Transcriptome Analysis Reveals MAPK/AMPK as a Key Regulator of the Inflammatory Response in PST Detoxification in Mytilus galloprovincialis and Argopecten irradians

 ,
,
Abstract
:
1. Introduction
2. Results
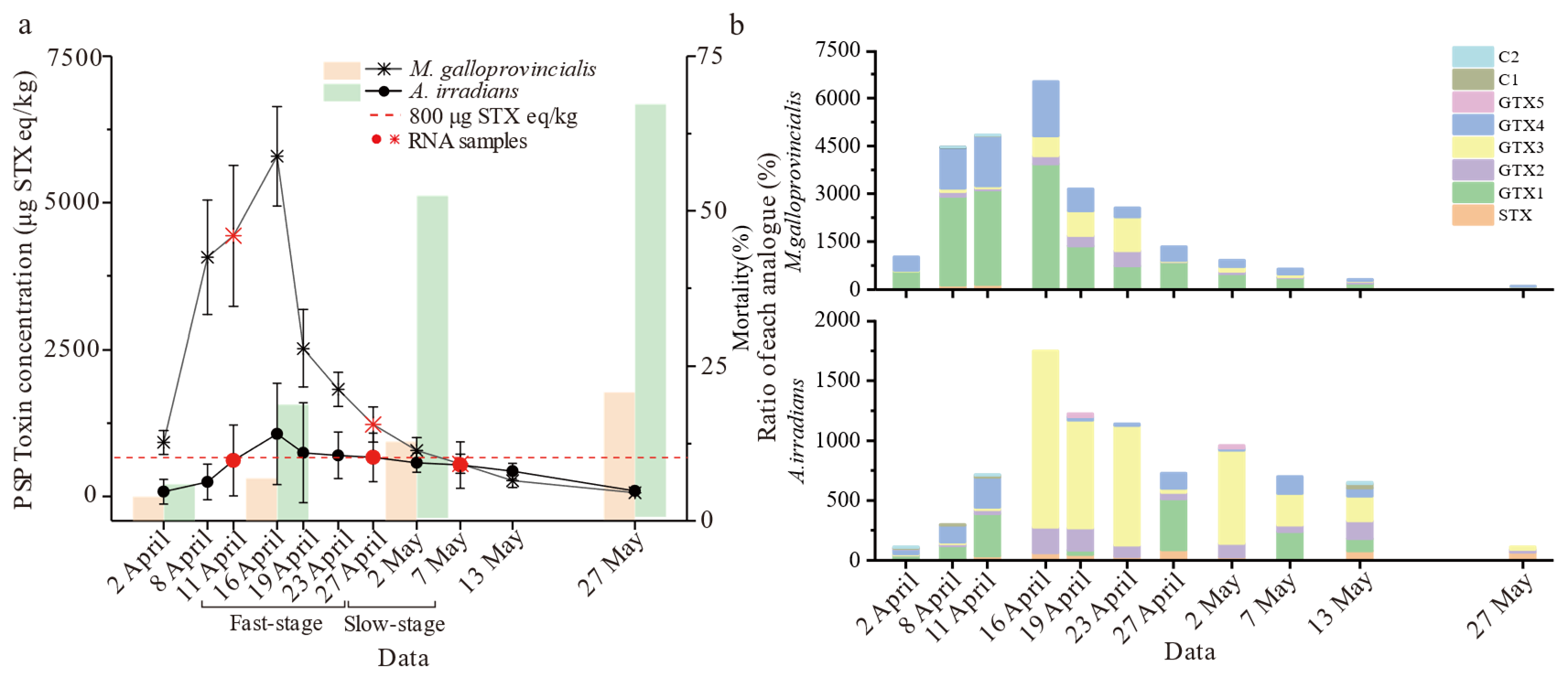
2.1. Analysis of PSTs Accumulation and Metabolism
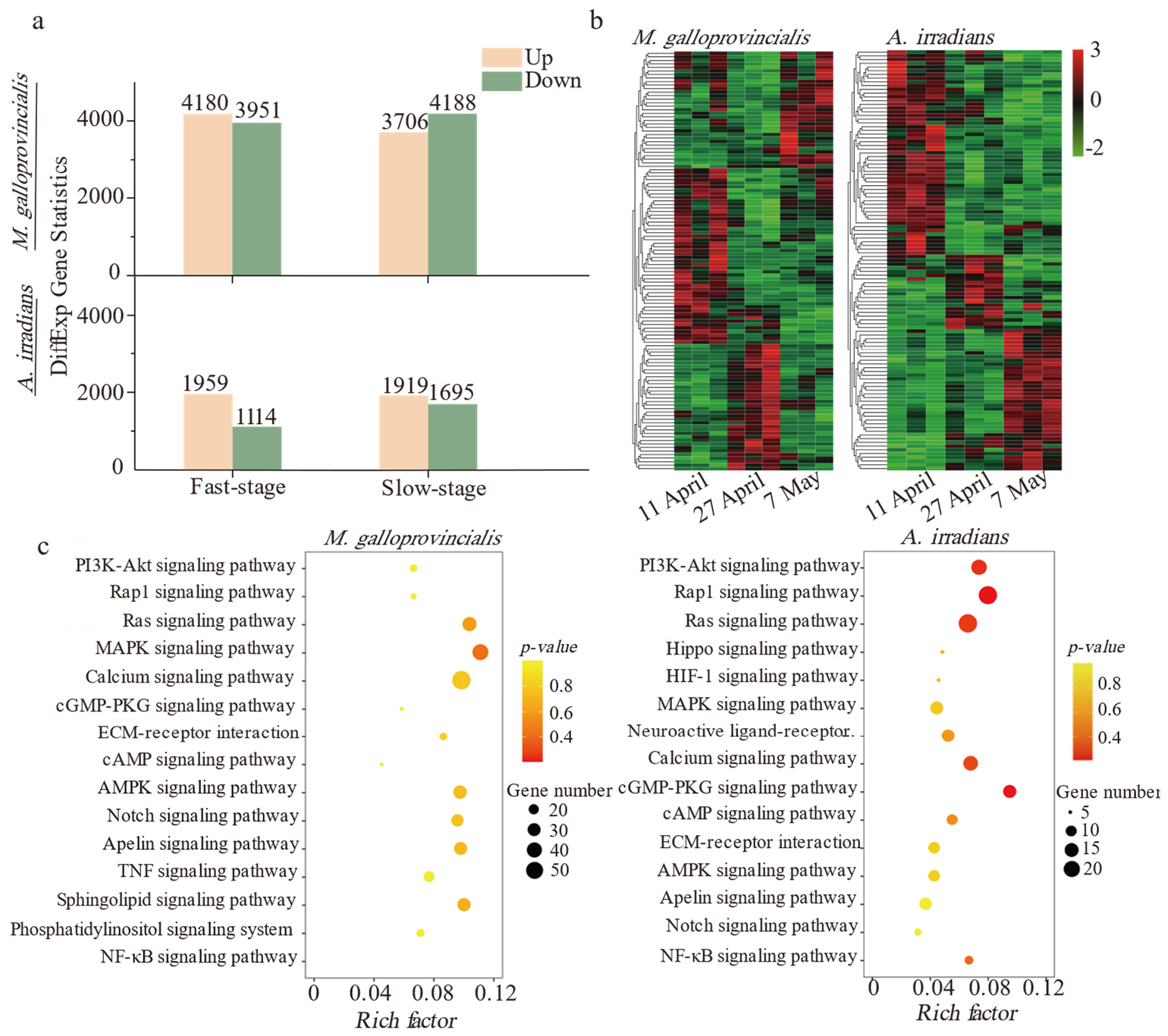
2.2. De Novo Assembly of Transcripts, Enrichment, and Annotation of Differential Genes
2.3. Mechanism of Inflammatory Responses Correlates with Elimination of PSTs
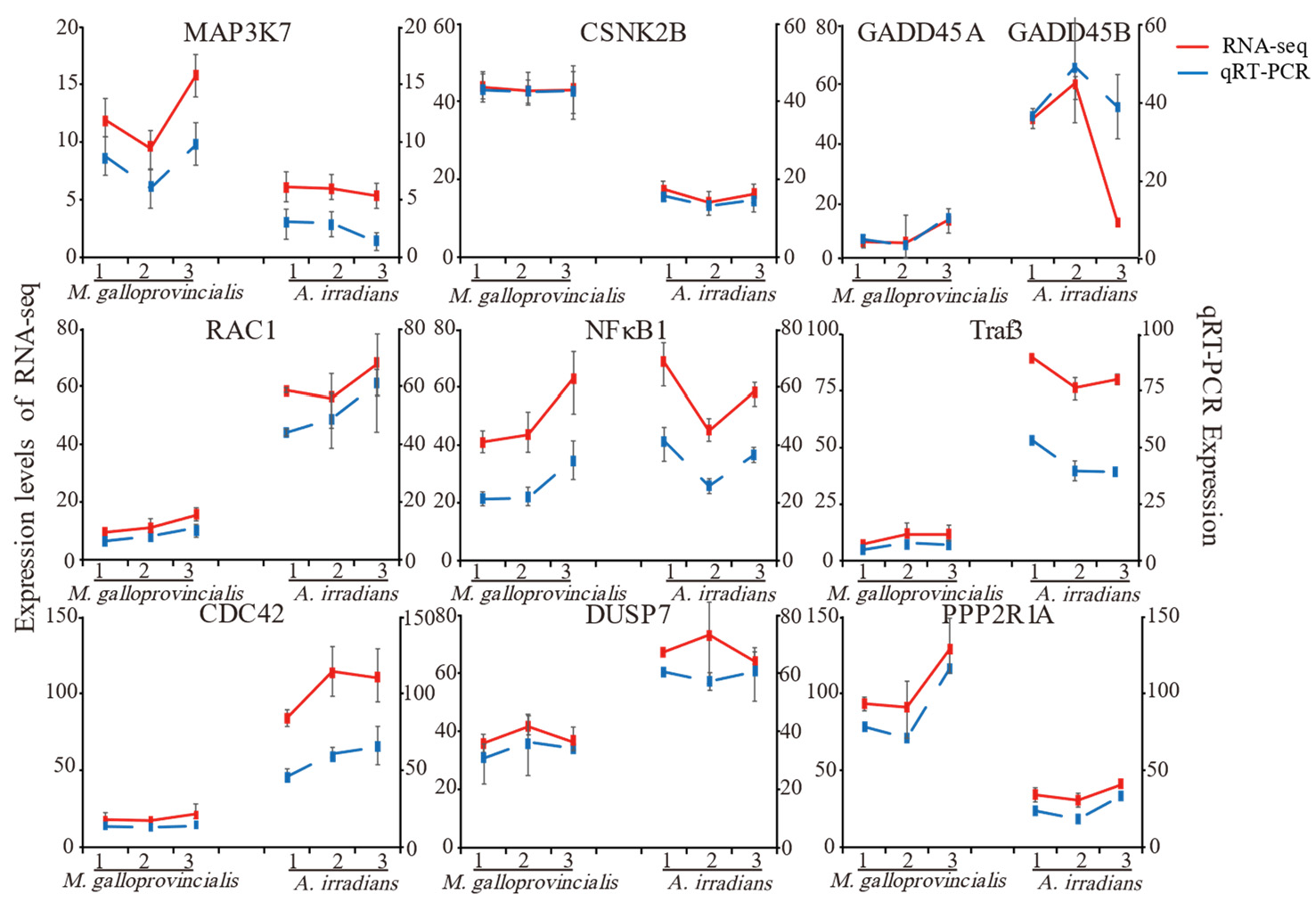
2.4. Validation of Important DEGs Related to the Inflammatory Reaction Using qRT-PCR
3. Discussion
3.1. Differences in Physiological Characteristics between M. galloprovincialis and A. irradians under PSTs Stress
3.2. Interference-Specific Differences in Inflammation between M. galloprovincialis and A. irradians under PSTs Stress
3.3. Importance of Adjusting the Immune Balance between Inflammation and Anti-Inflammatory Effects
4. Conclusions
5. Materials and Methods
5.1. Sample Collection and Preparation
5.2. Determination of PSTs Accumulation
5.3. LC-MS/MS Analysis
5.4. RNA Extraction, Library Construction, and Sequencing Analysis
5.5. Differentially Expressed Genes (DEGs) Analysis
5.6. Validation of Gene Expression by qRT-PCR
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lassudrie, M.; Hégaret, H.; Wikfors, G.H.; Silva, P. Effects of marine harmful algal blooms on bivalve cellular immunity and infectious diseases: A review. Dev. Comp. Immunol. 2020, 108, 103660. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Ye, R.-M.; Zheng, J.-W.; Luo, Z.-H.; Gu, H.-F.; Yang, W.-D.; Li, H.-Y.; Liu, J.-S. Molecular phylogeny and PSP toxin profile of the Alexandrium tamarense species complex along the coast of China. Mar. Pollut. Bull. 2014, 89, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Liu, T.; Vale, P.; Luo, Z. Morphology, phylogeny and toxin profiles of Gymnodinium inusitatum sp. nov., Gymnodinium catenatum and Gymnodinium microreticulatum (Dinophyceae) from the Yellow Sea, China. Harmful Algae 2013, 28, 97–107. [Google Scholar] [CrossRef]
