The Crystal Structure of Bacillus thuringiensis Tpp80Aa1 and Its Interaction with Galactose-Containing Glycolipids

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Tpp80Aa1 Has Activity against C. quinquefasciatus, Ae. aegypti, and An. gambiae Larvae and Mosquito Cell Lines
2.2. Tpp80Aa1 Binding Occurs Predominantly in the Posterior Midgut

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anopheles gambiae | Culex quinquefasciatus | Aedes aegypti | |
|---|---|---|---|
| Tpp80Aa1 | 8.6 | 24.6 | 29.6 |
| Cry48/Tpp49 | NT * | 0.02/0.006 [24] | NT * |
| Tpp1/Tpp2 | 0.013–0.03 [20] | 0.013–0.03 [24] | No toxicity to very low toxicity–depending on variant [25,26] |
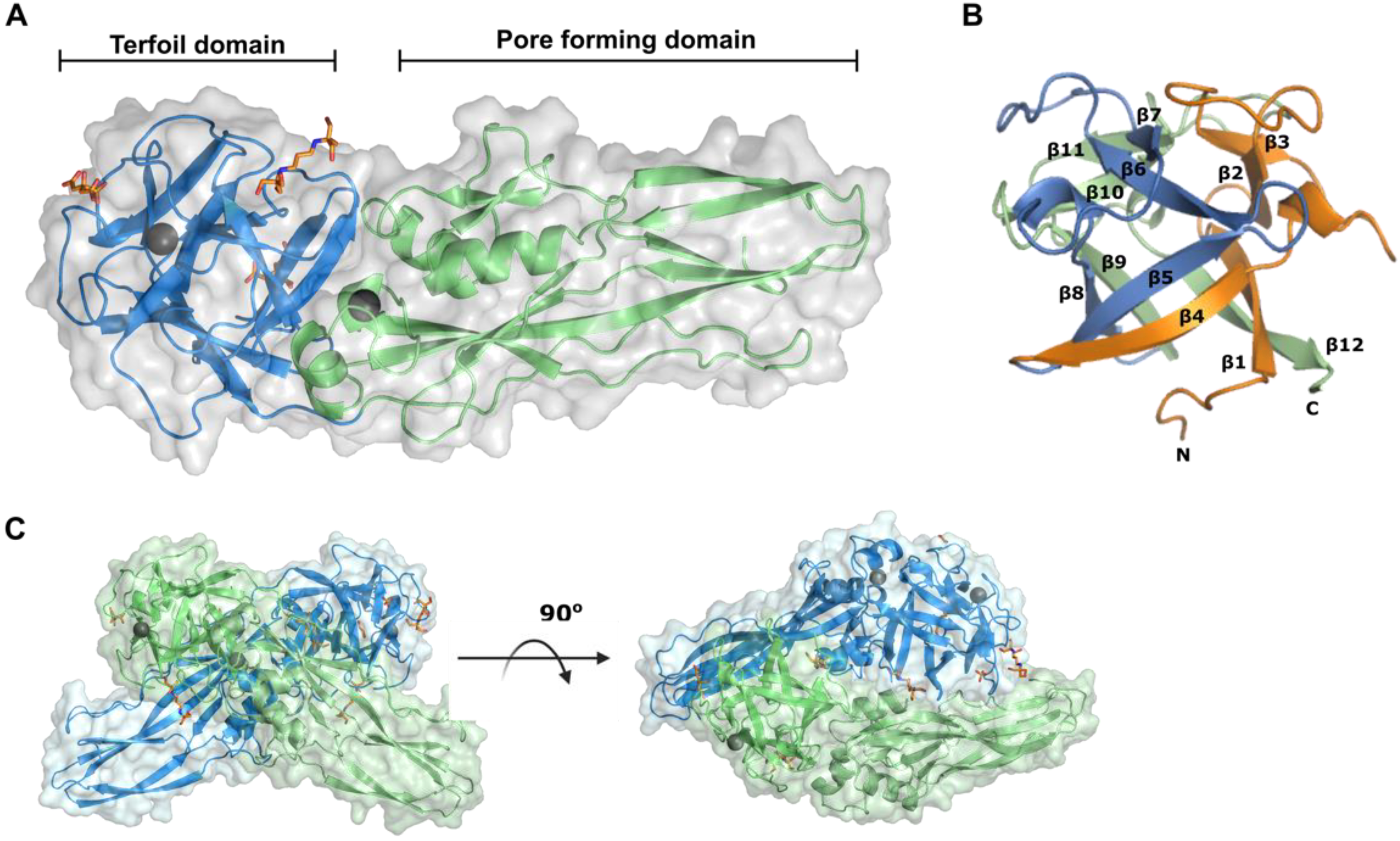
2.3. Tpp80Aa1 Structure Description
2.4. Interface Analysis
2.5. Tpp80Aa1 Has Structural Similarity with Other Tpp Proteins and Ricin B-like Lectin Domain-Containing Proteins
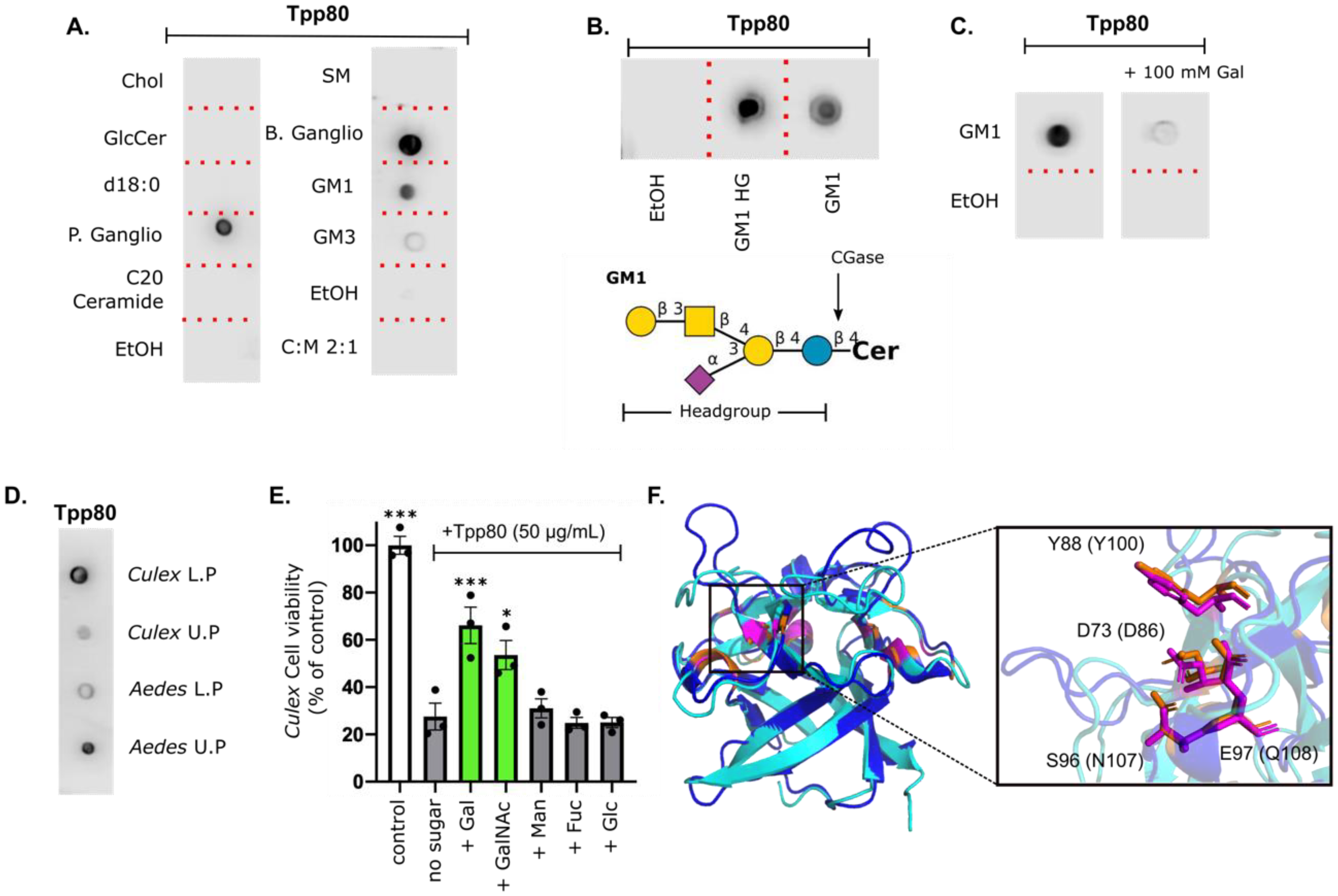
2.6. Tpp80Aa1 Binds Galactose-Containing Glycolipids and Lipids from Target Species
2.7. Galactose Competition Reduces Tpp80Aa1 Toxicity in Mosquito Cell Lines
3. Discussion
4. Materials and Methods
4.1. Tpp80Aa1 Expression and Purification
4.2. Bioassays (Insects & Cells)
4.3. Tpp80Aa1 Labelling
4.4. In Vivo Midgut Imaging
4.5. Crystallisation
4.6. Data Collection and Structure Determination
4.7. Structural Analysis
4.8. Lipid Extractions
4.9. Lipid Dot Blots
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [PubMed]
- Silva-Filha, M.; Romao, T.P.; Rezende, T.M.T.; Carvalho, K.D.S.; Gouveia de Menezes, H.S.; Alexandre do Nascimento, N.; Soberon, M.; Bravo, A. Bacterial Toxins Active against Mosquitoes: Mode of Action and Resistance. Toxins 2021, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, Z.Q.; Zhang, J.; Wan, Y.S.; Jin, W.J.; Li, Y.Z.; Fang, X.J. Cry80Aa1, a novel Bacillus thuringiensis toxin with mosquitocidal activity to Culex pipiens pallens. J. Invertebr. Pathol. 2020, 173, 107386. [Google Scholar] [CrossRef]
