Development of an Antifungal and Antimycotoxigenic Device Containing Allyl Isothiocyanate for Silo Fumigation

 , , ,
, , , 
Abstract
1. Introduction
2. Results
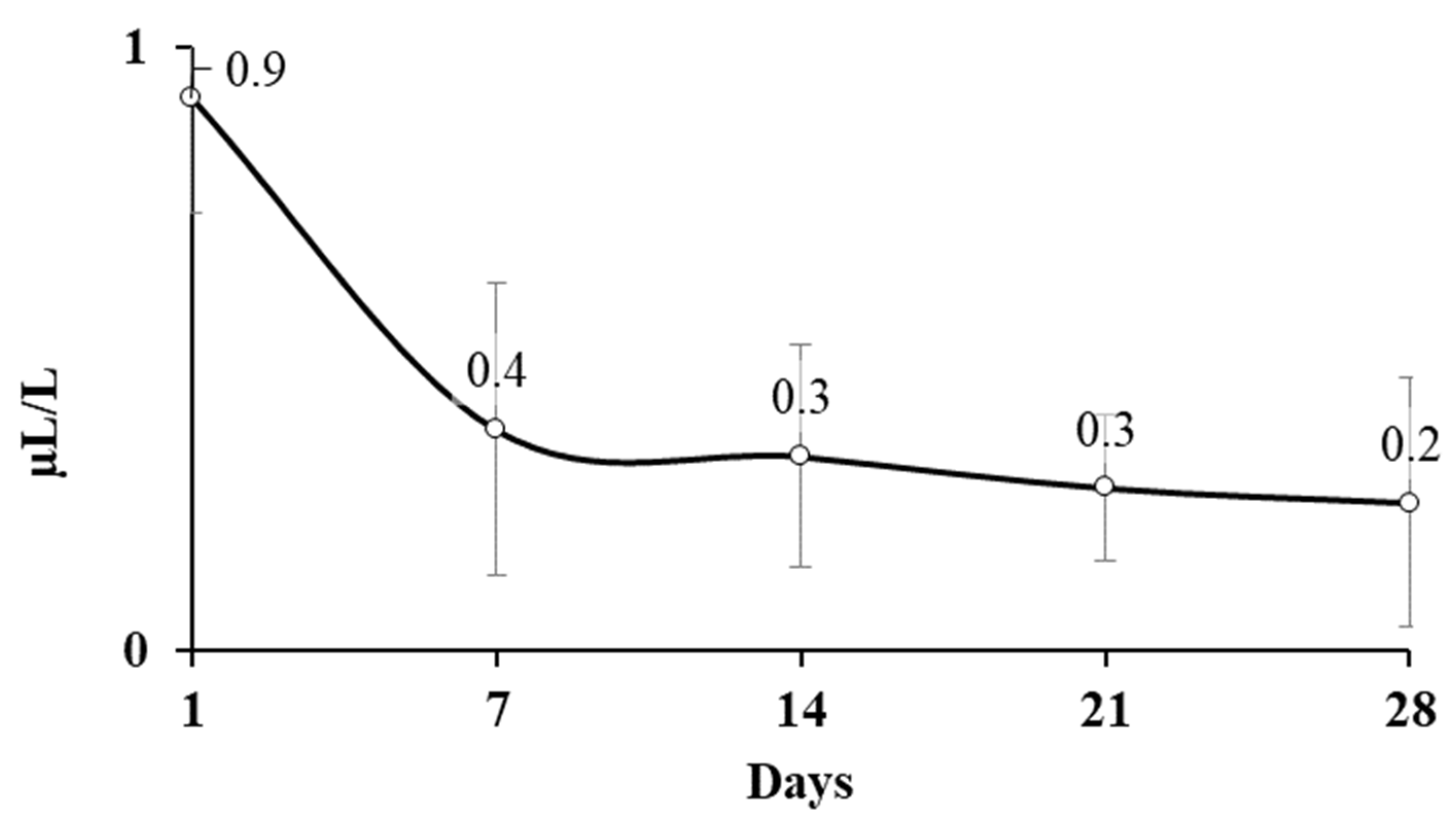
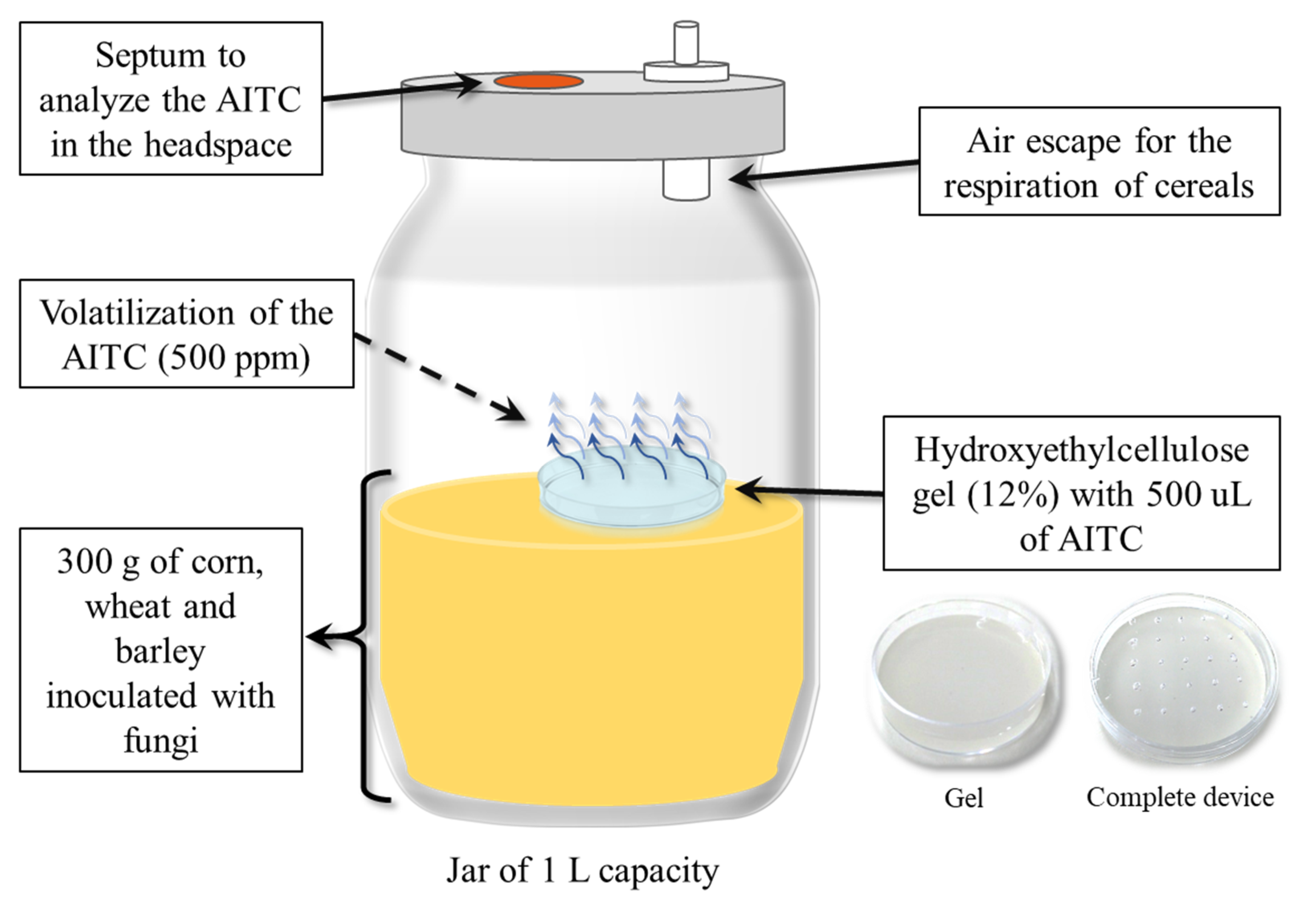
2.1. AITC Concentration in Headspace and Cereals in Laboratory Scale Silo
2.2. Validation Method for the Analysis of Mycotoxins in Cereals
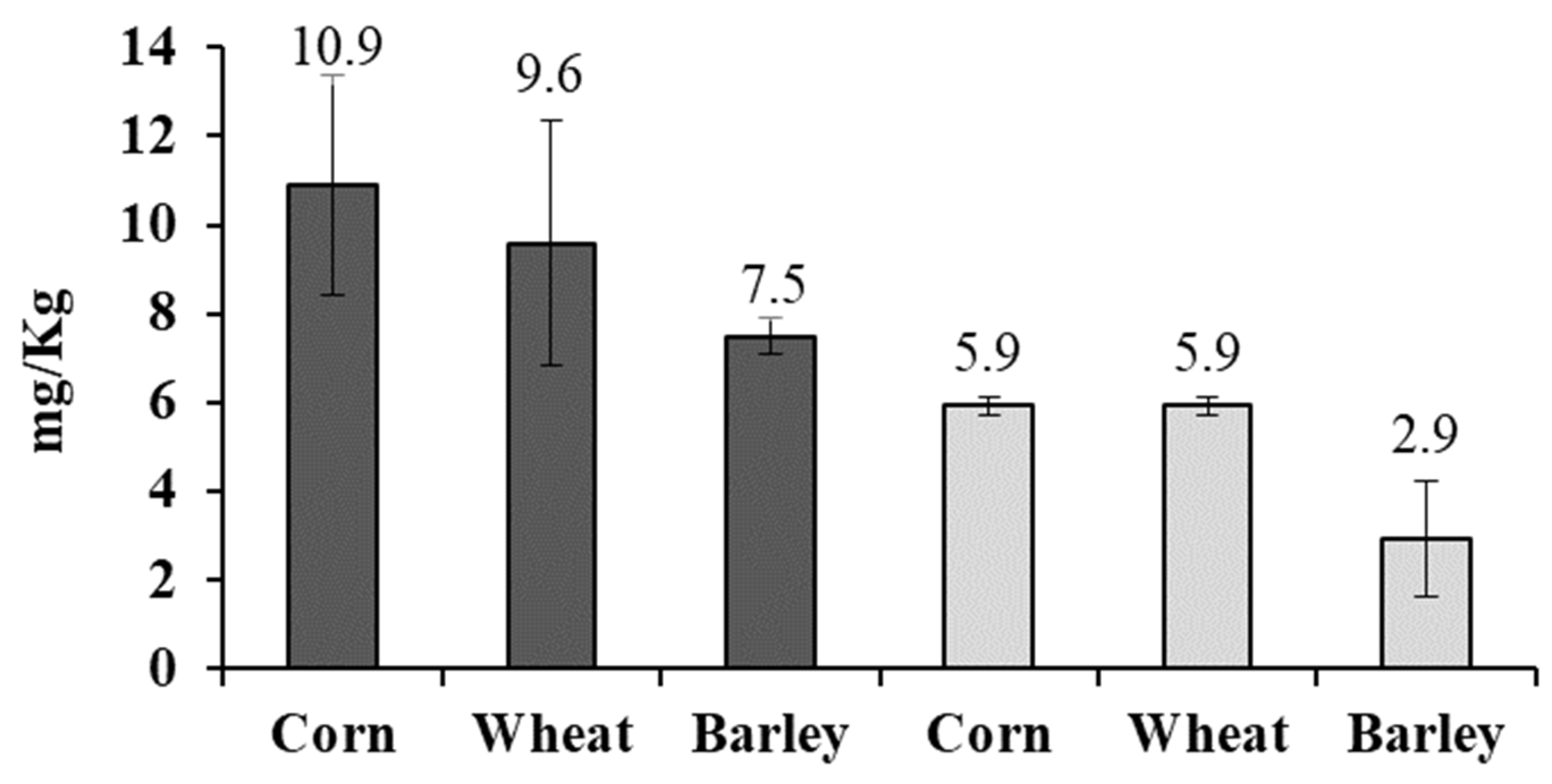
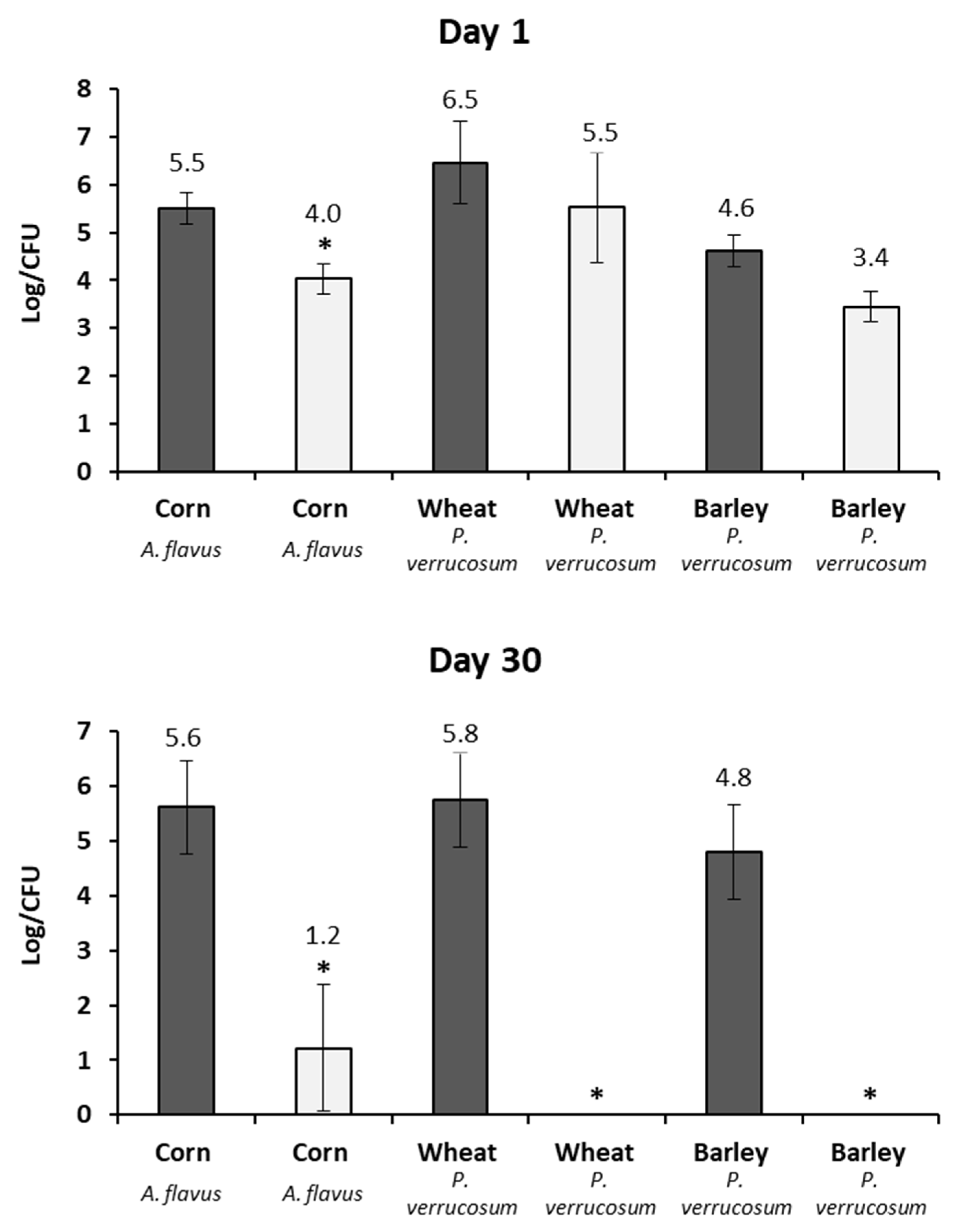
2.3. Fungal Growth and Mycotoxin Production in Lab Scale Silo System
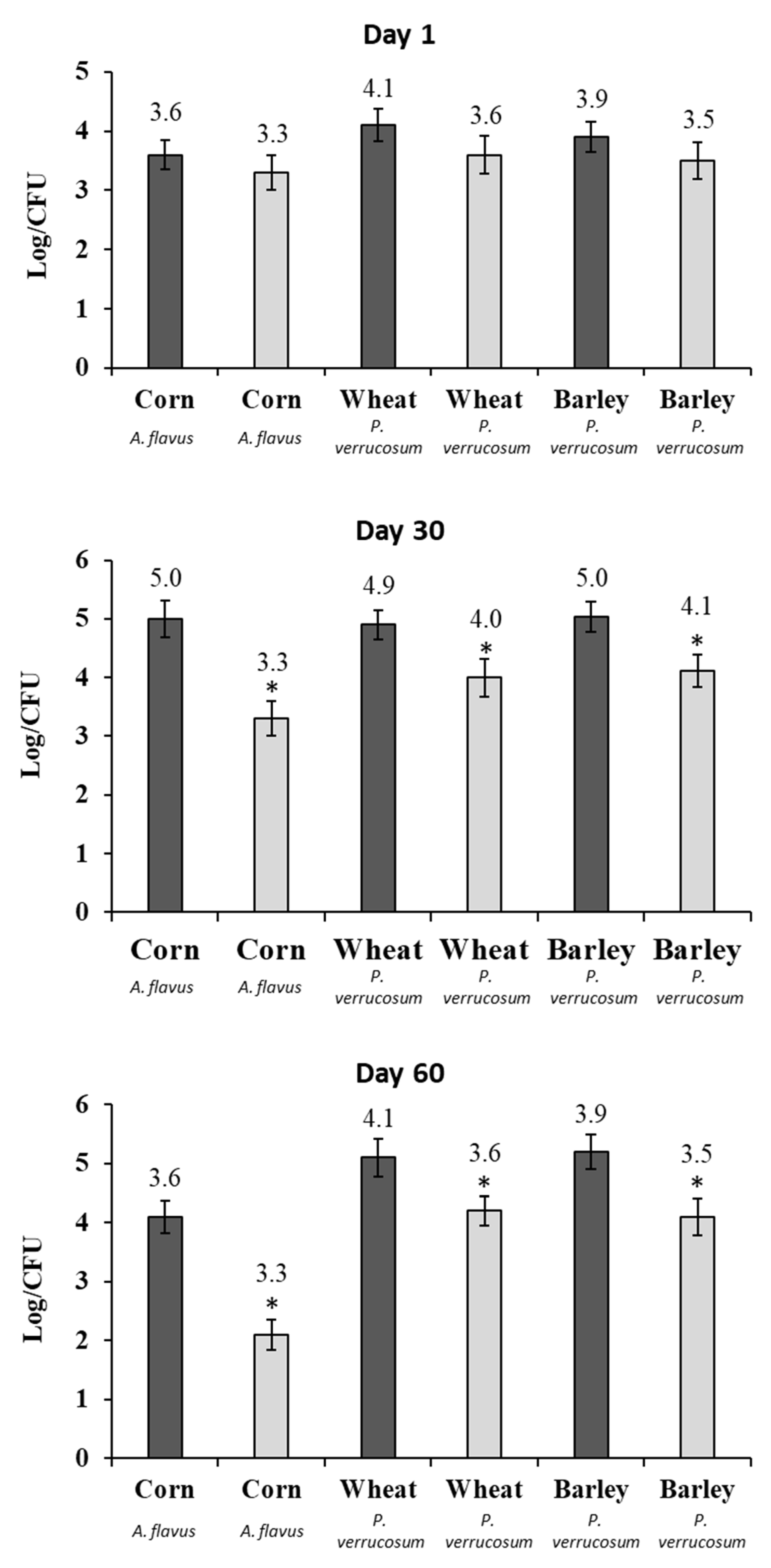
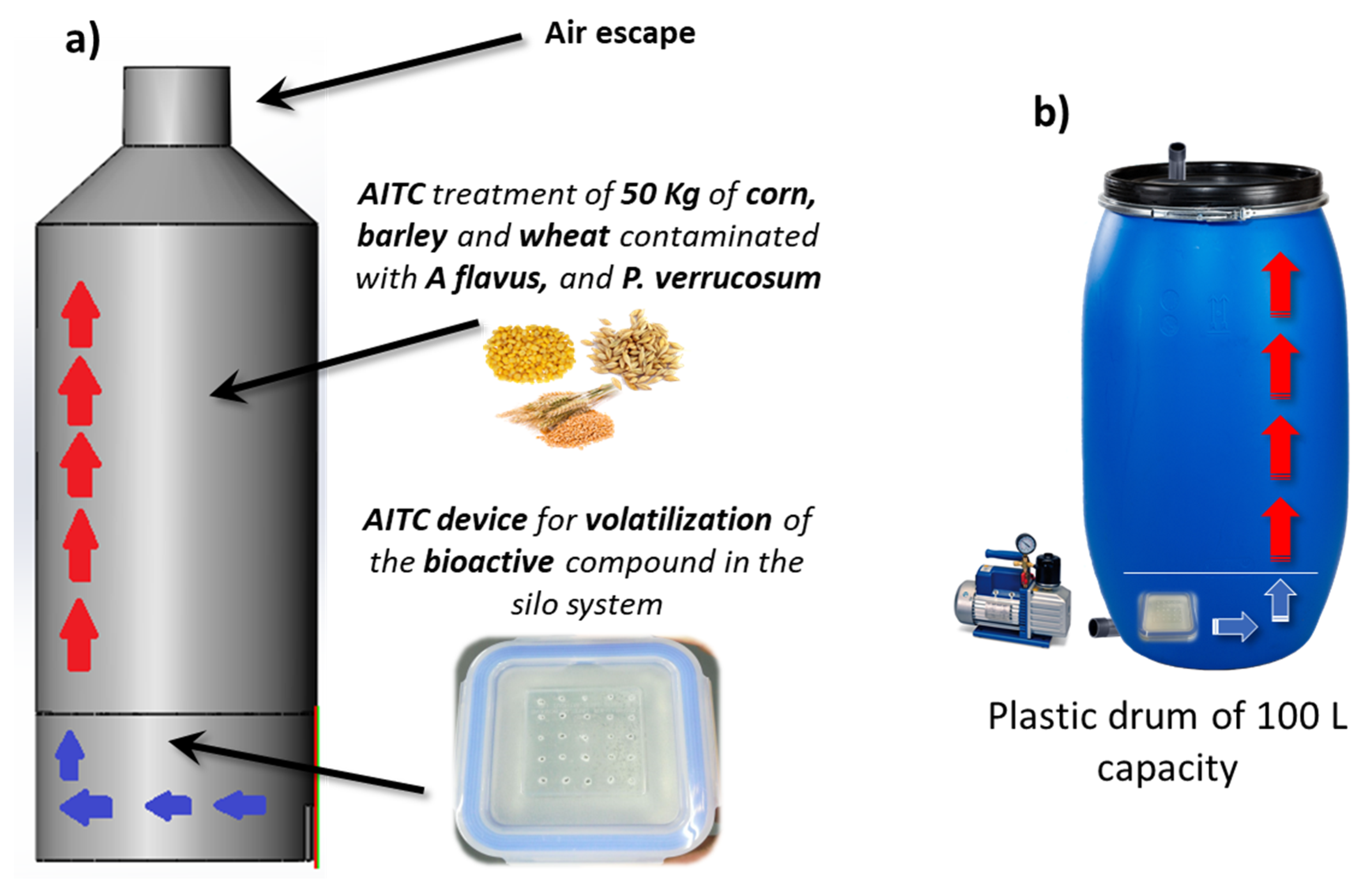
2.4. Fungal Growth and Mycotoxin Production in a Small-Scale Silo System
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Chemicals and Microbial Strains
5.2. Laboratory Scale Silo System and Antifungal Treatment with the AITC Device
5.3. AITC Device Application in a Small-Scale Silo System
5.4. Determination of AITC Concentration in the Headspace of the Laboratory Scale Silo
5.5. Determination of AITC Concentration in the Cereals of the Laboratory Scale Silo
5.6. Mycotoxin Extraction and LC-MS/MS Analysis of Corn, Barley, and Wheat
5.7. Determination of the Fungal Population
