The Toxicological Intersection between Allergen and Toxin: A Structural Comparison of the Cat Dander Allergenic Protein Fel d1 and the Slow Loris Brachial Gland Secretion Protein

 , , ,
, , , _Kwok.png) and
and 
Abstract
1. Introduction
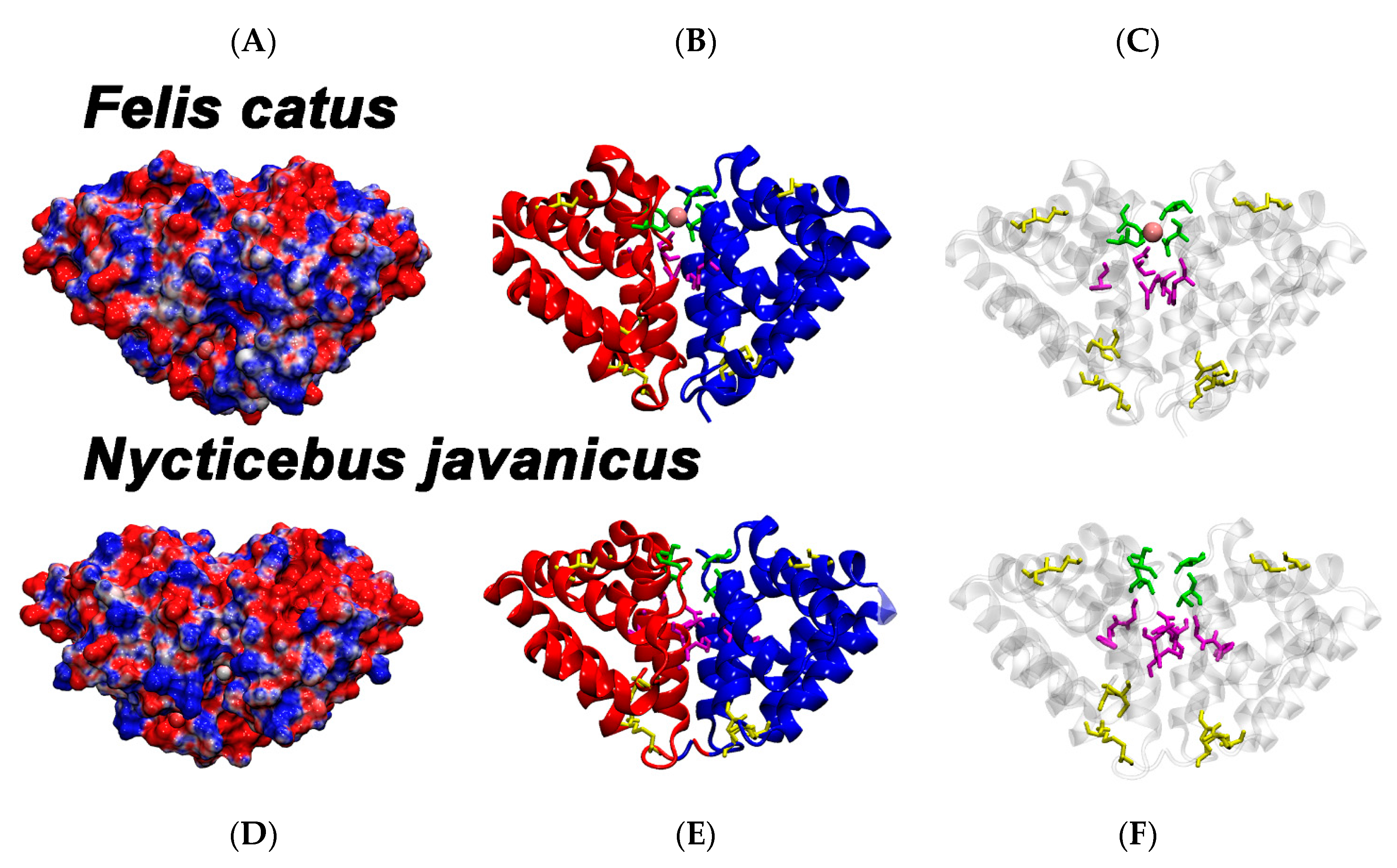
2. Results and Discussion
3. Materials and Methods
3.1. Brachial Gland Secretion
3.2. MS/MS
3.3. LC-MS/MS Analysis
3.4. mRNA
3.5. Modeling
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Petras, D.; Card, D.C.; Suranse, V.; Mychajliw, A.M.; Richards, D.; Koludarov, I.; Albulescu, L.O.; Slagboom, J.; Hempel, B.F.; et al. Solenodon genome reveals convergent evolution of venom in eulipotyphlan mammals. Proc. Nat. Acad. Sci. USA 2019, 116, 25745–25755. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Kakumanu, R.; Hodgson, W.C.; Ravi, R.; Alagon, A.; Harris, R.J.; Brust, A.; Alewood, P.F.; Kemp-Harper, B.K.; Fry, B.G. Vampire Venom: Vasodilatory Mechanisms of Vampire Bat (Desmodus rotundus) Blood Feeding. Toxins 2019, 11. [Google Scholar] [CrossRef]
- Ligabue-Braun, R.; Verli, H.; Carlini, C.R. Venomous mammals: A review. Toxicon 2012, 59, 680–695. [Google Scholar] [CrossRef]
- Low, D.H.; Sunagar, K.; Undheim, E.A.; Ali, S.A.; Alagon, A.C.; Ruder, T.; Jackson, T.N.; Pineda Gonzalez, S.; King, G.F.; Jones, A.; et al. Dracula’s children: Molecular evolution of vampire bat venom. J. Proteom. 2013, 89, 95–111. [Google Scholar] [CrossRef]
- Rode-Margono, J.E.; Nekaris, K.A. Cabinet of Curiosities: Venom Systems and Their Ecological Function in Mammals, with a Focus on Primates. Toxins 2015, 7, 2639–2658. [Google Scholar] [CrossRef]
- Alterman, L. Toxins and toothcombs: Potential allospecific chemical defenses in Nycticebus and Perodicticus. In Creatures of the Dark: The Nocturnal Prosimians; Alterman, L., Doyle, G.A., Izard, M.K., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 413–424. [Google Scholar]
- Nekaris, K.A.; Moore, R.S.; Rode, E.J.; Fry, B.G. Mad, bad and dangerous to know: The biochemistry, ecology and evolution of slow loris venom. J. Venom. Anim. Toxins Incl. Trop Dis. 2013, 19, 21. [Google Scholar] [CrossRef]
- Fuller, G.; Lukas, K.E.; Kuhar, C.; Dennis, P.M. A retrospective review of mortality in lorises and pottos in North American zoos 1980–2010. Endanger. Species Res. 2014, 23, 205–217. [Google Scholar] [CrossRef]
- Streicher, U. Aspects of Ecology and Conservation of the Pygmy Loris Nycticebus Pygmaeus in Vietnam; Ludwig-Maximilian-Universitat Munchen: Munich, Germany, 2004. [Google Scholar]
- Fung, H.T.; Wong, O.F. Clinical quiz: A potentially toxic primate bite. Hong Kong J. Emerg. Med. 2016, 23, 301–303. [Google Scholar] [CrossRef]