- Wang, D.Z.; Lin, L.; Gu, H.F.; Chan, L.L.; Hong, H.S. Comparative studies on morphology, ITS sequence and protein profile of Alexandrium tamarense and A catenella isolated from the China Sea. Harmful Algae 2008, 7, 106–113. [Google Scholar] [CrossRef]
- Ding, L.; Qiu, J.; Li, A. Proposed biotransformation pathways for new metabolites of paralytic shellfish toxins based on field and experimental mussel samples. J. Agric. Food Chem. 2017, 65, 5494–5502. [Google Scholar] [CrossRef]
- Yu, R.C.; Zhang, Q.C.; Liu, Y.; Chen, Z.F.; Zhou, M.J. The dinoflagellate Alexandrium catenella producing only carbamate toxins may account for the seafood poisonings in Qinhuangdao, China. Harmful Algae 2021, 103, 101980. [Google Scholar] [CrossRef]
- Andres, J.K.; Yñiguez, A.T.; Maister, J.M.; Turner, A.D.; Olano, D.E.B.; Mendoza, J.; Salvador-Reyes, L.; Azanza, R.V. Paralytic shellfish toxin uptake, assimilation, depuration, and transformation in the southeast Asian green-lipped mussel (Perna viridis). Toxins 2019, 11, 468. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.A.; Cmb, C.; Qi, L. Effects of toxic dinoflagellate Alexandrium catenella on sexual maturation and reproductive output in the Pacific oyster Crassostrea gigas. Aquat. Toxicol. 2021, 232, 105745. [Google Scholar]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramón, M.; Wikfors, G.H. Pathology and immune response of the blue mussel (Mytilus edulis L.) after an exposure to the harmful dinoflagellate Prorocentrum minimum. Harmful Algae 2008, 7, 630–638. [Google Scholar] [CrossRef]
- Liu, Y.; Kong, F.-Z.; Xun, X.-G.; Dai, L.; Geng, H.-X.; Hu, X.-L.; Yu, R.-C.; Bao, Z.-M.; Zhou, M.-J. Biokinetics and biotransformation of paralytic shellfish toxins in different tissues of Yesso scallops, Patinopecten yessoensis. Chemosphere 2020, 261, 128063. [Google Scholar] [CrossRef]
- Mjba, B.; Fm, A.; Rf, C.; Ap, C.; Ep, D.; Cv, B. Paralytic shellfish toxin profiles in mussel, cockle and razor shell under post-bloom natural conditions: Evidence of higher biotransformation in razor shells and cockles. Mar. Environ. Res. 2020, 154, 104839. [Google Scholar]
- Vale, P. Metabolites of saxitoxin analogues in bivalves contaminated by Gymnodinium catenatum. Toxicon 2010, 55, 162–165. [Google Scholar] [CrossRef]