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Majeed, S.; Iqbal, M.; Asi, M.R.; Iqbal, S.Z. Aflatoxins and ochratoxin A contamination in rice, corn and corn products from Punjab, Pakistan. J. Cereal Sci. 2013, 58, 446–450. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Asi, M.R.; Ariño, A.; Akram, N.; Zuber, M. Aflatoxin contamination in different fractions of rice from Pakistan and estimation of dietary intakes. Mycotoxin Res. 2012, 28, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Pittet, A. Natural occurrence of mycotoxins in foods and feeds—An updated review. Rev. Med. Vet. 1998, 149, 479–492. [Google Scholar]
- IARC-International Agency for Research on Cancer. Monographs on the evaluation of carcinogenic risks to humans. In A Review of Biological Agents for Human Carcinogens; IARC-International Agency for Research on Cancer: Lyon, France, 2012. [Google Scholar]
- Iqbal, S.Z.; Asi, M.R.; Ariño, A. Aflatoxins. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Science Direct: London, UK, 2013; pp. 43–47. [Google Scholar]
- Hammami, W.; Fiori, S.; Al Thani, R.; Ali Kali, N.; Balmas, V.; Migheli, Q.; Jaoua, S. Fungal and aflatoxin contamination of marketed spices. Food Control 2014, 37, 177–181. [Google Scholar] [CrossRef]