- Valtierra-de-Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance. Toxins 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A. Bacillus thuringiensis serovariety israelensis and Bacillus sphaericus for mosquito control. J. Am. Mosq. Control Assoc. 2007, 23, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, N.; Bone, E.J.; Williams, J.A.; Ellar, D.J. Contribution of the Individual Components of the Delta-Endotoxin Crystal to the Mosquitocidal Activity of Bacillus thuringiensis Subsp Israelensis. Fems. Microbiol. Lett. 1995, 131, 249–254. [Google Scholar]
- Oei, C.; Hindley, J.; Berry, C. Binding of purified Bacillus sphaericus binary toxin and its deletion derivatives to Culex quinquefasciatus gut: Elucidation of functional binding domains. J. Gen. Microbiol. 1992, 138, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Wirth, M.C.; Jiannino, J.A.; Federici, B.A.; Walton, W.E. Synergy between toxins of Bacillus thuringiensis subsp. israelensis and Bacillus sphaericus. J. Med. Entomol. 2004, 41, 935–941. [Google Scholar] [CrossRef][Green Version]
- Wirth, M.C.; Federici, B.A.; Walton, W.E. Cyt1A from Bacillus thuringiensis synergizes activity of Bacillus sphaericus against Aedes aegypti (Diptera: Culicidae). Appl. Envion. Microbiol. 2000, 66, 1093–1097. [Google Scholar] [CrossRef]
- Wirth, M.C.; Walton, W.E.; Federici, B.A. Cyt1A from Bacillus thuringiensis restores toxicity of Bacillus sphaericus against resistant Culex quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2000, 37, 401–407. [Google Scholar] [CrossRef]
- Mulla, M.S.; Thavara, U.; Tawatsin, A.; Chomposrf, J.; Su, T.Y. Emergence of resistance and resistance management in field populations of tropical Culex quinquefasciatus to the microbial control agent Bacillus sphaericus. J. Am. Mosq. Contr. 2003, 19, 39–46. [Google Scholar]
