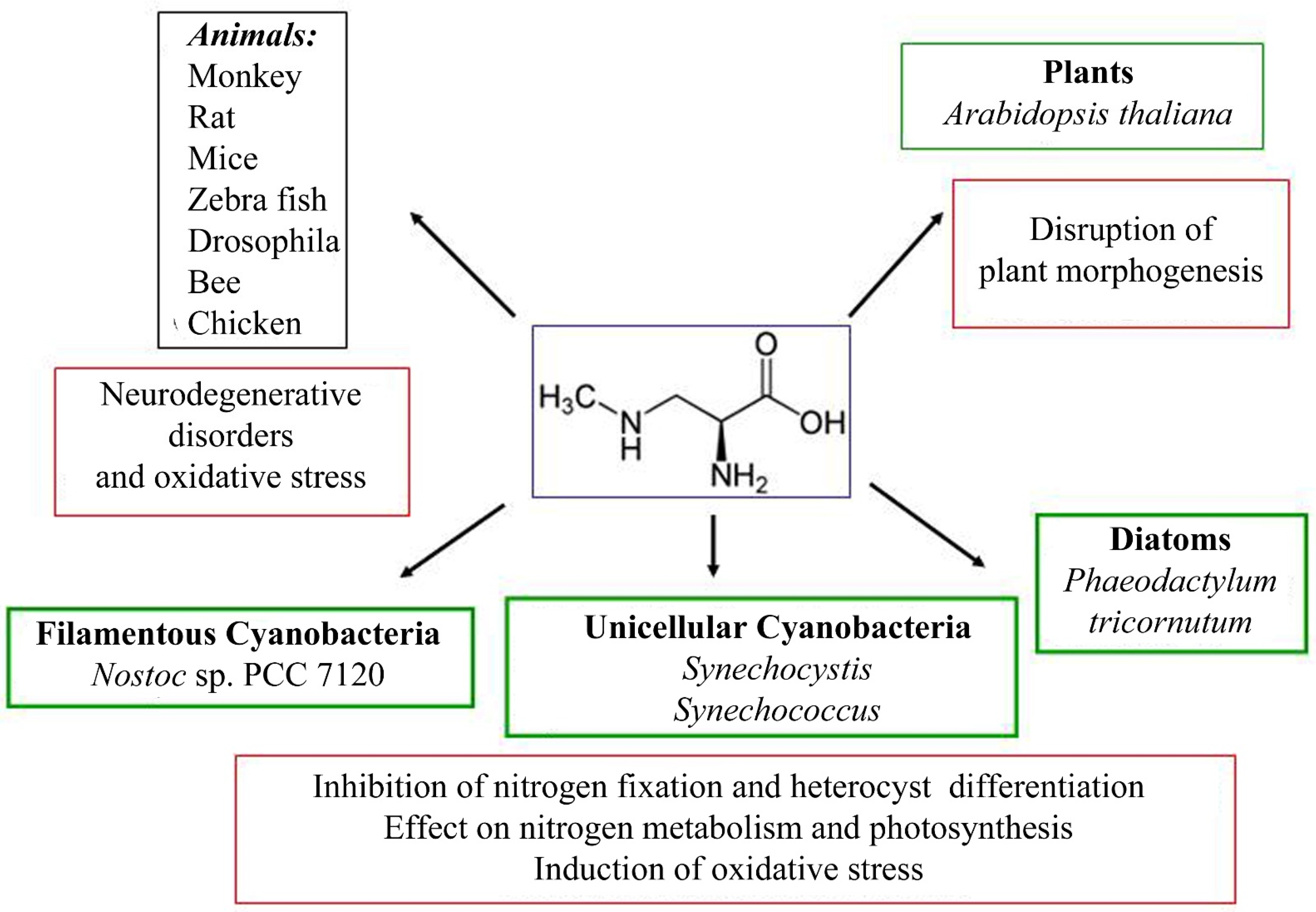
4.1. The Effect of BMAA on Various Organisms: Pleiotropic Toxic Effect
4.2. BMAA Neurotoxicity Hypothesis: BMAA as a Glutamate Receptors Agonist
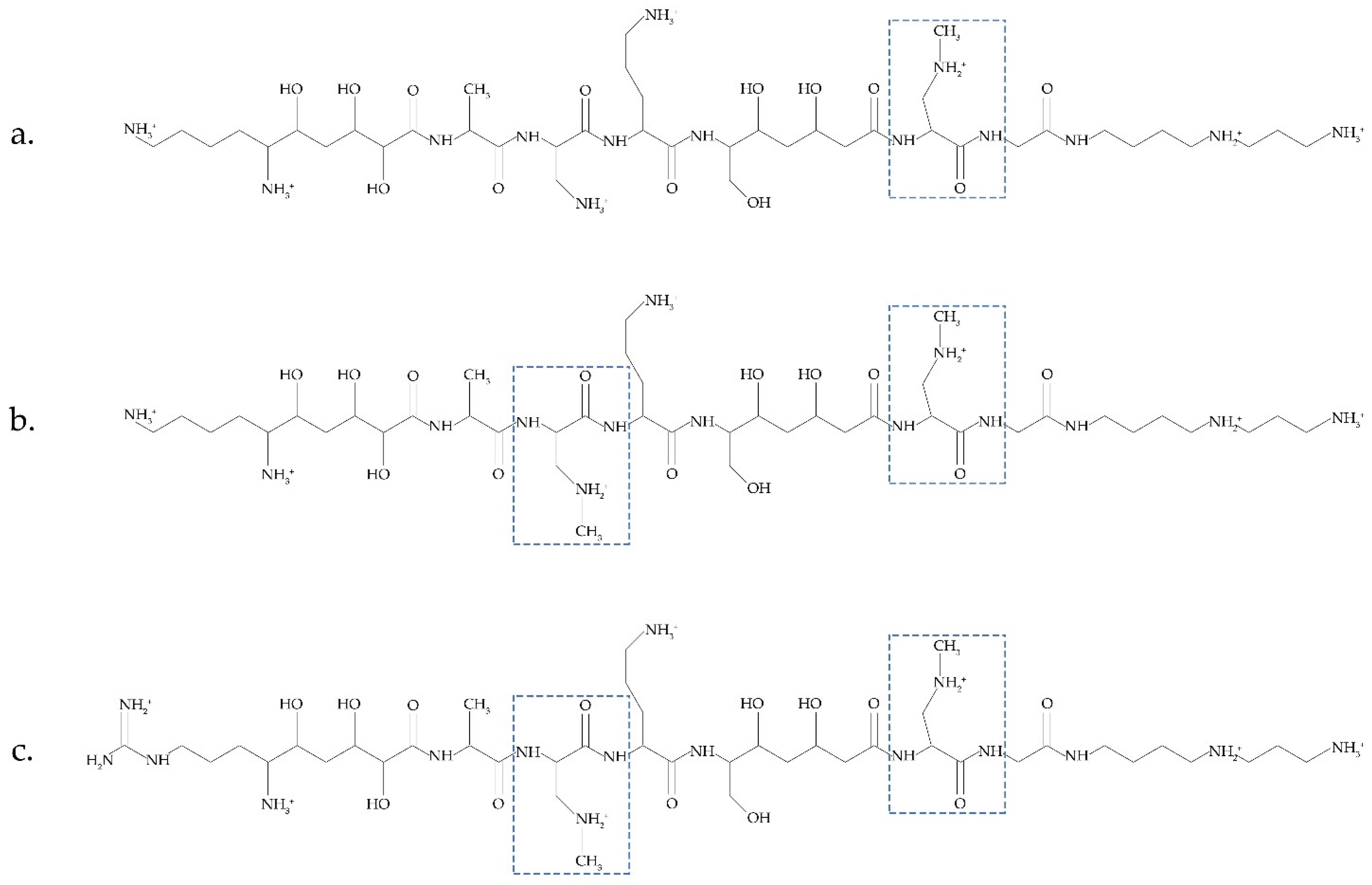
4.3. BMAA and Proteins: Misincorporation and Electrostatic Binding
4.4. BMAA as a Metal-Binding Amino Acid
4.5. The Effect of BMAA on Nitrogen Metabolism in Eukaryotic and Prokaryotic Cells
4.6. Can BMAA Induce Oxidative Stress and Apoptosis?
5. The Biological Significance of BMAA and Further Research Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vega, A.; Bell, E.A. α-Amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Nunn, P.B. 50 years of research on α-amino-β-methylaminopropionic acid (β-methylaminoalanine). Phytochemistry 2017, 144, 271–281. [Google Scholar] [CrossRef]
- Vranova, V.; Rejsek, K.; Skene, K.R.; Formanek, P. Non-protein amino acids: Plant, soil and ecosystem interactions. Plant Soil 2011, 342, 31–48. [Google Scholar] [CrossRef]
- Shenker, M.; Fan, T.W.M.; Crowley, D.E. Phytosiderophores influence on cadmium mobilization and uptake by wheat and barley plants. J. Environ. Qual. 2001, 30, 2091–2098. [Google Scholar] [CrossRef]
- Casagrande, D.J.; Given, P.H. Geochemistry of amino acids in some Florida peat accumulation-II. Amino acid distributions. Geochim. Cosmochim. Acta 1980, 44, 1493–1507. [Google Scholar] [CrossRef]
- Walsh, C.T.; O’Brien, R.V.; Khosla, C. Nonproteinogenic Amino Acid Building Blocks for Nonribosomal Peptide and Hybrid Polyketide Scaffolds. Angew. Chem. Int. Ed. 2013, 52, 7098–7124. [Google Scholar] [CrossRef]
- Hirano, A.; Malamud, N.; Elizan, T.S.; Kurland, L.T. Amyotrophic lateral sclerosis and Parkinsonism-dementia complex on Guam. Further pathologic studies. Arch. Neurol. 1966, 15, 35–51. [Google Scholar] [CrossRef]
- Spencer, P.S.; Nunn, P.B.; Hugon, J.; Ludolph, A.C.; Ross, S.M.; Roy, D.N.; Robertson, R.C. Guam amyotrophic lateral sclerosis-parkinsonism-dementia linked to a plant excitant neurotoxin. Science 1987, 237, 517–522. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of beta-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Actaneurologica Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef]
- Bradley, W.G.; Mash, D.C. Beyond Guam: The cyanobacteria/BMAA hypothesis of the cause of ALS and other neurodegenerative diseases. Amyotroph. Lateral Scler. 2009, 10, 7–20. [Google Scholar] [CrossRef]
- Popova, A.A.; Koksharova, O.A. Neurotoxic non-proteinogenic amino acid β-N-methylamino-L-alanine and its role in biological systems. Biochemistry 2016, 81, 794–805. [Google Scholar] [CrossRef]
- Nunn, P.B.; Codd, G.A. Environmental distribution of the neurotoxin l-BMAA in Paenibacillus species. Toxicol. Res. 2019, 8, 781–783. [Google Scholar] [CrossRef]
- Brody, J.A.; Stanhope, J.M.; Kurland, L.T. Patterns of amyotrophic lateral sclerosis and parkinsonism-dementia on Guam. Contemp Neurol Ser. 1975, 12, 45–70. [Google Scholar]
- Pablo, J.; Banack, S.A.; Cox, P.A.; Johnson, T.E.; Papapetropoulos, S.; Bradley, W.G.; Buck, A.; Mash, D.C. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s disease. Acta Neurol Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce beta-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Downing, T.G. β-N-methylamino-L-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Banack, S.A.; Johnson, H.E.; Cheng, R.; Cox, P.A. Production of the neurotoxin BMAA by a marine cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of beta-N-methylamino-L-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin beta-N-methylamino-L-alanine (BMAA) in South Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spacil, Z.; Ilag, L.L.; Ronnevi, L.O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Cervantes Cianca, R.C.; Baptista, M.S.; Lopes, V.R.; Vasconcelos, V.M. The non-protein amino acid β-N-methylamino-L-alanine in Portuguese cyanobacterial isolates. Amino Acids 2012, 42, 2473–2479. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Sebastian Funke, M.; Peuthert, A.; Pflugmacher, S. β-N-Methylamino-L-alanine exposure alters defense against oxidative stress in aquatic plants Lomariopsis lineata, Fontinalis antipyretica, Riccia fluitans and Taxiphyllum barbieri. Ecotoxicol. Environ. Saf. 2013, 88, 72–78. [Google Scholar] [CrossRef]
- Spacil, Z.; Eriksson, J.; Jonasson, S.; Rasmussen, U.; Ilag, L.L.; Bergman, B. Analytical protocol for identification of BMAA and DAB in biological samples. Analyst 2009, 135, 127–132. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-L-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Violi, J.P.; Mitrovic, S.M.; Colville, A.; Main, B.J.; Rodgers, K.J. Prevalence of β-methylamino-L-alanine (BMAA) and its isomers in freshwater cyanobacteria isolated from eastern Australia. Ecotoxicol Environ Saf. 2019, 172, 72–81. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Wilbraham, J.; Banack, S.A.; Metcalf, J.S.; Codd, G.A. Microcystins, BMAA and BMAA isomers in 100-year-old Antarctic cyanobacterial mats collected during Captain R.F. Scott’s Discovery Expedition. Eur. J. Phycol. 2018, 53, 115–121. [Google Scholar] [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and bacteria isolated from a diatom culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-L-alanine (BMAA) and Its Isomers by Freshwater Diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.S.; Lundholm, N.; Hubbard, K.A.; Montresor, M.; Leaw, C.P. Toxic and harmful marine diatoms. In Diatoms: Fundamentals and Applications; Seckbach, J., Gordon, R., Eds.; Scrivener Publishing LLC: Salem, MA, USA, 2019; pp. 389–434. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef]