- Blesa, J.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Limited survey for the presence of aflatoxins in foods from local markets and supermarkets in Valencia, Spain. Food Addit. Contam. 2004, 21, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Van de Perre, E.; Jacxsens, L.; Lachat, C.; El Tahan, F.; De Meulenaer, B. Impact of maximum levels in European legislation on exposure of mycotoxins in dried products: Case of aflatoxin B1 and ochratoxin A in nuts and dried fruits. Food Chem. Toxicol. 2015, 75, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins. II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus Wilh. J. Chem. Soc. Perkin 1965, 7083–7088. [Google Scholar] [CrossRef]
- Téren, J.; Varga, J.; Hamari, Z.; Rinyu, E.; Kevei, F. Immunochemical detection of ochratoxin A in black Aspergillus strains. Mycopathologia 1996, 134, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Abarca, M.L.; Bragulat, M.R.; Castellá, G.; Cabañes, F.J. Impact of some environmental factors on growth and ochratoxin A production by Aspergillus niger and Aspergillus welwitschiae. Int. J. Food Microbiol. 2018, 291, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Bode, H.; Raupp, F.; Geisen, R. Influence of light on ochratoxin biosynthesis by Penicillium. Mycotoxin Res. 2010, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.; Bresch, H.; Cholmakov-Bodechtel, C.; Engel, G.; Garais, M.; Majerus, P.; Rosner, H.; Scheuer, R. Ochratoxin A: Contamination of foods and consumer exposure. Arch. Lebensmittelhyg. 2000, 51, 81–128. [Google Scholar]
- Petzinger, E.; Weidenbach, A. Mycotoxins in the food chain: The role of ochratoxins. Livestock Prod. Sci. 2002, 76, 245–250. [Google Scholar] [CrossRef]
- Rizzo, A.; Eskola, M.; Atroshi, F. Ochratoxin a in Cereals, Foodstuffs and Human Plasma. Eur. J. Plant Pathol. 2002, 108, 631–637. [Google Scholar] [CrossRef]
- Denli, M.; Perez, J.F. Ochratoxins in feed, a risk for animal and human health: Control strategies. Toxins 2010, 2, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.; Bhoola, K. Ochratoxins-food contaminants: Impact on human health. Toxins 2010, 2, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.S.; Katta, S.K.; Fingerhut, D.D.; DeVries, J.W.; Bullerman, L.B. Effects of baking and frying on the fumonisin B1 content of corn-based foods. J. Agric. Food Chem. 1997, 45, 4800–4805. [Google Scholar] [CrossRef]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar] [CrossRef]
