Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications

 ,
, 
Abstract
1. Introduction
2. Results
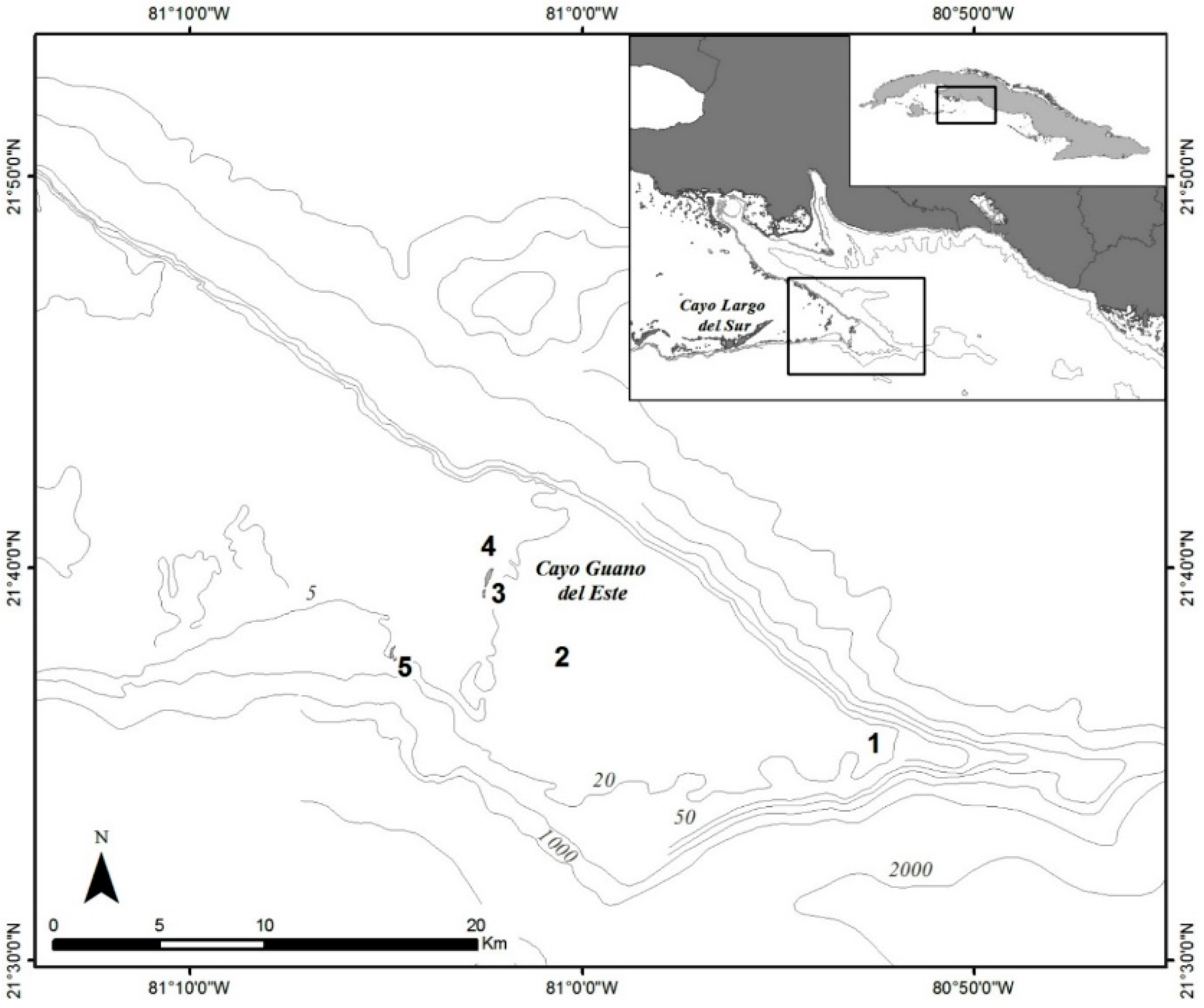
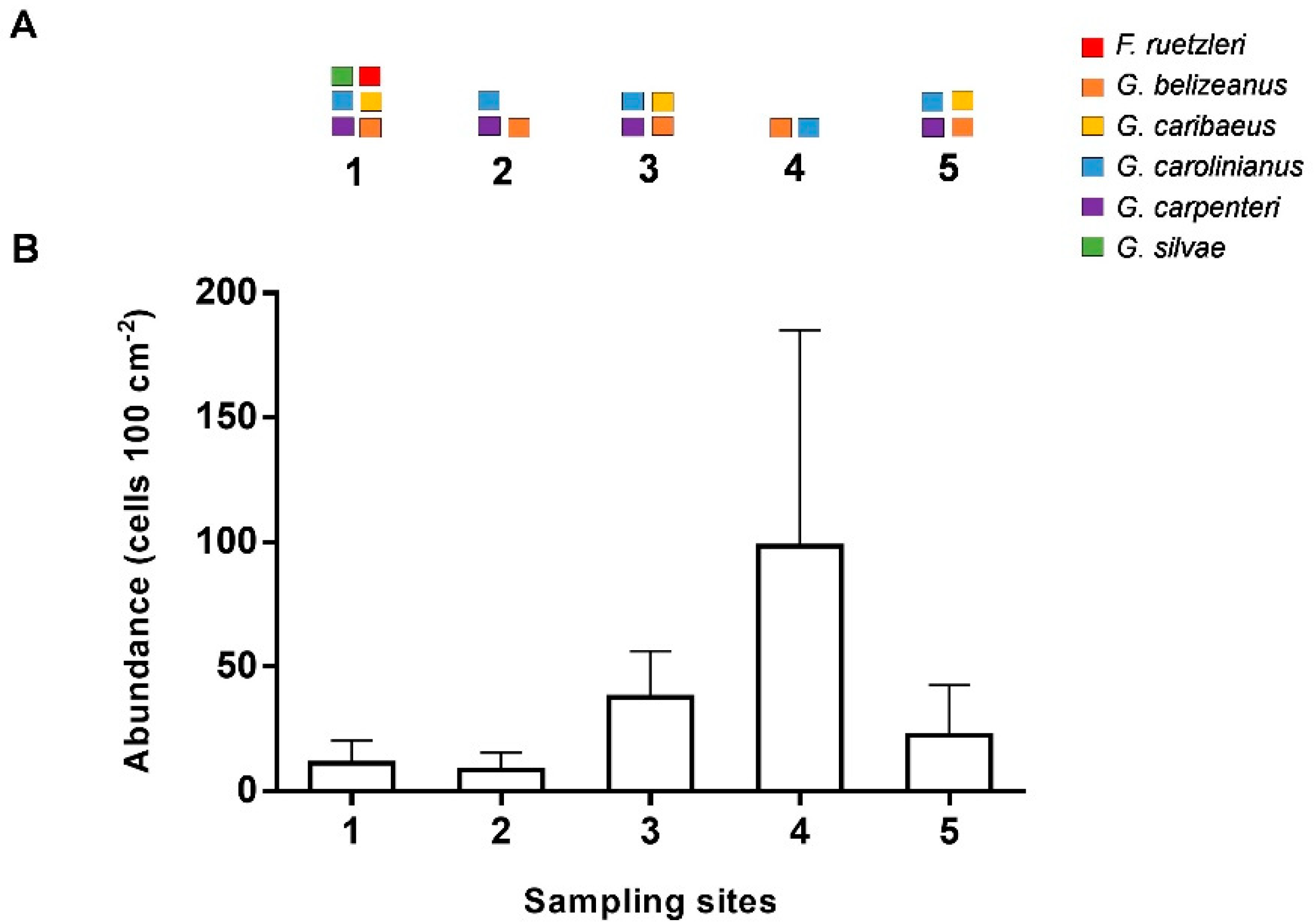
2.1. Benthic Dinoflagellate Composition
2.2. Identification and Toxicity of Gambierdiscus in Culture
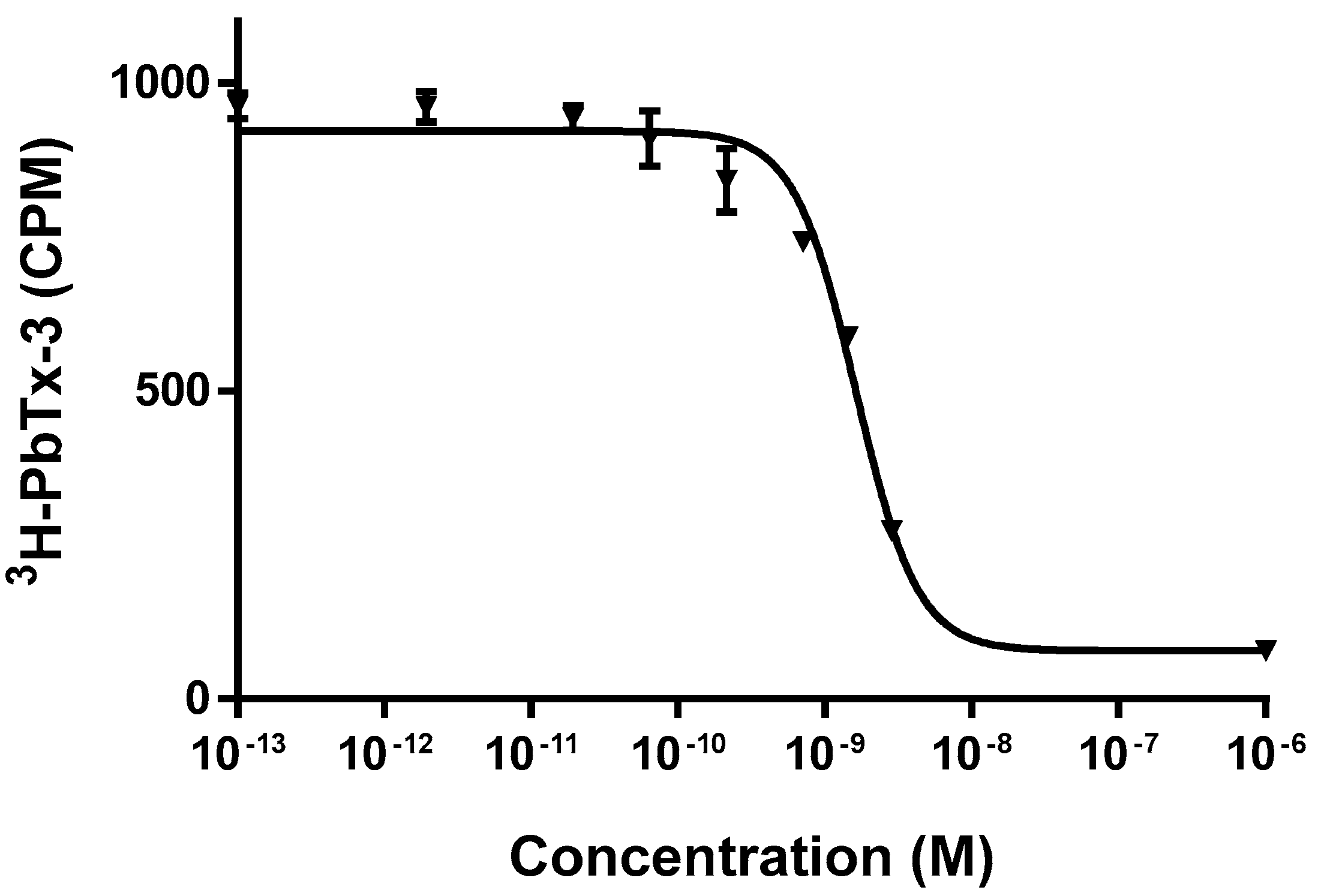
2.3. Toxicity Assessment of Invertebrate and Fish Samples
3. Discussion
4. Materials and Methods
4.1. Sample Collection
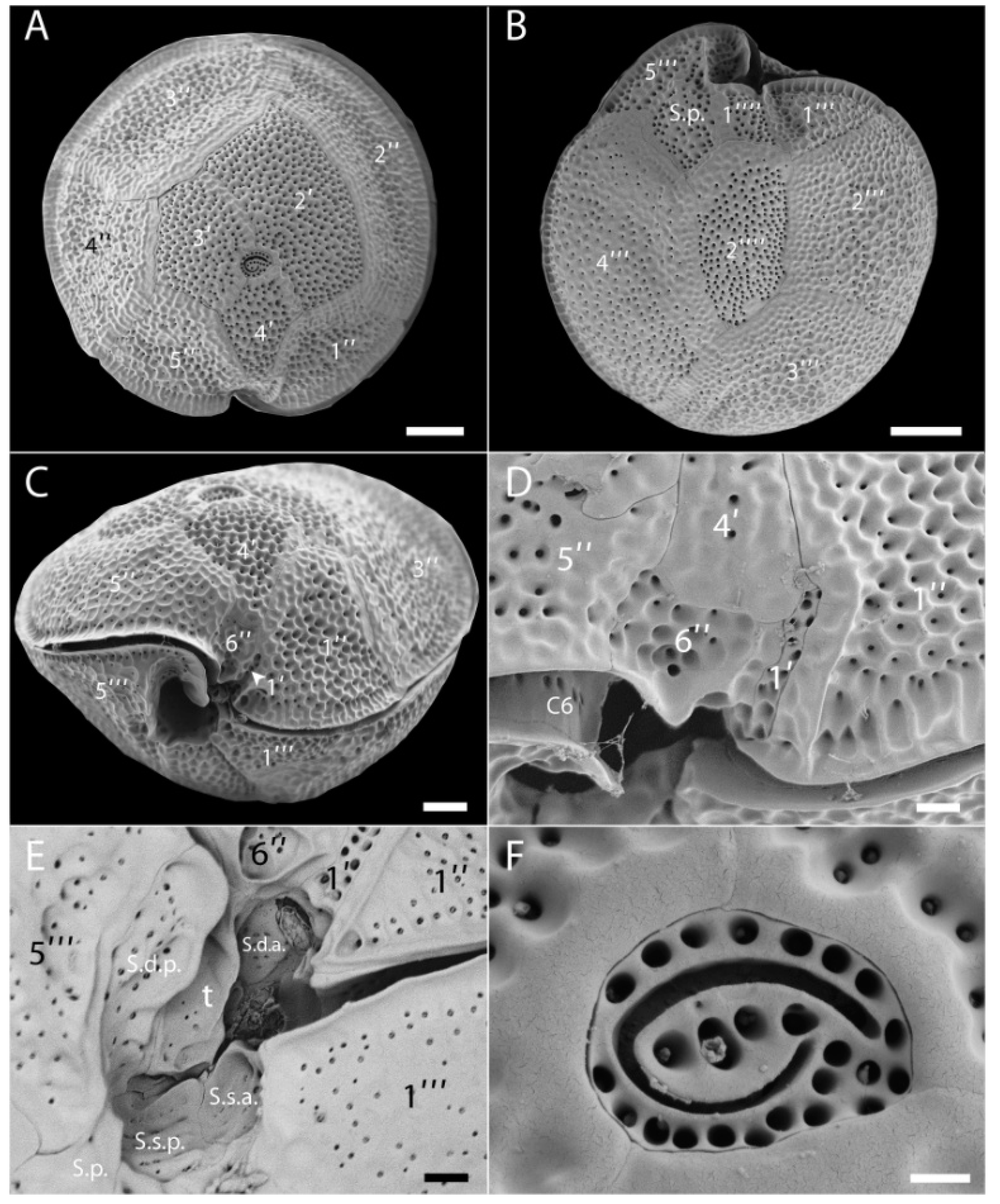
4.2. Gambierdiscus/Fukuyoa Identification and Counting
4.3. Molecular Identification of Gambierdiscus and Fukuyoa Species in Field Samples
4.4. Gambierdiscus Isolation and Culture
Morphological Identification
4.5. Ciguatoxin Analysis in Cell and Fish Samples
4.5.1. Toxin Extraction
4.5.2. Toxin Measurement Using a Radioligand-Receptor Binding Assay
4.5.3. Toxin Measurement Using a Cell-Based Assay (CBA-N2A)
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Friedman, M.; Fernandez, M.; Backer, L.; Dickey, R.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Dechraoui Bottein, M.-Y.; Kashinsky, L.; Wang, Z.; Littnan, C.; Ramsdell, J.S. Identification of ciguatoxins in Hawaiian monk seals Monachus schauinslandi from the Northwestern and main Hawaiian islands. Environ. Sci. Technol. 2011, 45, 5403–5409. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food. Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Uehara, K.; Shah, M.M.R.; Suda, S.; Yasumoto, T.; Taira, Y.; Yamaguchi, H.; et al. Genetic diversity and distribution of the ciguatera-causing dinoflagellate Gambierdiscus spp. (Dinophyceae) in coastal areas of Japan. PLoS ONE 2013, 8, e60882. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Murray, S.; Harwood, D.; Trnski, T.; Munday, R. The Epiphytic Genus Gambierdiscus (Dinophyceae) in the Kermadec Islands and Zealandia Regions of the Southwestern Pacific and the Associated Risk of Ciguatera Fish Poisoning. Mar. Drugs 2017, 15, 219. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Model. 2015, 316, 194–210. [Google Scholar] [CrossRef]
- Hossen, V.; Soliño, L.; Leroy, P.; David, E.; Velge, P.; Dragacci, S.; Krys, S.; Flores Quintana, H.; Diogène, J. Contribution to the risk characterization of ciguatoxins: LOAEL estimated from eight ciguatera fish poisoning events in Guadeloupe (French West Indies). Environ. Res. 2015, 143, 100–108. [Google Scholar] [CrossRef]
- Pottier, I.; Vernoux, J.-P.; Lewis, R.J. Ciguatera Fish Poisoning in the Caribbean Islands and Western Atlantic. In Reviews of Environmental Contamination and Toxicology: Continuation of Residue Reviews; Ware, G.W., Ed.; Springer: New York, NY, USA, 2001; pp. 99–141. [Google Scholar]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.