- Ding, J.; Sun, C.; He, C.; Li, J.; Ju, P.; Li, F. Microplastics in four bivalve species and basis for using bivalves as bioindicators of microplastic pollution. Sci. Total Environ. 2021, 782, 146830. [Google Scholar] [CrossRef]
- Strehse, J.S.; Maser, E. Marine bivalves as bioindicators for environmental pollutants with focus on dumped munitions in the sea: A review. Mar. Environ. Res. 2020, 158, 105006. [Google Scholar] [CrossRef]
- Comeau, L.A.; Babarro, J.M.; Riobó, P.; Scarratt, M.; Starr, M.; Tremblay, R. PSP-producing dinoflagellate Alexandrium minutum induces valve microclosures in the mussel Mytilus galloprovincialis. Aquaculture 2019, 500, 407–413. [Google Scholar] [CrossRef]
- Braga, A.C.; Pereira, V.; Marçal, R.; Marques, A.; Guilherme, S.; Costa, P.R.; Pacheco, M. DNA damage and oxidative stress responses of mussels Mytilus galloprovincialis to paralytic shellfish toxins under warming and acidification conditions—Elucidation on the organ-specificity. Aquat. Toxicol. 2020, 228, 105619. [Google Scholar] [CrossRef]
- Prego-Faraldo, M.V.; Martínez, L.; Méndez, J. RNA-seq analysis for assessing the early response to DSP toxins in Mytilus galloprovincialis digestive gland and gill. Toxins 2018, 10, 417. [Google Scholar] [CrossRef] [Green Version]
- Pazos, A.J.; Ventoso, P.; Martínez-Escauriaza, R.; Pérez-Parallé, M.L.; Blanco, J.; Triviño, J.C.; Sánchez, J.L. Transcriptional response after exposure to domoic acid-producing Pseudo-nitzschia in the digestive gland of the mussel Mytilus galloprovincialis. Toxicon 2017, 140, 60–71. [Google Scholar]
- Suarez-Ulloa, V.; Fernandez-Tajes, J.; Aguiar-Pulido, V.; Prego-Faraldo, M.V.; Eirin-Lopez, J.M. Unbiased high-throughput characterization of mussel transcriptomic responses to sublethal concentrations of the biotoxin okadaic acid. PeerJ 2015, 3, e1429. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; De Moro, G.; Manfrin, C.; Milandri, A.; Riccardi, E.; Beran, A.; Venier, P.; Pallavicini, A. RNA sequencing and de novo assembly of the digestive gland transcriptome in Mytilus galloprovincialis fed with toxinogenic and non-toxic strains of Alexandrium minutum. BMC Res. Notes 2014, 7, 722. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.B. Metabolic Transformation and Physiological and Biochemical Response of Bivalves to Paralytic Shellfish Poisoning; Ocean University of China: Qingdao, China, 2014. [Google Scholar]
