Anthrax Edema and Lethal Toxins Differentially Target Human Lung and Blood Phagocytes

 and
and 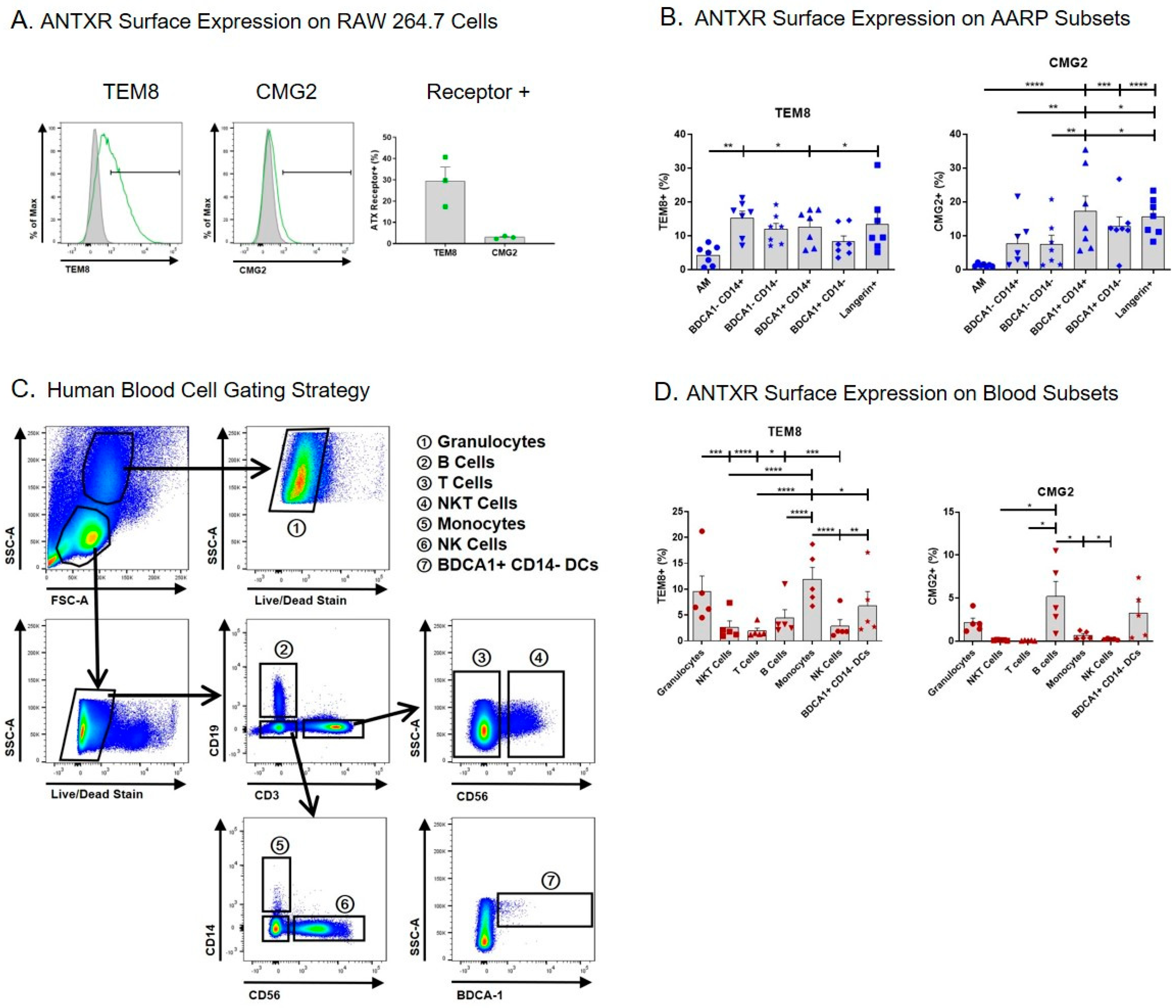{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Small Percentages of Human AARP Subsets Express Surface TEM8 and CMG2
2.2. B. anthracis PA Binds to Human AARP Subsets in a Dose-dependent but ANTXR-independent Manner
2.3. Human AARP Subsets are Resistant to LT-induced Apoptosis and Necrosis
2.4. Anthrax ET Decreases B. anthracis Spore Internalization by Human AARP Subsets, While LT Decreases Spore Internalization by Human Leukocytes
3. Discussion
4. Materials and Methods
4.1. Collection of Human Bronchoalveolar Lavage (BAL) Cells
4.2. Collection of Human Leukocytes
4.3. Flow Cytometry
4.4. Preparation of Bacillus anthracis Spores
4.5. Anthrax Receptor Surface Expression
4.6. Protective Antigen Binding Assays
4.7. Apoptosis and Necrosis Induction
4.8. Toxin Effects on B. anthracis Spore Internalization
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dixon, T.C.; Meselson, M.; Guillemin, J.; Hanna, P.C. Anthrax. N. Engl. J. Med. 1999, 341, 815–826. [Google Scholar] [CrossRef]
- Jernigan, D.B.; Raghunathan, P.L.; Bell, B.P.; Brechner, R.; Bresnitz, E.A.; Butler, J.C.; Cetron, M.; Cohen, M.; Doyle, T.; Fischer, M.; et al. Investigation of Bioterrorism-Related Anthrax, United States, 2001: Epidemiologic Findings. Emerg. Infect. Dis. 2002, 8, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Meselson, M.; Guillemin, J.; Hugh-Jones, M.; Langmuir, A.; Popova, I.; Shelokov, A.; Yampolskaya, O. The Sverdlovsk anthrax outbreak of 1979. Science 1994, 266, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Driks, A. The Bacillus anthracis spore. Mol. Asp. Med. 2009, 30, 368–373. [Google Scholar] [CrossRef]
- Ochs, M.; Nyengaard, J.R.; Jung, A.; Knudsen, L.; Voigt, M.; Wahlers, T.; Richter, J.; Gundersen, H.J.G. The Number of Alveoli in the Human Lung. Am. J. Respir. Crit. Care Med. 2004, 169, 120–124. [Google Scholar] [CrossRef]
- Ross, J.M. The pathogenesis of anthrax following the administration of spores by the respiratory route. J. Pathol. Bacteriol. 1957, 73, 485–494. [Google Scholar] [CrossRef]
- Lincoln, R.E.; Hodges, D.R.; Klein, F.; Mahlandt, B.G.; Jones, W.I.; Haines, B.W.; Rhian, M.A.; Walker, J.S. Role of the Lymphatics in the Pathogenesis of Anthrax. J. Infect. Dis. 1965, 115, 481–494. [Google Scholar] [CrossRef]
- Guidi-Rontani, C. The alveolar macrophage: The Trojan horse of Bacillus anthracis. Trends Microbiol. 2002, 10, 405–409. [Google Scholar] [CrossRef]
- Chakrabarty, K.; Wu, W.; Booth, J.L.; Duggan, E.S.; Coggeshall, K.M.; Metcalf, J. Bacillus anthracis Spores Stimulate Cytokine and Chemokine Innate Immune Responses in Human Alveolar Macrophages through Multiple Mitogen-Activated Protein Kinase Pathways. Infect. Immun. 2006, 74, 4430–4438. [Google Scholar] [CrossRef]
- Patel, V.I.; Booth, J.L.; Duggan, E.S.; Cate, S.; White, V.L.; Hutchings, D.; Kovats, S.; Burian, D.M.; Dozmorov, M.; Metcalf, J. Transcriptional Classification and Functional Characterization of Human Airway Macrophage and Dendritic Cell Subsets. J. Immunol. 2016, 198, 1183–1201. [Google Scholar] [CrossRef]
- Jakubzick, C.; Tacke, F.; Llodra, J.; van Rooijen, N.; Randolph, G.J. Modulation of Dendritic Cell Trafficking to and from the Airways. J. Immunol. 2006, 176, 3578–3584. [Google Scholar] [CrossRef]
- Kirby, A.; Coles, M.C.; Kaye, P.M. Alveolar macrophages transport pathogens to lung draining lymph nodes. J. Immunol. 2009, 183, 1983–1989. [Google Scholar] [CrossRef]