- Marler, T.E.; Snyder, L.R.; Shaw, C.A. Cycas micronesica (Cycadales) plants devoid of endophytic cyanobacteria increase in β-methylamino-L-alanine. Toxicon 2010, 56, 563–568. [Google Scholar] [CrossRef]
- Downing, T.G.; Phelan, R.R.; Downing, S. A potential physiological role for cyanotoxins in cyanobacteria of arid environments. J. Arid Environ. 2015, 112, 147–151. [Google Scholar] [CrossRef]
- Downing, S.; Downing, T.G. The metabolism of the non proteinogenic amino acid β-N-methylamino-L-alanine (BMAA) in the cyanobacterium Synechocystis PCC 6803. Toxicon 2016, 115, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; Codd, G.A. Metabolic solutions to the biosynthesis of some diaminomonocarboxylic acids in nature: Formation in cyanobacteria of the neurotoxins 3-N-methyl-2,3-diaminopropanoic acid (BMAA) and 2,4-diaminobutanoic acid (2,4-DAB). Phytochemistry 2017, 144, 253–270. [Google Scholar] [CrossRef]
- Jiang, L.; Kiselova, N.; Rosén, J.; Ilag, L.L. Quantification of neurotoxin BMAA (β-N-methylamino-L-alanine) in seafood from Swedish markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef]
- Salomonsson, M.L.; Fredriksson, E.; Alfjorden, A.; Hedeland, M.; Bondesson, U. Seafood sold in Sweden contains BMAA: A study of free and total concentrations with UHPLC-MS/MS and dansyl chloride derivatization. Toxicol. Rep. 2015, 2, 1473–1481. [Google Scholar] [CrossRef]
- Hammerschlag, N.; Davis, D.A.; Mondo, K.; Seely, M.S.; Murch, S.J.; Glover, W.B.; Divoll, T.; Evers, D.C.; Mash, D.C. Cyanobacterial Neurotoxin BMAA and Mercury in Sharks. Toxins 2016, 8, 238. [Google Scholar] [CrossRef]
- Regueiro, J.; Negreira, N.; Carreira-Casais, A.; Pérez-Lamela, C.; Simal-Gándara, J. Dietary exposure and neurotoxicity of the environmental free and bound toxin β-N-methylamino-l-alanine. Food Res. Int. 2017, 100, 1–13. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Schwanemann, T.; Esterhuizen-Londt, M.; Pflugmacher, S. Protein association of β-N-methylamino-L-alanine in Triticum aestivum via irrigation. Food Addit. Contam. Part A 2018, 35, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen-Londt, M.; Pflugmacher, S. Vegetables cultivated with exposure to pure and naturally occurring β-N-methylamino-L-alanine (BMAA) via irrigation. Environ. Res. 2019, 169, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Rydberg, S. Transfer of the Neurotoxin β-N-methylamino-l-alanine (BMAA) in the Agro–Aqua Cycle. Mar. Drugs 2020, 18, 244. [Google Scholar] [CrossRef] [PubMed]
- Brenner, E.D.; Stevenson, D.W.; McCombie, R.W.; Katari, M.; A Rudd, S.; Mayer, K.F.X.; Palenchar, P.M.; Runko, S.J.; Twigg, R.W.; Dai, G.; et al. Expressed sequence tag analysis in Cycas, the most primitive living seed plant. Genome Biol. 2003, 4, R78. [Google Scholar] [CrossRef]
- Mantas, M.J.Q.; Nunn, P.B.; Codd, G.A.; Barker, D. Genomic insights into the biosynthesis and physiology of the cyanobacterial neurotoxin 3-N-methyl-2,3-diaminopropanoic acid (BMAA). Phytochemistry 2022, 200, 113198. [Google Scholar] [CrossRef]
- Müller, S.; Garcia-Gonzalez, E.; Mainz, A.; Hertlein, G.; Heid, N.C.; Mösker, E.; van den Elst, H.V.; Overkleeft, H.S.; Genersch, E.; Süssmuth, R.D.; et al. Paenilamicin: Structure and biosynthesis of a hybrid nonribosomal peptide/polyketide antibiotic from the bee pathogen Paenibacillus larvae. Angew. Chem. Int. Ed. 2014, 53, 10821–10825. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.; Guo, H.; Zhang, H. The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef]
- Berstad, K.; Berstad, J.E.R. Parkinson’s disease; the hibernating spore hypothesis. Med. Hypotheses 2017, 104, 48–53. [Google Scholar] [CrossRef]
- Brenner, S. Blue-green algae or cyanobacteria in the intestinal micro-flora may produce neurotoxins such as β-N-Methylamino-L-Alanine (BMAA) which may be related to development of amyotrophic lateral sclerosis, Alzheimer’s disease and Parkinson-Dementia-Complex in humans and equine motor neuron disease in horses. Med. Hypotheses 2013, 80, 103. [Google Scholar] [CrossRef]
- Nunes-Costa, D.; Magalhães, J.D.; G-Fernandes, M.; Cardoso, S.M.; Empadinhas, N. Microbial BMAA and the Pathway for Parkinson’s Disease Neurodegeneration. Front. Aging Neurosci. 2020, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzi, S.C.; Sharon, I.; Wrighton, K.C.; Koren, O.; Hug, L.A.; Thomas, B.C.; Goodrich, J.K.; Bell, J.; Spector, T.D.; Banfield, J.T.; et al. The human gut and groundwater harbor non-photosynthetic bacteria belonging to a new candidate phylum sibling to Cyanobacteria. Elife 2013, 2, e01102. [Google Scholar] [CrossRef] [PubMed]
- Soo, R.M.; Skennerton, C.T.; Sekiguchi, Y.; Imelfort, M.; Paech, S.J.; Dennis, P.G.; Steen, J.A.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. An expanded genomic representation of the phylum cyanobacteria. Genome. Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.; Hellenas, K.E. Determination of the neurotoxin BMAA (beta-N-methylamino-L-alanine) in cycad seed and cyanobacteria by LC-MS/MS (liquid chromatography tandem mass spectrometry). Analyst 2008, 133, 1785–1789. [Google Scholar] [CrossRef]
- Bishop, S.L.; Murch, S.J. A systematic review of analytical methods for the detection and quantification of β-N-methylamino-l-alanine (BMAA). Analyst 2020, 145, 13–28. [Google Scholar] [CrossRef]
- Faassen, E.J. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef]
- Faassen, E.; Antoniou, M.; Beekman-Lukassen, W.; Bláhová, L.; Chernova, E.; Christophoridis, C.; Combes, A.; Edwards, C.; Fastner, J.; Harmsen, J.; et al. A Collaborative Evaluation of LC-MS/MS Based Methods for BMAA Analysis: Soluble Bound BMAA Found to Be an Important Fraction. Mar. Drugs 2016, 14, 45. [Google Scholar] [CrossRef]