- Engel, E.; Baty, C.; Le Corre, D.; Souchon, I.; Martin, N. Flavor-active compounds potentially implicated in cooked cauliflower acceptance. J. Agric. Food Chem. 2002, 50, 6459–6467. [Google Scholar] [CrossRef] [PubMed]
- Luciano, F.B.; Holley, R.A. Enzymatic inhibition by allyl isothiocyanate and factors affecting its antimicrobial action against Escherichia coli O157:H7. Int. J. Food Microbiol. 2009, 131, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Isshiki, K.; Tokuoka, K.; Mori, R.; Chiba, S. Preliminary examination of allyl-isothiocyanate vapor for food preservation. Biosci. Biotechnol. Biochem. 1992, 56, 1476–1477. [Google Scholar] [CrossRef]
- Manyes, L.; Luciano, F.B.; Mañes, J.; Meca, G. In vitro antifungal activity of allyl isothiocyanate (AITC) against Aspergillus parasiticus and Penicillium expansum and evaluation of the AITC estimated daily intake. Food Chem. Toxicol. 2015, 83, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, T.M.; Bordin, K.; Manyes, L.; Meca, G.; Mañes, J.; Luciano, F.B. Gaseous allyl isothiocyanate to inhibit the production of aflatoxins, beauvericin and enniatins by Aspergillus parasiticus and Fusarium poae in wheat flour. Food Control 2016, 62, 317–321. [Google Scholar] [CrossRef]
