Mycotoxins of Concern in Children and Infant Cereal Food at European Level: Incidence and Bioaccessibility


Abstract
:1. Introduction
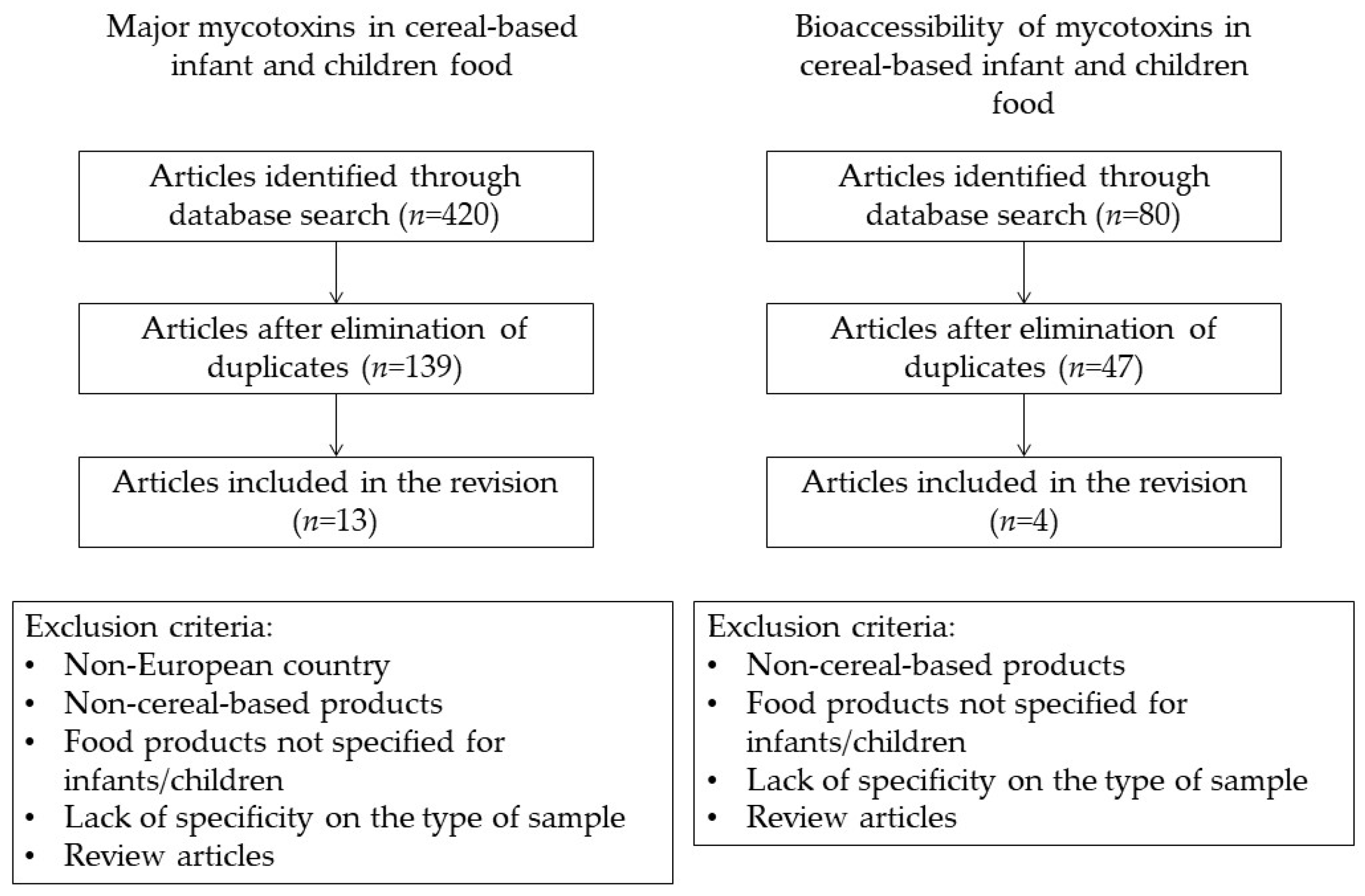
2. Materials and Methods
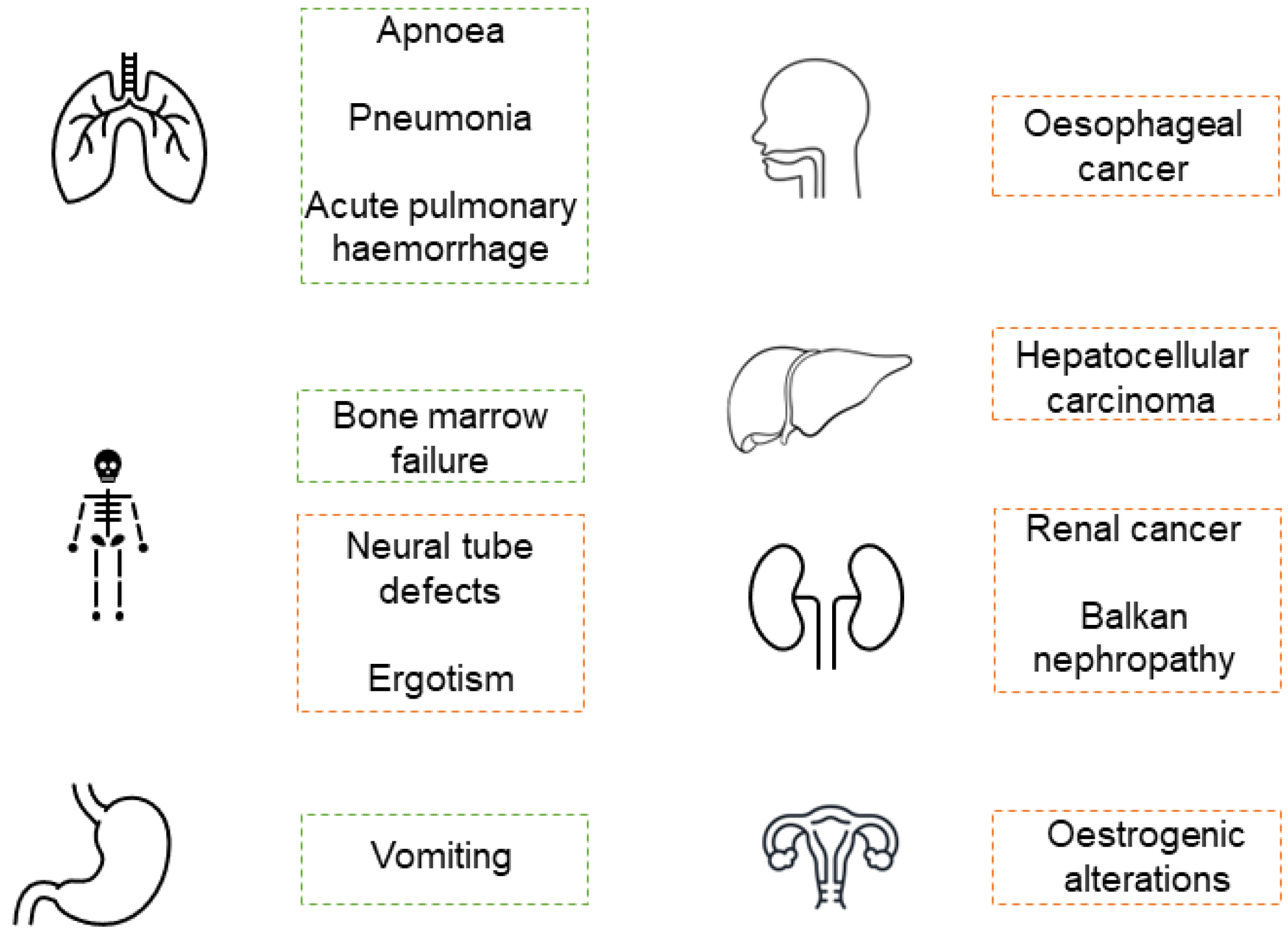
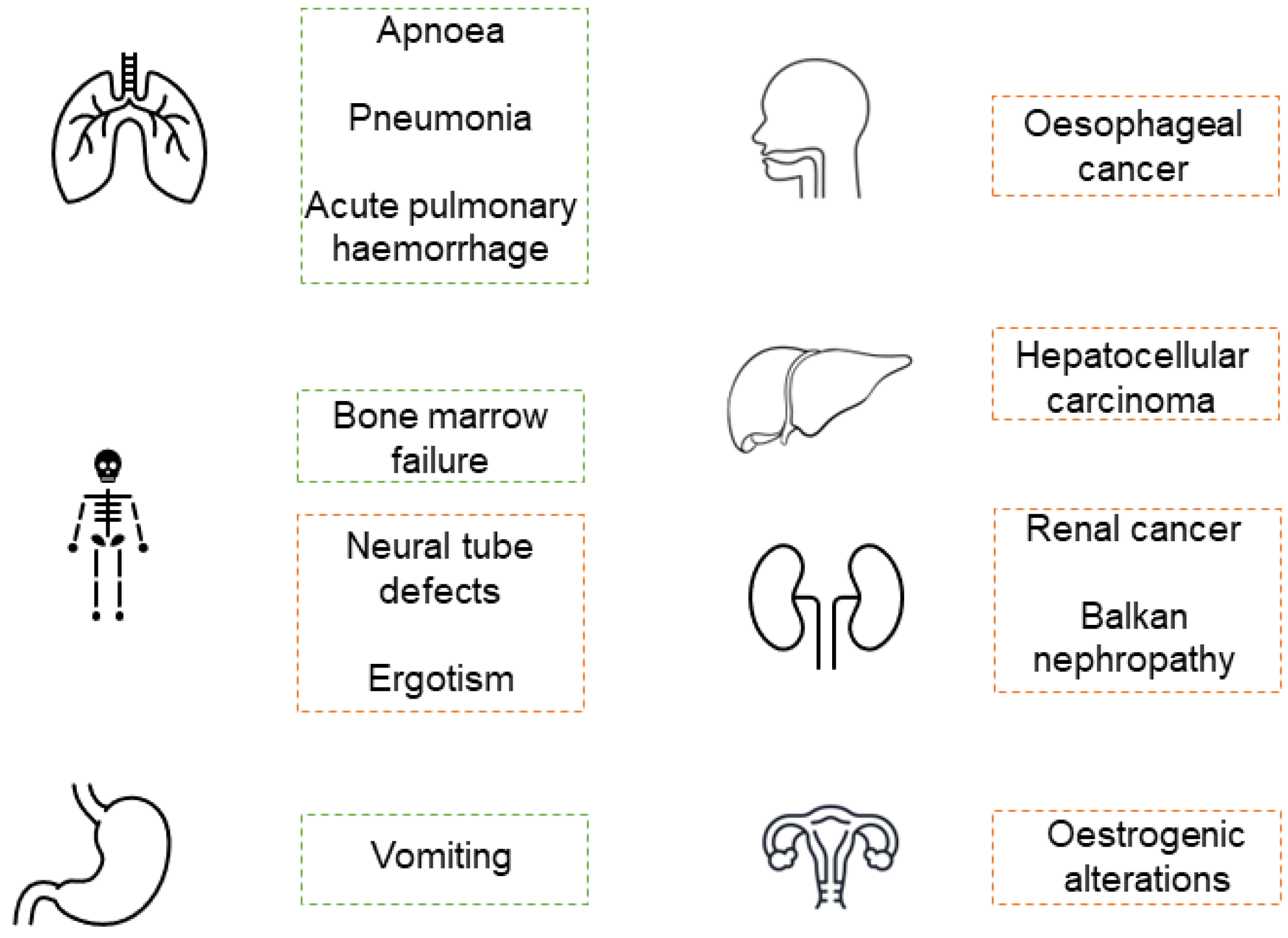
3. Mycotoxicosis in Infants and Children
4. Protective Effects of Cereals against Mycotoxin Exposure
5. Major Mycotoxins in Cereals
6. Bioaccessibility of Mycotoxins in Cereal-Based Infants’ and Children’s Food and Strategies for Its Reduction
7. Methods for Mycotoxin Analysis in Food and Bioaccessibility Studies
7.1. Analysis of Mycotoxin in Food
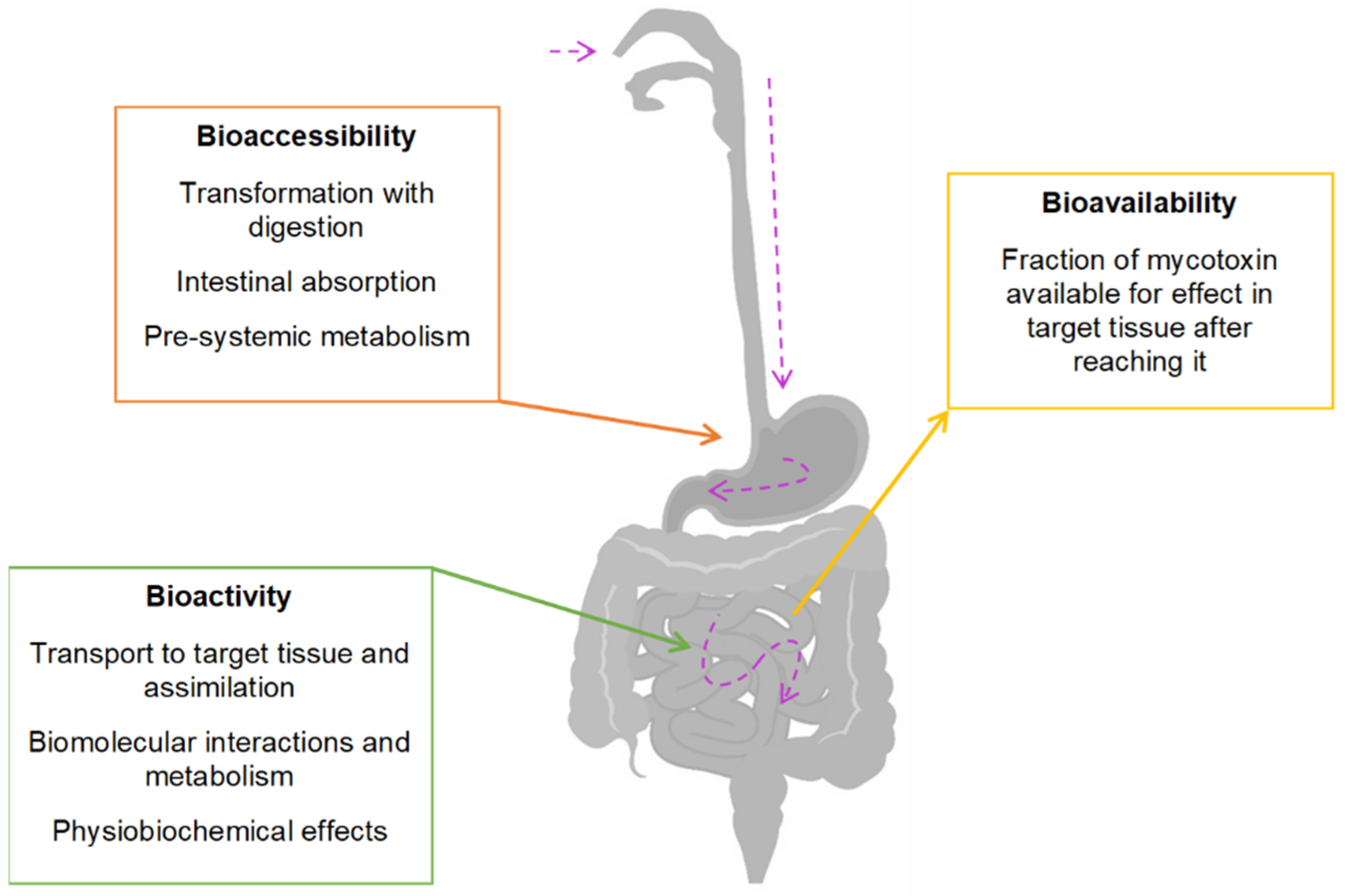
7.2. Bioavailability and Bioaccessibility Analysis
8. Gaps in the Research of Mycotoxins in Infant/Children Cereal-Based Food Matrices
9. Final Considerations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cascant, M.M.; Garrigues, S.; de la Guardia, M. Direct determination of major components in human diets and baby foods. Anal. Bioanal. Chem. 2015, 407, 1961–1972. [Google Scholar] [CrossRef] [PubMed]
- Amezdroz, E.; Carpenter, L.; O’Callaghan, E.; Johnson, S.; Waters, E. Transition from milks to the introduction of solid foods across the first 2 years of life: Findings from an Australian birth cohort study. J. Hum. Nutr. Diet. 2015, 28, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, M.E.; Nulty, A.K. When Does It All Begin: What, When, and How Young Children Are Fed. Nestle Nutr. Inst. Workshop Ser. 2020, 93, 15–24. [Google Scholar] [PubMed]
- UNICEF. From the First Hour of Life; Data and Analytics, Division of Data, Research and Policy and Nutrition Section, Programe Division—UNICEF: New York, USA, 2016; ISBN 978-92-806-4852-2. [Google Scholar]
- Klerks, M.; Bernal, M.J.; Roman, S.; Bodenstab, S.; Gil, A.; Sanchez-Siles, L.M. Infant Cereals: Current Status, Challenges, and Future Opportunities for Whole Grains. Nutrients 2019, 11, 473. [Google Scholar] [CrossRef] [Green Version]
- Doughari, J.H. The Occurrence, Properties and Significance of Citrinin Mycotoxin. J. Plant Pathol. Microbiol. 2015, 6, 11. [Google Scholar]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef]
- Peraica, M.; Richter, D.; Rašić, D. Mycotoxicoses in children. Arch. Ind. Hyg. Toxicol. 2014, 65, 347–363. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Gelderblom, W.C.; Marasas, W.F. Controversies in fumonisin mycotoxicology and risk assessment. Hum. Exp. Toxicol. 2012, 31, 215–235. [Google Scholar] [CrossRef]
- Alizadeh, A.M.; Rohandel, G.; Roudbarmohammadi, S.; Roudbary, M.; Sohanaki, H.; Ghiasian, S.A.; Taherkhani, A.; Semnani, S.; Aghasi, M. Fumonisin B1 contamination of cereals and risk of esophageal cancer in a high risk area in northeastern Iran. Asian Pac. J. Cancer Prev. 2012, 13, 2625–2628. [Google Scholar] [CrossRef] [Green Version]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. ‘Emerging’ mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef]
- Sherif, S.O.; Salama, E.E.; Abdel-Wahhab, M.A. Mycotoxins and child health: The need for health risk assessment. Int. J. Hyg. Environ. Health 2009, 212, 347–368. [Google Scholar] [CrossRef]
- Raiola, A.; Tenore, G.C.; Manyes, L.; Meca, G.; Ritieni, A. Risk analysis of main mycotoxins occurring in food for children: An overview. Food Chem. Toxicol. 2015, 84, 169–180. [Google Scholar] [CrossRef]
- Hulin, M.; Bemrah, N.; Nougadère, A.; Volatier, J.L.; Sirot, V.; Leblanc, J.C. Assessment of infant exposure to food chemicals: The French Total Diet Study design. Food Addit. Contam. Part A 2014, 31, 1226–1239. [Google Scholar] [CrossRef]
- Arce-López, B.; Lizarraga, E.; López de Mesa, R.; González-Peñas, E. Assessment of Exposure to Mycotoxins in Spanish Children through the Analysis of Their Levels in Plasma Samples. Toxins 2021, 13, 150. [Google Scholar] [CrossRef]
