Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity


Abstract
1. Introduction
2. Bacterial Metabolites
3. Fungal Metabolites
4. Plant Metabolites
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations(FAO). Future of Food and Agriculture-Trends and Challenges; Food and Agriculture Organization of the United Nations(FAO): Rome, Italy, 2017. [Google Scholar]
- Rosegrant, M.W.; Cline, S.A. Global food security: Challenges and policies. Science 2003, 302, 1917–1919. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Caicedo, J.C.; Villamizar, S. Xanthomonas citri ssp. citri pathogenicity, a review. Citrus Res. Dev. Biotech. 2021, 135. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action mechanisms of effectors in plant-pathogen interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Lonigro, S.L.; Evidente, A.; Andolfi, A. Bacteriocin production by Pseudomonas syringae pv. ciccaronei NCPPB2355. Isolation and partial characterization of the antimicrobial compound. J. Appl. Microbiol. 1999, 86, 257–265. [Google Scholar]
- Evidente, A.; Abouzeid, A.M.; Andolfi, A.; Cimmino, A. Recent achievements in the bio-control of Orobanche infesting important crops in the Mediterranean basin. J. Agric. Sci. Technol. 2011, 1, 461–483. [Google Scholar]
- Pires, D.; Vicente, C.S.; Menéndez, E.; Faria, J.M.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Lorenzen, K.; Anke, K. Basidiomycetes as a source for new bioactive natural products. In Current Organic Chemistry; Mori, K., Ed.; Bentham Science Publisher: Miami, FL, USA, 1998; pp. 329–364. [Google Scholar]
- Anke, H.; Sterner, O. Insecticidal and nematocidal metabolites from fungi. In The Mycota; Osiewacz, H.D., Ed.; Springer: Berlin, Germany, 2002; Volume 10, pp. 109–127. [Google Scholar]
- Osmanova, N.; Schultze, W.; Ayoub, N. Azaphilones: A class of fungal metabolites with diverse biological activities. Phytochem. Rev. 2010, 9, 315–342. [Google Scholar] [CrossRef]
- Shen, W.; Mao, H.; Huang, Q.; Dong, J. Benzenediol lactones: A class of fungal metabolites with diverse structural features and biological activities. Eur. J. Med. Chem. 2015, 97, 747–777. [Google Scholar] [CrossRef]
- Mao, Z.; Sun, W.; Fu, L.; Luo, H.; Lai, D.; Zhou, L. Natural dibenzo-α-pyrones and their bioactivities. Molecules 2014, 19, 5088–5108. [Google Scholar] [CrossRef] [PubMed]
- Ghisalberti, E.L. Secondary metabolites with antinematodal activity. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 26, pp. 425–506. [Google Scholar]
- D’addabbo, T.; Carbonara, T.; Leonetti, P.; Radicci, V.; Tava, A.; Avato, P. Control of plant parasitic nematodes with active saponins and biomass from Medicago sativa. Phytochem. Rev. 2011, 10, 503–519. [Google Scholar] [CrossRef]
- Hernández-Carlos, B.; Gamboa-Angulo, M. Insecticidal and nematicidal contributions of Mexican flora in the search for safer biopesticides. Molecules 2019, 24, 897. [Google Scholar] [CrossRef] [PubMed]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the extremes of life: An untapped treasure for bioactive compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Martínez-Gómez, Á.; Fenoll, C.; Escobar, C. The use of biochar for plant pathogen control. Phytopathology 2021, 111, 1490–1499. [Google Scholar] [CrossRef]
- Snoop, W.; Mrozik, H.; Fisher, M.H. Structure and activity of avermectins and milbemycins in animal health. Vet. Parsitol. 1995, 59, 139–156. [Google Scholar]
- Hoshino, T. Violacein and related tryptophan metabolites produced by Chromobacterium violaceum: Biosynthetic mechanism and pathway for construction of violacein core. Appl. Microbiol. Biotechnol. 2011, 91, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Konzen, M.; De Marco, D.; Cordova, C.A.; Vieira, T.O.; Antonio, R.V.; Creczynski-Pasa, T.B. Antioxidant properties of violacein: Possible relation on its biological function. Bioorg. Med. Chem. 2006, 14, 8307–8313. [Google Scholar] [CrossRef]
- Becker, M.H.; Brucker, R.M.; Schwantes, C.R.; Harris, R.N.; Minbiole, K.P. The bacterially produced metabolite violacein is associated with survival of amphibians infected with a lethal fungus. Appl. Environ. Microbiol. 2009, 75, 6635–6638. [Google Scholar] [CrossRef]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef]
- Zeng, L.; Jin, H.; Lu, D.; Yang, X.; Pan, L.; Cui, H.; He, X.; Qiu, H.; Qin, B. Isolation and identification of chemical constituents from the bacterium Bacillus sp. and their nematicidal activities. J. Basic Microbiol. 2015, 55, 1239–1244. [Google Scholar] [CrossRef]