- Brittingham, K.C.; Ruthel, G.; Panchal, R.G.; Fuller, C.L.; Ribot, W.J.; Hoover, T.A.; Young, H.A.; Anderson, A.O.; Bavari, S. Dendritic cells endocytose Bacillus anthracis spores: Implications for anthrax pathogenesis. J. Immunol. 2005, 174, 5545–5552. [Google Scholar] [CrossRef] [PubMed]
- Cleret, A.; Quesnel-Hellmann, A.; Vallon-Eberhard, A.; Verrier, B.; Jung, S.; Vidal, D.; Mathieu, J.; Tournier, J.-N. Lung dendritic cells rapidly mediate anthrax spore entry through the pulmonary route. J. Immunol. 2007, 178, 7994–8001. [Google Scholar] [CrossRef] [PubMed]
- Desch, A.N.; Gibbings, S.L.; Goyal, R.; Kolde, R.; Bednarek, J.; Bruno, T.; Slansky, J.E.; Jacobelli, J.; Mason, R.; Ito, Y.; et al. Flow Cytometric Analysis of Mononuclear Phagocytes in Nondiseased Human Lung and Lung-Draining Lymph Nodes. Am. J. Respir. Crit. Care Med. 2016, 193, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.H.; Vasan, R.; Keene, D.R.; Koehler, T.M.; Xu, Y. Potential dissemination of Bacillus anthracis utilizing human lung epithelial cells. Cell. Microbiol. 2008, 10, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.H.; Liu, Q.; Jenkins, S.A.; Tuvim, M.J.; Dickey, B.; Xu, Y. In Vivo Demonstration and Quantification of Intracellular Bacillus anthracis in Lung Epithelial Cells. Infect. Immun. 2008, 76, 3975–3983. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Gu, C.; Höök, M.; Chen, X.; Rivera, J.; Pozzi, A.; Xu, Y. Entry of Bacillus anthracis spores into epithelial cells is mediated by the spore surface protein BclA, integrinα2β1 and complement component C1q. Cell. Microbiol. 2010, 13, 620–634. [Google Scholar] [CrossRef] [PubMed]
- Weiner, Z.P.; Glomski, I.J. Updating Perspectives on the Initiation of Bacillus anthracis Growth and Dissemination through Its Host. Infect. Immun. 2012, 80, 1626–1633. [Google Scholar] [CrossRef]
- Fish, D.C.; Mahlandt, B.G.; Dobbs, J.P.; Lincoln, R.E. Purification and Properties of In Vitro–produced Anthrax Toxin Components. J. Bacteriol. 1968, 95, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Leppla, S.H. Anthrax toxin edema factor: A bacterial adenylate cyclase that increases cyclic AMP concentrations of eukaryotic cells. Proc. Natl. Acad. Sci. USA 1982, 79, 3162–3166. [Google Scholar] [CrossRef]
- Duesbery, N. Proteolytic Inactivation of MAP-Kinase-Kinase by Anthrax Lethal Factor. Science 1998, 280, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Roth, I.L.; Williams, R.P. Culture of the Cytopathic Area Surrounding Virulent Cells of Bacillus Anthracis in Mouse Spleen. J. Bacteriol. 1964, 88, 523–530. [Google Scholar] [CrossRef]
- Guidi-Rontani, C.; Weber-Levy, M.; Labruyère, E.; Mock, M. Germination of Bacillus anthracis spores within alveolar macrophages. Mol. Microbiol. 1999, 31, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Gordon, V.M.; Klimpel, K.R.; Arora, N.; Henderson, M.A.; Leppla, S.H. Proteolytic activation of bacterial toxins by eukaryotic cells is performed by furin and by additional cellular proteases. Infect. Immun. 1995, 63, 82–87. [Google Scholar] [CrossRef]
- Klimpel, K.R.; Molloy, S.S.; Thomas, G.; Leppla, S.H. Anthrax toxin protective antigen is activated by a cell surface protease with the sequence specificity and catalytic properties of furin. Proc. Natl. Acad. Sci. USA 1992, 89, 10277–10281. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Klimpel, K.R.; Goel, S.; Swain, P.K.; Leppla, S.H. Oligomerization of Anthrax Toxin Protective Antigen and Binding of Lethal Factor during Endocytic Uptake into Mammalian Cells. Infect. Immun. 1999, 67, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Kintzer, A.; Sterling, H.J.; Tang, I.I.; Williams, E.R.; Krantz, B.A. Anthrax Toxin Receptor Drives Protective Antigen Oligomerization and Stabilizes the Heptameric and Octameric Oligomer by a Similar Mechanism. PLoS ONE 2010, 5, e13888. [Google Scholar] [CrossRef] [PubMed]