- Yuan, Z.M.; Zhang, Y.M.; Cai, Q.X.; Liu, E.Y. High-level field resistance to Bacillus sphaericus C3-41 in Culex quinquefasciatus from southern China. Biocontrol. Sci. Technol. 2000, 10, 41–49. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 186, 107438. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; Crickmore, N. Structural classification of insecticidal proteins—Towards an in silico characterisation of novel toxins. J. Invertebr. Pathol. 2017, 142, 16–22. [Google Scholar] [CrossRef]
- Cao, B.; Nie, Y.; Guan, Z.; Chen, C.; Wang, N.; Wang, Z.; Shu, C.; Zhang, J.; Zhang, D. The crystal structure of Cry78Aa from Bacillus thuringiensis provides insights into its insecticidal activity. Commun. Biol. 2022, 5, 801. [Google Scholar] [CrossRef] [PubMed]
- De Melo, J.V.; Jones, G.W.; Berry, C.; Vasconcelos, R.H.T.; de Oliveira, C.M.F.; Furtado, A.F.; Peixoto, C.A.; Silva, M.H.N.L. Cytopathological Effects of Bacillus sphaericus Cry48Aa/Cry49Aa Toxin on Binary Toxin-Susceptible and -Resistant Culex quinquefasciatus Larvae. Appl. Envion. Microb. 2009, 75, 4782–4789. [Google Scholar] [CrossRef]
- Opota, O.; Gauthier, N.C.; Doye, A.; Berry, C.; Gounon, P.; Lemichez, E.; Pauron, D. Bacillus sphaericus binary toxin elicits host cell autophagy as a response to intoxication. PLoS ONE 2011, 6, e14682. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.J.; Gill, S.S. An electron microscope study of the toxic action of Bacillus sphaericus in Culex quinquefasciatus larvae. J. Invertebr. Pathol. 1988, 52, 237–247. [Google Scholar] [CrossRef]
- De Melo, J.V.; Vasconcelos, R.H.; Furtado, A.F.; Peixoto, C.A.; Silva-Filha, M.H. Ultrastructural analysis of midgut cells from Culex quinquefasciatus (Diptera: Culicidae) larvae resistant to Bacillus sphaericus. Micron 2008, 39, 1342–1350. [Google Scholar] [CrossRef]
- Davidson, E.W. Variation in binding of Bacillus sphaericus toxin and wheat germ agglutinin to larval midgut cells of six species of mosquitoes. J. Invertebr. Pathol. 1989, 53, 251–259. [Google Scholar] [CrossRef]
