New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1


 and
and
Abstract
:1. Introduction
2. Results and Discussion
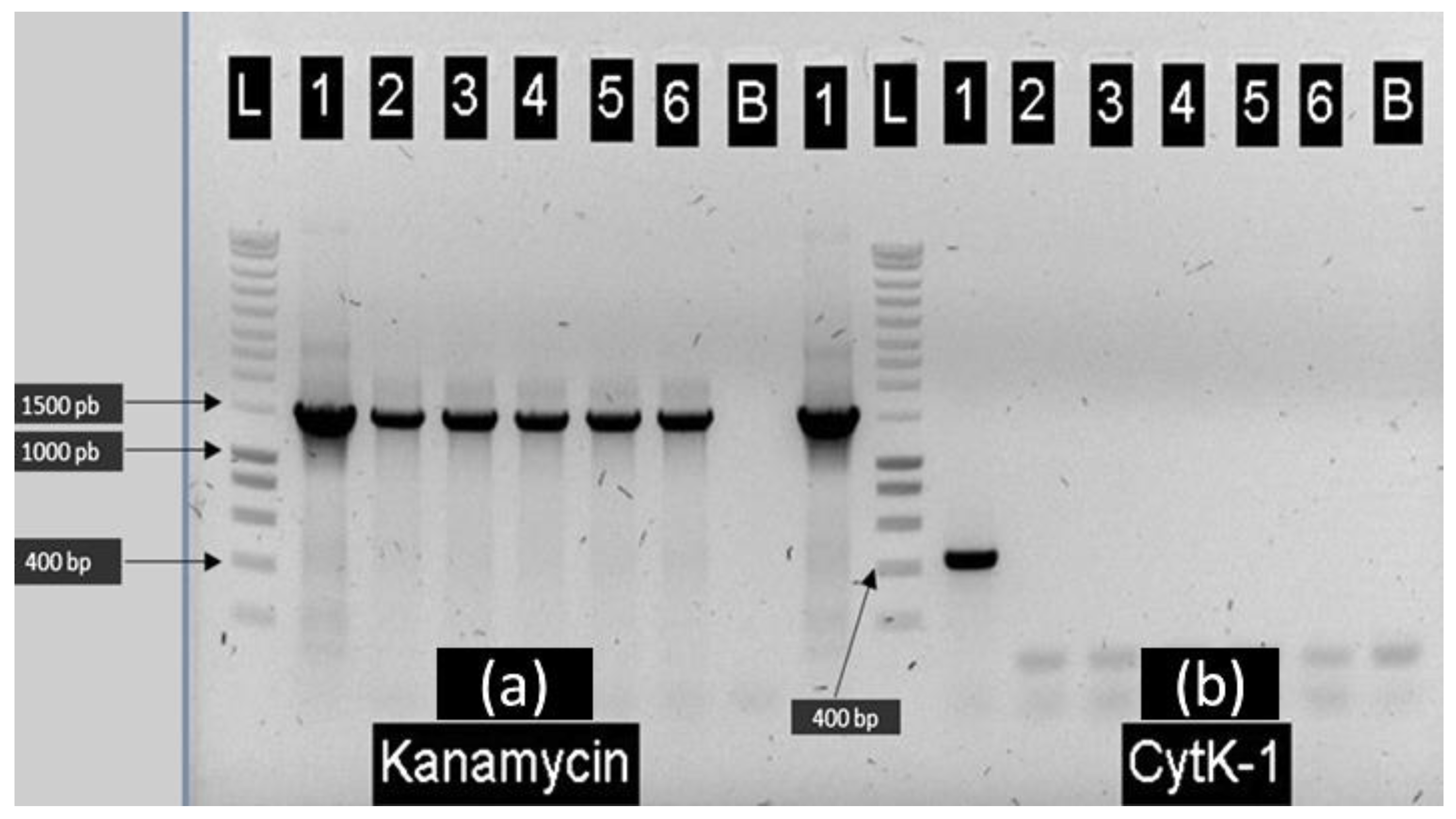
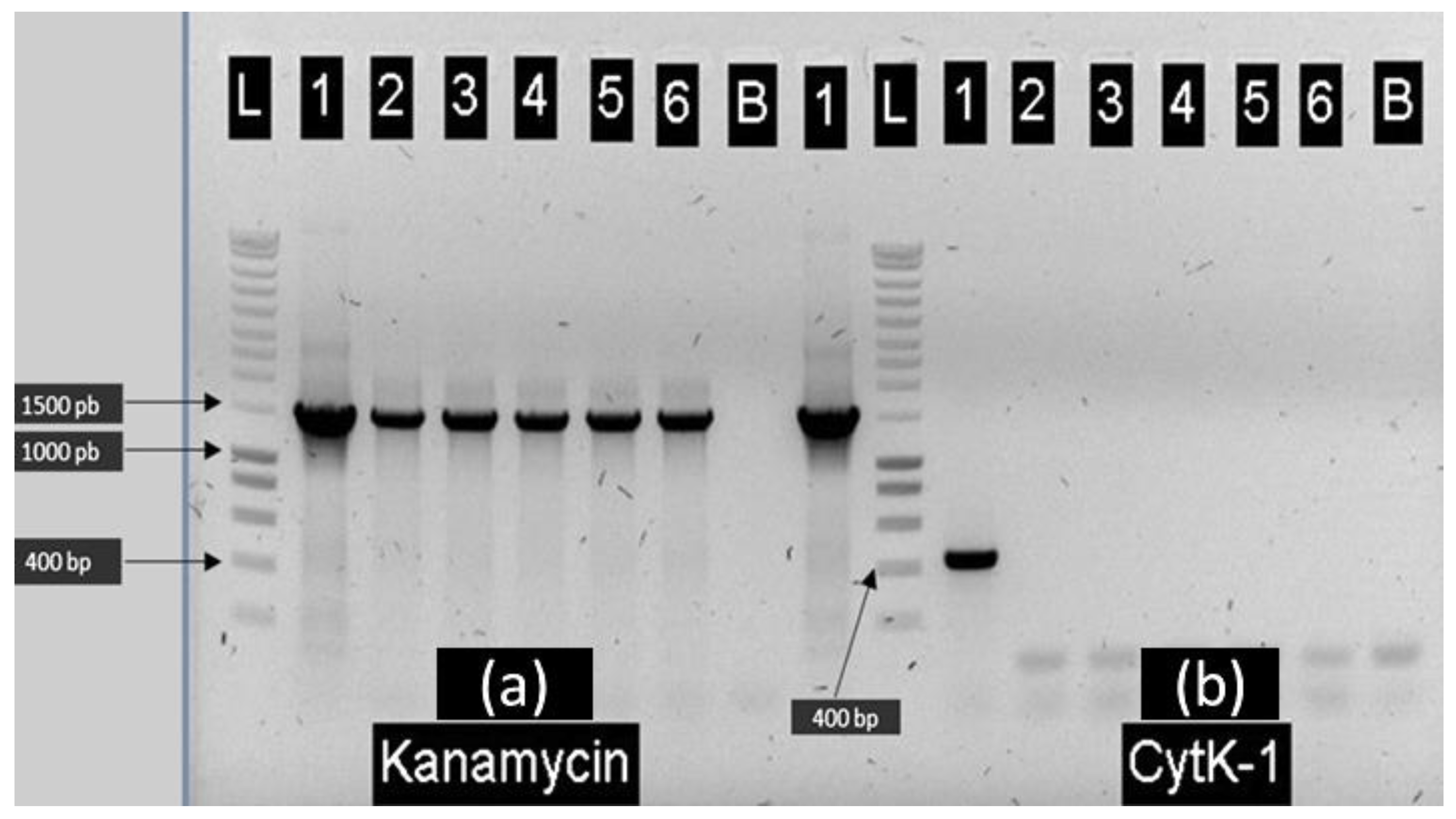
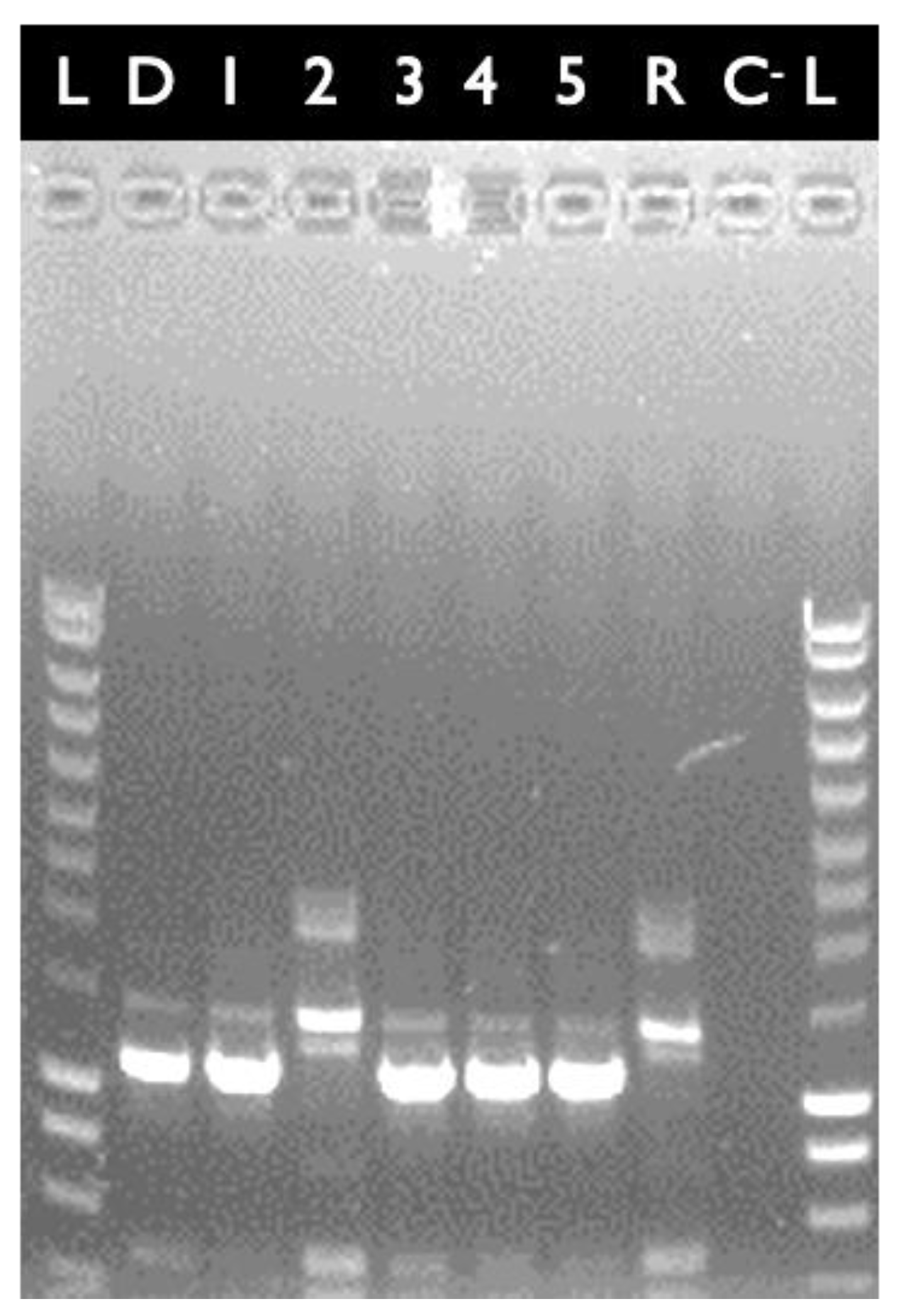
2.1. Construction of E28.3cytK-1 KO-A and E8.1cytK-1 KO B. cytotoxicus Mutants Lacking cytK-1
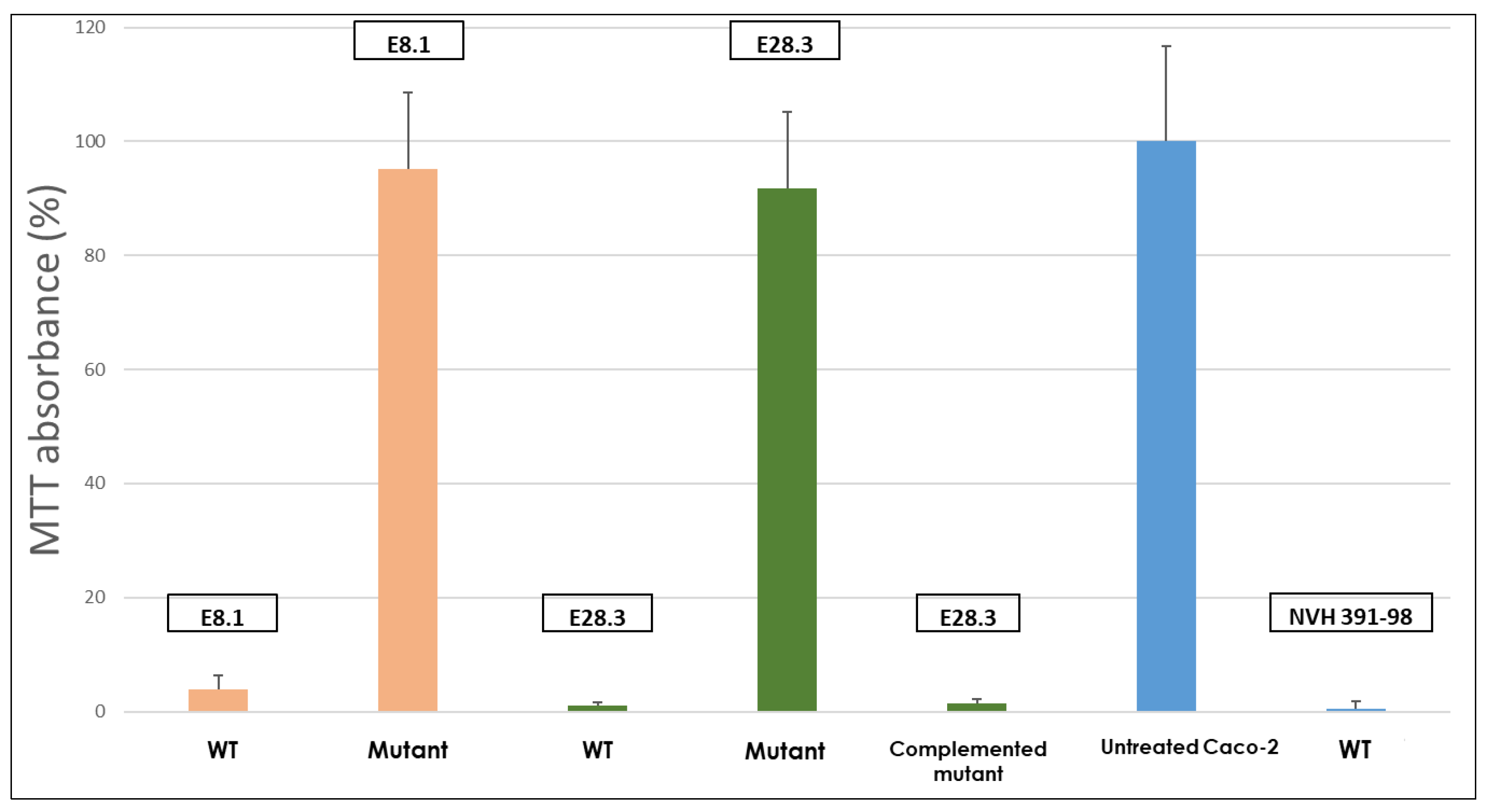
2.2. Cytotoxicity of B. cytotoxicus Wild-Type Strains E28.3 and E8.1, Their Derived Mutants Lacking the cytK-1 Gene and the Complemented Mutant
3. Conclusions
4. Materials and Methods
4.1. Bacterial Strains, Growth Media and Plasmids
4.2. Construction of the E28.3 Mutant Strain Lacking the cytK-1 Gene

{kind=link}
{kind=link}
{kind=link}
| Strains | Main Features | References |
|---|---|---|
| B. cytotoxicus strains | ||
| E8.1 | Wild-type strain isolated from potato flakes; RAPD pattern A, plasmid profile PP10 | [23] |
| E8.1cytK−1-KO-B | cytK-1 knockout mutant of E8.1 | This work |
| E28.3 | Wild-type strain isolated from potato flakes; RAPD pattern A and plasmid profile PP8 | [23] |
| E28.3cytK−1-KO-A | cytK-1 knockout mutant of E28.3 | This work |