- Bradley, K.A.; Mogridge, J.; Mourez, M.; Collier, R.J.; Young, J.A.T. Identification of the cellular receptor for anthrax toxin. Nature 2001, 414, 225–229. [Google Scholar] [CrossRef]
- Scobie, H.M.; Rainey, G.J.A.; Bradley, K.A.; Young, J.A.T. Human capillary morphogenesis protein 2 functions as an anthrax toxin receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 5170–5174. [Google Scholar] [CrossRef]
- Liu, S.; Crown, D.; Miller-Randolph, S.; Moayeri, M.; Wang, H.; Hu, H.; Morley, T.; Leppla, S.H. Capillary morphogenesis protein-2 is the major receptor mediating lethality of anthrax toxin in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 12424–12429. [Google Scholar] [CrossRef] [PubMed]
- Martchenko, M.; Candille, S.I.; Tang, H.; Cohen, S.N. Human genetic variation altering anthrax toxin sensitivity. Proc. Natl. Acad. Sci. USA 2012, 109, 2972–2977. [Google Scholar] [CrossRef] [PubMed]
- Abrami, L.; Bischofberger, M.; Kunz, B.; Groux, R.; van der Goot, F.G. Endocytosis of the Anthrax Toxin Is Mediated by Clathrin, Actin and Unconventional Adaptors. PLoS Pathog. 2010, 6, e1000792. [Google Scholar] [CrossRef] [PubMed]
- Abrami, L.; Liu, S.; Cosson, P.; Leppla, S.H.; van der Goot, F.G. Anthrax toxin triggers endocytosis of its receptor via a lipid raft–mediated clathrin-dependent process. J. Cell Boil. 2003, 160, 321–328. [Google Scholar] [CrossRef]
- Scobie, H.M.; Marlett, J.M.; Rainey, G.J.A.; Lacy, D.B.; Collier, R.J.; Young, J.A. Anthrax Toxin Receptor 2 Determinants that Dictate the pH Threshold of Toxin Pore Formation. PLoS ONE 2007, 2, e329. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, A.M. Macrophages are sensitive to anthrax lethal toxin through an acid-dependent process. J. Boil. Chem. 1986, 261, 7123–7126. [Google Scholar]
- Popov, S.G.; Villasmil, R.; Bernardi, J.; Grene, E.; Cardwell, J.; Wu, A.; Alibek, D.; Bailey, C.; Alibek, K. Lethal toxin of Bacillus anthracis causes apoptosis of macrophages. Biochem. Biophys. Res. Commun. 2002, 293, 349–355. [Google Scholar] [CrossRef]
- Lin, C.-G.; Kao, Y.-T.; Liu, W.-T.; Huang, H.-H.; Chen, K.-C.; Wang, T.-M.; Lin, H.-C. Cytotoxic effects of anthrax lethal toxin on macrophage-like cell line J774A.1. Curr. Microbiol. 1996, 33, 224–227. [Google Scholar] [CrossRef]
- Moayeri, M.; Martinez, N.W.; Wiggins, J.; Young, H.A.; Leppla, S.H. Mouse Susceptibility to Anthrax Lethal Toxin Is Influenced by Genetic Factors in Addition to Those Controlling Macrophage Sensitivity. Infect. Immun. 2004, 72, 4439–4447. [Google Scholar] [CrossRef]
- Haines, B.W.; Klein, F.; Lincoln, R.E. Quantitative Assay for Crude Anthrax Toxins. J. Bacteriol. 1965, 89, 74–83. [Google Scholar] [CrossRef]
- Wu, W.; Mehta, H.; Chakrabarty, K.; Booth, J.L.; Duggan, E.S.; Patel, K.B.; Ballard, J.D.; Coggeshall, K.M.; Metcalf, J. Resistance of Human Alveolar Macrophages to Bacillus anthracis Lethal Toxin. J. Immunol. 2009, 183, 5799–5806. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, M.T.; Navarro, A.; Arico, C.D.; Li, J.; Alkhatib, O.; Chen, S.; Díaz-Arévalo, D.; Zeng, M. Targeted Silencing of Anthrax Toxin Receptors Protects against Anthrax Toxins. J. Boil. Chem. 2014, 289, 15730–15738. [Google Scholar] [CrossRef] [PubMed]
- Premanandan, C.; Lairmore, M.D.; Fernandez, S.; Phipps, A.J. Quantitative measurement of anthrax toxin receptor messenger RNA in primary mononuclear phagocytes. Microb. Pathog. 2006, 41, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Banks, D.J.; Barnajian, M.; Maldonado-Arocho, F.J.; Sanchez, A.M.; Bradley, K.A. Anthrax toxin receptor 2 mediates Bacillus anthracis killing of macrophages following spore challenge. Cell. Microbiol. 2005, 7, 1173–1185. [Google Scholar] [CrossRef]