- Lekakarn, H.; Promdonkoy, B.; Boonserm, P. Interaction of Lysinibacillus sphaericus binary toxin with mosquito larval gut cells: Binding and internalization. J. Invertebr. Pathol. 2015, 132, 125–131. [Google Scholar] [CrossRef]
- Dadd, R.H. Alkalinity within the midgut of mosquito larvae with alkaline-active digestive enzymes. J. Insect Physiol. 1975, 21, 1847–1853. [Google Scholar] [CrossRef] [PubMed]
- Boudko, D.Y.; Moroz, L.L.; Linser, P.J.; Trimarchi, J.R.; Smith, P.J.; Harvey, W.R. In situ analysis of pH gradients in mosquito larvae using non-invasive, self-referencing, pH-sensitive microelectrodes. J. Exp. Biol. 2001, 204, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.W.; Nielsen-Leroux, C.; Yang, Y.; Yuan, Z.; Dumas, V.F.; Monnerat, R.G.; Berry, C. A new Cry toxin with a unique two-component dependency from Bacillus sphaericus. Faseb. J. 2007, 21, 4112–4120. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; Hindley, J.; Ehrhardt, A.F.; Grounds, T.; de Souza, I.; Davidson, E.W. Genetic determinants of host ranges of Bacillus sphaericus mosquito larvicidal toxins. J. Bacteriol. 1993, 175, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Bideshi, D.K.; Federici, B.A. Recombinant strain of Bacillus thuringiensis producing Cyt1A, Cry11B, and the Bacillus sphaericus binary toxin. Appl. Envion. Microbiol. 2003, 69, 1331–1334. [Google Scholar] [CrossRef]
- Cummings, R.D.; Etzler, M.E. R-type Lectins. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor: New York, NY, USA, 2009. [Google Scholar]
- Cirauqui, N.; Abriata, L.A.; van der Goot, F.G.; Dal Peraro, M. Structural, physicochemical and dynamic features conserved within the aerolysin pore-forming toxin family. Sci. Rep. 2017, 7, 13932. [Google Scholar] [CrossRef]
- Szczesny, P.; Iacovache, I.; Muszewska, A.; Ginalski, K.; van der Goot, F.G.; Grynberg, M. Extending the aerolysin family: From bacteria to vertebrates. PLoS ONE 2011, 6, e20349. [Google Scholar] [CrossRef]
- Schrödinger, L.; DeLano, W. PyMOL. 2020. Available online: http://www.pymol.org/pymol (accessed on 1 November 2022).