| E28.3.1 | Derivative of E28.3 containing pXO16::Tn5401, TetR | [29] |
| E28.3.1cytK−1-KO-A | Derivative of E28.3cytK−1-KO-A containing pXO16::Tn5401, TetR | This work |
| E28.3cytK−1-KO-A(pHT304-18Z::cytK-1) | Complementation of the mutant lacking cytK-1 gene | This work |
| NVH 391-98 | Reference strain; highly cytotoxic | [1] |
| E. colistrains | ||
| E. coli C2925 dam−/dcm− | K12 derivative deficient in adenine and cytosine methyl-transferases, chemically competent | New England Biolabs™ |
| E. coli NEB 5-alpha | DH5-alpha derivative, T1 phage resistant and endA1 deficient, chemically competent | New England Biolabs™ |
| Plasmids | ||
| pHT304-18Z | Shuttle vector used in the complementation; EryR | [30] |
| pHT304-18Z::cytK-1 | CytK-1 gene of strain E28.3 cloned into the pHT304-18Z shuttle vector | This work |
| pMAD | Gram+/Gram- shuttle vector containing a thermo-sensitive Gram+ replicon; EryR, AmpR | [27] |
| pMAD::UD | pMAD derivative containing the KanR cassette with up- and down-stream regions of the cytK-1 gene, EryR, AmpR, KanR | This work |
| pDG783 | Plasmid carrying a KanR cassette; KanR | [28] |