- Mabry, R.; Brasky, K.; Geiger, R.; Carrion, R.; Hubbard, G.B.; Leppla, S.; Patterson, J.L.; Georgiou, G.; Iverson, B.L. Detection of Anthrax Toxin in the Serum of Animals Infected with Bacillus anthracis by Using Engineered Immunoassays. Clin. Vaccine Immunol. 2006, 13, 671–677. [Google Scholar] [CrossRef]
- Ingram, R.; Harris, A.; Ascough, S.; Metan, G.; Doganay, M.; Ballie, L.; Williamson, E.D.; Dyson, H.; Robinson, J.H.; Sriskandan, S.; et al. Exposure to anthrax toxin alters human leucocyte expression of anthrax toxin receptor 1. Clin. Exp. Immunol. 2013, 173, 84–91. [Google Scholar] [CrossRef]
- Normolle, D.; Donnenberg, V.S.; Donnenberg, A.D. Statistical classification of multivariate flow cytometry data analyzed by manual gating: Stem, progenitor, and epithelial marker expression in nonsmall cell lung cancer and normal lung. Cytom. Part A 2012, 83, 150–160. [Google Scholar] [CrossRef]
- Piasecka, J.; Thornton, C.A.; Rees, P.; Summers, H.D. Diffusion Mapping of Eosinophil-Activation State. Cytom. Part A 2019, 97, 253–258. [Google Scholar] [CrossRef]
- de la Calle, C.; Joubert, P.-E.; Law, H.K.W.; Hasan, M.; Albert, M.L. Simultaneous assessment of autophagy and apoptosis using multispectral imaging cytometry. Autophagy 2011, 7, 1045–1051. [Google Scholar] [CrossRef][Green Version]
- Alileche, A.; Serfass, E.R.; Muehlbauer, S.M.; Porcelli, S.A.; Brojatsch, J. Anthrax Lethal Toxin-Mediated Killing of Human and Murine Dendritic Cells Impairs the Adaptive Immune Response. PLoS Pathog. 2005, 1, e19. [Google Scholar] [CrossRef]
- Muehlbauer, S.M.; Evering, T.H.; Bonuccelli, G.; Squires, R.C.; Ashton, A.W.; Porcelli, S.A.; Lisanti, M.P.; Brojatsch, J. Anthrax Lethal Toxin Kills Macrophages in a Strain-Specific Manner by Apoptosis or Caspase-1-Mediated Necrosis. Cell Cycle 2007, 6, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Booth, J.L.; Duggan, E.S.; Patel, V.I.; Langer, M.; Wu, W.; Braun, A.; Coggeshall, K.M.; Metcalf, J. Bacillus anthracis spore movement does not require a carrier cell and is not affected by lethal toxin in human lung models. Microbes Infect. 2016, 18, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Yang, W.-L.; Wang, P. Measurement of Phagocytic Engulfment of Apoptotic Cells by Macrophages Using pHrodo Succinimidyl Ester. Curr. Protoc. Immunol. 2013, 100, 14.31.1–14.31.8. [Google Scholar] [CrossRef] [PubMed]
- Lenzo, J.; O’Brien-Simpson, N.M.; Cecil, J.; Holden, J.A.; Reynolds, E.C. Determination of Active Phagocytosis of Unopsonized Porphyromonas gingivalis by Macrophages and Neutrophils Using the pH-Sensitive Fluorescent Dye pHrodo. Infect. Immun. 2016, 84, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Jung, K.H.; Chai, Y.G. The Role of NF-κB and H3K27me3 Demethylase, Jmjd3, on the Anthrax Lethal Toxin Tolerance of RAW 264.7 Cells. PLoS ONE 2010, 5, e9913. [Google Scholar] [CrossRef]
- Ngundi, M.M.; Meade, B.D.; Lin, T.-L.; Tang, W.-J.; Burns, D.L. Comparison of Three Anthrax Toxin Neutralization Assays. Clin. Vaccine Immunol. 2010, 17, 895–903. [Google Scholar] [CrossRef]
- Ahn, H.C.; Kim, N.Y.; Hur, G.H.; Yang, J.-S.; Shin, S. Role of chondroitin sulfate C in the action of anthrax toxin. Toxicology 2012, 297, 10–16. [Google Scholar] [CrossRef]
- Gut, I.M.; Tamilselvam, B.; Prouty, A.M.; Stojkovic, B.; Czeschin, S.; van der Donk, W.A.; Blanke, S.R. Bacillus anthracis spore interactions with mammalian cells: Relationship between germination state and the outcome of in vitro. BMC Microbiol. 2011, 11, 46. [Google Scholar] [CrossRef]