- Porojan, C.; Mitrovic, S.M.; Yeo, D.C.J.; Furey, A. Overview of the potent cyanobacterial neurotoxin β-methylamino-L-alanine (BMAA) and its analytical determination. Food Addit. Contam. Part A 2016, 33, 1570–1586. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Banack, S.A.; Bishop, S.L.; Metcalf, J.S.; Murch, S.J.; Davis, D.A.; Stommel, E.W.; Karlsson, O.; Brittebo, E.B.; Chatziefthimiou, A.D.; et al. Is exposure to BMAA a risk factor for neurodegenerative diseases? A response to a critical review of the BMAA hypothesis. Neurotox. Res. 2021, 39, 81–106. [Google Scholar] [CrossRef]
- Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Tien Do, D.; Simon, D.F.; Sauveґ, S. Analysis of the neurotoxin β-N-methylamino-L-alanine (BMAA) and isomers in surface water by FMOC derivatization liquid chromatography high resolution mass spectrometry. PLoS ONE 2019, 14, e0220698. [Google Scholar] [CrossRef]
- Banack, S.A. Second laboratory validation of β-N-methylamino-L-alanine, N-(2aminoethyl)glycine, and 2,4-diaminobuytric acid by ultra-performance liquid chromatography and tandem mass spectrometry. Neurotox. Res. 2020, 39, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Johnston, E.; Aberg, K.M.; Nilsson, U.; Ilag, L.L. Strategy for quantifying trace levels of BMAA in cyanobacteria by LC/MS/MS. Anal. Bioanal. Chem. 2013, 405, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Mabry, T.J.; Cao, P.; Moini, M. Identification of nonprotein amino acids from cycad seeds as N-ethoxycarbonyl ethyl ester derivatives by positive chemical-ionization gas chromatography-mass spectrometry. J. Chromatogr. A 1997, 787, 288–294. [Google Scholar] [CrossRef]
- Guo, T.; Geis, S.; Hedman, C.; Arndt, M.; Krick, W.; Sonzogni, W. Characterization of ethyl chloroformate derivative of beta-methylamino-L-alanine. J. Am. Soc. Mass. Spectr. 2007, 18, 817–825. [Google Scholar] [CrossRef][Green Version]
- Baptista, M.S.; Cianca, R.C.; Lopes, V.R.; Almeida, C.M.; Vasconcelos, V.M. Determination of the nonprotein amino acid beta-N-methylamino-L-alanine in estuarine cyanobacteria by capillary electrophoresis. Toxicon 2011, 58, 410–414. [Google Scholar] [CrossRef]
- Beach, D.G.; Kerrin, E.S.; Quilliam, M.A. Selective quantitation of the neurotoxin BMAA by use of hydrophilic-interaction liquid chromatography-differential mobility spectrometry-tandem mass spectrometry (HILIC-DMS-MS/MS). Anal. Bioanal. Chem. 2015, 407, 8397–8409. [Google Scholar] [CrossRef]
- Beach, D.G.; Kerrin, E.S.; Giddings, S.D.; Quilliam, M.A.; McCarron, P. Differential mobility-mass spectrometry double spike isotope dilution study of release of beta-methylaminoalanine and proteinogenic amino acids during biological sample hydrolysis. Sci. Rep. 2018, 8, 117. [Google Scholar] [CrossRef]
- Moura, S.; De Almeida Ultramari, M.; Mendes Louzada de Paula, D.; Yonamine, M.; Pinto, E. 1H NMR determination of β-N-methylamino-l-alanine (l-BMAA) in environmental and biological samples. Toxicon 2009, 53, 578–583. [Google Scholar] [CrossRef]
- Cohen, S.A. Analytical techniques for the detection of α-amino-β-methylaminopropionic acid. Analyst 2012, 137, 1991–2005. [Google Scholar] [CrossRef]
- Masseret, E.; Banack, S.A.; Boumediene, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Detection of BMAA in the marine environment of an ALS cluster in Southern France. PLoS ONE 2013, 8, e83406. [Google Scholar]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T.G. The effect of β-N-methylamino-L-alanine (BMAA) on oxidative stress response enzymes of the macrophyte Ceratophyllum demersum. Toxicon 2011, 57, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Salomonsson, M.; Hansson, A.; Bondesson, U. Development and in-house validation of a method for quantification of BMAA in mussels using dansyl chloride derivatization and ultra-performance liquid chromatography tandem mass spectrometry. Anal. Methods 2013, 5, 4865–4874. [Google Scholar] [CrossRef]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The fate of the cyanobacterial toxin β-N-methylamino-L-alanine in freshwater mussels. Ecotoxicol. Environ. Saf. 2014, 101, 51–58. [Google Scholar] [CrossRef]
- Zurita, J.; Zguna, N.; Andrýs, R.; Strzelczak, A.; Jiang, L.; Thorsen, G.; Ilag, L.L. Chiral analysis of β-methylamino alanine (BMAA) enantiomers after (+)-1-(9-fluorenyl)-ethyl chloroformate (FLEC) derivatization and LC-MS/MS. Anal. Methods 2019, 11, 432–442. [Google Scholar] [CrossRef]
- Banack, S.A.; Metcalf, J.S.; Spacil, Z.; Downing, T.G.; Downing, S.; Long, A.; Nunn, P.B.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin beta-N-methylamino-L-alanine (BMAA) from other diamino acids. Toxicon 2011, 57, 730–738. [Google Scholar] [CrossRef]
- Lage, S.; Burian, A.; Rasmussen, U.; Costa, P.R.; Annadotter, H.; Godhe, A.; Rydberg, S. BMAA extraction of cyanobacteria samples: Which method to choose? Environ. Sci. Pollut. Res. Int. 2016, 23, 338–350. [Google Scholar] [CrossRef]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-methylamino-l-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef]
- Davis, D.A.; Cox, P.A.; Banack, S.A.; Lecusay, P.D.; Garamszegi, S.P.; Hagan, M.J.; Powell, J.T.; Metcalf, J.S.; Palmour, R.M.; Beierschmitt, A.; et al. L-serine reduces spinal cord pathology in a vervet model of preclinical ALS/MND. J. Neuropathol. Exp. Neurol. 2020, 79, 393–406. [Google Scholar] [CrossRef]
- Weiss, J.H.; Christine, C.W.; Choi, D.W. Bicarbonate dependence of glutamate receptor activation by beta-N-methylamino-L-alanine: Channel recording and study with related compounds. Neuron 1989, 3, 321–326. [Google Scholar] [CrossRef]