- Zhang, Y. Cancer-preventive isothiocyanates: Measurement of human exposure and mechanism of action. Mutat. Res. 2004, 555, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Cejpek, K.; Valusek, J.; Velísek, J. Reactions of allyl isothiocyanate with alanine, glycine, and several peptides in model systems. J. Agric. Food Chem. 2000, 48, 3560–3565. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Ose, A.; Takai, M.; Kaneko, M.; Nishioka, C.; Ohzu, Y.; Odano, M.; Sekiyama, Y.; Mizukami, Y.; Nakamura, N.; et al. Inhibition of aflatoxin production and fungal growth on stored corn by allyl isothiocyanate vapor. J. Food Hyg. Soc. Jpn. 2015, 56, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Delaquis, P.J.; Sholberg, P.L. Antimicrobial activity of gaseous allyl isothiocyanate. J. Food Prot. 1997, 60, 943. [Google Scholar] [CrossRef]
- Suhr, K.I.; Nielsen, P.V. Antifungal activity of essential oils evaluated by two different application techniques against rye bread spoilage fungi. J. Appl. Microbiol. 2003, 94, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Winther, M.; Nielsen, P.V. Active packaging of cheese with allyl isothiocyanate, an alternative to modified atmosphere packaging. J. Food Prot. 2006, 69, 2430–2435. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.M.; Manyes, L.; Luciano, F.; Manes, J.; Meca, G. Antimicrobial compound allylisothiocyanate against the Aspergillus parasiticus growth and its aflatoxins production in pizza crust. Food Chem. Toxicol. 2015, 83, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Tracz, B.L.; Bordina, K.; Nazareth, T.M.; Costa, L.B.; Macedo, R.E.F.; Meca, G.; Luciano, F.B. Assessment of allyl isothiocyanate as a fumigant to avoid mycotoxin production during corn storage. LWT 2017, 75, 692–696. [Google Scholar] [CrossRef]
- Saladino, F.; Quiles, J.M.; Luciano, F.B.; Mañes, J.; Fernandez-Franzón, M.; Meca, G. Shelf life improvement of the loaf bread using allyl, phenyl and benzyl isothiocyanates against Aspergillus parasiticus. LWT 2017, 78, 208–214. [Google Scholar] [CrossRef]
- Serrano, A.B.; Font, G.; Mañes, J.; Ferrer, E. Emerging Fusarium mycotoxins in organic and conventional pasta collected in Spain. Food Chem. Toxicol. 2013, 51, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Hocking, A.D. Fungi and FoodSpoilage; Blackie Academic and Professional: London, UK, 1997; Volume II. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | LOD (μg/Kg) | LOQ (μg/Kg) | Recovery (%) | ME (%) |
|---|---|---|---|---|
| AFB1 | 0.08 | 0.27 | 70.4 | 78.2 |