- Leverone, J.R.; Shumway, S.E.; Blake, N.J. Comparative effects of the toxic dinoflagellate Karenia brevis on clearance rates in juveniles of four bivalve molluscs from Florida, USA. Toxicon 2007, 49, 634–645. [Google Scholar] [CrossRef]
- LeveroneÃ, J.R.; Blake, N.J.; Pierce, R.H.; Shumway, S.E. Effects of the dinoflagellate Karenia brevis on larval development in three species of bivalve mollusc from Florida. Toxicon 2006, 48, 75–84. [Google Scholar] [CrossRef]
- Shan, W.; Rui, H.; Bao, Z.; Du, H.; He, Y.; Su, H.; Zhang, Y.; Fu, X.; Jiao, W.; Li, Y. Transcriptome sequencing of zhikong scallop (Chlamys farreri) and comparative transcriptomic analysis with yesso scallop (Patinopecten yessoensis). PLoS ONE 2013, 8, 63927. [Google Scholar]
- Lk, A.; Cbb, C.; Tb, C.; Vs, D.; Mhde, F.; Fm, B.; Hs, A. Comparative de novo assembly and annotation of mantle tissue transcriptomes from the Mytilus edulis species complex (M. edulis, M. galloprovincialis, M. trossulus). Mar. Genom. 2020, 51, 100700. [Google Scholar]
- Song, L.; Wang, L.; Zhang, H.; Wang, M. The immune system and its modulation mechanism in scallop. Fish Shellfish. Immunol. 2015, 46, 65–78. [Google Scholar] [CrossRef]
- Marco, G.; Venier, P. An updated molecular basis for mussel immunity. Fish Shellfish Immunol. 2015, 46, 17–38. [Google Scholar]
- Shk, A.; Kwn, A.; Ba, B.; Ksc, C.; Khp, A.; Kip, A. Quantification of the inflammatory responses to pro-and anti-Inflammatory agents in manila clam, Ruditapes philippinarum. Fish Shellfish Immunol. 2021, 115, 22–26. [Google Scholar]
- Ugo, M.; Sergiy, K.; Baldur, S. The role of mitogen-activated protein kinase-activated protein kinases (MAPKAPKs) in inflammation. Genes 2013, 4, 101–133. [Google Scholar]
- Wei, X.; Lu, M.; Duan, G.; Li, H.; Liu, J.; Yang, W. Responses of CYP450 in the mussel Perna viridis after short-term exposure to the DSP toxins-producing dinoflagellate Prorocentrum lima. Ecotoxicol. Environ. Saf. 2019, 176, 178–185. [Google Scholar] [CrossRef]