- Kosaka, H.; Aikawa, T.; Ogura, N.; Tabata, K.; Kiyohara, T. Pine wilt disease caused by the pine wood nematode: The induced resistance of pine trees by the avirulent isolates of nematode. Eur. J. Plant Pathol. 2001, 107, 667–675. [Google Scholar] [CrossRef]
- Song, M.J.; Bae, J.; Lee, D.S.; Kim, C.H.; Kim, J.S.; Kim, S.W.; Hong, S.I. Purification and characterization of prodigiosin produced by integrated bioreactor from Serratia sp. KH-95. JBB 2006, 101, 157–161. [Google Scholar]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zeng, Z.; Xue, B.; Deng, Y.; Sun, M.; Tang, Y.-J.; Ruana, L. Bacillus thuringiensis produces the lipopeptide thumolycin to antagonize microbes and nematodes. Microbiol. Res. 2018, 215, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Cabras, A.; Mannoni, M.A.; Serra, S.; Andolfi, A.; Fiore, M.; Evidente, A. Occurrence, isolation and biological activity of phytotoxic metabolites produced in vitro by Sphaeropsis sapinea, pathogenic fungus of Pinus radiata. Eur. J. Plant Pathol. 2006, 115, 187–193. [Google Scholar] [CrossRef]
- Kang, M.K.; Kim, J.H.; Liu, M.J.; Jin, C.Z.; Park, D.; Kim, J.; Sung, B.-H.; Kim, C.J.; Son, K.-H. New discovery on the nematode activity of aureothin and alloaureothin isolated from endophytic bacteria Streptomyces sp. AE170020. Sci. Rep. 2022, 12, 3947. [Google Scholar] [CrossRef]
- Anke, H.; Stadler, M.; Mayer, A.; Sterner, O. Secondary metabolites with nematicidal and antimicrobial activity from nematophagous fungi and Ascomycetes. Can. J. Bot. 1995, 73, 932–939. [Google Scholar] [CrossRef]
- Dasenbrock, J. Isolierung und Strukturautlclarung neuer Wirkstoffe aus Hijheren Pilzen. Ph.D. Thesis, University of Bonn, Bonn, Germany, 1994. [Google Scholar]
- Lorenzen, K.; Anke, T.; Anders, U.; Hindermayr, H.; Hansske, F. 14-Epidihydrocochlioquinone B, and 14-epicochlioquinone B, antibiotics from fermentations of the Ascomycete Neobulgaria pura: Structure elucidation and effects on platelet aggregation. Z. Naturforsch. 1994, 49C, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.M.; Frazier, E.G.; Bergstrom, A.R.; Williamson, J.M.; Liesch, J.M.; Goetz, M.A. Cochlioquinone A, a nematocidal agent which competes for specific [3H]ivermectin binding sites. J. Antibiot. 1990, 43, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C. Ivermectin: An update. Parasitol. Today 1985, 1, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Wada, K.; Munakata, K. New nematicidal metabolites from a fungus, Irpex lacteus. Agric. Biol. Chem. 1981, 45, 1527–1529. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. VII. Structure determination of brominated lachnumon and mycorrhizin A derivatives. J. Antibiot. 1995, 48, 158–161. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. 111. Production of novel isocoumarin derivatives. Isolation and biological activities. J. Antibiot. 1995, 48, 261–266. [Google Scholar] [CrossRef]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceurn (Karst.) Karst. IV. Structural elucidation of novel isocoumarin derivatives. J. Antibiot. 1995, 48, 267–270. [Google Scholar]
- Chexal, K.K.; Tamm, C. Biosynthesis of mikrolin. Helv. Chim. Acta 1978, 61, 2002–2018. [Google Scholar] [CrossRef]
- Chexal, K.K.; Tamm, C.; Clardy, J.; Hirotsu, K. Gilmicolin and mycorrhizinol, two new metabolites of Gilmaniella humicola Barron. Helv. Chim. Acta 1979, 62, 1130–1142. [Google Scholar] [CrossRef]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. V. Production, isolation and biological activities of bromine-containing mycorrhizin and lachnumon derivatives and four additional new bioactive metabolites. J. Antibiot. 1995, 48, 149–153. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. VI. Structure determination of non-halogenated metabolites structurally related to mycorrhizin A. J. Antibiot. 1995, 48, 154–157. [Google Scholar]
- Mayer, A.; Anke, H.; Sterner, O. Omphalotin, a new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat. Prod. Lett. 1997, 10, 25–32. [Google Scholar] [CrossRef]
- Kohno, J.; Nishio, M.; Sukarai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kish, N.; Yamashita, T.; Okuda, T.; Komatsubara, S. Isolation and structure determination of TMC-151s: Novel polyketide antibiotics from Gliocladium catenulatum Gilman & Abbott TC 1280. Tetrahedron 1999, 55, 7771–7786. [Google Scholar]
- Kohno, J.; Nishio, M.; Kish, M.N.; Komatsubara, S. Biosynthesis of the fungal polyketide antibiotics TMC151s: Origin of the carbon skeleton. J. Antibiot. 2000, 53, 1301–1304. [Google Scholar] [CrossRef]