- Kushnir-Sukhov, N.M. A Novel Link between Early Life Allergen Exposure and Neuroimmune Development in Children. J. Clin. Exp. Immunol. 2020, 5, 188–195. [Google Scholar]
- Etzel, R.A. What the primary care pediatrician should know about syndromes associated with exposures to mycotoxins. Curr. Probl. Pediatr. Adolesc. Health Care 2006, 36, 282–305. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Ryu, D. Advances in Mycotoxin Research: Public Health Perspectives. J. Food Sci. 2015, 80, T2970–T2983. [Google Scholar] [CrossRef]
- Guerre, P. Mycotoxin and Gut Microbiota Interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef]
- Schöneberg, T.; Kibler, K.; Sulyok, M.; Musa, T.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Voegele, R.T.; Vogelgsang, S. Can plant phenolic compounds reduce Fusarium growth and mycotoxin production in cereals? Food Addit. Contam. Part A 2018, 35, 2455–2470. [Google Scholar] [CrossRef] [Green Version]
- Stuper-Szablewska, K.; Kurasiak-Popowska, D.; Nawracała, J.; Perkowski, J. Quantitative profile of phenolic acids and antioxidant activity of wheat grain exposed to stress. Eur. Food Res. Technol. 2019, 245, 1595–1603. [Google Scholar] [CrossRef] [Green Version]
- Ferruz, E.; Loran, S.; Herrera, M.; Gimenez, I.; Bervis, N.; Barcena, C.; Carramiñana, J.J.; Juan, T.; Herrera, A.; Ariño, A. Inhibition of Fusarium Growth and Mycotoxin Production in Culture Medium and in Maize Kernels by Natural Phenolic Acids. J. Food Prot. 2016, 79, 1753–1758. [Google Scholar] [CrossRef]
- Atanasova-Penichon, V.; Barreau, C.; Richard-Forget, F. Antioxidant Secondary Metabolites in Cereals: Potential Involvement in Resistance to Fusarium and Mycotoxin Accumulation. Front. Microbiol. 2016, 7, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fardet, A.; Rock, E.; Rémésy, C. Is the in vitro antioxidant potential of whole-grain cereals and cereal products well reflected in vivo? J. Cereal Sci. 2008, 48, 258–276. [Google Scholar] [CrossRef]
- Slavin, J. Why whole grains are protective: Biological mechanisms. Proc. Nutr. Soc. 2003, 62, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Eaton, J.W. Antioxidant functions of phytic acid. Free Radic. Biol. Med. 1990, 8, 61–69. [Google Scholar] [CrossRef]
- Feizollahi, E.; Mirmahdi, R.S.; Zoghi, A.; Zijlstra, R.T.; Roopesh, M.S.; Vasanthan, T. Review of the beneficial and anti-nutritional qualities of phytic acid, and procedures for removing it from food products. Food Res. Int. 2021, 143, 110284. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, R.; Yang, Y.; Xie, H.; Huang, Y.; Chen, X.; Wang, D.; Zhang, Z. Protective Effect of Organic Selenium on Oxidative Damage and Inflammatory Reaction of Rabbit Kidney Induced by T-2 Toxin. Biol. Trace Elem. Res. 2021, 199, 1833–1842. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [Green Version]
- Slavin, J.; Jacobs, D.; Marquart, L. Whole-grain consumption and chronic disease: Protective mechanisms. Nutr. Cancer 1997, 27, 14–21. [Google Scholar] [CrossRef]
- Aoudia, N.; Callu, P.; Grosjean, F.; Larondelle, Y. Effectiveness of mycotoxin sequestration activity of micronized wheat fibres on distribution of ochratoxin A in plasma, liver and kidney of piglets fed a naturally contaminated diet. Food Chem. Toxicol. 2009, 47, 1485–1489. [Google Scholar] [CrossRef]
- Fardet, A. 16—New Concepts and Paradigms for the Protective Effects of Plant-Based Food Components in Relation to Food Complexity. In Vegetarian and Plant-Based Diets in Health and Disease Prevention; Mariotti, F., Ed.; Academic Press: London, UK, 2017; pp. 293–312. [Google Scholar]
- Hernández, M.; Juan-García, A.; Moltó, J.C.; Mañes, J.; Juan, C. Evaluation of Mycotoxins in Infant Breast Milk and Infant Food, Reviewing the Literature Data. Toxins 2021, 13, 535. [Google Scholar] [CrossRef]