- Gardiner, M.; Weldon, A.; Poindexter, S.A.; Gibson, N.; Nekaris, K.A.I. Survey of practitioners handling slow lorises (Primates: Nycticebus): An assessment of the harmful effects of slow loris bites. J. Venom. Res. 2018, 9, 1–7. [Google Scholar] [PubMed]
- Madani, G.; Nekaris, K.A. Anaphylactic shock following the bite of a wild Kayan slow loris (Nycticebus kayan): Implications for slow loris conservation. J. Venom. Anim. Toxins Incl. Trop Dis. 2014, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- Hagey, L.; Fry, B.G.; Snyder, H. Talking Defensively: A Dual Use for the Brachial Gland Exudate of Slow and Pygmy Lorises. In Developments in Primatology; Gursky, N., Ed.; Kluwer: South Holland, The Netherlands, 2007. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.C.; Baer, H.; Ohman, J.L., Jr. A comparative study of the allergens of cat urine, serum, saliva, and pelt. J. Allergy Clin. Immunol. 1985, 76, 563–569. [Google Scholar] [CrossRef]
- Charpin, C.; Mata, P.; Charpin, D.; Lavaut, M.N.; Allasia, C.; Vervloet, D. Fel d I allergen distribution in cat fur and skin. J. Allergy Clin. Immunol. 1991, 88, 77–82. [Google Scholar] [CrossRef]
- Klug, J.; Beier, H.M.; Bernard, A.; Chilton, B.S.; Fleming, T.P.; Lehrer, R.I.; Miele, L.; Pattabiraman, N.; Singh, G. Uteroglobin/Clara cell 10-kDa family of proteins: Nomenclature committee report. Ann. N. Y. Acad. Sci. 2000, 923, 348–354. [Google Scholar] [CrossRef]
- van Milligen, F.J.; Vroom, T.M.; Aalberse, R.C. Presence of Felis domesticus allergen I in the cat’s salivary and lacrimal glands. Int. Arch. Allergy Appl. Immunol. 1990, 92, 375–378. [Google Scholar] [CrossRef]
- Duffort, O.A.; Carreira, J.; Nitti, G.; Polo, F.; Lombardero, M. Studies on the biochemical structure of the major cat allergen Felis domesticus I. Mol. Immunol. 1991, 28, 301–309. [Google Scholar] [CrossRef]
- Kristensen, A.K.; Schou, C.; Roepstorff, P. Determination of isoforms, N-linked glycan structure and disulfide bond linkages of the major cat allergen Fel d1 by a mass spectrometric approach. Biol. Chem. 1997, 378, 899–908. [Google Scholar] [CrossRef]
- Griffith, I.J.; Craig, S.; Pollock, J.; Yu, X.B.; Morgenstern, J.P.; Rogers, B.L. Expression and genomic structure of the genes encoding FdI, the major allergen from the domestic cat. Gene 1992, 113, 263–268. [Google Scholar] [CrossRef]
- Kaiser, L.; Gronlund, H.; Sandalova, T.; Ljunggren, H.G.; van Hage-Hamsten, M.; Achour, A.; Schneider, G. The crystal structure of the major cat allergen Fel d 1, a member of the secretoglobin family. J. Biol. Chem. 2003, 278, 37730–37735. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, L.; Velickovic, T.C.; Badia-Martinez, D.; Adedoyin, J.; Thunberg, S.; Hallen, D.; Berndt, K.; Gronlund, H.; Gafvelin, G.; van Hage, M.; et al. Structural characterization of the tetrameric form of the major cat allergen Fel d 1. J. Mol. Biol. 2007, 370, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Gronlund, H.; Bergman, T.; Sandstrom, K.; Alvelius, G.; Reininger, R.; Verdino, P.; Hauswirth, A.; Liderot, K.; Valent, P.; Spitzauer, S.; et al. Formation of disulfide bonds and homodimers of the major cat allergen Fel d 1 equivalent to the natural allergen by expression in Escherichia coli. J. Biol. Chem. 2003, 278, 40144–40151. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.K.; Leung, D.Y.M. Dog and Cat Allergies: Current State of Diagnostic Approaches and Challenges. Allergy Asthma Immunol. Res. 2018, 10, 97–105. [Google Scholar] [CrossRef]
- Smith, W.; O’Neil, S.E.; Hales, B.J.; Chai, T.L.; Hazell, L.A.; Tanyaratsrisakul, S.; Piboonpocanum, S.; Thomas, W.R. Two newly identified cat allergens: The von Ebner gland protein Fel d 7 and the latherin-like protein Fel d 8. Int. Arch. Allergy Immunol. 2011, 156, 159–170. [Google Scholar] [CrossRef]
- Smith, W.; Butler, A.J.; Hazell, L.A.; Chapman, M.D.; Pomes, A.; Nickels, D.G.; Thomas, W.R. Fel d 4, a cat lipocalin allergen. Clin. Exp. Allergy 2004, 34, 1732–1738. [Google Scholar] [CrossRef]
- Ohman, J.L.; Lowell, F.C.; Bloch, K.J. Allergens of mammalian origin: Characterization of allergen extracted from cat pelts. J. Allergy Clin. Immunol. 1973, 52, 231–241. [Google Scholar] [CrossRef]
- van Ree, R.; van Leeuwen, W.A.; Bulder, I.; Bond, J.; Aalberse, R.C. Purified natural and recombinant Fel d 1 and cat albumin in in vitro diagnostics for cat allergy. J. Allergy Clin. Immunol. 1999, 104, 1223–1230. [Google Scholar] [CrossRef]
- Custovic, A.; Simpson, A.; Pahdi, H.; Green, R.M.; Chapman, M.D.; Woodcock, A. Distribution, aerodynamic characteristics, and removal of the major cat allergen Fel d 1 in British homes. Thorax 1998, 53, 33–38. [Google Scholar] [CrossRef]
- Jalil-Colome, J.; de Andrade, A.D.; Birnbaum, J.; Casanova, D.; Mege, J.L.; Lanteaume, A.; Charpin, D.; Vervloet, D. Sex difference in Fel d 1 allergen production. J. Allergy Clin. Immunol. 1996, 98, 165–168. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Morgenstern, J.P.; Griffith, I.J.; Brauer, A.W.; Rogers, B.L.; Bond, J.F.; Chapman, M.D.; Kuo, M.C. Amino acid sequence of Fel dI, the major allergen of the domestic cat: Protein sequence analysis and cDNA cloning. Proc. Nat. Acad. Sci. USA 1991, 88, 9690–9694. [Google Scholar] [CrossRef] [PubMed]
- Barnes, H.J.; Nordlund-Moller, L.; Nord, M.; Gustafsson, J.; Lund, J.; Gillner, M. Structural basis for calcium binding by uteroglobins. J. Mol. Biol. 1996, 256, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Beato, M. Binding of steroids to uteroglobin. J. Steroid Biochem 1976, 7, 327–334. [Google Scholar] [CrossRef]