| pXO16::Tn5401 | Large conjugative plasmid from B. thuringiensis sv. israelensis, tagged with a Tetracycline-resistance gene; TetR | [31] |
| Designation | Primers | Sequences (5′ to 3′) | Tm (°C) | Amplicon (pb) | Reference |
|---|---|---|---|---|---|
| KanR gene | Kana_R | GTTTTTTACTATCGATACAAATTCCTCGTAG | 60 | 1.490 | This study |
| Kana_F | ACATATATCGTGATAAACCCAGCGAACC | ||||
| CytK-1 upstream region | cytK-1_Up_R | GGGTTTATCACGATATATGTCGTATTTCACATATATC | 58 | 987 | This study |
| cytK-1_Up_F | AGATCTATCGATGCATGCCAGAAGTTTTAGGTTCATACATTTG | ||||
| CytK-1 downstream region | cytK-1_Down_R | CGGATCCATATGACGTCGACAATGCGAGAGACGTTGCG | 62 | 1.001 | This study |
| cytK-1_Down_F | TTGTATCGATAGTAAAAAACAACACTGACAAACTC | ||||
| CytK-1 gene | cytK-1_R | CCTCGTGCATCTGTTTCATGAG | 61 | 436 | [33] |
| cytK-1_F | CAATTCCAGGGGCAAGTGTC |
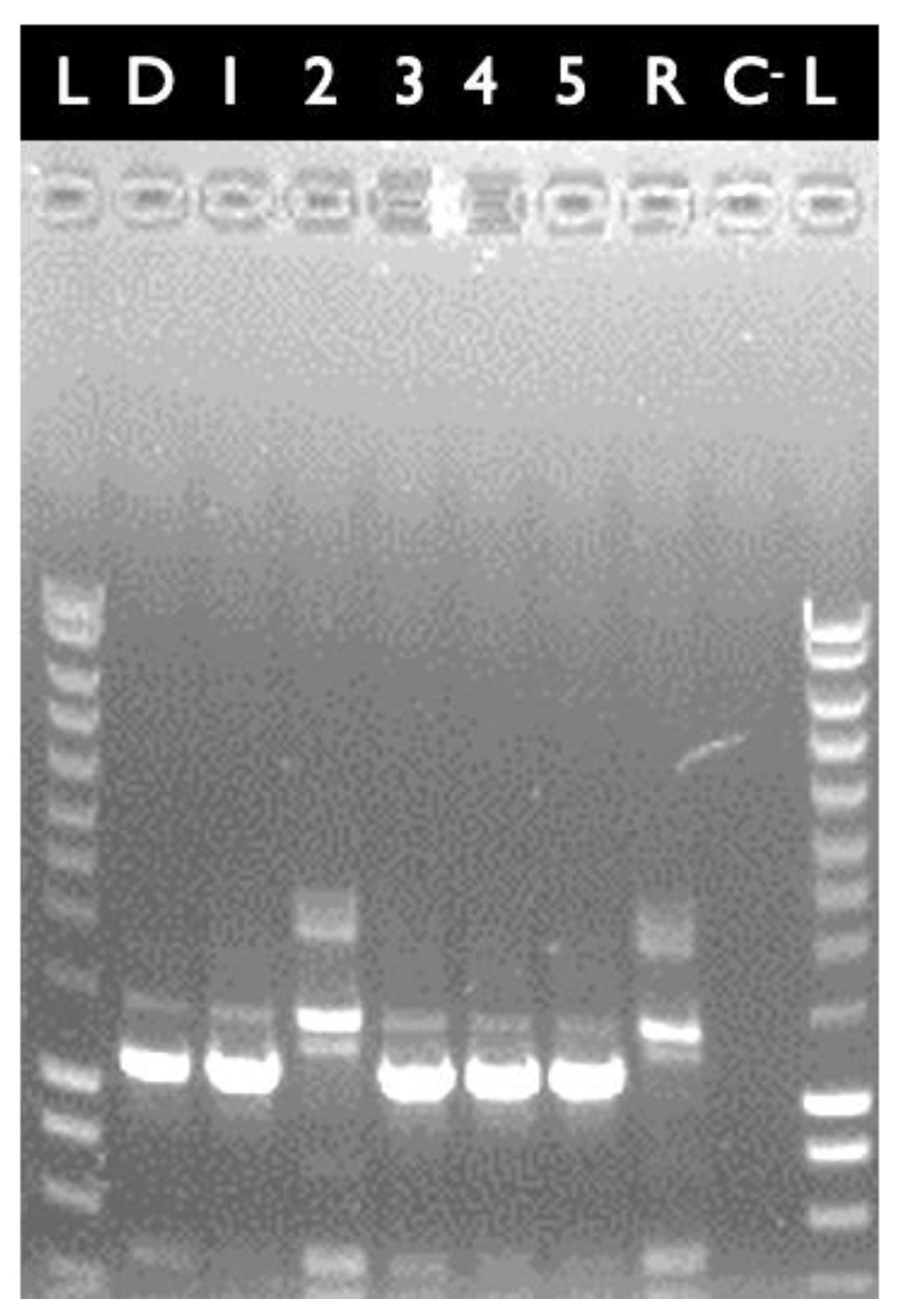
4.3. Transfer of the cytK-1 KO Locus from Strain E28.3 to Strain E8.1
4.4. Complementation of the cytK-1 KO Mutant
4.5. Cytotoxicity of E28.3, E8.1, Their Mutants Lacking cytK-1 and the Complemented Strain
4.5.1. Cell Culture
4.5.2. Preparation of Bacterial Cell-Free Supernatants