- Weiss, J.H.; Koh, J.-Y.; Choi, D.W. Neurotoxicity of β-N-methylamino-L-alanine (BMAA) and β-N-oxalylamino-L-alanine (BOAA) on cultured cortical neurons. Brain Res. 1989, 497, 64–71. [Google Scholar] [CrossRef]
- Seawright, A.A.; Brown, A.W.; Nolan, C.C.; Cavanagh, J.B. Selective degeneration of cerebellar cortical neurons caused by cycad neurotoxin, L-β-methylaminoalanine (L-BMAA), in rats. Neuropathol. Appl. Neurobiol. 1990, 16, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Purdie, E.L.; Samsudin, S.; Eddy, F.B.; Codd, G.A. Effects of the cyanobacterial neurotoxin β-N-methylamino-L-alanine on the early-life stage development of zebrafish (Danio rerio). Aquat. Toxicol. 2009, 95, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Purdie, E.L.; Metcalf, J.S.; Kashmiri, S.; Codd, G.A. Toxicity of the cyanobacterial neurotoxin β -N-methylamino-L-alanine to three aquatic animal species. Amyotroph. Lateral Scler. 2009, 10, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Escala, W.; Papapetropoulos, S.; Bradley, W.G.; Zhai, R.G. BMAA neurotoxicity in Drosophila. Amyotroph Lateral Scler. 2009, 10 (Suppl. S2), 61–66. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Berg, A.L.; Lindstrom, A.K.; Hanrieder, J.; Arnerup, G.; Roman, E.; Bergquist, J.; Lindquist, N.G.; Brittebo, E.B.; Andersson, M. Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression, and neurodegeneration in adult hippocampus. Toxicol. Sci. 2012, 130, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.Z.; Yu, S.; Hsu, C.-I.; Liu, J.; Acab, A.; Wu, R.; Tao, A.; Chiang, B.J.; Weiss, J.H. Intrathecal infusion of BMAA induces selective motor neuron damage and astrogliosis in the ventral horn of the spinal cord. Exp. Neurol. 2014, 261, 1–9. [Google Scholar] [CrossRef]
- Wang, S.; Qiu, J.; Zhao, M.; Li, F.; Yu, R.; Li, A. Accumulation and distribution of neurotoxin BMAA in aquatic animals and effect on the behavior of zebrafish in a T-maze test. Toxicon 2020, 173, 39–47. [Google Scholar] [CrossRef]
- Nunn, P.B. Three phases of research on β-N-methylamino-L-alanine (BMAA)—A neurotoxic amino acid. Amyotroph. Lateral Scler. 2009, 10 (Suppl. S2), 26–33. [Google Scholar] [CrossRef]
- De Munck, E.; Muñoz-Sáez, E.; Miguel, B.G.; Solas, M.T.; Martínez, A.; Arahuetes, R.M. β-N-methylamino-l-alanine causes neurological and pathological phenotypes mimicking Amyotrophic Lateral Sclerosis (ALS): The first step towards an experimental model for sporadic ALS. Environ. Toxicol. Pharmacol. 2013, 36, 243–255. [Google Scholar] [CrossRef]
- De Munck, E.; Muñoz-Sáez, E.; Miguel, B.G.; Solas, M.T.; Martínez, A.; Arahuetes, R.M. Morphometric and neurochemical alterations found in l-BMAA treated rats. Environ. Toxicol. Pharmacol. 2015, 39, 1232–1245. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Wiegand, C.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) uptake by the animal model, Daphnia magna and subsequent oxidative stress. Toxicon 2015, 100, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sáez, E.; de Munck García, E.; Arahuetes Portero, R.M.; Vicente, F.; Ortiz-López, F.J.; Cantizani, J.; Miguel, B.G. Neuroprotective role of sphingosine-1-phosphate in L-BMAA treated neuroblastoma cells (SH-SY5Y). Neurosci. Lett. 2015, 593, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sáez, E.; de Munck García, E.; Arahuetes Portero, R.M.; Martínez, A.; Solas Alados, M.T.; Miguel, B.G. Analysis of β-N-methylamino-l-alanine (L-BMAA) neurotoxicity in rat cerebellum. NeuroToxicology 2015, 48, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Kim, K.; Oh, Y.; Yoon, K.S.; Baik, H.H.; Kim, S.S.; Ha, J.; Kang, I.; Choe, W. Neurotoxin β-N-methylamino-L-alanine induces endoplasmic reticulum stress-mediated neuronal apoptosis. Mol. Med. Rep. 2016, 14, 4873–4880. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef]
- Brooke-Jones, M.; Gáliková, M.; Dircksen, H. Cyanobacterial neurotoxin beta-methyl-amino-l-alanine affects dopaminergic neurons in optic ganglia and brain of Daphnia magna. Toxins 2018, 10, 527. [Google Scholar] [CrossRef]
- Brenner, E.D.; Martinez-Barboza, N.; Clark, A.P.; Liang, Q.S.; Stevenson, D.W.; Coruzzi, G.M. Arabidopsis mutants resistant to S(+)-beta-methyl-alpha, beta-diaminopropionic acid, a cycad-derived glutamate receptor agonist. Plant Physiol. 2000, 124, 1615–1624. [Google Scholar] [CrossRef]
- Brenner, E.D.; Stahlberg, R.; Mancuso, S.; Vivanco, J.; Baluška, F.; Van Volkenburgh, E. Plant neurobiology: An integrated view of plant signaling. Trends Plant Sci. 2006, 11, 413–419. [Google Scholar] [CrossRef]
- Brenner, E.D.; Feinberg, P.; Runko, S.; Coruzzi, G.M. A mutation in the Proteosomal Regulatory Particle AAA-ATPase-3 in Arabidopsis impairs the light-specific hypocotyl elongation response elicited by a glutamate receptor agonist, BMAA. Plant Mol Biol. 2009, 70, 523–533. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Pflugmacher, S.; Downing, T.G. β-N-Methylamino-l-alanine (BMAA) uptake by the aquatic macrophyte Ceratophyllum demersum. Ecotoxicol. Environ. Saf. 2011, 74, 74–77. [Google Scholar] [CrossRef]
- Downing, S.; Esterhuizen-Londt, M.; Grant Downing, T. β-N-methylamino-L-alanine (BMAA) metabolism in the aquatic macrophyte Ceratophyllum demersum. Ecotoxicol. Environ. Saf. 2015, 120, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Ström, L.; Godhe, A.; Rydberg, S. The effect of exogenous β-N-methylamino-l-alanine (BMAA) on the diatoms Phaeodactylum tricornutum and Thalassiosira weissflogii. Harmful Algae 2016, 58, 85–92. [Google Scholar] [CrossRef] [PubMed]
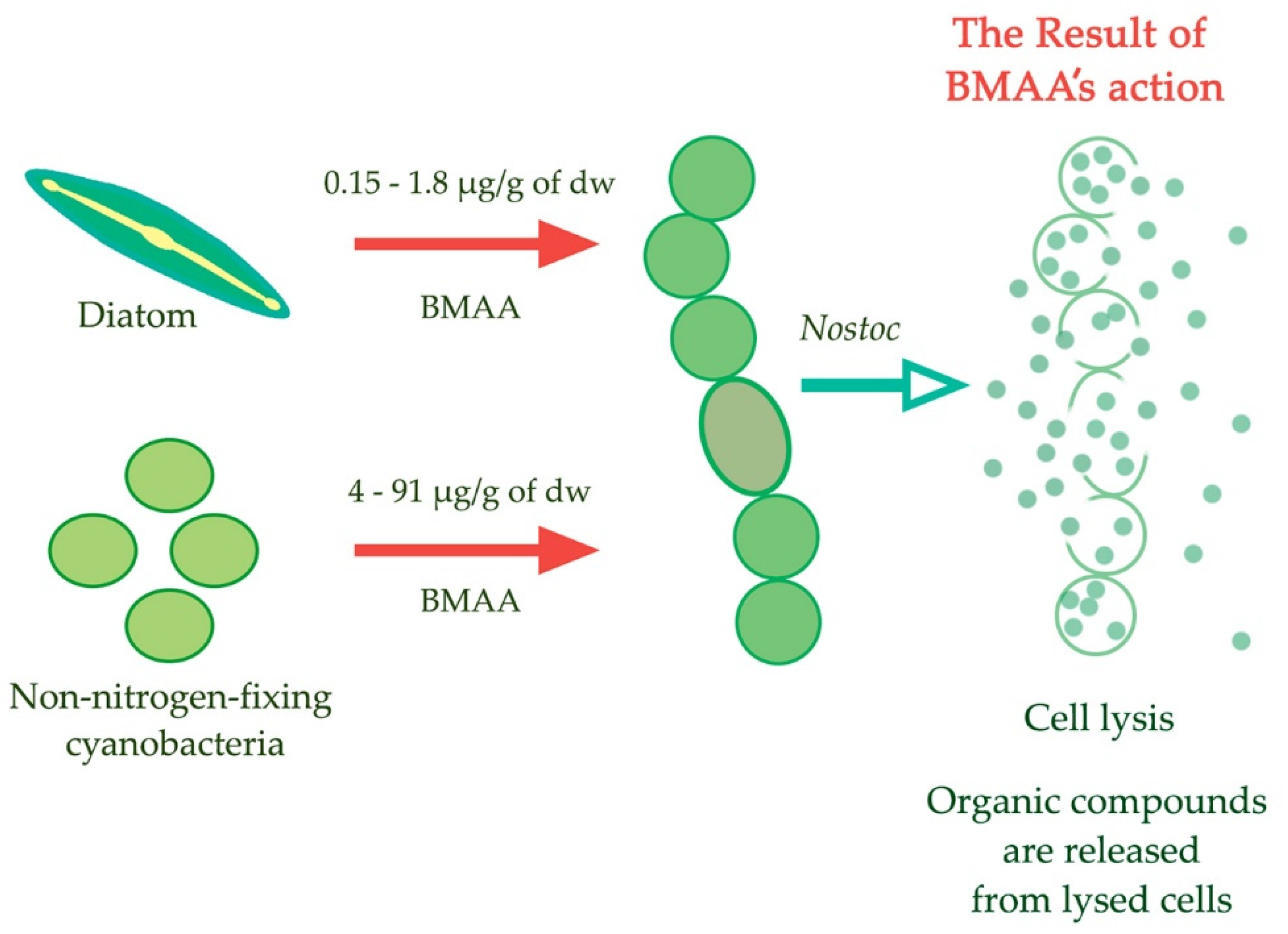
- Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA inhibits nitrogen fixation in the cyanobacterium Nostoc sp. PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.; Rasmussen, U.; Semashko, T.; Govorun, V.; Koksharova, O. Stress effects of cyanotoxin β-methylamino-L-alanine (BMAA) on cyanobacterial heterocyst formation and functionality Environ. Microbiol. Rep. 2018, 10, 369–377. [Google Scholar] [CrossRef]
- Popova, A.; Semashko, T.; Kostina, N.; Rasmussen, U.; Govorun, V.; Koksharova, O. The cyanotoxin BMAA induces heterocyst specific gene expression in Anabaena sp. PCC 7120 under repressive conditions. Toxins 2018, 10, 478. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Butenko, I.O.; Pobeguts, O.V.; Safronova, N.A.; Govorun, V.M. The first proteomic study of Nostoc sp. PCC 7120 exposed to cyanotoxin BMAA under nitrogen starvation. Toxins 2020, 12, 310. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Butenko, I.O.; Pobeguts, O.V.; Safronova, N.A.; Govorun, V.M. Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC7120 under BMAA Treatment. Toxins 2020, 12, 372. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Butenko, I.O.; Pobeguts, O.V.; Safronova, N.A.; Govorun, V.M. β-N-Methylamino-L-Alanine (BMAA) Causes Severe Stress in Nostoc sp. PCC 7120 Cells under Diazotrophic Conditions: A Proteomic Study. Toxins 2021, 13, 325. [Google Scholar] [CrossRef]
- Vergou, Y.; Touraki, M.; Paraskevopoulou, A.; Triantis, T.M.; Hiskia, A.; Gkelis, S. β-Ν-Methylamino-L-alanine interferes with nitrogen assimilation in the cyanobacterium, non-BMAA producer, Synechococcus sp. TAU-MAC 0499. Toxicon 2020, 185, 147–155. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Wang, S.; Zhang, J.-Y.; Lin, G.-M.; Gan, N.; Song, L.; Zeng, X.; Zhang, C.-C. The Proposed Neurotoxin β-N-Methylamino-L-Alanine (BMAA) Is Taken up through Amino-Acid Transport Systems in the Cyanobacterium Anabaena PCC 7120. Toxins 2020, 12, 518. [Google Scholar] [CrossRef]
- Banack, S.A.; Caller, T.A.; Stommel, E.W. The cyanobacteria derived toxin beta-N-methylamino-L-alanine and amyotrophic lateral sclerosis. Toxins 2010, 2, 2837–2850. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Basile, M.; Mash, D.C. Cerebral uptake and protein incorporation of cyanobacterial toxin β-N-methylamino-L-alanine. Neuroreport 2013, 24, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.B.; Mash, D.C.; Murch, S.J. The natural non-protein amino acid N-β-methylamino-L-alanine (BMAA) is incorporated into protein during synthesis. Amino Acids 2014, 46, 2553–2559. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Bergquist, J.; Andersson, M. Quality measures of imaging mass spectrometry aids in revealing long-term striatal protein changes induced by neonatal exposure to the cyanobacterial toxin β-N-methylamino-L-alanine (BMAA). Mol. Cell Proteom. 2014, 13, 93–104. [Google Scholar] [CrossRef]
- Van Onselen, R.; Downing, T.G. BMAA-protein interactions: A possible new mechanism of toxicity. Toxicon 2018, 143, 74–80. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The non-protein amino acid BMAA is misincorporated into human proteins in place of L-serine causing protein misfolding and aggregation. PLoS ONE 2013, 8, e0075376. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Guillemin, G.J. The Cyanotoxin and non-protein amino acid β-methylamino-L-alanine (L-BMAA) in the food chain: Incorporation into proteins and its impact on human health. Neurotox. Res. 2019, 36, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Proctor, E.A.; Mowrey, D.D.; Dokholyan, N.V. β-Methylamino-L-alanine substitution of serine in SOD1 suggests a direct role in ALS etiology. PLOS Comput. Biol. 2019, 15, e1007225. [Google Scholar] [CrossRef]
- Korn, A.; Höfling, C.; Zeitschel, U.; Krueger, M.; Roßner, S.; Huster, D. Incorporation of the Non-Proteinaceous Amino Acid β-Methyl-Amino-Alanine Affects Amyloid β Fibril Properties and Toxicity. ACS Chem. Neurosci. 2020, 11, 1038–1047. [Google Scholar] [CrossRef]
- Han, N.C.; Bullwinkle, T.J.; Loeb, K.F.; Faull, K.F.; Mohler, K.; Rinehart, J.; Ibba, M. The mechanism of β-N-methylamino-L-alanine inhibition of tRNA aminoacylation and its impact on misincorporation. J. Biol. Chem. 2020, 295, 1402–1410. [Google Scholar] [CrossRef]
- Beri, J.; Nash, T.; Martin, R.M.; Bereman, M.S. Exposure to BMAA mirrors molecular processes linked to neurodegenerative disease. Proteomics 2017, 17, 1700161. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhang, Y.; Inoue, H.; Yogo, S.; Hirasawa, E. Purification and molecular analysis of a monoamine oxidase isolated from Narcissus tazetta. Biosci. Biotechnol. Biochem. 2013, 77, 1728–1733. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.F.; Candeias, E.; Esteves, A.R.; Magalhães, J.D.; Ferreira, I.L.; Nunes-Costa, D.; Rego, A.C.; Empadinhas, N.; Cardoso, S.M. Microbial BMAA elicits mitochondrial dysfunction, innate immunity activation, and Alzheimer’s disease features in cortical neurons. J. Neuroinflamm. 2020, 17, 332. [Google Scholar] [CrossRef]
- Soto, T.; Buzzi, E.D.; Rotstein, N.P.; German, O.L.; Politi, L.E. Damaging effects of BMAA on retina neurons and Müller glial cells. Exp. Eye Res. 2021, 202, 108342. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; O’Brien, P.; Pettit, L.D.; Pyburn, S.I. Complexes of zinc, copper, and nickel with the nonprotein amino acid L-α-amino-β-methylaminopropionic acid: A naturally occurring neurotoxin. J. Inorg. Biochem. 1989, 37, 175–183. [Google Scholar] [CrossRef]
- Weiss, J.H.; Sensi, S.L. Ca2+,Zn2+ permeable AMPA or kainate receptors: Possible key factors in selective neurodegeneration. Trends Neurosci. 2000, 23, 365–371. [Google Scholar] [CrossRef]
- Pochwat, B.; Nowak, G.; Szewczyk, B. Relationship between Zinc (Zn2+) and Glutamate Receptors in the Processes Underlying Neurodegeneration. Neural Plast. 2015, 2015, 591563. [Google Scholar] [CrossRef]