- Kohno, J.; Asai, Y.; Nishio, M.; Sakurai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kishi, N.; Okuda, T.; Komatsubara, S. TMC-171A, B, C and TMC-154, novel polyketide antibiotics produced by Gliocladium sp. TC 1304 and TC 1282. J. Antibiot. 1999, 52, 1114–1123. [Google Scholar]
- Omura, S.; Tomoda, H.; Tabata, N.; Ohyama, Y.; Abe, T.; Namikoshi, M. Roselipins, novel fungal metabolites having a highly methylated fatty acid modified with a monnose and an arabinitol. J. Antibiot. 1999, 52, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, H.; Ohyama, Y.; Abe, T.; Tabata, N.; Namikoshi, M.; Yamaguchi, Y.; Masuma, R.; Omura, S. Roselipins, inhibitors of diacylglycerol acyltransferase, produced by Gliocladium roseum KF-1040. J. Antibiot. 1999, 52, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Tabata, N.; Ohyama, Y.; Tomoda, H.; Abe, T.; Namikoshi, M.; Omura, S. Structure elucidation of roselipins, inhibitors of diacylglycerol acyltransferase, produced by Gliocladium roseum KF-1040. J. Antibiot. 1999, 52, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, S.; Kanda, M.; Utagawa, M.; Chiba, N.; Ohtani, H.; Mikawa, T. MK7924, a novel metabolite with nematocidal activity from Coronophora gregaria. J. Antibiot. 2003, 56, 652–654. [Google Scholar] [CrossRef][Green Version]
- Zhu, Y.; Dong, J.; Wang, L.; Zhou, W.; Li, L.; He, H.; Liu, H.; Zhang, K. Screening and isolation of antinematodal metabolites against Bursaphelenchus xylophilus produced by fungi. Ann. Microbiol. 2008, 58, 375–380. [Google Scholar] [CrossRef]
- Mamiya, Y. The pine wood nematode. In Plant and Insect Nematodes; Nickle, W.R., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1984. [Google Scholar]
- Sutherland, J.R.; Webster, J.M. Nematode pests of forest trees. In Plant-Parasitic Nematodes in Temperate Agriculture; Evans, K., Trudgill, D.L., Webster, J.M., Eds.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Ferris, H.; Zheng, L. Plant sources of Chinese Herbal Remedies: Effectis on Pratylenchus vulnus and Meloidogyne javanica. J. Nematol. 1999, 31, 241–263. [Google Scholar] [PubMed]
- Dong, J.Y.; Song, H.C.; Li, J.H.; Tang, Y.S.; Sun, R.; Wang, L.; Zhou, Y.P.; Wang, M.L.; Shen, K.Z.; Wang, C.R.; et al. Ymf 1029A-E, preussomerin analogues from the fresh-water-derived fungus YMF 1.01029. J. Nat. Prod. 2008, 71, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.S.; Niu, X.M.; Wang, Y.L.; Guo, J.P.; Pan, W.Z.; Huang, X.-W.; Zhang, K.-Q. Isolation of putative biosynthetic intermediates of prenylated indole alkaloids from a thermophilic fungus Talaromyces thermophilus. Org. Lett. 2010, 12, 4356–4359. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.P.; Zhu, C.Y.; Zhang, C.P.; Chu, Y.S.; Wang, Y.L.; Zhang, J.-X.; Wu, D.-K.; Zhang, K.-Q.; Niu, X.-M. Thermolides, potent nematocidal PKS-NRPS hybrid metabolites from thermophilic fungus Talaromyces thermophilus. J. Am. Chem. Soc. 2012, 134, 20306–20309. [Google Scholar] [CrossRef]
- Zhang, J.M.; Wang, H.H.; Liu, X.; Hu, C.H.; Zou, Y. Heterologous and engineered biosynthesis of nematocidal polyketide-nonribosomal peptide hybrid macrolactone from extreme thermophilic fungi. J. Am. Chem. Soc. 2020, 142, 1957–1965. [Google Scholar] [CrossRef]
- Meng, X.; Mao, Z.; Lou, J.; Xu, L.; Zhong, L.; Peng, Y.; Zhou, L.; Wang, M. Benzopyranones from the endophytic fungus Hyalodendriella sp. Ponipodef12 and their bioactivities. Molecules 2012, 17, 11303–11314. [Google Scholar] [CrossRef]
- Liu, T.; Meyer, S.L.; Chitwood, D.J.; Chauhan, K.R.; Dong, D.; Zhang, T.; Li, J.; Liu, W.C. New nematotoxic indoloditerpenoid produced by Gymnoascus reessii za-130. J. Agric. Food Chem. 2017, 65, 3127–3132. [Google Scholar]
- Ashrafi, S.; Helaly, S.; Schroers, H.J.; Stadler, M.; Richert-Poeggeler, K.R.; Dababat, A.A.; Maier, W. Ijuhya vitellina sp. nov., a novel source for chaetoglobosin A, is a destructive parasite of the cereal cyst nematode Heterodera filipjevi. PLoS ONE 2017, 12, e0180032. [Google Scholar] [CrossRef]
- Kim, T.Y.; Jang, J.Y.; Yu, N.H.; Chi, W.J.; Bae, C.H. Nematicidal activity of grammicin produced by Xylaria grammica KCTC 13121BP against Meloidogyne incognita. Pest Manag. Sci. 2018, 74, 384–391. [Google Scholar] [CrossRef]
- Edwards, R.L.; Maitland, D.J.; Pittayakhajonwut, A.J.S.; Whalley, J. Metabolites of the higher fungi. Part 33. Grammicin, a novel bicyclic C7H6O4 furanopyranol from the fungus Xylaria grammica (Mont.) Fr. J. Chem. Soc. Perkin Trans. 2001, 1, 1296–1299. [Google Scholar] [CrossRef]