4.5.3. MTT Assay
4.5.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lund, T.; De Buyser, M.L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef]
- Guinebretière, M.H.; Auger, S.; Galleron, N.; Contzen, M.; De Sarrau, B.; De Buyser, M.L.; Lamberet, G.; Fagerlund, A.; Granum, P.E.; Lereclus, D.; et al. Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of the Bacillus cereus Group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 2013, 63, 31–40. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federici, B.A. Insecticidal bacteria: An overwhelming success for invertebrate pathology. J. Invert. Pathol. 2005, 89, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.C. Bacillus anthracis. J. Clin. Pathol. 2003, 56, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoton, F.M.; Andrup, L.; Swiecicka, I.; Mahillon, J. The cereulide genetic determinants of emetic Bacillus cereus are plasmid-borne. Microbiology 2005, 151, 2121–2124. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Fricker, M.; Scherer, S. Bacillus cereus, the causative agent of an emetic type of foodborne illness. Mol. Nutr. Food Res. 2004, 48, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Senesi, S.; Ghelardi, E. Production, secretion and biological activity of Bacillus cereus enterotoxins. Toxins 2010, 2, 1690–1703. [Google Scholar] [CrossRef]
- Gohar, M.; Faegri, K.; Perchat, S.; Ravnum, S.; Okstad, O.A.; Gominet, M.; Kolsto, A.B.; Lereclus, D. The PlcR virulence regulon of Bacillus cereus. PLoS ONE 2008, 3, e2793. [Google Scholar] [CrossRef]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.-L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.-A.; Ramarao, N.; et al. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: Epidemiology and genetic characterisation. Eur. Commun. Dis. Bull. 2016, 21, 30413. [Google Scholar] [CrossRef] [Green Version]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ding, S.; Shi, P.; Dietrich, R.; Martlbauer, E.; Zhu, K. Non-hemolytic enterotoxin of Bacillus cereus induces apoptosis in Vero cells. Cell Microbiol. 2017, 19, e12684. [Google Scholar] [CrossRef]