- Zhang, L.-C.; Shao, Y.; Gao, L.; Liu, J.; Xi, Y.-Y.; Xu, Y.; Wu, C.-T.; Chen, W.; Chen, H.-P.; Wang, Y.-L.; et al. TEM8 functions as a receptor for uPA and mediates uPA-stimulated EGFR phosphorylation. Cell Commun. Signal. 2018, 16, 62. [Google Scholar] [CrossRef]
- Chaudhary, A.; Hilton, M.B.; Seaman, S.; Haines, D.C.; Stevenson, S.; Lemotte, P.K.; Tschantz, W.R.; Zhang, X.M.; Saha, S.; Fleming, T.; et al. TEM8/ANTXR1 blockade inhibits pathological angiogenesis and potentiates tumoricidal responses against multiple cancer types. Cancer Cell 2012, 21, 212–226. [Google Scholar] [CrossRef]
- Bürgi, J.; Kunz, B.; Abrami, L.; Deuquet, J.; Piersigilli, A.; Scholl-Bürgi, S.; Lausch, E.; Unger, S.; Superti-Furga, A.; Bonaldo, P.; et al. CMG2/ANTXR2 regulates extracellular collagen VI which accumulates in hyaline fibromatosis syndrome. Nat. Commun. 2017, 8, 15861. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.E.; Mavila, A.; Salazar, R.; Bayless, K.J.; Kanagala, S.; Maxwell, S.A.; Davis, G.E. Differential gene expression during capillary morphogenesis in 3D collagen matrices: Regulated expression of genes involved in basement membrane matrix assembly, cell cycle progression, cellular differentiation and G-protein signaling. J. Cell Sci. 2001, 114, 2755–2773. [Google Scholar] [PubMed]
- Nanda, A.; Carson-Walter, E.B.; Seaman, S.; Barber, T.D.; Stampfl, J.; Singh, S.; Vogelstein, B.; Kinzler, K.W.; Croix, B.S. TEM8 Interacts with the Cleaved C5 Domain of Collagen α3(VI). Cancer Res. 2004, 64, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, K.; Basile, C.; Spring, S.; Bonuccelli, G.; Lisanti, M.; Terman, B.I. TEM8 expression stimulates endothelial cell adhesion and migration by regulating cell? Matrix interactions on collagen. Exp. Cell Res. 2005, 305, 133–144. [Google Scholar] [CrossRef]
- Werner, E.; Kowalczyk, A.P.; Faundez, V. Anthrax toxin receptor 1/tumor endothelium marker 8 mediates cell spreading by coupling extracellular ligands to the actin cytoskeleton. J. Biol. Chem. 2006, 281, 23227–23236. [Google Scholar] [CrossRef] [PubMed]
- Martchenko, M.; Jeong, S.Y.; Cohen, S.N. Heterodimeric integrin complexes containing beta1-integrin promote internalization and lethality of anthrax toxin. Proc. Natl. Acad. Sci. USA 2010, 107, 15583–15588. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Hoover, B.; Leppla, S.H. The Receptors that Mediate the Direct Lethality of Anthrax Toxin. Toxins 2012, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Friedlander, A.; Dreier, T.; Ezzell, J.; Leppla, S. Effects of anthrax toxin components on human neutrophils. Infect. Immun. 1985, 47, 306–310. [Google Scholar] [CrossRef]
- Liu, S.; Miller-Randolph, S.; Crown, D.; Moayeri, M.; Sastalla, I.; Okugawa, S.; Leppla, S.H. Anthrax Toxin Targeting of Myeloid Cells through the CMG2 Receptor Is Essential for Establishment of Bacillus anthracis Infections in Mice. Cell Host Microbe 2010, 8, 455–462. [Google Scholar] [CrossRef]
- Ethier, C.; Lacy, P.; Davoine, F. Identification of Human Eosinophils in Whole Blood by Flow Cytometry. Adv. Struct. Saf. Stud. 2014, 1178, 81–92. [Google Scholar] [CrossRef]
- Rühle, P.F.; Fietkau, R.; Gaipl, U.S.; Frey, B. Development of a Modular Assay for Detailed Immunophenotyping of Peripheral Human Whole Blood Samples by Multicolor Flow Cytometry. Int. J. Mol. Sci. 2016, 17, 1316. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-R.A.; Hotten, D.F.; Malakhau, Y.; Volker, E.; Ghio, A.J.; Noble, P.W.; Kraft, M.; Hollingsworth, J.W.; Gunn, M.D.; Tighe, R.M. Flow Cytometric Analysis of Myeloid Cells in Human Blood, Bronchoalveolar Lavage, and Lung Tissues. Am. J. Respir. Cell Mol. Boil. 2016, 54, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Arocho, F.J.; Fulcher, J.A.; Lee, B.; Bradley, K.A. Anthrax oedema toxin induces anthrax toxin receptor expression in monocyte-derived cells. Mol. Microbiol. 2006, 61, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Pentelute, B.L.; Collier, R.J.; Zhou, Z.H. Atomic structure of anthrax protective antigen pore elucidates toxin translocation. Nature 2015, 521, 545–549. [Google Scholar] [CrossRef]