- Williamson, L.J.; Galchenkova, M.; Best, H.L.; Bean, R.J.; Munke, A.; Awel, S.; Pena, G.; Knoska, J.; Schubert, R.; Doerner, K.; et al. Structure of the Lysinibacillus sphaericus Tpp49Aa1 pesticidal protein elucidated from natural crystals using MHz-SFX. bioRxiv 2022. [Google Scholar] [CrossRef]
- Colletier, J.P.; Sawaya, M.R.; Gingery, M.; Rodriguez, J.A.; Cascio, D.; Brewster, A.S.; Michels-Clark, T.; Hice, R.H.; Coquelle, N.; Boutet, S.; et al. De novo phasing with X-ray laser reveals mosquito larvicide BinAB structure. Nature 2016, 539, 43–47. [Google Scholar] [CrossRef]
- Ponstingl, H.; Henrick, K.; Thornton, J.M. Discriminating between homodimeric and monomeric proteins in the crystalline state. Proteins 2000, 41, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, L.; Lecroisey, A.; Charles, J.F. Role of the gut proteinases from mosquito larvae in the mechanism of action and the specificity of the Bacillus sphaericus toxin. Can. J. Microbiol. 1990, 36, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Broadwell, A.H.; Baumann, P. Proteolysis in the gut of mosquito larvae results in further activation of the Bacillus sphaericus toxin. Appl. Envion. Microbiol. 1987, 53, 1333–1337. [Google Scholar] [CrossRef]
- Treiber, N.; Reinert, D.J.; Carpusca, I.; Aktories, K.; Schulz, G.E. Structure and mode of action of a mosquitocidal holotoxin. J. Mol. Biol. 2008, 381, 150–159. [Google Scholar] [CrossRef]
- Matsushima-Hibiya, Y.; Watanabe, M.; Hidari, K.I.P.J.; Miyamoto, D.; Suzuki, Y.; Kasama, T.; Kanazawa, T.; Koyama, K.; Sugimura, T.; Wakabayashi, K. Identification of glycosphingolipid receptors for pierisin-1, a guanine-specific ADP-ribosylating toxin from the cabbage butterfly. J. Biol. Chem. 2003, 278, 9972–9978. [Google Scholar] [CrossRef] [PubMed]
- Cordara, G.; van Eerde, A.; Grahn, E.M.; Winter, H.C.; Goldstein, I.J.; Krengel, U. An Unusual Member of the Papain Superfamily: Mapping the Catalytic Cleft of the Marasmius oreades agglutinin (MOA) with a Caspase Inhibitor. PLoS ONE 2016, 11, e0149407. [Google Scholar] [CrossRef]
- Ichinose, H.; Kuno, A.; Kotake, T.; Yoshida, M.; Sakka, K.; Hirabayashi, J.; Tsumuraya, Y.; Kaneko, S. Characterization of an exo-beta-1,3-galactanase from Clostridium thermocellum. Appl. Env. Microb. 2006, 72, 3515–3523. [Google Scholar] [CrossRef]
- Candy, L.; Peumans, W.J.; Menu-Bouaouiche, L.; Astoul, C.H.; Van Damme, J.; Van Damme, E.J.; Erard, M.; Rouge, P. The Gal/GalNAc-specific lectin from the plant pathogenic basidiomycete Rhizoctonia solani is a member of the ricin-B family. Biochem. Biophys. Res. Commun. 2001, 282, 655–661. [Google Scholar] [CrossRef]
- Hazes, B. The (QxW)(3) domain: A flexible lectin scaffold. Protein Sci. 1996, 5, 1490–1501. [Google Scholar] [CrossRef]
- Rutenber, E.; Ready, M.; Robertus, J.D. Structure and evolution of ricin B chain. Nature 1987, 326, 624–626. [Google Scholar] [CrossRef]