- Kang, H.A.; Yu, Z.A.; Ht, A.; Zz, A.; Hz, B.; Fh, A.; Fy, A. Macrophage immunomodulatory effects of low molecular weight peptides from Mytilus coruscus via NF-κB/MAPK signaling pathways. J. Funct. Foods 2021, 83, 104562. [Google Scholar]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chovatiya, R.; Medzhitov, R. Stress, inflammation, and defense of homeostasis. Mol. Cell 2014, 54, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yu, Y.; Guo, M.; Lin, Y.; Tan, Z. Integrated analysis of physiological, transcriptomics and metabolomics provides insights into detoxication disruption of PFOA exposure in Mytilus edulis. Ecotoxicol. Environ. Saf. 2021, 214, 112081. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Review article origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joy, M. High-value compounds from the molluscs of marine and estuarine ecosystems as prospective functional food ingredients: An overview. Food Res. Int. 2020, 137, 109637. [Google Scholar] [CrossRef]
- Theoharides, T.C. Stress, inflammation, and autoimmunity: The 3 modern erinyes. Clin. Ther. 2020, 42, 742–744. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, H.; Wang, Y.; Fan, C.; Shen, X. Combined effects of temperature and toxic algal abundance on paralytic shellfish toxic accumulation, tissue distribution and elimination dynamics in mussels Mytilus coruscus. Toxins 2021, 13, 425. [Google Scholar] [CrossRef]
- Guan, C.; Guo, H.; Jiao, X.; Zhao, W. Studies on the time of accumulation and elimination of Alexandrium tamarense toxins in Argopectens irradias. J. Earth. Sci.-China 2012, 2, 112–117. [Google Scholar]
- Bianchi, V.A.; Langeloh, H.; Tillmann, U.; Krock, B.; Müller, A.; Bickmeyer, U.; Abele, D. Separate and combined effects of neurotoxic and lytic compounds of Alexandrium strains on Mytilus edulis feeding activity and hemocyte function. Fish Shellfish Immunol. 2019, 84, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Zhou, M.; Fu, M.; Yu, R.; Wang, Y.; Li, J. Effects of the dinoflagellate Alexandrium tamarense on early development of the scallop Argopecten irradians concentricus. Aquaculture 2003, 217, 167–178. [Google Scholar] [CrossRef]
- Gerdol, M.; Moreira, R.; Cruz, F.; Gómez-Garrido, J.; Figueras, A. Massive gene presence-absence variation shapes an open pan-genome in the Mediterranean mussel. Genome Biol. 2020, 21, 275. [Google Scholar] [CrossRef]