- Naik, S.; Brock, S.; Akkaladevi, N.; Tally, J.; McGinn-Straub, W.; Zhang, N.; Gao, P.; Gogol, E.P.; Pentelute, B.L.; Collier, R.J.; et al. Monitoring the Kinetics of the pH-Driven Transition of the Anthrax Toxin Prepore to the Pore by Biolayer Interferometry and Surface Plasmon Resonance. Biochemistry 2013, 52, 6335–6347. [Google Scholar] [CrossRef]
- Norbury, C.C.; Hewlett, L.J.; Prescott, A.R.; Shastri, N.; Watts, C. Class I MHC presentation of exogenous soluble antigen via macropinocytosis in bone marrow macrophages. Immunity 1995, 3, 783–791. [Google Scholar] [CrossRef]
- Sallusto, F.; Cella, M.; Danieli, C.; Lanzavecchia, A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: Downregulation by cytokines and bacterial products. J. Exp. Med. 1995, 182, 389–400. [Google Scholar] [CrossRef]
- Norbury, C.C.; Chambers, B.J.; Prescott, A.R.; Ljunggren, H.-G.; Watts, C. Constitutive macropinocytosis allows TAP-dependent major histocompatibility compex class I presentation of exogenous soluble antigen by bone marrow-derived dendritic cells. Eur. J. Immunol. 1997, 27, 280–288. [Google Scholar] [CrossRef]
- Racoosin, E.L.; Swanson, J.A. Macropinosome maturation and fusion with tubular lysosomes in macrophages. J. Cell Boil. 1993, 121, 1011–1020. [Google Scholar] [CrossRef]
- van Haarst, J.M.; Hoogsteden, H.C.; de Wit, H.J.; Verhoeven, G.T.; Havenith, C.E.; Drexhage, H.A. Dendritic cells and their precursors isolated from human bronchoalveolar lavage: Immunocytologic and functional properties. Am. J. Respir. Cell Mol. Boil. 1994, 11, 344–350. [Google Scholar] [CrossRef]
- Li, F.; Yang, M.; Wang, L.; Williamson, I.; Tian, F.; Qin, M.; Shah, P.K.; Sharifi, B.G. Autofluorescence contributes to false-positive intracellular Foxp3 staining in macrophages: A lesson learned from flow cytometry. J. Immunol. Methods 2012, 386, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, J.C.; Chen, D.; Lapinski, P.E.; Turner, J.; Grigorova, I.; Swanson, J.A.; King, P.D. Macropinocytosis drives T cell growth by sustaining the activation of mTORC1. Nat. Commun. 2020, 11, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Reyes, R.; Pérez-López, A.; Sánchez-Gómez, C.; Hernández-Mote, R.R.; Castro-Eguiluz, D.; Ortiz-Navarrete, V.; Alpuche-Aranda, C.M.; Castro-Eguiluz, A.D. Salmonella infects B cells by macropinocytosis and formation of spacious phagosomes but does not induce pyroptosis in favor of its survival. Microb. Pathog. 2012, 52, 367–374. [Google Scholar] [CrossRef]
- Steinman, R.M.; Brodie, S.E.; Cohn, Z.A. Membrane flow during pinocytosis. A stereologic analysis. J. Cell Biol. 1976, 68, 665–687. [Google Scholar] [CrossRef]
- Redka, D.S.; Gütschow, M.; Grinstein, S.; Canton, J. Differential ability of proinflammatory and anti-inflammatory macrophages to perform macropinocytosis. Mol. Boil. Cell 2018, 29, 53–65. [Google Scholar] [CrossRef]
- Comer, J.E.; Galindo, C.L.; Zhang, F.; Wenglikowski, A.M.; Bush, K.L.; Garner, H.R.; Peterson, J.W.; Chopra, A.K. Murine macrophage transcriptional and functional responses to Bacillus anthracis edema toxin. Microb. Pathog. 2006, 41, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Larabee, J.L.; de Giusti, K.; Regens, J.L.; Ballard, J.D. Bacillus anthracis Edema Toxin Activates Nuclear Glycogen Synthase Kinase 3β. Infect. Immun. 2008, 76, 4895–4904. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Popov, S.G.; Villasmil, R.; Bernardi, J.; Grene, E.; Cardwell, J.; Popova, T.; Wu, A.; Alibek, D.; Bailey, C.; Alibek, K. Effect of Bacillus anthracis lethal toxin on human peripheral blood mononuclear cells. FEBS Lett. 2002, 527, 211–215. [Google Scholar] [CrossRef]