- Emes, R.D.; Riley, M.C.; Laukaitis, C.M.; Goodstadt, L.; Karn, R.C.; Ponting, C.P. Comparative evolutionary genomics of androgen-binding protein genes. Genome Res. 2004, 14, 1516–1529. [Google Scholar] [CrossRef]
- Hard, T.; Barnes, H.J.; Larsson, C.; Gustafsson, J.A.; Lund, J. Solution structure of a mammalian PCB-binding protein in complex with a PCB. Nat. Struct. Biol. 1995, 2, 983–989. [Google Scholar] [CrossRef]
- Lopez de Haro, M.S.; Perez Martinez, M.; Garcia, C.; Nieto, A. Binding of retinoids to uteroglobin. FEBS Lett. 1994, 349, 249–251. [Google Scholar] [CrossRef]
- Negi, S.S.; Schein, C.H.; Oezguen, N.; Power, T.D.; Braun, W. InterProSurf: A web server for predicting interacting sites on protein surfaces. Bioinformatics 2007, 23, 3397–3399. [Google Scholar] [CrossRef]
- Gould, H.J.; Sutton, B.J.; Beavil, A.J.; Beavil, R.L.; McCloskey, N.; Coker, H.A.; Fear, D.; Smurthwaite, L. The biology of IGE and the basis of allergic disease. Annu. Rev. Immunol. 2003, 21, 579–628. [Google Scholar] [CrossRef]
- Kleine-Tebbe, J.; Kleine-Tebbe, A.; Jeep, S.; Schou, C.; Lowenstein, H.; Kunkel, G. Role of the major allergen (Fel d I) in patients sensitized to cat allergens. Int. Arch. Allergy Immunol. 1993, 100, 256–262. [Google Scholar] [CrossRef]
- Satyaraj, E.; Gardner, C.; Filipi, I.; Cramer, K.; Sherrill, S. Reduction of active Fel d1 from cats using an antiFel d1 egg IgY antibody. Immun. Inflamm. Dis. 2019, 7, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Uermosi, C.; Zabel, F.; Manolova, V.; Bauer, M.; Beerli, R.R.; Senti, G.; Kundig, T.M.; Saudan, P.; Bachmann, M.F. IgG-mediated down-regulation of IgE bound to mast cells: A potential novel mechanism of allergen-specific desensitization. Allergy 2014, 69, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Kepley, C.L.; Zhang, K.; Terada, T.; Yamada, T.; Saxon, A. A chimeric human-cat fusion protein blocks cat-induced allergy. Nat. Med. 2005, 11, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, S.A.; Nekaris, K.A.I. Vertical clingers and gougers: Rapid acquisition of adult limb proportions facilitates feeding behaviours in young Javan slow lorises (Nycticebus javanicus). Mammalian Biology-Zeitschrift für Säugetierkunde 2017, 87, 40–49. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30 (Suppl. 1), S162–S173. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput Chem 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Negri, L.; Lattanzi, R.; Giannini, E.; Melchiorri, P. Bv8/Prokineticin proteins and their receptors. Life Sci. 2007, 81, 1103–1116. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Nat. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Enzyme Digest | Initial de novo Sequence Returned by Peaks Studio | ALC (%) | m/z | z | Final Correct Sequence |