- Figueras, A.; Moreira, R.; Sendra, M.; Novoa, B. Genomics and immunity of the mediterranean mussel Mytilus galloprovincialis in a changing environment. Fish Shellfish Immunol. 2019, 90, 440–445. [Google Scholar] [CrossRef]
- Cappello, T.; Maisano, M.; D’Agata, A.; Natalotto, A.; Mauceri, A.; Fasulo, S. Effects of environmental pollution in caged mussels (Mytilus galloprovincialis). Mar. Environ. Res. 2013, 91, 52–60. [Google Scholar] [CrossRef]
- Chi, C.; Giri, S.S.; Jun, J.W.; Yun, S.; Kim, H.J.; Kim, S.G.; Park, S.C. Immune response of the bay scallop, Argopecten irradians, after exposure to the algicide palmitoleic acid. Fish Shellfish Immunol. 2016, 57, 371–378. [Google Scholar] [CrossRef]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Yun, S.; Kim, S.G.; Park, S.C. Marine toxin okadaic acid affects the immune function of bay scallop (Argopecten irradians). Molecules 2016, 21, 1108. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.; Zhang, C.; Liu, J.; Zheng, X. Effects of marine toxin domoic acid on innate immune responses in bay scallop Argopecten irradians. J. Mar. Sci. Eng. 2019, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- Hermabessiere, L.; Fabioux, C.; Lassudrie, M.; Boullot, F.; Long, M.; Lambert, C.; Le Goïc, N.; Gouriou, J.; Le Gac, M.; Chapelle, A.; et al. Influence of gametogenesis pattern and sex on paralytic shellfish toxin levels in triploid Pacific oyster Crassostrea gigas exposed to a natural bloom of Alexandrium minutum. Aquaculture 2016, 455, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Yun, S.; Park, S.C. Effects of algal toxin okadaic acid on the non-specific immune and antioxidant response of bay scallo(Argopecten irradians. Fish Shellfish Immunol. 2017, 65, 111–117. [Google Scholar] [CrossRef]
- Qiu, J.B.; Ma, F.; Fan, H.; Li, A. Effects of feeding Alexandrium tamarense, a paralytic shellfish toxin producer, on antioxidant enzymes in scallops (Patinopecten yessoensis) and mussels (Mytilus galloprovincialis). Aquaculture 2013, 396–399, 76–81. [Google Scholar] [CrossRef]