- Hazes, B.; Read, R.J. A mosquitocidal toxin with a ricin-like cell-binding domain. Nat. Struct. Biol. 1995, 2, 358–359. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Hui, F.; Scheib, U.; Hu, Y.; Sommer, R.J.; Aroian, R.V.; Ghosh, P. Structure and glycolipid binding properties of the nematicidal protein Cry5B. Biochemistry 2012, 51, 9911–9921. [Google Scholar] [CrossRef]
- Rupar, M.J.; Donovan, W.P.; Groat, R.G.; Slaney, A.C.; Mattison, J.W.; Johnson, T.B.; Charles, J.F.; Dumanoir, V.C.; de Barjac, H. Two novel strains of Bacillus thuringiensis toxic to coleopterans. Appl. Envion. Microbiol. 1991, 57, 3337–3344. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Zhang, J.; Crickmore, N.; Song, F.; Gao, J.; Shu, C. Cry78Aa, a novel Bacillus thuringiensis insecticidal protein with activity against Laodelphax striatellus and Nilaparvata lugens. J. Invertebr. Pathol. 2018, 158, 1–5. [Google Scholar] [CrossRef]
- Cao, B.; Shu, C.; Geng, L.; Song, F.; Zhang, J. Cry78Ba1, One Novel Crystal Protein from Bacillus thuringiensis with High Insecticidal Activity against Rice Planthopper. J. Agric. Food Chem. 2020, 68, 2539–2546. [Google Scholar] [CrossRef]
- Nascimento, N.A.; Torres-Quintero, M.C.; Molina, S.L.; Pacheco, S.; Romao, T.P.; Pereira-Neves, A.; Soberon, M.; Bravo, A.; Silva-Filha, M. Functional Bacillus thuringiensis Cyt1Aa Is Necessary To Synergize Lysinibacillus sphaericus Binary Toxin (Bin) against Bin-Resistant and -Refractory Mosquito Species. Appl. Envion. Microbiol. 2020, 86, e02770-19. [Google Scholar] [CrossRef]
- Burton, S.L.; Ellar, D.J.; Li, J.; Derbyshire, D.J. N-acetylgalactosamine on the putative insect receptor aminopeptidase N is recognised by a site on the domain III lectin-like fold of a Bacillus thuringiensis insecticidal toxin. J. Mol. Biol. 1999, 287, 1011–1022. [Google Scholar] [CrossRef]
- Derbyshire, D.J.; Ellar, D.J.; Li, J. Crystallization of the Bacillus thuringiensis toxin Cry1Ac and its complex with the receptor ligand N-acetyl-D-galactosamine. Acta Cryst. D Biol. Cryst. 2001, 57, 1938–1944. [Google Scholar] [CrossRef]
- Sharma, M.; Gupta, G.D.; Kumar, V. Mosquito-larvicidal BinA toxin displays affinity for glycoconjugates: Proposal for BinA mediated cytotoxicity. J. Invertebr. Pathol. 2018, 156, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Helms, B. Host-pathogen interactions: Lipids grease the way. Eur. J. Lipid Sci. Technol. 2006, 108, 895–897. [Google Scholar] [CrossRef]
- Holmgren, J.; Lonnroth, I.; Mansson, J.; Svennerholm, L. Interaction of cholera toxin and membrane GM1 ganglioside of small intestine. Proc. Natl. Acad. Sci. USA 1975, 72, 2520–2524. [Google Scholar] [CrossRef] [PubMed]
- Scobie, H.M.; Rainey, G.J.; Bradley, K.A.; Young, J.A. Human capillary morphogenesis protein 2 functions as an anthrax toxin receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 5170–5174. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.S.; Ishii, S.; Shin, J.S.; Sung, K.I.; Park, B.S.; Jang, H.Y.; Kim, B.W. Globotriaosylceramide (Gb3) content in HeLa cells is correlated to Shiga toxin-induced cytotoxicity and Gb3 synthase expression. BMB Rep. 2009, 42, 310–314. [Google Scholar] [CrossRef]