- Birkinshaw, J.H.; Michael, S.E.; Bracken, A.; Raistrick, H. Patulin in the common cold collaborative research on a derivative of Penicillium patulum Bainier. II. Biochem. Chem. Lancet 1943, 245, 625. [Google Scholar]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. Food Saf. 2005, 1, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Puel, O.; Galtie, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; Ariyawansa, H.A.; Phillips, A.J.; Wanasinghe, D.N.; Bhat, D.J.; McKenzie, E.H.C.; Camporesi, E.; Hide, K.D. Additions to Sporormiaceae: Introducing two novel genera, Sparticola and Forliomyces, from Spartium. Cryptogam. Mycol. 2016, 37, 75–97. [Google Scholar]
- Phukhamsakda, C.; Macabeo, A.P.G.; Huch, V.; Cheng, T.; Hyde, K.D. Stadler, M. Sparticolins A-G, biologically active oxidized spirodioxynaphthalene derivatives from the ascomycete. J. Nat. Prod. 2019, 82, 2878–2885. [Google Scholar] [CrossRef]
- Du, F.Y.; Ju, G.L.; Xiao, L.; Zhou, Y.M.; Wu, X. Sesquiterpenes and cyclodepsipeptides from marine-derived fungus Trichoderma longibrachiatum and their antagonistic activities against soil-borne pathogens. Mar. Drugs 2020, 18, 165. [Google Scholar] [CrossRef]
- Gonzalez, J.A.; Estevez-Braun, A.; Estevez-Reyes, R.; Bazzocchi, I.L.; Moujir, L.; Jimenez, I.A.; Ravelo, A.G.; Gonzalez, A.G. Biological activity of secondary metabolites from Bupleurum salicifolium (Umbelliferae). Experientia 1995, 51, 35–39. [Google Scholar]
- Bramwell, D.; BramwelI, Z. Wíld Flowers of the Canary Islands; Cabíldo Insular de Tenerífe: Santa Cruz de Tenerife, Spain, 1974. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Chicester, UK, 2002. [Google Scholar]
- Atkinson, H.J.; Taylor, J.D. Evidence for a calcium-binding site on the eggshell of Globodera rostochiensis with a role in hatching. Ann. Appl. Biol. 1980, 96, 307–315. [Google Scholar] [CrossRef]
- Atkinson, H.J.; Taylor, J.D. A calcium-binding sialoglycoprotein associated with an apparent eggshell membrane of Globodera rostochiensis. Ann. Appl. Biol. 1983, 102, 345–354. [Google Scholar] [CrossRef]
- Gonzalez, J.A.; Estevez-Braun, A. Phytonematicidal activity of aromatic compounds related to shikimate pathway. Pestic. Biochem. Phys. 1997, 58, 193–197. [Google Scholar] [CrossRef]
- Zasada, I.A.; Meyer, S.L.F.; Halbrendt, J.M.; Rice, C. Activity of hydroxamic acids from Secale cereale against the plant-parasitic nematodes Meloidogyne incognita and Xiphinema americanum. Phytopathology 2005, 95, 1116–1121. [Google Scholar] [CrossRef]
- Razavi, S.M. Plant counnarins as allelopathic agents. Int. J. Biol. Chem. 2011, 5, 86–90. [Google Scholar] [CrossRef]
- Cui, H.; Jin, H.; Liu, Q.; Yan, Z.; Ding, L.; Qin, B. Nematicidal metabolites from roots of Stellera chamaejasme against Bursaphelenchus xylophilus and Bursaphelenchus mucronatus. Pest Manag. Sci. 2014, 70, 827–835. [Google Scholar] [CrossRef]
- Kumarasingha, R.; Karpe, A.V.; Preston, S.; Yeo, T.C.; Lim, D.S.; Tu, C.L.; Kavlene, J.L.; Simpson, J.; Gasser, R.B.; Beale, P.D.; et al. Metabolic profiling and in vitro assessment of anthelmintic fractions of Picria fel-terrae Lour. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 171–178. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, G.; Huang, X.; Wang, Z.; Tan, N. 1, 4-naphthoquinone triggers nematode lethality by inducing oxidative stress and activating insulin/IGF signaling pathway in Caenorhabditis elegans. Molecules 2017, 22, 798. [Google Scholar] [CrossRef]
- Fang, N.; Casida, J.E. Anticancer action of cube’ insecticide: Correlation for rotenoid constituents between inhibition of NADH:ubiquinone oxidoreductase and induced ornithine decarboxylase activities. Proc. Natl. Acad. Sci. USA 1998, 95, 3380–3384. [Google Scholar] [CrossRef]
- Cabizza, M.; Angioni, A.; Melis, M.; Cabras, M.; Tuberoso, C.V.; Cabras, P. Rotenone and rotenoids in cub’e resins, formulations, and residues on olives. J. Agric. Food Chem. 2004, 52, 288–293. [Google Scholar] [CrossRef]
- Wenjie, J.; Fang, Y.; Gan, C.; Wu, Y.; Pang, J. Extraction and purification of deguelin from Derris trifoliata Lour root. Int. J. Agric. Biol. Eng. 2009, 2, 98–103. [Google Scholar]
- Chen, C.S.; Ho, D.R.; Chen, F.Y.; Chen, C.R.; Ke, Y.D.; Su, J.G.J. AKT mediates actinomycin D-induced p53 expression. Oncotarget 2014, 5, 693–703. [Google Scholar] [CrossRef]
- Vats, S.; Kamal, R. Cassia occidentalis L. (a new source of rotenoids): Itsin vitro regulation by feeding precursors and larvicidal efficacy. Plant Cell Tissue Organ Cult. 2014, 116, 403–409. [Google Scholar] [CrossRef]
- Ashack, R.J.; McCarty, L.P.; Malek, R.S.; Goodman, F.R.; Peet, N.P. Evaluation of rotenone and related compounds as antagonists of slow-reacting substance of anaphylaxis. J. Med. Chem. 1980, 23, 1022–1026. [Google Scholar] [CrossRef]