- Coppa, C.F.S.C.; Khaneghah, A.M.; Alvito, P.; Assunção, R.; Martins, C.; Eş, I.; Gonçalves, B.L.; Valganon de Neeff, D.; Sant’Ana, A.S.; Corassin, C.H.; et al. The occurrence of mycotoxins in breast milk, fruit products and cereal-based infant formula: A review. Trends Food Sci. Technol. 2019, 92, 81–93. [Google Scholar] [CrossRef]
- De Sá, S.V.M.; Monteiro, C.; Fernandes, J.O.; Pinto, E.; Faria, M.A.; Cunha, S.C. Emerging mycotoxins in infant and children foods: A review. Crit. Rev. Food Sci. Nutr. 2021, 1–15. [Google Scholar] [CrossRef]
- Juan, C.; Raiola, A.; Mañes, J.; Ritieni, A. Presence of mycotoxin in commercial infant formulas and baby foods from Italian market. Food Control 2014, 39, 227–236. [Google Scholar] [CrossRef]
- Pereira, V.L.; Fernandes, J.O.; Cunha, S.C. Comparative assessment of three cleanup procedures after QuEChERS extraction for determination of trichothecenes (type A and type B) in processed cereal-based baby foods by GC–MS. Food Chem. 2015, 182, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Assunção, R.; Vasco, E.; Nunes, B.; Loureiro, S.; Martins, C.; Alvito, P. Single-compound and cumulative risk assessment of mycotoxins present in breakfast cereals consumed by children from Lisbon region, Portugal. Food Chem. Toxicol. 2015, 86, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Hampikyan, H.; Bingol, E.B.; Colak, H.; Cetin, O.; Bingol, B. Determination of ochratoxin a in baby foods by ELISA and HPLC. Acta Aliment. 2015, 44, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Assunção, R.; Martins, C.; Dupont, D.; Alvito, P. Patulin and ochratoxin A co-occurrence and their bioaccessibility in processed cereal-based foods: A contribution for Portuguese children risk assessment. Food Chem. Toxicol. 2016, 96, 205–214. [Google Scholar] [CrossRef]
- Martins, C.; Assunção, R.; Cunha, S.C.; Fernandes, J.O.; Jager, A.; Petta, T.; Oliveira, C.A.; Alvito, P. Assessment of multiple mycotoxins in breakfast cereals available in the Portuguese market. Food Chem. 2018, 239, 132–140. [Google Scholar] [CrossRef]
- Assunção, R.; Martins, C.; Vasco, E.; Jager, A.; Oliveira, C.; Cunha, S.C.; José, O.F.; Nunes, B.; Loureiro, S.; Alvito, P. Portuguese children dietary exposure to multiple mycotoxins—An overview of risk assessment under MYCOMIX project. Food Chem. Toxicol. 2018, 118, 399–408. [Google Scholar] [CrossRef]
- Gotthardt, M.; Asam, S.; Gunkel, K.; Moghaddam, A.F.; Baumann, E.; Kietz, R.; Rychlik, M. Quantitation of Six Alternaria Toxins in Infant Foods Applying Stable Isotope Labeled Standards. Front. Microbiol. 2019, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Bervis, N.; Carraminana, J.J.; Juan, T.; Herrera, A.; Arino, A.; Loran, S. Occurrence and Exposure Assessment of Aflatoxins and Deoxynivalenol in Cereal-Based Baby Foods for Infants. Toxins 2019, 11, 150. [Google Scholar] [CrossRef] [Green Version]
- Postupolski, J.; Starski, A.; Ledzion, E.; Kurpinska-Jaworska, J.; Szczesna, M. Exposure assessment of infants and young children on selected Fusarium toxins. Rocz. Panstw. Zakl. Hig. 2019, 70, 5–14. [Google Scholar] [CrossRef]
- Capei, R.; Pettini, L.; Mando Tacconi, F. Occurrence of Ochratoxin A in breakfast cereals and sweet snacks in Italy: Dietary exposure assessment. Ann Ig 2019, 31, 130–139. [Google Scholar] [PubMed]
- Braun, D.; Eiser, M.; Puntscher, H.; Marko, D.; Warth, B. Natural contaminants in infant food: The case of regulated and emerging mycotoxins. Food Control 2021, 123, 107676. [Google Scholar] [CrossRef]
- Mruczyk, K.; Cisek-Woźniak, A.; Mizgier, M.; Wójciak, R.W. Natural Occurrence of Deoxynivalenol in Cereal-Based Baby Foods for Infants from Western Poland. Toxins 2021, 13, 777. [Google Scholar] [CrossRef] [PubMed]
- González-Arias, C.A.; Marín, S.; Sanchis, V.; Ramos, A.J. Mycotoxin bioaccessibility/absorption assessment using in vitro digestion models: A review. World Mycotoxin J. 2013, 6, 167–184. [Google Scholar] [CrossRef] [Green Version]