- Li, X.; Du, X.; Ni, J. Zn2+ Aggravates Tau Aggregation and Neurotoxicity. Int. J. Mol. Sci. 2019, 20, 487. [Google Scholar] [CrossRef]
- Diaz-parga, P.; Goto, J.J.; Krishnan, V.V. On the differential roles of Mg2+, Zn2+, and Cu2+ in the equilibrium of β-N-methyl-amino-L-alanine (BMAA) and its carbamates. Neurotox. Res. 2020, 39, 6–16. [Google Scholar] [CrossRef]
- Lepoutre, A.; Milliote, N.; Bonnard, M.; Palos-Ladeiro, M.; Rioult, D.; Bonnard, I.; Bastien, F.; Faassen, E.; Geffard, A.; Lance, E. Genotoxic and Cytotoxic Effects on the Immune Cells of the Freshwater Bivalve Dreissena polymorpha Exposed to the Environmental Neurotoxin BMAA. Toxins 2018, 10, 106. [Google Scholar] [CrossRef]
- Gerić, M.; Gajski, G.; Domijan, A.-M.; Garaj-Vrhovac, V.; Filipič, M.; Žegura, B. Genotoxic effects of neurotoxin ß-N-methylamino-L-alanine in human peripheral blood cells. Chemosphere 2018, 214, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Spencer, P.S.; Palmer, V.S.; Kisby, G.E. Cycad β-N-methylamino-L-alanine (BMAA), methylazoxymethanol, genotoxicity, and neurodegeneration. Toxicon. 2018, 155, 49–50. [Google Scholar] [CrossRef] [PubMed]
- Pierozan, P.; Cattani, D.; Karlsson, O. Hippocampal neural stem cells are more susceptible to the neurotoxin BMAA than primary neurons: Effects on apoptosis, cellular differentiation, neurite outgrowth, and DNA methylation. Cell Death Dis. 2020, 11, 910. [Google Scholar] [CrossRef] [PubMed]
- Polsky, F.I.; Nunn, P.B.; Bell, E.A. Distribution and toxicity of alpha-amino-beta-methylaminopropionic acid. Fed. Proc. 1972, 31, 1473–1475. [Google Scholar]
- Rodgers, K.J. Non-protein amino acids and neurodegeneration: The enemy within. Exp. Neurol. 2014, 253, 192–196. [Google Scholar] [CrossRef]
- Davis, D.A.; Garamszegi, S.P.; Banack, S.A.; Dooley, P.D.; Coyne, T.M.; McLean, D.W.; Rotstein, D.S.; Mash, D.C.; Cox, P.A. BMAA, Methylmercury, and Mechanisms of Neurodegeneration in Dolphins: A Natural Model of Toxin Exposure. Toxins 2021, 13, 697. [Google Scholar] [CrossRef]
- Duncan, M.W.; Villacreses, N.E.; Pearson, P.G.; Wyatt, L.; Rapoport, S.I.; Kopin, I.J.; Markey, S.P.; Smith, Q.R. 2-Amino-3-(methylamino)-propanoic acid (BMAA) pharmacokinetics and blood-brain barrier permeability in the rat. J. Pharmacol. Exp. Ther. 1991, 258, 27–35. [Google Scholar]
- Berntzon, L.; Ronnevi, L.O.; Bergman, B.; Eriksson, J. Detection of BMAA in the human central nervous system. Neuroscience 2015, 292, 137–147. [Google Scholar] [CrossRef]
- Frøyset, A.K.; Khan, E.A.; Fladmark, K.E. Quantitative proteomics analysis of zebrafish exposed to sub-lethal dosages of β-methyl-amino-L-alanine (BMAA). Sci. Rep 2016, 6, 29631. [Google Scholar] [CrossRef]
- Goto, J.J.; Koenig, J.H.; Ikeda, K. The physiological effect of ingested β-N-methylamino-L-alanine on a glutamatergic synapse in an in vivo preparation. Comp. Biochem. Physiol. C Toxicol. Pharm. 2012, 156, 171–177. [Google Scholar] [CrossRef]
- Okle, O.; Rath, L.; Galizia, C.G.; Dietrich, D.R. The cyanobacterial neurotoxin beta-N-methylamino-l-alanine (BMAA) induces neuronal and behavioral changes in honeybees. Toxicol. Appl. Pharm. 2013, 270, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Rakonczay, Z.; Matsuoka, Y.; Giacobini, E. Effects of L-beta-N-methylamino-Lalanine (L-BMAA) on the cortical cholinergic and glutamatergic systems of the rat. J Neurosci Res 1991, 29, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.N.; Omelchenko, I.; Ross, S.M.; Spencer, P. The neurotoxin, β-N-methylamino-l-alanine (BMAA) interacts with the strychnine-insensitive glycine modulatory site of the N-methyl-d-aspartate receptor. Neuropharmacology 1995, 34, 651–658. [Google Scholar] [CrossRef]
- Brownson, D.M.; Mabry, T.J.; Leslie, S.W. The cycad neurotoxic amino acid, beta-N-methylamino-L-alanine (BMAA), elevates intracellular calcium levels in dissociated rat brain cells. J. Ethnopharmacol. 2002, 82, 159–167. [Google Scholar] [CrossRef]
- Rao, S.D.; Banack, S.A.; Cox, P.A.; Weiss, J.H. BMAA selectively injures motor neurons via AMPA/kainate receptor activation. Exp. Neurol. 2006, 201, 244–252. [Google Scholar] [CrossRef]
- Lopicic, S.; Nedeljkov, V.; Cemerikic, D. Augmentation and ionic mechanism of effect of beta-N-methylamino-L-alanine in presence of bicarbonate on membrane potential of Retzius nerve cells of the leech Haemopis sanguisuga. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 153, 284–292. [Google Scholar] [CrossRef]
- Chiu, A.; Gehringer, M.; Braidy, N.; Guillemin, G.; Welch, J.H.; Neilan, B.A. Gliotoxicity of the cyanotoxin, β-methyl-amino-L-alanine (BMAA). Sci. Rep. 2013, 3, 1482. [Google Scholar] [CrossRef]
- Dennison, K.L.; Spalding, E.P. Glutamate-Gated Calcium Fluxes in Arabidopsis. Plant Physiol. 2000, 124, 1511–1514. [Google Scholar] [CrossRef]
- Davenport, R. Glutamate receptors in plants. Ann. Bot. 2002, 90, 549–557. [Google Scholar] [CrossRef]
- Kwaaitaal, M.; Huisman, R.; Maintz, J.; Reinstädler, A.; Panstruga, R. Ionotropic glutamate receptor (iGluR)-like channels mediate MAMP-induced calcium influx in Arabidopsis thaliana. Biochem. J. 2011, 440, 355–373. [Google Scholar] [CrossRef]
- Price, M.B.; Jelesko, J.; Okumoto, S. Glutamate Receptor Homologs in Plants: Functions and Evolutionary Origins. Front. Plant Sci. 2012, 3, 235. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G.; Roberts, M.R. Glutamate receptor-like channels in plants: A role as amino acid sensors in plant defence? F1000Prime Rep. 2014, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- De Bortoli, S.; Teardo, E.; Szabò, I.; Morosinotto, T.; Alboresi, A. Evolutionary insight into the ionotropic glutamate receptor superfamily of photosynthetic organisms. Biophys. Chem. 2016, 218, 14–26. [Google Scholar] [CrossRef]