| AFB2 | 0.08 | 0.27 | 64.2 | 76.5 |
| AFG1 | 0.16 | 0.53 | 62.8 | 65.3 |
| AFG2 | 0.30 | 1.00 | 66.1 | 60.9 |
| OTA | 0.05 | 0.17 | 75.6 | 89.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiles, J.M.; Nazareth, T.d.M.; Luz, C.; Luciano, F.B.; Mañes, J.; Meca, G. Development of an Antifungal and Antimycotoxigenic Device Containing Allyl Isothiocyanate for Silo Fumigation. Toxins 2019, 11, 137. https://doi.org/10.3390/toxins11030137
Quiles JM, Nazareth TdM, Luz C, Luciano FB, Mañes J, Meca G. Development of an Antifungal and Antimycotoxigenic Device Containing Allyl Isothiocyanate for Silo Fumigation. Toxins. 2019; 11(3):137. https://doi.org/10.3390/toxins11030137
Chicago/Turabian StyleQuiles, Juan Manuel, Tiago de Melo Nazareth, Carlos Luz, Fernando Bittencourt Luciano, Jordi Mañes, and Giuseppe Meca. 2019. "Development of an Antifungal and Antimycotoxigenic Device Containing Allyl Isothiocyanate for Silo Fumigation" Toxins 11, no. 3: 137. https://doi.org/10.3390/toxins11030137
APA StyleQuiles, J. M., Nazareth, T. d. M., Luz, C., Luciano, F. B., Mañes, J., & Meca, G. (2019). Development of an Antifungal and Antimycotoxigenic Device Containing Allyl Isothiocyanate for Silo Fumigation. Toxins, 11(3), 137. https://doi.org/10.3390/toxins11030137