- Arce-López, B.; Lizarraga, E.; Vettorazzi, A.; González-Peñas, E. Human Biomonitoring of Mycotoxins in Blood, Plasma and Serum in Recent Years: A Review. Toxins 2020, 12, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, C.; Afonso, C.; Lourenço, H.; Costa, S.; Nunes, M.L. Bioaccessibility assessment methodologies and their consequences for the risk–benefit evaluation of food. Trends Food Sci. Technol. 2015, 41, 5–23. [Google Scholar] [CrossRef]
- Rebellato, A.P.; dos Santos Caramês, E.T.; Pallone, J.A.L.; de Oliveira Rocha, L. Mycotoxin bioaccessibility in baby food through in vitro digestion: An overview focusing on risk assessment. Curr. Opin. Food Sci. 2021, 41, 107–115. [Google Scholar] [CrossRef]
- Raiola, A.; Meca, G.; Mañes, J.; Ritieni, A. Bioaccessibility of Deoxynivalenol and its natural co-occurrence with Ochratoxin A and Aflatoxin B1 in Italian commercial pasta. Food Chem. Toxicol. 2012, 50, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Kabak, B.; Brandon, E.F.A.; Var, I.; Blokland, M.; Sips, A.J.A.M. Effects of probiotic bacteria on the bioaccessibility of aflatoxin B1 and ochratoxin A using an in vitro digestion model under fed conditions. J. Environ. Sci. Health Part B 2009, 44, 472–480. [Google Scholar] [CrossRef]
- Prosperini, A.; Meca, G.; Font, G.; Ruiz, M.-J. Bioaccessibility of Enniatins A, A1, B, and B1 in Different Commercial Breakfast Cereals, Cookies, and Breads of Spain. J. Agric. Food Chem. 2013, 61, 456–461. [Google Scholar] [CrossRef]
- Tran, V.N.; Viktorová, J.; Ruml, T. Mycotoxins: Biotransformation and Bioavailability Assessment Using Caco-2 Cell Monolayer. Toxins 2020, 12, 628. [Google Scholar] [CrossRef]
- Assunção, R.; Ferreira, M.; Martins, C.; Diaz, I.; Padilla, B.; Dupont, D.; Bragança, M.; Alvito, P. Applicability of In Vitro Methods to Study Patulin Bioaccessibility and Its Effects on Intestinal Membrane Integrity. J. Toxicol. Environ. Health Part A 2014, 77, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusariummycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Horn, N.; Ajuwon, K.M. Mechanisms of deoxynivalenol-induced endocytosis and degradation of tight junction proteins in jejunal IPEC-J2 cells involve selective activation of the MAPK pathways. Arch. Toxicol. 2021, 95, 2065–2079. [Google Scholar] [CrossRef]
- Wen, J.; Mu, P.; Deng, Y. Mycotoxins: Cytotoxicity and biotransformation in animal cells. Toxicol. Res. 2016, 5, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Gross-Steinmeyer, K.; Eaton, D.L. Dietary modulation of the biotransformation and genotoxicity of aflatoxin B1. Toxicology 2012, 299, 69–79. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, L.; Zhang, N.-Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.-S.; Sun, L.-H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res./Rev. Mutat. Res. 2018, 778, 79–89. [Google Scholar] [CrossRef]
- Gratz, S.; Wu, Q.K.; El-Nezami, H.; Juvonen, R.O.; Mykkänen, H.; Turner, P.C. Lactobacillus rhamnosus strain GG reduces aflatoxin B1 transport, metabolism, and toxicity in Caco-2 Cells. Appl. Environ. Microbiol. 2007, 73, 3958–3964. [Google Scholar] [CrossRef] [Green Version]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.-R.; Anadón, A.; Martínez, M.-A. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology 2016, 353–354, 21–33. [Google Scholar] [CrossRef]
- Gao, Y.; Bao, X.; Meng, L.; Liu, H.; Wang, J.; Zheng, N. Aflatoxin B1 and Aflatoxin M1 Induce Compromised Intestinal Integrity through Clathrin-Mediated Endocytosis. Toxins 2021, 13, 184. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures. Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins 2019, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- Kőszegi, T.; Poór, M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins 2016, 8, 111. [Google Scholar] [CrossRef]