- Li, Y.H.; Yu, X.Z.; Mo, L.Y.; Lin, Y.J.; Zhang, Q. Involvement of glutamate receptors in regulating calcium influx in rice seedlings under Cr exposure. Ecotoxicology 2019, 28, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Lazebny, O.E.; Lazebnaya, I.V.; Koksharova, O.A. Phylogenetic Analysis of Cyanobacterial Glutamate-like Receptors: The First Overlook. In Proceedings of the 5th Moscow International Conference “Molecular Phylogenetics and Biodiversity Biobanking”, Moscow, Russia, 25–28 August 2018; A. Troitsk, A., Rusin, L., Eds.; p. 85. [Google Scholar] [CrossRef]
- Van Onselen, R.; Cook, N.A.; Phelan, R.R.; Downing, T.G. Bacteria do not incorporate β-N-methylamino-l-alanine into their proteins. Toxicon 2015, 102, 55–61. [Google Scholar] [CrossRef]
- Novak, M.; Hercog, K.; Žegura, B. Assessment of the mutagenic and genotoxic activity of cyanobacterial toxin beta-N-methyl-amino-L-alanine in Salmonella typhimurium. Toxicon 2016, 118, 134–140. [Google Scholar] [CrossRef]
- Downing, S.; Van de Venter, M.; Downing, T.G. The effect of exogenous β-N-methylamino-L-alanine on the growth of Synechocystis PCC 6803. Microb. Ecol 2012, 63, 149–156. [Google Scholar] [CrossRef]
- Richter, K.E.; Mena, E.E. L-beta-methylaminoalanine inhibits [3H]glutamate binding in the presence of bicarbonate ions. Brain Res. 1989, 492, 385–388. [Google Scholar] [CrossRef]
- Diaz-parga, P.; Goto, J.J.; Krishnan, V.V. Chemistry and chemical equilibrium dynamics of BMAA and its carbamate adducts. Neurotox. Res. 2018, 33, 76–86. [Google Scholar] [CrossRef]
- Nunn, P.B.; Ponnusamy, M. β-N-methylaminoalanine (BMAA): Metabolism and metabolic effects in model systems and in neural and other tissues of the rat in vitro. Toxicon 2009, 54, 85–94. [Google Scholar] [CrossRef]
- Liu, X.; Rush, T.; Zapata, J.; Lobner, D. β-N-methylamino-L-alanine induces oxidative stress and glutamate release through action on system Xc(−). Exp. Neurol 2009, 217, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Stavans, J.; Flores, E. The multicellular nature of filamentous heterocyst-forming cyanobacteria. FEMS Microbiol. Rev. 2016, 40, 831–854. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbio. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Kaneko, T.; Nakamura, Y.; Wolk, C.P.; Kuritz, T.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Ishikawa, A.; Kawashima, K.; Kimura, T.; et al. Complete genomic sequence of the filamentous nitrogen-fixing Cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. 2001, 8, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Castenholz, R.W. Phylum BX. Cyanobacteria (Oxygenic Photosynthetic Bacteria). Bergey’s Manual® of Systematic Bacteriology, Vol. 1: The Archaea and the Deeply Branching and Phototrophic Bacteria; Springer: New York, NY, USA, 2001; pp. 473–599. [Google Scholar]
- Singh, H.N.; Rai, U.N.; Rao, V.V.; Bagchi, S.N. Evidence for ammonia as an inhibitor of heterocyst and nitrogenase formation in the cyanobacterium Anabaena cycadeae. Biochem. Biophys. Res. Commun. 1983, 111, 180–187. [Google Scholar] [CrossRef]
- Olney, J.W.; Zorumski, C.; Price, M.T.; Labruyere, J. L-cysteine, a bicarbonate-sensitive endogenous excitotoxin. Science 1990, 248, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Flory, W.; Koeppe, R.E. Variation of neurotoxicity of L- and D-2,4-diaminobutyric acid with route of administration. Toxicol. Appl. Pharm. 1972, 23, 334–338. [Google Scholar] [CrossRef]
- Rawson, D.M. The effects of exogenous amino-acids on growth and nitrogenase activity in the cyanobacterium Anabaena cylindrica PCC 7122. J. Gen. Microbiol. 1985, 131, 2549–2554. [Google Scholar] [CrossRef]
- Weiss, J.H.; Choi, D.W. β-N-methylamino-L-alanine neurotoxicity: Requirement for bicarbonate as a cofactor. Science 1988, 241, 973–975. [Google Scholar] [CrossRef]
- Forchhammer, K.; Selim, K.A. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 2019, 44, 33–53. [Google Scholar] [CrossRef]
- Fessenden, R.J.; Fessenden, J.S. Organic Chemistry; Fessenden, J.S., Ed.; Brooks/Cole Publishing Company: Belmont, CA, USA, 1998. [Google Scholar]
- Gubisne-Haberle, D.; Hill, W.; Kazachkov, M.; Richardson, J.S.; Yu, P.H. Protein cross-linkage induced by formaldehyde derived from semicarbazide-sensitive amine oxidase-mediated deamination of methylamine. J. Pharmacol. Exp. Ther. 2012, 310, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Boomsma, F.; Van Dijk, J.; Bhaggoe, U.M.; Bouhuizen, A.M.; Van den Meiracker, A.H. Variation in semicarbazide-sensitive amine oxidase activity in plasma and tissues of mammals. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2000, 126, 69–78. [Google Scholar] [CrossRef]
- Percival, F.W.; Purves, W.K. Multiple amine oxidases in cucumber seedlings. Plant Physiol. 1974, 54, 601–607. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Takser, L.; Benachour, N.; Husk, B.; Cabana, H.; Gris, D. Cyanotoxins at low doses induce apoptosis and inflammatory effects in murine brain cells: Potential implications for neurodegenerative diseases. Toxicol. Rep. 2016, 3, 180–189. [Google Scholar] [CrossRef]
- Santucci, S.; Zsürger, N.; Chabry, J. β- N -methylamino-l-alanine induced in vivo retinal cell death. J. Neurochem. 2009, 109, 819–825. [Google Scholar] [CrossRef]
- Azam, F.; Worden, A.Z. Microbes, Molecules, and Marine Ecosystems. Science 2004, 303, 1622–1624. [Google Scholar] [CrossRef]