- Nagahama, M.; Shibutani, M.; Seike, S.; Yonezaki, M.; Takagishi, T.; Oda, M.; Kobayashi, K.; Sakurai, J. The p38 MAPK and JNK pathways protect host cells against clostridium perfringens beta-toxin. Infect. Immun. 2013, 81, 3703–3708. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Park, S.C. Detoxification and immune transcriptomic response of the gill tissue of bay scallop (Argopecten irradians) following exposure to the algicide palmitoleic acid. Biomolecules 2018, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.; Yun, S.; Giri, S.S.; Kim, H.J.; Kang, J.W.; Park, S.C. Effect of the algicide thiazolidinedione 49 on immune responses of bay scallop Argopecten Irradians. Molecules 2019, 24, 3579. [Google Scholar] [CrossRef] [Green Version]
- Song, J.A.; Choi, C.Y.; Park, H. Exposure of bay scallop Argopecten irradians to micro-polystyrene: Bioaccumulation and toxicity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 236, 108801. [Google Scholar] [CrossRef]
- Huang, B.; Lin, C.; Chen, H.; Lin, J.; Cheng, Y.; Kao, S. AMPK activation inhibits expression of proinflammatory mediators through downregulation of PI3K/p38 MAPK and NF-κB signaling in murine macrophages. DNA Cell Biol. 2015, 34, 133–141. [Google Scholar] [CrossRef]
- Bouallegui, Y. Immunity in mussels: An overview of molecular components and mechanisms with a focus on the functional defenses. Fish Shellfish Immunol. 2019, 89, 158–169. [Google Scholar] [CrossRef]
- Romero, A.; Novoa, B.; Figueras, A. The complexity of apoptotic cell death in mollusks: An update. Fish Shellfish Immunol. 2015, 46, 79–87. [Google Scholar] [CrossRef]
- Hardie, D.G.; Schaffer, B.E.; Brunet, A. AMPK: An energy-sensing pathway with multiple inputs and outputs. Trends Cell Biol. 2015, 26, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, C.; Speirs, C.; Williams, J.J.L.; Ewart, M.-A.; Mancini, S.J.; Hawley, S.A.; Delles, C.; Viollet, B.; Costa-Pereira, A.P.; Baillie, G.S.; et al. Phosphorylation of Janus kinase 1 (JAK1) by AMP-activated protein kinase (AMPK) links energy sensing to anti-inflammatory signaling. Sci. Signal. 2016, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Qiu, J.; Wang, X.; Zhang, Y.; Xia, M. AMP-activated protein kinase suppresses endothelial cell inflammation through phosphorylation of transcriptional coactivator. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2897–2908. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Wu, H.; Guo, M.; Bing. X.; Zheng. G.; Peng. J.; Tan. Z.; Zhai. Y. Simultaneous identification and detection of paralytic shellfish toxin in bivalve mollusks by liquid chromatography coupled with quadrupole/linear ion trap tandem mass apectrometry. Oceanol. Limnol. Sin. 2017, 48, 623–632. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M. galloprovincialis | A. irridians | |||||||
|---|---|---|---|---|---|---|---|---|
| Fast-Stage | Slow-Stage | Fast-Stage | Slow-Stage | |||||
| NES | p-Value | NES | p-Value | NES | p-Value | NES | p-Value | |
| MAPK | 1.13 | - | 1.82 | *** | 1.06 | - | −1.31 | * |
| TNF | 0.88 | - | 2.01 | *** | 1.24 | * | −1.28 | * |
| Ras | 1.04 | - | 1.50 | *** | 1.01 | - | −0.87 | - |
| NF-κB | 0.82 | - | 1.74 | *** | 1.38 | * | 0.85 | - |
| AMPK | −1.55 | *** | 1.85 | *** | −1.27 | - | −1.15 | - |
| PI3K-Akt | - | - | - | - | 1.12 | - | 1.11 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Wu, H.; Zheng, G.; Peng, J.; Guo, M.; Tan, Z. Transcriptome Analysis Reveals MAPK/AMPK as a Key Regulator of the Inflammatory Response in PST Detoxification in Mytilus galloprovincialis and Argopecten irradians. Toxins 2022, 14, 516. https://doi.org/10.3390/toxins14080516
Dong C, Wu H, Zheng G, Peng J, Guo M, Tan Z. Transcriptome Analysis Reveals MAPK/AMPK as a Key Regulator of the Inflammatory Response in PST Detoxification in Mytilus galloprovincialis and Argopecten irradians. Toxins. 2022; 14(8):516. https://doi.org/10.3390/toxins14080516
Chicago/Turabian StyleDong, Chenfan, Haiyan Wu, Guanchao Zheng, Jixing Peng, Mengmeng Guo, and Zhijun Tan. 2022. "Transcriptome Analysis Reveals MAPK/AMPK as a Key Regulator of the Inflammatory Response in PST Detoxification in Mytilus galloprovincialis and Argopecten irradians" Toxins 14, no. 8: 516. https://doi.org/10.3390/toxins14080516
APA StyleDong, C., Wu, H., Zheng, G., Peng, J., Guo, M., & Tan, Z. (2022). Transcriptome Analysis Reveals MAPK/AMPK as a Key Regulator of the Inflammatory Response in PST Detoxification in Mytilus galloprovincialis and Argopecten irradians. Toxins, 14(8), 516. https://doi.org/10.3390/toxins14080516