- Okombe Embeya, V.; Lumbu Simbi, J.B.; Stevigny, C.; Vandenput, S.; Pongombo Shongo, C.; Duez, P. Traditional plantbased remedies to control gastrointestinal disorders in livestock in the regions of Kamina and Kaniama (Katanga province, DemocraticRepublic of Congo. J. Ethnopharmacol. 2014, 153, 686–693. [Google Scholar] [CrossRef]
- Preston, S.; Korhonen, P.K.; Mouchiroud, L.; Cornaglia, M.; McGee, S.L.; Young, N.D.; Davis, R.A.; Crawford, S.; Nowell, C.; Ansell, B.R.E.; et al. Deguelin exerts potent nematocidal activity via the mitochondrial respiratory chain. FASEB J. 2017, 31, 4515–4532. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Rufino-González, Y.; Ponce-Macotela, M.; Delgado, G. In vitro activity of “Mexican Arnica” Heterotheca inuloides Cass natural products and some derivatives against Giardia intestinalis. Parasitology 2015, 142, 576–584. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Coballase-Urrutia, E.; Sicilia-Argumedo, G.; Ramírez-Apan, T.; Delgado, G. Toxicological evaluation of thenatural products and some semisynthetic derivatives of Heterotheca inuloides Cass (Asteraceae). J. Ethnopharmacol. 2015, 175, 256–265. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Franco-Navarro, F.; Delgado, G. In vitro nematicidal activity of natural and semisynthetic cadinenes from Heterotheca inuloides against the plant-parasitic nematode Nacobbus aberrans (Tylenchida: Pratylenchidae). Pest Manag. Sci. 2019, 75, 1734–1742. [Google Scholar] [CrossRef]
- Jang, J.Y.; Le Dang, Q.; Choi, G.J.; Park, H.W.; Kim, J.C. Control of root-knot nematodes using Waltheria indica producing 4-quinolone alkaloids. Pest Manag. Sci. 2019, 75, 2264–2270. [Google Scholar]
- Ismail, M.; Kowsar, A.; Javed, S.; Choudhary, M.I.; Khan, S.W.; Abbas, Q.; Tang, Y.; Wang, W. The antibacterial, insecticidal and nematocidal cctivities and toxicity studies of Tanacetum falconeri Hook. f. Turk. J. Pharm. Sci. 2021, 18, 744. [Google Scholar] [CrossRef]
- Velasco-Azorsa, R.; Zeferino-Díaz, R.; Alvarado-Rodríguez, J.G.; López-Ruiz, H.; Rojas-Lima, S.; Flores-Castro, K.; Cid del Prado-Vera, I.; Alatorre-Rosas, R.; Tut-Pech, F.; Carrillo-Benítez, M.G.; et al. Nematicidal activity of furanoeremophilenes against Meloidogyne incognita and Nacobbus aberrans. Pest Manag. Sci. 2022, 78, 2571–2580. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Source | Biological Activity | References |
|---|---|---|---|
| Metabolites Produced by Bacteria | |||
| Avermectin B1a (1) | Streptomyces avermentilis | Nematocidal activity against Haemonchus contortus, Ostertagia circumcincta; Trichostrongylus axei; Trichostrongylus colubriformis; Cooperia spp.; Oesophagostomum columbianu, (LD50 values in mice approximatively ranking between 15 and 50 mg/kg) and Meloidogyne incognita or Rotylenchulus reniformis (LD50/2 h 1.56 and 32.9 µg/mL , respectively) | [22] |
| milbemycin D (2) | Streptomyces hygroscopicus and Streptomyces cyaneogriseus | Nematocidal activity against Caenorhabditis elegans (EC50 89 nM) | [22] |
| Violacein (3) | Chromobacterium violaceum, Janthinobacterium lividum, and Pseudoalteromonas tunicata D2 Collimonas sp., Duganella sp. and Pseudoalteromonas spp. | Antibacterial, antitrypanocidal, anti-ulcerogenic, anticancer, antioxidant, leishmanicidal, antifungal, and antiviral | [24] [27] |
| Oxidative stress resistance, | [24] | ||
| defense of amphibians from fungal disease | [25] | ||
| Pseudoalteromonas tunicate | Antipredator defense mechanism against protozoan grazers | [26] | |
| Microbulbifer sp. D250 | Nematocidal activity against C. elegans (LC50 > 30 nM) | [27] | |
| 4-Oxabicyclo[3.2.2]nona-1(7), 5,8-triene (4) | Bacillus strain SMrs28 | Nematocidal activity against B. sxylophilus and D. destructor (LD50 904.12 and 1594.0 μg/mL/72 h, respectively) | [28] |
| (3S,8aS)-Hexahydro-3-methylpyrro[1,2-a]pyrazine-1,4-dione (5) | “ | Nematocidal activity against B. sxylophilus and D. destructor (LD50 451.26 and 366.62 μg/mL/72 h, respectively) | “ |
| Phenylacetamide (6) | “ | Nematocidal activity against B. sxylophilus and D. destructor (LD50 232.98 and 206.38 μg/mL/72 h, respectively) “ | “ |
| Cyclo(L-Pro-L-Val) (7) | “ | No tested | “ |
| Lauric acid (8) | “ | “ | “ |
| Methyl elaidate (9) | “ | “ | “ |
| Prodigiosin (10) | Serratia marcescens | Nematocidal activity against R. similis and M. javanica (LC50 83 and 79 μg/mL, respectively) | [31] |
| Thumolycin (11) | Bacillus thuringiensis | Nematocidal activity against C. elegans (600 U) | [32] |
