Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position

 , , and
, , and
Abstract

1. Introduction
2. Results and Discussion
2.1. Preparation of NIV
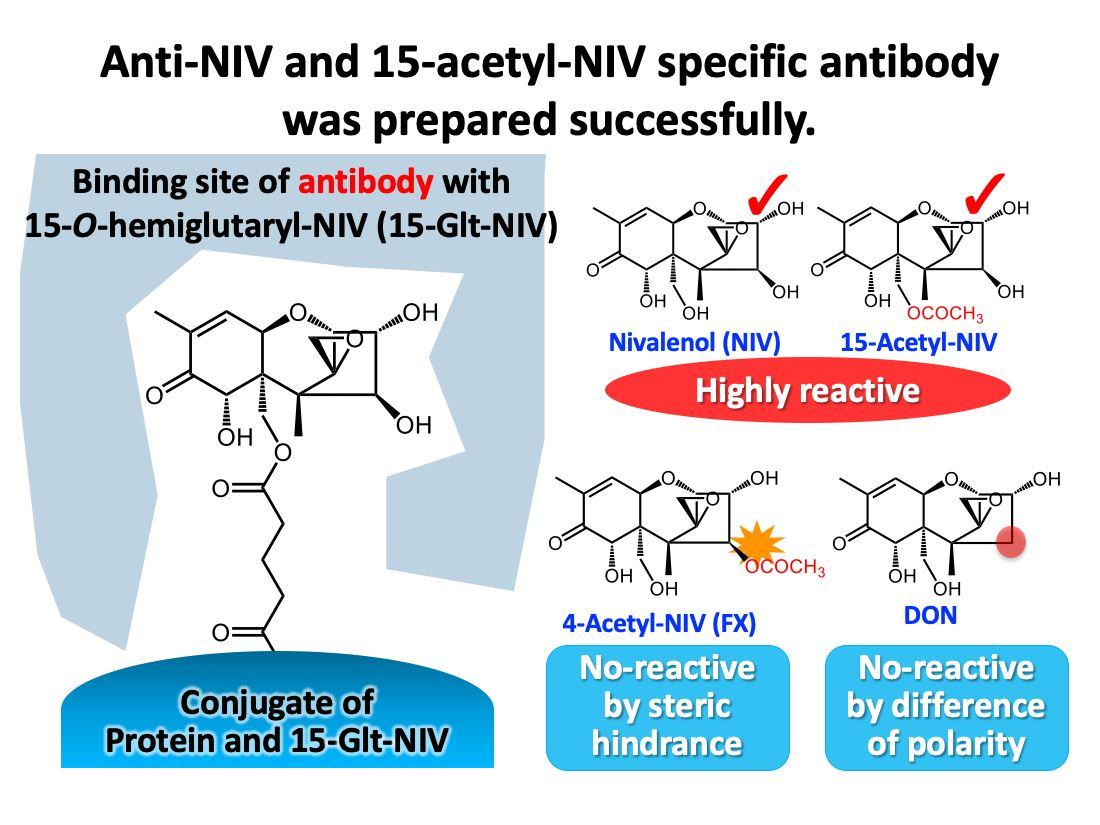
2.2. Introduction of a Carboxy Group into NIV
2.3. Preparation of PoAbs That React with NIV
2.4. Preparation of MoAbs That React with NIV
2.5. Reactivity of PoAbs and MoAbs with NIV Based on dc-ELISA
2.6. Reactivity of PoAbs and MoAbs with 15-Ac-NIV and Other Analogues of NIV
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Culture of Fusarium sp. for NIV Production
4.3. Preparation of NIV
4.4. HPLC Analysis of NIV
4.5. Synthesis of 15-Glt-NIV and 15-Ac-NIV
4.6. Preparation of 15-Glt-NIV–Protein Conjugate
4.7. Preparation of PoAbs and MoAbs
4.8. d-ELISA and dc-ELISA Creation
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAO Cereal Supply and Demand Brief. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 9 September 2022).