- Ringot, D.; Chango, A.; Schneider, Y.-J.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem.-Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Alizadeh, A.; Akbari, P.; Varasteh, S.; Braber, S.; Malekinejad, H.; Fink-Gremmels, J. Ochratoxin A challenges the intestinal epithelial cell integrity: Results obtained in model experiments with Caco-2 cells. World Mycotoxin J. 2019, 12, 399–407. [Google Scholar] [CrossRef]
- Wang, H.; Zhai, N.; Chen, Y.; Fu, C.; Huang, K. OTA induces intestinal epithelial barrier dysfunction and tight junction disruption in IPEC-J2 cells through ROS/Ca2+-mediated MLCK activation. Environ. Pollut. 2018, 242, 106–112. [Google Scholar] [CrossRef]
- Maresca, M. From the Gut to the Brain: Journey and Pathophysiological Effects of the Food-Associated Trichothecene Mycotoxin Deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef]
- Wu, Q.-H.; Wang, X.; Yang, W.; Nüssler, A.K.; Xiong, L.-Y.; Kuča, K.; Dohnal, V.; Zhang, X.-J.; Yuan, Z.-H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Sergent, T.; Parys, M.; Garsou, S.; Pussemier, L.; Schneider, Y.-J.; Larondelle, Y. Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular metabolism at realistic intestinal concentrations. Toxicol. Lett. 2006, 164, 167–176. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Gremmels, H.; Koelink, P.J.; Verheijden, K.A.T.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414–2429. [Google Scholar] [CrossRef] [Green Version]
- Kadota, T.; Furusawa, H.; Hirano, S.; Tajima, O.; Kamata, Y.; Sugita-Konishi, Y. Comparative study of deoxynivalenol, 3-acetyldeoxynivalenol, and 15-acetyldeoxynivalenol on intestinal transport and IL-8 secretion in the human cell line Caco-2. Toxicol. In Vitro 2013, 27, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Terciolo, C.; Neves, M.; Puel, S.; Naylies, C.; Lippi, Y.; Pinton, P.; Oswald, I.P. Comparative sensitivity of proliferative and differentiated intestinal epithelial cells to the food contaminant, deoxynivalenol. Environ. Pollut. 2021, 277, 116818. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.-K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018, 16, e05242. [Google Scholar]
- Malekinejad, H.; Maas-Bakker, R.F.; Fink-Gremmels, J. Bioactivation of zearalenone by porcine hepatic biotransformation. Vet. Res. 2005, 36, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. 2006, 172, 96–102. [Google Scholar] [CrossRef]
- Videmann, B.; Mazallon, M.; Tep, J.; Lecoeur, S. Metabolism and transfer of the mycotoxin zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2008, 46, 3279–3286. [Google Scholar] [CrossRef]
- Sforza, S.; Dall’asta, C.; Marchelli, R. Recent advances in mycotoxin determination in food and feed by hyphenated chromatographic techniques/mass spectrometry. Mass Spectrom. Rev. 2006, 25, 54–76. [Google Scholar] [CrossRef]
- Meca, G.; Ritieni, A.; Mañes, J. Influence of the heat treatment on the degradation of the minor Fusarium mycotoxin beauvericin. Food Control 2012, 28, 13–18. [Google Scholar] [CrossRef]
- Luz, C.; Saladino, F.; Luciano, F.B.; Mañes, J.; Meca, G. Occurrence, toxicity, bioaccessibility and mitigation strategies of beauvericin, a minor Fusarium mycotoxin. Food Chem. Toxicol. 2017, 107, 430–439. [Google Scholar] [CrossRef]
- Saladino, F.; Posarelli, E.; Luz, C.; Luciano, F.B.; Rodriguez-Estrada, M.T.; Mañes, J.; Meca, G. Influence of probiotic microorganisms on aflatoxins B1 and B2 bioaccessibility evaluated with a simulated gastrointestinal digestion. J. Food Compos. Anal. 2018, 68, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Luciano, F.B.; Meca, G.; Manyes, L.; Mañes, J. A chemical approach for the reduction of beauvericin in a solution model and in food systems. Food Chem. Toxicol. 2014, 64, 270–274. [Google Scholar] [CrossRef]
- Zheng, M.Z.; Richard, J.L.; Binder, J. A Review of Rapid Methods for the Analysis of Mycotoxins. Mycopathologia 2006, 161, 261–273. [Google Scholar] [CrossRef]