| Aureothin (12) | Streptomyces sp. AE170020 | Nematocidal activity against B. xylophilus J2s, J3s and J4s (LC50 0.81, 1.15 and 1.54 μg/ L) | [34] |
| Alloaureothin (13) | “ | Nematocidal activity against B. xylophilus J2s, J3s and J4s (LC50 0.83, 1.10 and 1.47 μg/ L) | “ |
| Metabolites produced by fungi | |||
| 1-Methoxy-8-hydroxynaphthalene (14), | Daldinia concentrica | Cytotoxic, antibiotic, and nematocidal activity against C. elegans (LD50 10 μg/mL) | [35] [36] |
| 1,8-Dimethoxynaphthalene (15) | “ | Cytotoxic, antibiotic, and nematocidal activity against C. elegans (LD50 25 μg/mL) | “ |
| 5-Hydroxy-2-methylchromanone (16) | “ | Nematocidal activity C. elegans (LD50 > 100 μg/mL) | [36] |
| 14-epi-Dihydrocochlioquinone B (17) | Neobulgaria pura | Nematocidal against C. elegans (LD50 10 μg/mL) and M. incognita (LD30 100 μg/mL) | [35,37] |
| 14-epi-Cochlioquinone B (18) | “ | Nematocidal activity against C. elegans and M. incognita | “ |
| Cochlioquinone A (19) | Helminthosporium sp. | “ | [38] |
| Ivermectin (20) | Streptomyces avermitilis | “ | [39] |
| 5-Pentyl-2-furaldehyde (21) | Dermateaceae sp. Irpex lacteus | Nematocidal activity against C. elegans (LD50 60 μg/mL) and M. incognita (LD50 75 μg/mL) | [40] |
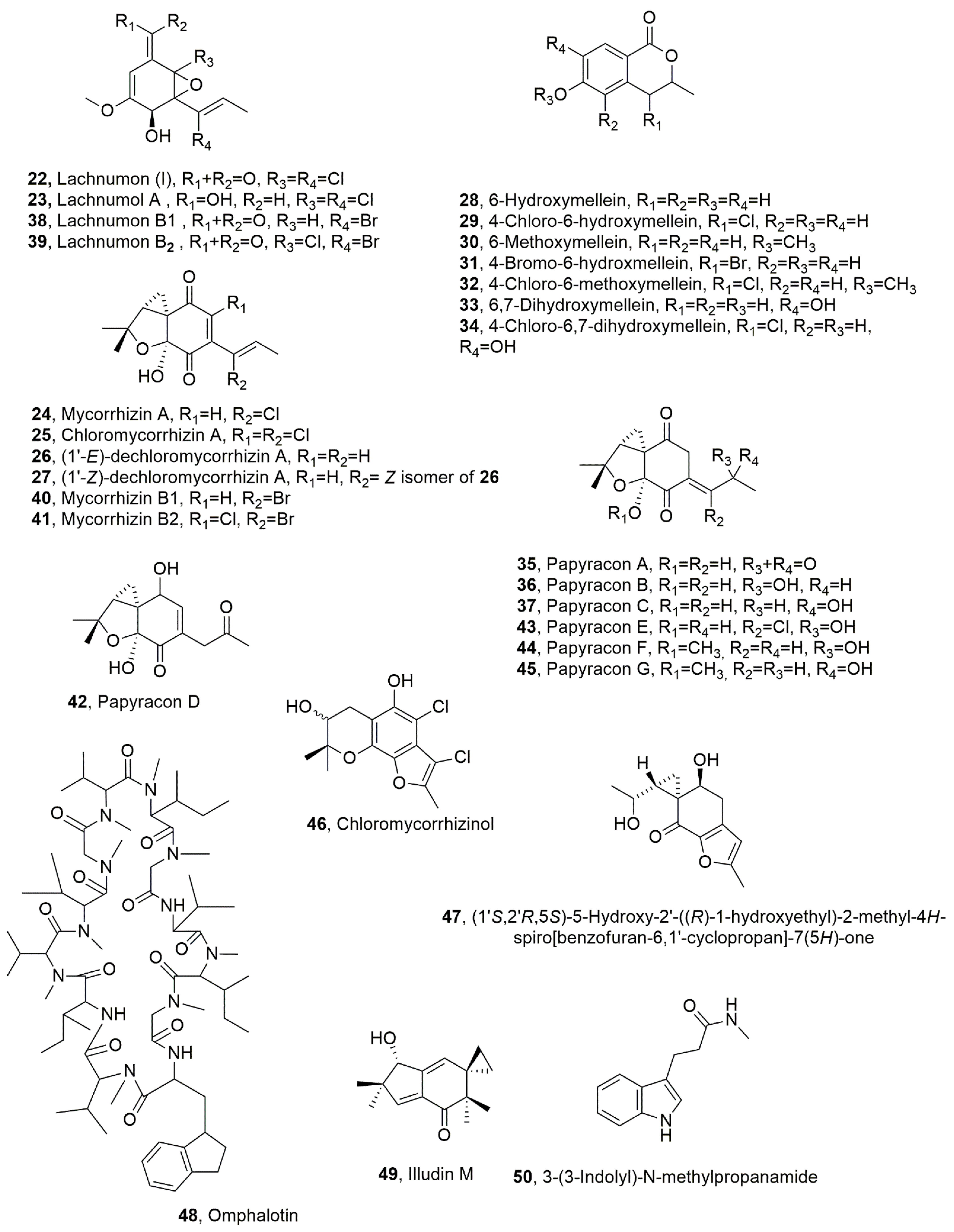
| Lachnumon (I) (22) | Lachnum papyraceum | Nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (LD50 > 100μg/mL) | [41,42] |
| Lachnumol A (23) | “ | Nematocidal activity against C. elegans (LD50 5 μg/mL) and M.incognita (LD50 100 μg/mL) | “ |
| Mycorrhizin A (24), | “ | Nematocidal activity against C. elegans (LD50 1 μg/mL) and M. incognita (LD50 100 μg/mL) | “ |
| Chloromycorrhizin A (25) | “ | Nematocidal activity against C. elegans (LD50 100 μg/mL) and M. incognita (LD50 100 μg/mL) | “ |
| (1’-E)-dechloromycorrhizin A (26) | “ | Nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100μg/mL) | “ |
| (1’-Z)-dechloromycorrhizin A (27) | “ | Nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| 6-Hydroxymellein (28) | “ | No nematocidal activity | [35] |
| 4-Chloro-6-hydroxymellein (29) | “ | “ | “ |
| 6-Methoxymellein (30) | “ | “ | “ |
| 4-Bromo-6-hydroxmellein (31) | “ | “ | “ |
| 4-Chloro-6-methoxymellein (23) | “ | “ | “ |
| 6,7-dihydroxymellein (33) | “ | ” | “ |
| 4-Chloro-6,7-dihydroxymellein (34) | “ | Nematocidal activity against C. elegans | “ |
| Papyracon A (35) | “ | Moderate nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon B (36) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon C (37) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Lachnumon B1 (38) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Lachnumon B2 (39) | “ | Moderate nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (Not tested) | “ |
| Mycorrhizin B1 (40) | “ | Moderate nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Mycorrhizin B2 (41) | “ | Moderate nematocidal activity against C. elegans (LD50 5 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon D (42) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon E (43) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon F (44) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon G (45) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Chloromycorrhizinol (46) | “ | Nematocidal activity against C. elegans (LD50 100 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| (1’S,2’R,5S)-5-Hydroxy-2’-((R)-1-hydroxyethyl)-2-methyl-4H-spiro[benzofuran-6,1’-cyclopropan]-7(5H)-one (47) | “ | Not tested | “ |
| Omphalotin (48) | Omphalotus olearius | Nematocidal activity against C. elegans (LD50 0.57 μg/mL) and M. incognita (LD50 18.95 μg/mL) | [48]. |
| Illudin M the (49) | “ | No activity | “ |
| 3-(3-Indolyl)-N-methylpropanamide (50) | “ | “ | “ |
| MK7924 (51) | Coronophora gregaria | Nematocidal activity against C. elegans at 100 μg/mL | [55] |
| 4,8-Dihydroxy-3,4-dihydronaphthalen-1(2H)-one (52), | Caryospora callicarpa | Nematocidal activity against B. xylophilus (LD50 540.2, 436.6 and 209.0 at 12, 24, and 36 h, respectively) | [56] |
| 4,5-Dihydroxy-3,4-dihydronaphthalen-1(2H)-one (53) | “ | Nematocidal activity against B. xylophilus (LD50 1169.8, 461.3, and 229.6 at 12, 24, and 36 h) | “ |
| 4,6,8-Trihydroxy-3,4-dihydronaphthalen-1(2H)-one (54) | “ | Nematocidal activity against B. xylophilus (LD50 1011.6, 522.5, and 220.3 at 12, 24, and 36 h) | “ |
| 3,4,6,8-Tetrahydroxy-3,4-dihydronaphthalen-1(2H)-one (55) | “ | Nematocidal activity against B. xylophilus (LD50 854, 468, and 206.1 at 12, 24 and 36 h) | “ |