- Wieser, H.; Koehler, P.; Scherf, K.A. The two faces of wheat. Front. Nutr. 2020, 7, 517313. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Nakajima, T. Deoxynivalenol and nivalenol accumulation in wheat infected with Fusarium graminearum during grain development. Phytopathology 2010, 100, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.C.; Ohtsubo, K.; Izumiyama, N.; Nakamura, K.; Tanaka, T.; Yamamura, H.; Ueno, Y. The acute and chronic toxicities of nivalenol in mice. Fundam. Appl. Toxicol. 1988, 11, 38–47. [Google Scholar] [CrossRef]
- Scientific Committee on Food in European commission. Opinion of the Scientific Committee on Food on Fusarium Toxins. Part 6: Group Evaluation of T-2 Toxin, HT-2 Toxin, Nivalenol and Deoxynivalenol. Available online: https://ec.europa.eu/food/fs/sc/scf/out123_en.pdf (accessed on 9 September 2022).
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- Morrison, E.; Rundberget, T.; Kosiak, B.; Aastveit, A.H.; Berbholt, A. Cytotoxicity of trichothecenes and fusarochromanone produced by Fusarium equiseti strains isolated from Norwegian cereals. Mycopathologia 2002, 153, 49–56. [Google Scholar] [CrossRef]
- Lauren, D.R.; Greenhalgh, R. Simultaneous analysis of nivalenol and deoxynivalenol in cereals by liquid chromatography. J. Assoc. Off. Anal. Chem. 1987, 70, 479–483. [Google Scholar] [CrossRef]
- Walker, F.; Meier, B. Determination of the Fusarium mycotoxins nivalenol, deoxynivalenol, 3-acetyldeoxynivalenol, and 15-O-acetyl-4-deoxynivalenol in contaminated whole wheat flour by liquid chromatography with diode array detection and gas chromatography with electron capture detection. J. AOAC Int. 1998, 81, 741–748. [Google Scholar] [CrossRef]
- Aoyama, K.; Akashi, H.; Mochizuki, N.; Ito, Y.; Miyashita, T.; Lee, S.; Ogiso, M.; Maeda, M.; Kai, S.; Tanaka, H.; et al. Interlaboratory study of LC-UV and LC-MS methods for the simultaneous determination of deoxynivalenol and nivalenol in wheat. Shokuhin Eiseigaku Zasshi 2012, 53, 152–156. [Google Scholar] [CrossRef]
- Girolamo, A.D.; Ciasca, B.; Pascale, M.; Lattanzio, V.M.T. Determination of zearalenone and trichothecenes, including deoxynivalenol and its acetylated derivatives, nivalenol, T-2 and HT-2 toxins, in wheat and wheat products by LC-MS/MS: A collaborative study. Toxins 2020, 12, 786. [Google Scholar] [CrossRef]
- Pestka, J.J. Enhanced surveillance of foodborne mycotoxins by immunochemical assay. J. Assoc. Off. Anal. Chem. 1988, 71, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Fremy, J.M.; Usleber, E. Policy on characterization of antibodies used in immunochemical methods of analysis for mycotoxins and phycotoxins. J. AOAC Int. 2003, 86, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.M.T.; von Holst, C.; Lippolis, V.; Girolamo, A.D.; Logrieco, A.F.; Mol, H.G.J.; Pascale, M. Evaluation of mycotoxin screening tests in a verification study involving first time users. Toxins 2019, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.C.; Zhang, G.S.; Chu, F.S. Enzyme-linked immunosorbent assay for deoxynivalenol in corn and wheat. J. Assoc. Off. Anal. Chem. 1988, 71, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Abouzied, M.M.; Beremand, M.N.; McCormick, S.P.; Pestka, J.J. Reactivity of deoxynivalenol (vomitoxin) monoclonal antibody towards putative trichothecene precursors and shunt metabolites. J. Food Prot. 1991, 54, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.C.; Savard, M.E.; Lau, R. Production of monoclonal antibodies for the specific detection of deoxynivalenol and 15-acetyldeoxynivalenol by ELISA. J. Agric. Food Chem. 1995, 43, 1740–1744. [Google Scholar] [CrossRef]
- Maragos, C.M.; McCormick, S.P. Monoclonal antibodies for the mycotoxins deoxynivalenol and 3-acetyl-deoxynivalenol. Food Agric. Immunol. 2000, 12, 181–192. [Google Scholar] [CrossRef]