- Pereira, V.L.; Fernandes, J.O.; Cunha, S.C. Mycotoxins in cereals and related foodstuffs: A review on occurrence and recent methods of analysis. Trends Food Sci. Technol. 2014, 36, 96–136. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Versantvoort, C.H.M.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.M.; Sips, A.J.A.M. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Zafrilla, P.; Tomás-Barberán, F.A. An in vitro method to simulate phenolic compound release from the food matrix in the gastrointestinal tract. Eur. Food Res. Technol. 2002, 214, 155–159. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, A.; Jinap, S.; Soleimany, F. Qualitative and Quantitative Analysis of Mycotoxins. Compr. Rev. Food Sci. Food Saf. 2009, 8, 202–251. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Romano, A.; Masi, P.; Cavella, S. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018, 98, 4003–4013. [Google Scholar] [CrossRef]
- Xie, L.; Chen, M.; Ying, Y. Development of Methods for Determination of Aflatoxins. Crit. Rev. Food Sci. Nutr. 2016, 56, 2642–2664. [Google Scholar] [CrossRef]
- Turner, N.W.; Bramhmbhatt, H.; Szabo-Vezse, M.; Poma, A.; Coker, R.; Piletsky, S.A. Analytical methods for determination of mycotoxins: An update (2009–2014). Anal. Chim. Acta 2015, 901, 12–33. [Google Scholar] [CrossRef]
- Bueno, D.; Istamboulie, G.; Muñoz, R.; Marty, J. Determination of Mycotoxins in Food: A Review of Bioanalytical to Analytical Methods. Appl. Spectrosc. Rev. 2015, 50, 728–774. [Google Scholar] [CrossRef]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical methods for determination of mycotoxins: A review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef]
- Krska, R.; Schubert-Ullrich, P.; Molinelli, A.; Sulyok, M.; MacDonald, S.; Crews, C. Mycotoxin analysis: An update. Food Addit. Contam. Part A 2008, 25, 152–163. [Google Scholar] [CrossRef]
- Wilson, T.J.; Romer, T.R. Use of the Mycosep Multifunctional Cleanup Column for Liquid Chromatographic Determination of Aflatoxins in Agricultural Products. J. Assoc. Off. Anal. Chem. 1991, 74, 951–956. [Google Scholar] [CrossRef]
- Cunha, S.C.; Fernandes, J.O. Development and validation of a method based on a QuEChERS procedure and heart-cutting GC-MS for determination of five mycotoxins in cereal products. J. Sep. Sci. 2010, 33, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Cunha, S.C.; Fernandes, J.O. Chapter 21—Application in Food Analysis. In Liquid-Phase Extraction; Poole, C.F., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 643–665. [Google Scholar]
- Tamura, M.; Uyama, A.; Mochizuki, N. Development of a Multi-mycotoxin Analysis in Beer-based Drinks by a Modified QuEChERS Method and Ultra-High-Performance Liquid Chromatography Coupled with Tandem Mass Spectrometry. Anal. Sci. 2011, 27, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shephard, G.S. Current Status of Mycotoxin Analysis: A Critical Review. J. AOAC Int. 2016, 99, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Boyer, R.F. Modern Expeimental Biochemestry, 3rd ed.; Benjamin Cummings: San Francisco, CA, USA, 2000. [Google Scholar]
- Bessaire, T.; Mujahid, C.; Mottier, P.; Desmarchelier, A. Multiple Mycotoxins Determination in Food by LC-MS/MS: An International Collaborative Study. Toxins 2019, 11, 658. [Google Scholar] [CrossRef] [Green Version]
- Korfmacher, W.A. Foundation review: Principles and applications of LC-MS in new drug discovery. Drug Discov. Today 2005, 10, 1357–1367. [Google Scholar] [CrossRef]
- Wang, X.; Wang, S.; Cai, Z. The latest developments and applications of mass spectrometry in food-safety and quality analysis. TrAC Trends Anal. Chem. 2013, 52, 170–185. [Google Scholar] [CrossRef]
- Santos Pereira, C.; Cunha, S.C.; Fernandes, J.O. Prevalent Mycotoxins in Animal Feed: Occurrence and Analytical Methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Mycotoxins | Processed Cereal-Based Foods and Baby Foods for Infants and Young Children (µg/kg) |