| Ymf 1029 A (56) | Unidentified freshwater fungus YMF 1.01029 | Nematocidal activity against B. xylophilus | [60] |
| Ymf 1029 B (57) | “ | “ | “ |
| Ymf 1029 C (58) | “ | “ | “ |
| Ymf 1029 D (59) | “ | “ | “ |
| Ymf 1029 E (60) | “ | “ | “ |
| Preussomerin C (61) | “ | “ | “ |
| Preussomerin D (62) | “ | “ | “ |
| (4RS)-4,8-dihydroxy-3,4-dihydronaphthalen-1(2H)-one (63) | “ | “ | “ |
| 4,6,8-Trihydroxy-3,4-dihydronaphthalen-1(2H)-one (64) | “ | “ | “ |
| Thermolide A (65) | Talaromyces thermophilus | Nematocidal activity against M. incognita, B. siylopilus, and P. redivevus | [62] |
| Thermolide B (66) | “ | “ | “ |
| Thermolide C (67) | “ | “ | “ |
| Thermolide D (68) | “ | “ | “ |
| Thermolide E (69) | “ | Not tested | “ |
| Thermolide F (70) | “ | “ | “ |
| Palmariol B (71) | Hyalodendriella sp. | Antimicrobial, inhibition of acetylcholinesterase, and nematocidal activity against C. elegans (IC50 56.21 μg/mL) | [64] |
| 4-Hydroxymellein (72) | “ | Antibacterial and nematocidal activity against C. elegans (IC50 86.86 μg/mL) | “ |
| Alternariol 9-methyl ether (73) | Nematocidal activity against C. elegans (IC50 93.99 μg/mL) | “ | |
| Botrallin (74) | “ | Nematocidal activity against C. elegans (IC50 84.51 μg/mL) | “ |
| Gymnoascole acetate (75)) | Gymnoascus reessii za-130 | Nematocidal activity against M. incognita J2s (EC50 47.5 μg/mL) | [65] |
| Chaetoglobosin A (76) | Ijuhya vitellina | Nematocidal activity against H. filipjevi (at 50 μg/mL, it paralyzed the nematode) | [66] |
| 19-O-Acetylchaetoglobosin A (77) | “ | Nematocidal activity against H. filipjevi (at 100 μg/mL, it paralyzed the nematode) | “ |
| Grammicin (78) | Xylaria grammica | Low antibacterial and cytotoxic activity, but nematocidal activity against M. incognita J2 juvenile mortality and egg-hatching inhibition (EC50 15.95 and 5.87 μg/mL) | [67] |
| Patulin (79) | Penicillium griseofulvum and Penicillium expansum, and other fungal genera | Antibacterial, cytotoxic, and dematocidal activity against M. incognita J2 juvenile mortality and eggs-hatching inhibition (EC50/72 h 115.67 μg/mL) | [67,69,70] |
| Sparticolin A (80) | Sparticola junci | Nematocidal activity against C. elegans (LD50 50 μg/mL) | [73] |
| Sparticolin B (81) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin C (82) | “ | Nematocidal activity against C. elegans (LD50 25 μg/mL) | “ |
| Sparticolin D (83) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin E (84) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin F (85) | “ | Nematocidal activity against C. elegans (LD50 12.5 μg/mL) | “ |
| Sparticolin G (86) | “ | Antifungal and cytotoxic activity, but not tested for nematocidal activity | “ |
| Trichodermene (87) | Trichoderma longibrachiatum | Antifungal activity against Colletotrichum lagenarium | [74] |
| Sesquiterpene (88) | “ | “ | “ |
| Sesquiterpene (89) | “ | “ | “ |
| Sesquiterpene (90) | “ | No activity | “ |
| Sesquiterpene (91) | “ | No activity | “ |
| Sesquiterpene (92) | “ | No activity | “ |
| Cyclodepsipeptide (93) | “ | Nematocidal against M. incognita (IC50 149.2 μg/mL) | “ |
| Cyclodepsipeptide (94) | “ | Nematocidal against M. incognita (IC50 140.6 μg/mL) | “ |
| Cyclodepsipeptide (95) | “ | Nematocidal against M. incognita (IC50 198.7 μg/mL) | “ |
| Metabolites produced by plants | |||
| Guayarol (96) | Bupleurum salicifolium | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | [80] |
| Buplerol (97) | “ | “ | “ |
| Matairesinol (98) | “ | Nematocidal activity against G. pallida and G. rostochiensis (70% hatching reduction) | “ |
| Matairesinoldimethyl ether (99) | “ | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | “ |
| Bursehernin (100) | “ | Nematocidal activity against G. pallida and G. rostochiensis (550% hatching reduction; HID 16.42) | “ |
| Pliviatolide (101) | “ | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | “ |
| Thujaplicatin, methyl ether (102) | “ | “ | “ |
| 2-Chloro-matairesinol (103) | “ | “ | “ |
| Nortrachelogenin (104) | “ | “ | “ |
| Nortrachelogenin triacetate (105) | “ | “ | “ |
| 2-Hydroxythujaplican methyl ether (106) | “ | “ | |
| DIBOA (107) | Plant of Poaceae family | Nematocidal activity against M. incognita (LD50/168 h 74.3 μg/mL; J2 mortality LD50 20.9 μg/mL) and X. americanum (LD50/24 h 18.4 μg/mL) | [81] |
| DIMBOA (108) | “ | Nematocidal activity against M. incognita J2 mortality (LD 50 46.1 μg/mL) and X. americanum (LD50/24 h 48.3 μg/mL) | “ |
| BOA (109) | “ | No activity | “ |
| MBOA (110) | Nematocidal activity against M. incognita J2 mortality (LD50 49.2 μg/mL) | “ | |