|---|---|---|---|---|---|
| Trypsin | N(+42.01) TLFYGVFGALVTGDK | 96 | 872.4631 | 2 | CPIFYGVFGAIVTGDK |
| Trypsin | NLLDSFLDKVSGTEPEK | 96 | 631.3182 | 2 | NIIDSFIDKVSGTEPEK |
| Trypsin | T(+42.01)TC(+71.06)KLQM(+15.99)AAFNEEGLTGK | 92 | 762.8611 | 2 | IQECFNEEGITGK |
| Trypsin | LTEEDQDNVQSGLDK | 97 | 564.2603 | 3 | LTEEDKDNVQSGLDK |
| Trypsin | Edman degradation | VKSSAVLENAK | |||
| Trypsin | Edman degradation | IYSSNFCPIFYGVFGAIVTGDK | |||
| AspN | DNKLTEEDKN(+.98)NVQSGL | 94 | 902.9345 | 2 | DNKLTEEDKDNVQSGL |
| LysC | LTEEDKDNVQSGLDK | 98 | 845.91 | 2 | LTEEDKDNVQSGLDK |
| LysC | AAFENLKEC (+57.02) FNEEGLTGK | 96 | 1028.9913 | 2 | AAFENIQECFNEEGITGK |
| Chain | Polar Area/Energy | Apolar Area/Energy | Total Area/Energy | Surface Atoms | Buried Atoms |
|---|---|---|---|---|---|
| Domain A | 2602.53 | 4497.99 | 7100.53 | 609 | 439 |
| Domain B | 2602.62 | 4497.99 | 7100.61 | 609 | 439 |
| Homodimer | 4730.58 | 7466.48 | 12197.06 | 1170 | 926 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheib, H.; Nekaris, K.A.-I.; Rode-Margono, J.; Ragnarsson, L.; Baumann, K.; Dobson, J.S.; Wirdateti, W.; Nouwens, A.; Nijman, V.; Martelli, P.; et al. The Toxicological Intersection between Allergen and Toxin: A Structural Comparison of the Cat Dander Allergenic Protein Fel d1 and the Slow Loris Brachial Gland Secretion Protein. Toxins 2020, 12, 86. https://doi.org/10.3390/toxins12020086
Scheib H, Nekaris KA-I, Rode-Margono J, Ragnarsson L, Baumann K, Dobson JS, Wirdateti W, Nouwens A, Nijman V, Martelli P, et al. The Toxicological Intersection between Allergen and Toxin: A Structural Comparison of the Cat Dander Allergenic Protein Fel d1 and the Slow Loris Brachial Gland Secretion Protein. Toxins. 2020; 12(2):86. https://doi.org/10.3390/toxins12020086
Chicago/Turabian StyleScheib, Holger, K. Anne-Isola Nekaris, Johanna Rode-Margono, Lotten Ragnarsson, Kate Baumann, James S. Dobson, Wirdateti Wirdateti, Amanda Nouwens, Vincent Nijman, Paolo Martelli, and et al. 2020. "The Toxicological Intersection between Allergen and Toxin: A Structural Comparison of the Cat Dander Allergenic Protein Fel d1 and the Slow Loris Brachial Gland Secretion Protein" Toxins 12, no. 2: 86. https://doi.org/10.3390/toxins12020086
APA StyleScheib, H., Nekaris, K. A.-I., Rode-Margono, J., Ragnarsson, L., Baumann, K., Dobson, J. S., Wirdateti, W., Nouwens, A., Nijman, V., Martelli, P., Ma, R., Lewis, R. J., Kwok, H. F., & Fry, B. G. (2020). The Toxicological Intersection between Allergen and Toxin: A Structural Comparison of the Cat Dander Allergenic Protein Fel d1 and the Slow Loris Brachial Gland Secretion Protein. Toxins, 12(2), 86. https://doi.org/10.3390/toxins12020086