|---|---|
| Aflatoxin B1 | 0.1 |
| Ochratoxin A | 0.5 |
| Patulin * | 10 |
| Deoxynivalenol | 200 |
| Sum T-2 and HT-2 toxin | 15 |
| Zearalenone | 20 |
| Sum Fumonisin B1 and Fumonisin B2 | 200 |
| Country | Sample | Mycotoxin | Total Samples | Positive Samples | References | ||
|---|---|---|---|---|---|---|---|
| % | Mean (µg/kg) | Range (µg/kg) | |||||
| Italy | Infant formulas and baby food | OTA | 75 | 20 | 0.06 | 0.050–0.120 | Juan et al., 2014 [42] |
| DON | 25.3 | 102.60 | 1–268 | ||||
| NIV | 4 | 19.91 | 5.5–235 | ||||
| FUS-X | 24 | 146.51 | 5.5–604 | ||||
| HT-2 | 2.7 | 12.65 | 2–151 | ||||
| β-ZOL | 6.7 | 2.5 | 2–23.2 | ||||
| ENB | 13.3 | 101.30 | 5–832 | ||||
| ENB1 | 1.3 | 7.80 | 5–117 | ||||
| ENB4 | 5.3 | 38.08 | 5–311 | ||||
| ENA1 | 4 | 6.58 | 5–125 | ||||
| BEA | 1.3 | 1.18 | 5–21.3 | ||||
| Portugal | Cereal baby food (maize, wheat, rice, barley, rye, oat, sorghum, millet, spelt) | DON | 9 | 44 | 173.13 | 0.37–270.57 | Pereira et al., 2015 [43] |
| 15AcDON | 11 | 30.94 | 2.50–30.94 | ||||
| T2-Tetrol | 11 | 112.18 | 10.48–112.18 | ||||
| NEO | 11 | 87.21 | 1.28–87.21 | ||||
| Portugal | Breakfast cereals for children (maize, wheat, rice, and multi-grain) | AFB1 | 26 | 0.028 | 0.040–0.400 | Assunção et al., 2015 [44] | |
| AFB2 | 0.002 | 0.030–0.300 | |||||
| AFG1 | 0.006 | 0.045–0.450 | |||||
| AFM1 | 0.012 | 0.100–1.000 | |||||
| OTA | 0.026 | 0.200–2.000 | |||||
| DON | 59 | 15–360 | |||||
| NIV | 6 | 25–360 | |||||
| FB1 | 13 | 2.5–8.0 | |||||
| FB2 | 3 | 2.5–8.0 | |||||
| Turkey | Baby food (cereal based supplementary foods for infants and children) | OTA | 50 | 68 | 0.034–0.374 | 0.042–0.380 | Hampikyan et al., 2015 [45] |
| Portugal | Children cereal-based food | PAT | 20 | 75 50 40 | 2.33 0.061 | 3.2–40.0 0.2–2.0 | Assunção et al., 2016 [46] |
| Portugal | Breakfast cereals | AFB1 | 26 | 69 | 0.013 | 0.003–0.130 | Martins et al., 2018 [47] |
| AFB2 | 27 | 0.004 | 0.001–0.011 | ||||
| AFG1 | 4 | 0.013 | 0.006–0.014 | ||||
| AFM1 | 12 | 0.017 | 0.011–0.240 | ||||
| OTA | 69 | 0.040 | 0.006–0.100 | ||||
| FB1 | 58 | 12.5 | 0.06–67.0 | ||||
| FB2 | 38 | 4.2 | 0.12–14.0 | ||||
| DON | 62 | 91.5 | 0.4–207.8 | ||||
| NIV | 4 | 27.1 | 5.6–27.1 | ||||
| ZEA | 19 | 0.7 | 0.12–5.6 | ||||
| Portugal | Cereal-based children food | AFB1 | 26 breakfast cereals | 73 | 0.036 | NM | Assunção et al., 2018 [48] |
| AFB2 | 46 | 0.07 | |||||
| AFG1 | 4 | NA | |||||
| AFM1 | 12 | 0.017 | |||||
| AFs | 73 | - | |||||
| OTA | 69 | 0.047 | |||||
| FB1 | 58 | 22.00 | |||||
| FB2 | 39 | 5.10 | |||||
| FMs | 58 | - | |||||
| ZEA | 73 | 1.20 | |||||
| DON | 62 | 95.9 | |||||
| NIV | 4 | NA | |||||
| AFB2 | 20 infant cereals (flours) | 5 | NA | NM | |||
| AFG1 | 10 | 0.014 | |||||
| AFM1 | 40 | 0.068 | |||||
| AFs | 45 | - | |||||
| OTA | 50 | 0.061 | |||||
| FB1 | 35 | 0.44 | |||||
| FMs | 35 | - | |||||
| ZEA | 30 | 0.48 | |||||
| DON | 20 | 41.8 | |||||
| OTAOTA | 6 biscuits | 100 | 0.086 | NM | |||
| DON | 50 | 43.8 | |||||
| Germany | Cereal-based baby food | AOH | 19 | 36.8 | 0.89 | 4.73–7.13 | Gotthardt et al., 2019 [49] |
| AME | 89.5 | 0.24 | 0.23–0.58 | ||||
| TEN | 94.7 | 1 | 0.18–7.53 | ||||
| ATX I | 15.8 | 0.17 | NA | ||||
| ATLP | 5.3 | 0.24 | NA | ||||
| TA | 50.2 | 5.66–221 | |||||
| Spain | Cereal-based baby food | AFB1 | 60 | 11 | 0.03 | 0.02–0.23 | Herrera et al., 2019 [50] |
| AFB2 | 1 | 0.01 | 0.02–0.20 | ||||
| AFG1 | 6 | 0.02 | 0.02–0.16 | ||||
| AFG2 | 1 | 0.01 | 0.02–0.11 | ||||
| DON | 12 | 37 | 33–245 | ||||
| Poland | Cereal-based infant and children food | DON | 302 | 17 | >LOD a <LOQ b | NM | Postupolski et al., 2019 [51] |
| NIV | 3 | ||||||
| ZEA | 14 | ||||||
| OTA | 4 | ||||||
| HT-2 | 0 | ||||||
| T-2 | 1 | ||||||
| FB1 | 3 | ||||||
| FB2 | 4 | ||||||
| Italy | Breakfast cereals Sweet cakes | OTA | 84 | 2.38 | 1 | NM | Capei et al., 2019 [52] |
| 35.7 | 1.34 | ||||||