- Mangan, D.F.; Mergenhagen, S.E.; Wahl, S.M. Apoptosis in human monocytes: Possible role in chronic inflammatory diseases. J. Periodontol. 1993, 64, 461–466. [Google Scholar]
- Munn, D.H.; Beall, A.C.; Song, D.; Wrenn, R.W.; Throckmorton, D.C. Activation-induced apoptosis in human macrophages: Developmental regulation of a novel cell death pathway by macrophage colony-stimulating factor and interferon gamma. J. Exp. Med. 1995, 181, 127–136. [Google Scholar] [CrossRef]
- Crawford, M.A.; Aylott, C.V.; Bourdeau, R.W.; Bokoch, G.M. Bacillus anthracis Toxins Inhibit Human Neutrophil NADPH Oxidase Activity. J. Immunol. 2006, 176, 7557–7565. [Google Scholar] [CrossRef] [PubMed]
- Kirby, J.E. Anthrax Lethal Toxin Induces Human Endothelial Cell Apoptosis. Infect. Immun. 2004, 72, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Greten, F.R.; Wong, A.; Westrick, R.J.; Arthur, J.S.C.; Otsu, K.; Hoffmann, A.; Montminy, M.; Karin, M. Signaling Pathways and Genes that Inhibit Pathogen-Induced Macrophage Apoptosis—CREB and NF-κB as Key Regulators. Immunity 2005, 23, 319–329. [Google Scholar] [CrossRef]
- Taft, S.C.; Weiss, A.A. Toxicity of Anthrax Toxin Is Influenced by Receptor Expression. Clin. Vaccine Immunol. 2008, 15, 1330–1336. [Google Scholar] [CrossRef]
- Raymond, B.; le Duc, D.; Ravaux, L.; le Goffic, R.; Candela, T.; Raymondjean, M.; Goossens, P.L.; Touqui, L. Edema Toxin Impairs Anthracidal Phospholipase A2 Expression by Alveolar Macrophages. PLoS Pathog. 2007, 3, e187. [Google Scholar] [CrossRef] [PubMed]
- Birmelin, M.; Decker, K. Synthesis of prostanoids and cyclic nucleotides by phagocytosing rat Kupffer cells. JBIC J. Boil. Inorg. Chem. 1984, 142, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Füöp, T.; Fóris, G.; Leövey, A. Age-related changes in cAMP and cGMP levels during phagocytosis in human polymorphonuclear leukocytes. Mech. Ageing Dev. 1984, 27, 233–237. [Google Scholar] [CrossRef]
- Pryzwansky, K.; Steiner, A.; Spitznagel, J.; Kapoor, C. Compartmentalization of cyclic AMP during phagocytosis by human neutrophilic granulocytes. Science 1981, 211, 407–410. [Google Scholar] [CrossRef]
- Ballinger, M.N.; Welliver, T.; Straight, S.; Peters-Golden, M.; Swanson, J.A. Transient Increase in Cyclic AMP Localized to Macrophage Phagosomes. PLoS ONE 2010, 5, e13962. [Google Scholar] [CrossRef]
- Aronoff, D.M.; Canetti, C.; Serezani, C.H.; Luo, M.; Peters-Golden, M. Cutting Edge: Macrophage Inhibition by Cyclic AMP (cAMP): Differential Roles of Protein Kinase A and Exchange Protein Directly Activated by cAMP-1. J. Immunol. 2005, 174, 595–599. [Google Scholar] [CrossRef]
- Makranz, C.; Cohen, G.; Reichert, F.; Kodama, T.; Rotshenker, S. cAMP cascade (PKA, Epac, adenylyl cyclase, Gi, and phosphodiesterases) regulates myelin phagocytosis mediated by complement receptor-3 and scavenger receptor-AI/II in microglia and macrophages. Glia 2006, 53, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.W.; Oestergaard, K.; Myers, J.T.; Swanson, J.A. Altered membrane trafficking in activated bone marrow-derived macrophages. J. Leukoc. Boil. 2000, 68, 487–494. [Google Scholar]
- Larabee, J.L.; Maldonado-Arocho, F.J.; Pacheco, S.; France, B.; de Giusti, K.; Shakir, S.M.; Bradley, K.A.; Ballard, J.D. Glycogen Synthase Kinase 3 Activation Is Important for Anthrax Edema Toxin-Induced Dendritic Cell Maturation and Anthrax Toxin Receptor 2 Expression in Macrophages. Infect. Immun. 2011, 79, 3302–3308. [Google Scholar] [CrossRef] [PubMed]