- Yamaji, A.; Sekizawa, Y.; Emoto, K.; Sakuraba, H.; Inoue, K.; Kobayashi, H.; Umeda, M. Lysenin, a novel sphingomyelin-specific binding protein. J. Biol. Chem. 1998, 273, 5300–5306. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef]
- Bravo, A.; Gomez, I.; Conde, J.; Munoz-Garay, C.; Sanchez, J.; Miranda, R.; Zhuang, M.; Gill, S.S.; Soberon, M. Oligomerization triggers binding of a Bacillus thuringiensis Cry1Ab pore-forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains. Biochim. Biophys. Acta 2004, 1667, 38–46. [Google Scholar] [CrossRef]
- Gomez, I.; Sanchez, J.; Munoz-Garay, C.; Matus, V.; Gill, S.S.; Soberon, M.; Bravo, A. Bacillus thuringiensis Cry1A toxins are versatile proteins with multiple modes of action: Two distinct pre-pores are involved in toxicity. Biochem. J. 2014, 459, 383–396. [Google Scholar] [CrossRef]
- Jenkins, J.L.; Lee, M.K.; Valaitis, A.P.; Curtiss, A.; Dean, D.H. Bivalent sequential binding model of a Bacillus thuringiensis toxin to gypsy moth aminopeptidase N receptor. J. Biol. Chem. 2000, 275, 14423–14431. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Cryst. D Biol. Cryst. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Cryst. D Biol. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubak, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Cryst. D Biol. Cryst. 2011, 67, 355–367. [Google Scholar] [CrossRef]
- Holm, L. Using Dali for Protein Structure Comparison. Methods Mol. Biol. 2020, 2112, 29–42. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Grahn, E.; Askarieh, G.; Holmner, A.; Tateno, H.; Winter, H.C.; Goldstein, I.J.; Krengel, U. Crystal structure of the Marasmius oreades mushroom lectin in complex with a xenotransplantation epitope. J. Mol. Biol. 2007, 369, 710–721. [Google Scholar] [CrossRef]
- Skamnaki, V.T.; Peumans, W.J.; Kantsadi, A.L.; Cubeta, M.A.; Plas, K.; Pakala, S.; Zographos, S.E.; Smagghe, G.; Nierman, W.C.; Van Damme, E.J.; et al. Structural analysis of the Rhizoctonia solani agglutinin reveals a domain-swapping dimeric assembly. FEBS J. 2013, 280, 1750–1763. [Google Scholar] [CrossRef]
- Fujimoto, Z.; Kuno, A.; Kaneko, S.; Kobayashi, H.; Kusakabe, I.; Mizuno, H. Crystal structures of the sugar complexes of Streptomyces olivaceoviridis E-86 xylanase: Sugar binding structure of the family 13 carbohydrate binding module. J. Mol. Biol. 2002, 316, 65–78. [Google Scholar] [CrossRef]
- Ichinose, H.; Fujimoto, Z.; Honda, M.; Harazono, K.; Nishimoto, Y.; Uzura, A.; Kaneko, S. A beta-l-Arabinopyranosidase from Streptomyces avermitilis is a novel member of glycoside hydrolase family 27. J. Biol. Chem. 2009, 284, 25097–25106. [Google Scholar] [CrossRef] [PubMed]
- Sulzenbacher, G.; Roig-Zamboni, V.; Peumans, W.J.; Rouge, P.; Van Damme, E.J.; Bourne, Y. Crystal structure of the GalNAc/Gal-specific agglutinin from the phytopathogenic ascomycete Sclerotinia sclerotiorum reveals novel adaptation of a beta-trefoil domain. J. Mol. Biol. 2010, 400, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Kurre, D.; Suguna, K. Crystal structures of a beta-trefoil lectin from Entamoeba histolytica in monomeric and a novel disulfide bond-mediated dimeric