- Fox, D.; Mathur, A.; Xue, Y.; Liu, Y.; Tan, W.H.; Feng, S.; Pandey, A.; Ngo, C.; Hayward, J.; Atmosukarto, I.I.; et al. Bacillus cereus non-haemolytic enterotoxin activates the NLRP3 inflammasome. Nat. Commun. 2020, 11, 760. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerlund, A.; Brillard, J.; Fürst, R.; Guinebretière, M.-H.; Granum, P.E. Toxin production in a rare and genetically remote cluster of strains of the Bacillus cereus group. BMC Microbiol. 2007, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.J.A.; Tasara, T.; Klumpp, J.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Whole-genome-based phylogeny of Bacillus cytotoxicus reveals different clades within the species and provides clues on ecology and evolution. Sci. Rep. 2019, 9, 1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.J.A.; Johler, S. Draft genome sequence of CH_213, a highly cytotoxic Bacillus cytotoxicus strain isolated from mashed potatoes. Microbiol. Resour. Announc. 2020, 9, e00836-20. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; Van De Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef]
- Heini, N.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Characterization of Bacillus cereus group isolates from powdered food products. Int. J. Food Microbiol. 2018, 283, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Jessberger, N.; Dietrich, R.; Granum, P.E.; Martlbauer, E. The Bacillus cereus food infection as multifactorial process. Toxins 2020, 12, 701. [Google Scholar] [CrossRef]
- Burtscher, J.; Etter, D.; Biggel, M.; Schlaepfer, J.; Johler, S. Further Insights into the Toxicity of Bacillus cytotoxicus Based on Toxin Gene Profiling and Vero Cell Cytotoxicity Assays. Toxins 2021, 13, 234. [Google Scholar] [CrossRef]
- Koné, K.M.; Douamba, Z.; Halleux, M.; Bougoudogo, F.; Mahillon, J. Prevalence and diversity of the thermotolerant bacterium Bacillus cytotoxicus among dried food products. J. Food Prot. 2019, 82, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Makart, L.; Commans, F.; Gillis, A.; Mahillon, J. Horizontal transfer of chromosomal markers mediated by the large conjugative plasmid pXO16 from Bacillus thuringiensis serovar israelensis. Plasmid 2017, 91, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Koné, K.M.; Fayad, N.; Gillis, A.; Mahillon, J. Bacillus Cytotoxicus Genomics: Chromosomal Diversity and Plasmidome Versatility. under revision.