- Shetron-Rama, L.M.; Herring-Palmer, A.C.; Huffnagle, G.B.; Hanna, P. Transport of Bacillus anthracis from the lungs to the draining lymph nodes is a rapid process facilitated by CD11c+ cells. Microb. Pathog. 2010, 49, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.K.; Lang, G.A.; Larabee, J.L.; Devera, T.S.; Aye, L.M.; Shah, H.B.; Ballard, J.D.; Lang, M.L. Bacillus anthracis lethal toxin disrupts TCR signaling in CD1d-restricted NKT cells leading to functional anergy. PLoS Pathog 2009, 5, e1000588. [Google Scholar] [CrossRef]
- Fang, H.; Xu, L.; Chen, T.Y.; Cyr, J.M.; Frucht, D.M. Anthrax Lethal Toxin Has Direct and Potent Inhibitory Effects on B Cell Proliferation and Immunoglobulin Production. J. Immunol. 2006, 176, 6155–6161. [Google Scholar] [CrossRef] [PubMed]
- During, R.L.; Li, W.; Hao, B.; Koenig, J.M.; Stephens, D.S.; Quinn, C.; Southwick, F.S. Anthrax Lethal Toxin Paralyzes Neutrophil Actin-Based Motility. J. Infect. Dis. 2005, 192, 837–845. [Google Scholar] [CrossRef]
- Chauncey, K.M.; Lopez, M.C.; Sidhu, G.; Szarowicz, S.E.; Baker, H.V.; Quinn, C.; Southwick, F.S. Bacillus anthracis’ lethal toxin induces broad transcriptional responses in human peripheral monocytes. BMC Immunol. 2012, 13, 33. [Google Scholar] [CrossRef]
- Kaplan, G. Differences in the Mode of Phagocytosis with Fc and C3 Receptors in Macrophages. Scand. J. Immunol. 1977, 6, 797–807. [Google Scholar] [CrossRef]
- Lowry, M.B.; Duchemin, A.M.; Robinson, J.M.; Anderson, C.L. Functional separation of pseudopod extension and particle internalization during Fc gamma receptor-mediated phagocytosis. J. Exp. Med. 1998, 187, 161–176. [Google Scholar] [CrossRef]
- May, R.C.; Caron, E.; Hall, A.; Machesky, L.M. Involvement of the Arp2/3 complex in phagocytosis mediated by FcgammaR or CR3. Nat. Cell Biol. 2000, 2, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Lewalle, P.; Rouas, R.; Lehmann, F.; Martiat, P. Freezing of dendritic cells, generated from cryopreserved leukaphereses, does not influence their ability to induce antigen-specific immune responses or functionally react to maturation stimuli. J. Immunol. Methods 2000, 240, 69–78. [Google Scholar] [CrossRef]
- Carr, K.A.; Lybarger, S.R.; Anderson, E.C.; Janes, B.K.; Hanna, P.C. The role of Bacillus anthracis germinant receptors in germination and virulence. Mol. Microbiol. 2010, 75, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, K.; Wu, W.; Booth, J.L.; Duggan, E.S.; Nagle, N.N.; Coggeshall, K.M.; Metcalf, J. Human Lung Innate Immune Response to Bacillus anthracis Spore Infection. Infect. Immun. 2007, 75, 3729–3738. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, V.I.; Booth, J.L.; Dozmorov, M.; Brown, B.R.; Metcalf, J.P. Anthrax Edema and Lethal Toxins Differentially Target Human Lung and Blood Phagocytes. Toxins 2020, 12, 464. https://doi.org/10.3390/toxins12070464
Patel VI, Booth JL, Dozmorov M, Brown BR, Metcalf JP. Anthrax Edema and Lethal Toxins Differentially Target Human Lung and Blood Phagocytes. Toxins. 2020; 12(7):464. https://doi.org/10.3390/toxins12070464
Chicago/Turabian StylePatel, Vineet I., J. Leland Booth, Mikhail Dozmorov, Brent R. Brown, and Jordan P. Metcalf. 2020. "Anthrax Edema and Lethal Toxins Differentially Target Human Lung and Blood Phagocytes" Toxins 12, no. 7: 464. https://doi.org/10.3390/toxins12070464
APA StylePatel, V. I., Booth, J. L., Dozmorov, M., Brown, B. R., & Metcalf, J. P. (2020). Anthrax Edema and Lethal Toxins Differentially Target Human Lung and Blood Phagocytes. Toxins, 12(7), 464. https://doi.org/10.3390/toxins12070464