- Kohno, H.; Yoshizawa, T.; Fukugi, M.; Miyoshi, M.; Sakamoto, C.; Hata, N.; Kawamura, O. Production and characterization of monoclonal antibodies against 3,4,15-triacetylnivalenol and 3,15-diacetyldeoxynivalenol. Food Agric. Immunol. 2003, 15, 243–254. [Google Scholar] [CrossRef]
- Santos, J.S.; Takabayashi, C.R.; Ono, E.Y.S.; Itano, E.N.; Mallmann, C.A.; Kawamura, O.; Hirooka, E.Y. Immunoassay based on monoclonal antibodies versus LC-MS: Deoxynivalenol in wheat and flour in Southern Brazil. Food Addit. Contam. Part A 2011, 28, 1083–1090. [Google Scholar] [CrossRef]
- Sanders, M.; Guo, Y.; Iyer, A.; García, Y.R.; Galvita, A.; Heyerick, A.; Deforce, D.; Risseeuw, M.D.P.; van Calenbergh, S.; Bracke, M.; et al. An immunogen synthesis strategy for the development of specific anti-deoxynivalenol monoclonal antibodies. Food Addit. Contam. Part A 2014, 31, 1751–1759. [Google Scholar] [CrossRef]
- Maragos, C.; Busman, M.; Sugita-Konishi, Y. Production and characterization of a monoclonal antibody that cross-reacts with the mycotoxins nivalenol and 4-deoxynivalenol. Food Addit. Contam. 2006, 23, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, T.; Fujimoto, Y.; Morita, Y. Toxicological research on substances from fusarium nivale III.: The structure of nivalenol and its monoacetate. Tetrahedron Lett. 1969, 10, 2823–2826. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Morooka, N. Deoxynivalenol and its monoacetate: New mycotoxins from Fusarium roseum and moldy barley. Agric. Biol. Chem. 1973, 37, 2933–2934. [Google Scholar] [CrossRef]
- Onji, Y.; Aoki, Y.; Yamazoe, Y.; Dohi, Y.; Moriyama, T. Isolation of nivalenol and fusarenon-X from pressed barley culture by centrifugal partition chromatography. J. Liq. Chromatogr. 1988, 11, 2537–2546. [Google Scholar] [CrossRef]
- Tanaka, T.; Hasegawa, A.; Matsuki, Y.; Ishii, K.; Ueno, Y. Improved methodology for the simultaneous detection of the trichothecene mycotoxins deoxynivalenol and nivalenol in cereals. Food Addit. Contam. 1985, 2, 125–137. [Google Scholar] [CrossRef]
- Fruhmann, P.; Skrinjar, P.; Weber, J.; Mikula, H.; Warth, B.; Sulyok, M.; Krska, R.; Adam, G.; Rosenberg, E.; Hametner, C.; et al. Sulfation of deoxynivalenol, its acetylated derivatives, and T2-toxin. Tetrahedron 2014, 70, 5260–5266. [Google Scholar] [CrossRef]
- Weber, J.; Fruhmann, P.; Hametner, C.; Schiessl, A.; Häubl, G.; Fröhlich, J.; Mikula, H. Synthesis of isotope-labeled deoxynivalenol-15-O-glycosides. Eur. J. Org. Chem. 2017, 2017, 7012–7018. [Google Scholar] [CrossRef]
- Uchigashima, M.; Saigusa, M.; Yamashita, H.; Miyake, S.; Fujita, K.; Nakajima, M.; Nishijima, M. Development of a novel immunoaffinity column for aflatoxin analysis using an organic solvent-tolerant monoclonal antibody. J. Agric. Food Chem. 2009, 57, 8728–8734. [Google Scholar] [CrossRef]
- Kondo, M.; Tsuzuki, K.; Hamda, H.; Yamaguchi, Y.; Uchigashima, M.; Saka, M.; Watanabe, E.; Iwasa, S.; Narita, H.; Miyake, S. Development of an enzyme-linked immunosorbent assay (ELISA) for residue analysis of the fungicide azoxystrobin in agricultural products. J. Agric. Food Chem. 2012, 60, 904–911. [Google Scholar] [CrossRef]
- Miyake, S.; Hayashi, A.; Kita, H.; Ohkawa, H. Polyclonal and monoclonal antibodies for the specific detection of the herbicide acifluorfen and related compounds. Pestic. Sci. 1997, 51, 49–55. [Google Scholar] [CrossRef]
- Miyake, S.; Hayashi, A.; Kumeta, T.; Kitajima, K.; Kita, H.; Ohkawa, H. Effectiveness of polyclonal and monoclonal antibodies prepared for an immunoassay of the etofenprox insecticide. Biosci. Biotechnol. Biochem. 1998, 62, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- CODEX Alimentarius. General Standard for Contaminations and Toxins in Food and Feed. CXS 193-1995. Available online: https://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B193-1995%252FCXS_193e.pdf (accessed on 9 September 2022).