- Pohnert, G. Influence of Algal Secondary Metabolites on Plankton Community Structure. In Algal Chemical Ecology; Amsler, C.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Koksharova, O.A.; Safronov, N.A. The effects of bacterial secondary metabolites on photosynthesis in microalgae cells. Biophys. Rev. 2022. [Google Scholar] [CrossRef]
- Warshan, D.; Espinoza, J.L.; Stuart, R.K.; Richter, R.A.; Kim, S.-Y.; Shapiro, N.; Woyke, T.; Kyrpides, N.; Barry, K.; Singan, V.; et al. Feathermoss and epiphytic Nostoc cooperate differently: Expanding the spectrum of plant-cyanobacteria symbiosis. ISME J. 2017, 11, 2821–2833. [Google Scholar] [CrossRef]
- Kim, S.Y.; Rasmussen, U.; Rydberg, S. Effect and function of β-N-methylamino-L-alanine in the diatom Phaeodactylum tricornutum. Sci. Total Environ. 2022, 24, 154778. [Google Scholar] [CrossRef]
- Nieves-Morión, M.; Flores, E.; Foster, R.A. Predicting substrate exchange in marine diatom-heterocystous cyanobacteria symbioses. Environ. Microbiol. 2020, 22, 2027–2052. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bereman, M.S.; Marsden, K.C. Exposure to a mixture of BMAA and MCLR synergistically modulates behavior in larval zebrafish while exacerbating molecular changes related to neurodegeneration. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kazemi Shariat Panahi, H.; Dehhaghi, M.; Heng, B.; Lane, D.J.R.; Bush, A.I.; Guillemin, G.J.; Tan, V.X. Neuropathological Mechanisms of β-N-Methylamino-L-Alanine (BMAA) with a Focus on Iron Overload and Ferroptosis. Neurotox. Res. 2022, 40, 614–635. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Concentration of BMAA μg/g Dry Weight of Sample | Method of Identification | Reference |
|---|---|---|---|
| Cyanobacteria Nostoc | 0.3 | HPLC-MS | [9] |
| Cycad/cyanobacterial symbiosis | 37–1161 | HPLC-MS | [9] |
| the immature male sporangi of Cycas micronesica | 1546 | HPLC-MS | [9] |
| the outer integument layer of the cycas seed sarcotesta | 1161 | HPLC-MS | [9] |
| Flying Foxes | 3556 | HPLC-MS | [9] |
| brain tissues from the frontal cortex of six Chamorro patients | 6 | HPLC-MS | [9] |
| brain tissues from the frontal cortex of two Alzheimer’s patients from Canada | 6.6 | HPLC-MS | [9] |
| Azolla filiculoides with cyanobacterial symbionts in its leaves | 2 | HPLC-MS | [9] |
| Gunnera kauaiensi with cyanobacterial symbionts in its leaves | 4 | HPLC-MS | [9] |
| Nostoc PCC 9305 (symbiont of Anthoceros ) | 156 (free) 1400 (protein-bound) | HPLC-MS | [17] |
| Nostoc PCC 7422 (symbiont of Cycas) | 962 (protein-bound) | HPLC-MS | [17] |
| Nostoc 8001 (symbiont of Gunnera monoica) | 203 (free) 664 (protein-bound) | HPLC-MS | [17] |
| Microcystis PCC 7820 (Freshwater, Scotland) | 6 (free) 12 (protein-bound) | HPLC-MS | [17] |
| Chroococcidiopsis indica GQ2-7 (Marine coral) | 435 (free) 76 (protein-bound) | HPLC-MS | [17] |
| Chroococcidiopsis indica GT-3-26 (Marine rock) | 1306 (free) 5415 (protein-bound) | HPLC-MS | [17] |
| Cylindrospermopsis raciborskii CR3 (Freshwater, Australia) | 6478 (free) 14 (protein-bound) | HPLC-MS | [17] |
| Nostoc 268 (Brackish Water, Baltic Sea) | 34 (free) 274 (protein-bound) | HPLC-MS | [17] |
| Nostoc sp. CMMED 01 (Marine, Hawaiian Islands) | 1243 (free) 1070 (protein-bound) | HPLC-MS | [17] |
| Chlorogloeopsis PCC 6912 (Soil, India) | 758 (free) | HPLC-MS | [17] |
| Fischerella PCC 7521 (Yellowstone, hot spring, USA) | 44 (free) 175 (protein-bound) | HPLC-MS | [17] |
| Scytonema PCC 7110 (Limestone cave, Bermuda) | 1733 (protein-bound) | HPLC-MS | [17] |
| Mytilus edulis (common mussel, Sweden’s west coast) | 0.15–0.2 | HPLC-MS/MS | [23] |
| Ostrea edulis (common oyster, Sweden’s west coast ) | 0.006–0.14 | HPLC-MS/MS | [23] |
| Scophthalmus maximus (turbot) (Brain tissue) | 0.047–1.29 | HPLC-MS/MS | [23] |
| Microcystis PCC7806 and Synechocystis J341 | the relative abundance of labeled amino acids based on LC/MS/MS peak areas | stable isotope 15N and UPLC-MS/MS | [27] |
| Thalassiosira sp. (CCAP 1085/15) diatom | 0.0033 | UHPLC-MS/MS | [30] |
| Skeletonema marinoi isolates (SAAE 08603) diatom | 0.0011 | UHPLC-MS/MS | [30] |
| Phaeodactylum tricornutum diatom | 0.20–1.4 | HPLC-MS/MS | [31] |
| Cyclotella (diatom from Lake Liddell ) | 0.103 (free) 0.154 (protein-bound) | LC-MS/MS | [32] |
| Navicula (from Lostock Dam) | 0.151 (free) 0.369 (protein-bound) | LC-MS/MS | [32] |
| Cycas micronesica K.D. Hill Gametophyte Southern | 1.94 | GC-MS | [35] |
| Cycas micronesica K.D. Hill Gametophyte Nothern | 3.32 | GC-MS | [35] |
| Cycas micronesica K.D. Hill total plant Southern | 3.44 | GC-MS | [35] |
| Cycas micronesica K.D. Hill total plant Nothern | 4.9 | GC-MS | [35] |
| Blue mussel (Mytilus edulis) Sweden (west coast) | 0.08–0.9 | UHPLC-MS/MS | [39] |
| Oyster (Ostrea edulis) Greece | 0.32 | UHPLC-MS/MS | [39] |
| Oyster (Crassostrea gigas) France | 0.66 | UHPLC-MS/MS | [39] |
| Triticum aestivum (mature seeds) via irrigation | 217 ± 150 ng g FW−1 (protein-bound) | UHPLC-MS/MS | [43] |
| chicken tissues (i.e., muscle, liver, brain, and eye) | 0.0045–0.03 (free) 0.03–0.23 (protein-bound) | UHPLC-MS/MS | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koksharova, O.A.; Safronova, N.A. Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance. Toxins 2022, 14, 539. https://doi.org/10.3390/toxins14080539
Koksharova OA, Safronova NA. Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance. Toxins. 2022; 14(8):539. https://doi.org/10.3390/toxins14080539
Chicago/Turabian StyleKoksharova, Olga A., and Nina A. Safronova. 2022. "Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance" Toxins 14, no. 8: 539. https://doi.org/10.3390/toxins14080539
APA StyleKoksharova, O. A., & Safronova, N. A. (2022). Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance. Toxins, 14(8), 539. https://doi.org/10.3390/toxins14080539