- Ramarao, N.; Lereclus, D. Adhesion and cytotoxicity of Bacillus cereus and Bacillus thuringiensis to epithelial cells are FlhA and PlcR dependent, respectively. Microbes Infect. 2006, 8, 1483–1491. [Google Scholar] [CrossRef]
- Arnaud, M.; Chastanet, A.; Debarbouillé, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, Gram-positive bacteria. Appl. Environ. Microbiol. 2004, 70, 6887–6891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerout-Fleury, A.M.; Shazand, K.; Frandsen, N.; Stragier, P. Antibiotic-resistance cassettes for Bacillus subtilis. Gene 1995, 167, 335–336. [Google Scholar] [CrossRef]
- Hinnekens, P.; Koné, K.M.; Fayad, N.; Leprince, A.; Mahillon, J. pXO16, the large conjugative plasmid from Bacillus thuringiensis serovar israelensis displays an extended host spectrum. Plasmid 2019, 102, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Agaisse, H.; Lereclus, D. Structural and functional analysis of the promoter region involved in full expression of the cryIIIA toxin gene of Bacillus thuringiensis. Mol. Microbiol. 1994, 13, 97–107. [Google Scholar] [CrossRef]
- Jensen, G.B.; Andrup, L.; Wilcks, A.; Smidt, L.; Poulsen, O.M. The aggregation-mediated conjugation system of Bacillus thuringiensis subsp. israelensis: Host range and kinetics of transfer. Cur. Microbiol. 1996, 33, 228–236. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., III; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Guinebretière, M.H.; Fagerlund, A.; Granum, P.E.; Nguyen-The, C. Rapid discrimination of cytK-1 and cytK-2 genes in Bacillus cereus strains by a novel duplex PCR system. FEMS Microbiol. Lett. 2006, 259, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahillon, J.; Lereclus, D. Electroporation of Bacillus thuringiensis and Bacillus cereus. In Electrotransformation of Bacteria; Eynard, N., Teissié, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 242–252. [Google Scholar]
- Turgeon, N.; Laflamme, C.; Ho, J.; Duchaine, C. Elaboration of an electroporation protocol for Bacillus cereus ATCC 14579. J. Microbiol. Methods 2006, 67, 543–548. [Google Scholar] [CrossRef]
- Makart, L.; Gillis, A.; Hinnekens, P.; Mahillon, J. A novel T4SS-mediated DNA transfer used by pXO16, a conjugative plasmid from Bacillus thuringiensis serovar israelensis. Environ. Microbiol. 2018, 20, 1550–1561. [Google Scholar] [CrossRef]
- Rajkovic, A.; Grootaert, C.; Butorac, A.; Cucu, T.; De Meulenaer, B.; van Camp, J.; Bracke, M.; Uyttendaele, M.; Bačun-Družina, V.; Cindrić, M. Sub-emetic toxicity of Bacillus cereus toxin cereulide on cultured human enterocyte-like Caco-2 cells. Toxins 2014, 6, 2270–2290. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koné, K.M.; Hinnekens, P.; Jovanovic, J.; Rajkovic, A.; Mahillon, J. New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1. Toxins 2021, 13, 698. https://doi.org/10.3390/toxins13100698
Koné KM, Hinnekens P, Jovanovic J, Rajkovic A, Mahillon J. New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1. Toxins. 2021; 13(10):698. https://doi.org/10.3390/toxins13100698
Chicago/Turabian StyleKoné, Klèma Marcel, Pauline Hinnekens, Jelena Jovanovic, Andreja Rajkovic, and Jacques Mahillon. 2021. "New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1" Toxins 13, no. 10: 698. https://doi.org/10.3390/toxins13100698
APA StyleKoné, K. M., Hinnekens, P., Jovanovic, J., Rajkovic, A., & Mahillon, J. (2021). New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1. Toxins, 13(10), 698. https://doi.org/10.3390/toxins13100698