forms. Glycobiology 2020, 30, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Renko, M.; Magister, S.; Smith, D.F.; Kunzler, M.; Strukelj, B.; Turk, D.; Kos, J.; Sabotic, J. Bivalent carbohydrate binding is required for biological activity of Clitocybe nebularis lectin (CNL), the N,N′-diacetyllactosediamine (GalNAcbeta1-4GlcNAc, LacdiNAc)-specific lectin from basidiomycete C. nebularis. J. Biol. Chem. 2012, 287, 10602–10612. [Google Scholar] [CrossRef] [PubMed]
- Terada, D.; Kawai, F.; Noguchi, H.; Unzai, S.; Hasan, I.; Fujii, Y.; Park, S.Y.; Ozeki, Y.; Tame, J.R. Crystal structure of MytiLec, a galactose-binding lectin from the mussel Mytilus galloprovincialis with cytotoxicity against certain cancer cell types. Sci. Rep. 2016, 6, 28344. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Yamasaki, T.; Eto, S.; Sugawara, H.; Kurisu, G.; Nakagawa, A.; Kusunoki, M.; Hatakeyama, T. Crystal structure of the hemolytic lectin CEL-III isolated from the marine invertebrate Cucumaria echinata: Implications of domain structure for its membrane pore-formation mechanism. J. Biol. Chem. 2004, 279, 37133–37141. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.; Lindqvist, Y.; Madrid, S.; Sandalova, T.; Fiskesund, R.; Yu, S.; Schneider, G. Crystal structure of bifunctional aldos-2-ulose dehydratase/isomerase from Phanerochaete chrysosporium with the reaction intermediate ascopyrone M. J. Mol. Biol. 2012, 417, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Kern, O.; Leon, P.C.V.; Gittis, A.G.; Bonilla, B.; Cruz, P.; Chagas, A.C.; Ganesan, S.; Ribeiro, J.M.C.; Garboczi, D.N.; Martin-Martin, I.; et al. The structures of two salivary proteins from the West Nile vector Culex quinquefasciatus reveal a beta-trefoil fold with putative sugar binding properties. Curr. Res. Struct. Biol. 2021, 3, 95–105. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Best, H.L.; Williamson, L.J.; Lipka-Lloyd, M.; Waller-Evans, H.; Lloyd-Evans, E.; Rizkallah, P.J.; Berry, C. The Crystal Structure of Bacillus thuringiensis Tpp80Aa1 and Its Interaction with Galactose-Containing Glycolipids. Toxins 2022, 14, 863. https://doi.org/10.3390/toxins14120863
Best HL, Williamson LJ, Lipka-Lloyd M, Waller-Evans H, Lloyd-Evans E, Rizkallah PJ, Berry C. The Crystal Structure of Bacillus thuringiensis Tpp80Aa1 and Its Interaction with Galactose-Containing Glycolipids. Toxins. 2022; 14(12):863. https://doi.org/10.3390/toxins14120863
Chicago/Turabian StyleBest, Hannah L., Lainey J. Williamson, Magdalena Lipka-Lloyd, Helen Waller-Evans, Emyr Lloyd-Evans, Pierre J. Rizkallah, and Colin Berry. 2022. "The Crystal Structure of Bacillus thuringiensis Tpp80Aa1 and Its Interaction with Galactose-Containing Glycolipids" Toxins 14, no. 12: 863. https://doi.org/10.3390/toxins14120863
APA StyleBest, H. L., Williamson, L. J., Lipka-Lloyd, M., Waller-Evans, H., Lloyd-Evans, E., Rizkallah, P. J., & Berry, C. (2022). The Crystal Structure of Bacillus thuringiensis Tpp80Aa1 and Its Interaction with Galactose-Containing Glycolipids. Toxins, 14(12), 863. https://doi.org/10.3390/toxins14120863