| 8-Geranylpsoralen, (111) | Apiaceae, Rutacea, Asteraceae and Fabaceae plant families | Nematocidal activity against B. xylophilus LD50/72 h 188.3 μg/mL) and P. redivivus (LD50/72 h 117.5 μg/mL) | [82] |
| Imperatorin (112) | “ | Nematocidal activity against B. xylophilus LD50/72 h 161.7 μg/mL) and P. redivivus (LD50/72 h 179.0 μg/mL) | “ |
| Heraclenin (113) | “ | Nematocidal activity against B. xylophilus LD50/72 h 114.7 μg/mL) and P. redivivus (LD50/72 h 184.7 μg/mL) | “ |
| Ruixianglangdusu B (114) | Stellera chamaejasme L. | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure at 227.4, 71.6, and 15.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure at 1.8 × 103, 160.2, and 0.6 μM, respectively) | [83] |
| Umbelliferone (115) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 1.3 × 107, 5.7 × 107, and 3.3 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.6 × 103, 851, and 33.4 μM, respectively) | “ |
| Daphnoretin (116) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure 47.8, 3.1, and 2.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.3 × 106, 169.9, and 3.1 μM, respectively) | “ |
| Chamaejasmenine C (117) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 1.7 × 104, 1.1 × 104, and 65.3 μM) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 463.5, 156.7, and 0.05 μM, respectively) | “ |
| 7-Methoxyneochaejasmin A (118) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24 and 72 h exposure <0.001, 3.4 and 167.3 μM) and B. mucronatus (LC50 at 12, 24 and 72 h exposure 1.8 × 104, 384.2, and 151.1 μM) | “ |
| (+)-Chamaejasmine (119) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 16.5, 8.8, and 4.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 1.8 × 103, 1.6 × 103, and 5.1 × 103 μM, respectively) | “ |
| Chamaechromone (120) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 0.7, 10.3, and 36.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 327, 5.7, and 0.003 μM, respectively) | “ |
| Isosikokianin A (121) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 147.7, 385.2, and 2.2 × 102 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.6 × 104, 32.5, and 2.3 μM, respectively) | |
| 1,4-Naphthoquinone (122) | Rubia wallichiana | Nematocidal activity against N2L4 (kill LC50 value 42.26 μg/mL), N2 (inhibition of egg hatching LC50 34.83 μg/mL), M. incognita (LC50 33.51 μg/mL) | [85] |
| Denguelin (123) | Leguminosae spp. | Nematocidal activity against H. contortus (L3 mortality IC50/24,48,72 h 81, 54 and 21 μM) (L4 mortality IC50/24,48,72 h 11.39, 25.4 and 0.004 μM) | [93] |
| 4-Methoxyisocadalene (124) | Heterotheca inuloides | No activity | [96] |
| 7-Hydroxycadalene (125) | “ | Nematocidal activity against N. aberrans (mortality at J2 stage (LC50 31.30 mg/mL) | “ |
| (4R)-7-hydroxy-3,4-dihydrocadalene (126) | “ | Nematocidal activity against N. aberrans (mortality at J2 stage LC50 26.30 mg/mL) | “ |
| 1𝛼-Hydroxy-1(4H)-isocadalen-4-one (127) | “ | No activity | “ |
| (1R,4R)-1-Hydroxy-4H-1,2, 3,4-tetrahydrocadalen-15-oic acid (128) | “ | “ | “ |
| rac-3,7-Dihydroxy 3(4H)-isocadalen-4-one (129) | “ | “ | |
| 5′-Methoxywaltherione A (130) | Waltheria indica | Nematocidal activity against M. arenaria (EC50 0.25 μg/mL), M. hapla (E50 0.09 μg/mL), M. incognita a(E50 0.09 μg/mL), and B. xylophilus (E50 2.13 μg/mL) | [97] |
| Waltherione A (131) | “ | Nematocidal activity against M. arenaria (EC50 0.63 μg/mL), M. hapla (E50 1.74 μg/mL), M. incognita a(E50 0.27 μg/mL), and B. xylophilus (E50 3.54 μg/mL) | “ |
| Waltherione C (132) | “ | Nematocidal activity against M. arenaria (EC50 10.67 μg/mL), M. hapla (E50 19.79 μg/mL), M. incognita (E50 16.59 μg/mL), and B. xylophilus (EC50 790.85 μg/mL) | “ |
| cis-Dehydromatricaria ester (133) | Tanacetum falconeri | Nematocidal activity against M. incognita (EC50/24,36, 72 h 3.4, 0.18, and 0.04 mg/L) | [98] |
| 3β-Angeloyloxy-6β-hydroxyfuranoeremophil-1(10)-ene (134) | Senecio sinuatos | Nematocidal activity against M. incognita (at 10 mg/mL) | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evidente, A. Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins 2022, 14, 849. https://doi.org/10.3390/toxins14120849
Evidente A. Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins. 2022; 14(12):849. https://doi.org/10.3390/toxins14120849
Chicago/Turabian StyleEvidente, Antonio. 2022. "Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity" Toxins 14, no. 12: 849. https://doi.org/10.3390/toxins14120849
APA StyleEvidente, A. (2022). Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins, 14(12), 849. https://doi.org/10.3390/toxins14120849