| Austria and Czech Republic | Processed cereal-based infant foods | AFL | 35 | 6 | - | <LOQ–1.1 | Braun et al., 2021 [53] * |
| AFB1 | - | - | |||||
| STG | 23 | - | <LOQ–0.5 | ||||
| ZEA | 3 | 0.24 | 1.2 | ||||
| DON | 6 | - | 25–62 | ||||
| NIV | 6 | 43 | <LOQ–20 | ||||
| T-2 | 26 | - | 0.8–3.0 | ||||
| BEA | 14 | 1.5 | <LOQ–3.1 | ||||
| ENA | 3 | −1.9 | <LOQ | ||||
| ENB | 11 | 0.7 | <LOQ–2.1 | ||||
| ENA1 | 60 | 5.9 | <LOQ–40 | ||||
| ENB1 | 26 | 3.9 | <LOQ–10 | ||||
| FB1 | 20 | 4.8 | <LOQ–8.3 | ||||
| AME | 20 | 0.6 | <LOQ–1.1 | ||||
| TA | 31 | 48 | <LOQ–124 | ||||
| TEN | 34 | 0.9 | <LOQ–1.5 | ||||
| ATPL | 23 | 11 | <LOQ–20 | ||||
| Poland | Cereal-based baby foods | DON | 110 | 9.09 | 107.8 | 62–148 | Mruczyk et al., 2021 [54] |
| Country | Sample | Mycotoxin | Total Samples | Bioaccessibility (%) | References |
|---|---|---|---|---|---|
| The Netherlands | Infant formula (spaghetti Bolognese) supplemented with 2 mL sunflower oil per 100 g of food | AFB1 OTA | 2 | 88 ± 16–94 ± 8 29 ± 6–32 ± 4 | Kabak et al., 2009 [60] |
| Italy | Commercial pasta | DON | 6 | 2.12–41.5 | Raiola et al., 2012 [59] |
| Spain | Breakfast cereals Cookies Breads | ENA ENA1 ENB ENB1 | 14 | 50 ± 3–80 ± 3 40 ± 2–64 ± 2 43 ± 3–70 ± 3 46 ± 3–74 ± 2 | Prosperini et al., 2013 [61] |
| Portugal | Cereal-based food | PAT OTA PAT+OTA PAT+OTA | 6 | 30 ± 3–77 ± 2 95 ± 0–105 ± 2 33 ± 1–64 ± 2 (PAT) 103 ± 1–109 ± 0 (OTA) | Assunção et al., 2016 [46] |
| Matrix | Mycotoxin | Digestion Model | Extraction | Detection Method | References |
|---|---|---|---|---|---|
| Infant formula | AFB1 OTA | Static in vitro digestion model: Oral phase (KCl/KSCN/NaH2PO4/NaSO4/NaCl/NaHCO3/urea/a-amylase/uric acid/mucin) Gastric phase (NaCl/NaH2PO4/KCl/CaCl2/NH4Cl/HCl/glucose/glucuronic acid/urea/glucosamine hydrochloride/BSA/pepsin/mucin) Intestinal phase (NaCl/NaHCO3/KCl/HCl/urea/CaCl2(2H2O)/BSA/bile) | Phosphoric acid/chloroform + IAC AflaOchra HPLCTM | HPLC-FD | Kabak et al., 2009 [60] |
| Commercial pasta | DON | Static in vitro digestion model: Oral phase (KCl/KSCN/NaH2PO4/NaSO4/NaCl/NaHCO3/urea/a-amylase)Gastric phase (HCl/pepsin) Intestinal phase (NaHCO3/pancreatin/bile salts/H2O) | ACN:water (84:16; v/v) | LC-MS/MS | Raiola et al., 2012 [59] |
| Breakfast cereals Cookies Breads | ENA ENA1 ENB ENB1 | Static in vitro digestion model: Oral phase (KCl/KSCN/NaH2PO4/NaSO4/NaCl/NaHCO3/urea/a-amylase) Gastric phase (HCl/pepsin) Intestinal phase (NaHCO3/pancreatin/bile salts/H2O) | Ethyl acetate | LC-DAD | Prosperini et al., 2013 [61] |
| Cereal-based food | PAT | Static in vitro digestion model: Oral phase (KCl/KH2PO4/NaHCO3/MgCl2(H2O)6/(NH4)2CO3) Gastric phase (KCl/KH2PO4/NaHCO3/NaCl/MgCl2(H2O)6/(NH4)2CO3/pepsin) Intestinal phase (KCl/KH2PO4/NaHCO3/NaCl/MgCl2(H2O)6/pancreatin/bile) | Ethyl acetate + sodium sulphate + sodium hydrogenocarbonate + SPE column | RP-HPLC-UV | Assunção et al., 2016 [46] |
| OTA | MeOH:water (80:20) + IAC AflaOchra | RP-HPLC-FD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, C.; Cunha, S.C.; Fernandes, J.O. Mycotoxins of Concern in Children and Infant Cereal Food at European Level: Incidence and Bioaccessibility. Toxins 2022, 14, 488. https://doi.org/10.3390/toxins14070488
Pereira C, Cunha SC, Fernandes JO. Mycotoxins of Concern in Children and Infant Cereal Food at European Level: Incidence and Bioaccessibility. Toxins. 2022; 14(7):488. https://doi.org/10.3390/toxins14070488
Chicago/Turabian StylePereira, Cheila, Sara C. Cunha, and José O. Fernandes. 2022. "Mycotoxins of Concern in Children and Infant Cereal Food at European Level: Incidence and Bioaccessibility" Toxins 14, no. 7: 488. https://doi.org/10.3390/toxins14070488
APA StylePereira, C., Cunha, S. C., & Fernandes, J. O. (2022). Mycotoxins of Concern in Children and Infant Cereal Food at European Level: Incidence and Bioaccessibility. Toxins, 14(7), 488. https://doi.org/10.3390/toxins14070488