- Aoki, T.; O’Donnell, K. Fusarium kyushuense sp. nov. from Japan. Mycoscience 1998, 39, 1–6. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Yamasaki, T.; Harada, A.; Ohtake, T.; Adachi, K.; Iwasa, S.; Narita, H.; Miyake, S. Analysis of the fungicide boscalid in horticultural crops using an enzyme-linked immunosorbent assay and an immunosensor based on surface plasmon resonance. J. Agric. Food Chem. 2015, 63, 8075–8082. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Education, Culture, Sports, Science and Technology of Japan. Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions; Ministry of Education, Culture, Sports, Science and Technology of Japan: Tokyo, Japan, 2006. Available online: https://www.mext.go.jp/b_menu/hakusho/nc/06060904.htm (accessed on 9 September 2022). (In Japanese)





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NIV and Its Derivatives | PoAb | MoAb | |
|---|---|---|---|
| MNV80 | MNV87 | ||
| NIV | 70 * | 36 | 37 |
| 4-Ac-NIV | >1000 | >1000 | >1000 |
| 15-Ac-NIV | 11 | 18 | 18 |
| DON | >1000 | >1000 | >1000 |
| 3-Ac-DON | NT | >1000 | >1000 |
| 15-Ac-DON | NT | >1000 | >1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noda, K.; Hirakawa, Y.; Nishino, T.; Sekizuka, R.; Kishimoto, M.; Furukawa, T.; Sawane, S.; Matsunaga, A.; Kobayashi, N.; Sugita, K.; et al. Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position. Toxins 2022, 14, 747. https://doi.org/10.3390/toxins14110747
Noda K, Hirakawa Y, Nishino T, Sekizuka R, Kishimoto M, Furukawa T, Sawane S, Matsunaga A, Kobayashi N, Sugita K, et al. Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position. Toxins. 2022; 14(11):747. https://doi.org/10.3390/toxins14110747
Chicago/Turabian StyleNoda, Kyoko, Yuki Hirakawa, Tomomi Nishino, Ritsuto Sekizuka, Marin Kishimoto, Tomohiro Furukawa, Sakiko Sawane, Ayu Matsunaga, Naoki Kobayashi, Kazutoshi Sugita, and et al. 2022. "Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position" Toxins 14, no. 11: 747. https://doi.org/10.3390/toxins14110747
APA StyleNoda, K., Hirakawa, Y., Nishino, T., Sekizuka, R., Kishimoto, M., Furukawa, T., Sawane, S., Matsunaga, A., Kobayashi, N., Sugita, K., Oonaka, K., Kawakami, H., Otsuka, Y., Yamamoto, T., Yamamoto, T., Yoshiya, T., Watanabe, M., Saka, M., Momma, K., ... Miyake, S. (2022). Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position. Toxins, 14(11), 747. https://doi.org/10.3390/toxins14110747