-P.; Brereton, I.M. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Robertson, A.; Quintana, H.A.F.; Silander, M.C.; Smith, T.B.; Olsen, D. Ciguatoxin prevalence in 4 commercial fish species along an oceanic exposure gradient in the US Virgin Islands. Environ. Toxicol. Chem. 2018, 37, 1852–1863. [Google Scholar] [CrossRef] [PubMed]
- Pottier, I.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Analysis of toxin profiles in three different fish species causing ciguatera fish poisoning in Guadeloupe, French West Indies. Food Addit. Contam. 2002, 19, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Soliño, L.; Widgy, S.; Pautonnier, A.; Turquet, J.; Loeffler, C.R.; Flores Quintana, H.A.; Diogène, J. Prevalence of ciguatoxins in lionfish (Pterois spp.) from Guadeloupe, Saint Martin, and Saint Barthélmy Islands (Caribbean). Toxicon 2015, 102, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Bogdanoff, A.K.; Morris, J.A.J.; Quintana, H.A.F.; Buddo, D.; et al. Ciguatoxin concentrations in invasive lionfish from the Caribbean region: Implications for fishery development and management. PLoS ONE 2018, 13, e0198358. [Google Scholar] [CrossRef]
- Pottier, I.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Characterisation of multiple Caribbean ciguatoxins and congeners in individual specimens of horse-eye jack (Caranx latus) by high-performance liquid chromatography/mass spectrometry. Toxicon 2002, 40, 929–939. [Google Scholar] [CrossRef]
- Dechraoui, M.Y.B.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- Lewis, R.J.; Jones, A.; Vernoux, J.-P. HPLC/Tandem Electrospray Mass Spectrometry for the determination of sub-ppblLevels of Pacific and Caribbean ciguatoxins in crude extracts of fish. Anal. Chem. 1999, 71, 247–250. [Google Scholar] [CrossRef]
- Pottier, I.; Hamilton, B.; Jones, A.; Lewis, R.J.; Vernoux, J.P. Identification of slow and fast-acting toxins in a highly ciguatoxic barracuda (Sphyraena barracuda) by HPLC/MS and radiolabelled ligand binding. Toxicon 2003, 42, 663–672. [Google Scholar] [CrossRef]
- Vernoux, J.-P.; Lewis, R.J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef]
- Litaker, R.W.; Holland, W.; Hardison, R.; Pisapia, F.; Hess, P.; Kibler, S.; Tester, P. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef]
- Dechraoui, M.-Y.; Naar, J.; Pauillac, S.; Legrand, A.-M. Ciguatoxins and brevetoxins, neurotoxic polyether compounds active on sodium channels. Toxicon 1999, 37, 125–143. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in Ciguatera Fish Poisoning in the Pacific by LC/MS. J. AOAC Int. 2014, 97, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Tester, P.A.; Chinain, M.; Fraga, S.; Lemée, R.; Litaker, W.; Penna, A.; Usup, G.; Vila, M.; Zingone, A. Harmful Algal Blooms in benthic systems: Recent progress and future research. Oceanogr 2017, 30, 36–45. [Google Scholar] [CrossRef]
- FAO Perfiles de Sobre la Pesca y Acuicultura por Paises. Cuba. Available online: http://www.fao.org/fishery/ (accessed on 28 May 2019).
- López, R.M.G. Felipe Poey y Aloy Obras; Ediciones Imagen Contemporánea: La Habana, Cuba, 1999; Volume 6. [Google Scholar]
- Pis, M. Experiencia Sobre Ciguatera en Islas del Caribe; OceanDocs; UNESCO/IOC Project office for IODE: Oostende, Belgium, 2010. [Google Scholar]
- Arencibia, G. Aspecto de interés sobre la ciguatera en Cuba. Rev. Electron. Vet. 2009, 10, 1–6. [Google Scholar]
- Morrison, K.E.; Aguiar, P.; Castro, A.; Waltner-Toews, D.; FitzGibbon, J. Ciguatera fish poisoning in La Habana, Cuba: A study of local social-ecological resilience. EcoHealth 2008, 5, 346–359. [Google Scholar] [CrossRef]
- Morrison, K.E.; Prieto, P.A.; Dominguez, A.C.; Waltner-Toews, D. An ecosystem approach to ciguatera fish poisoning in Cuba: Preliminary results. In Proceedings of the OCEANS 2005 MTS/IEEE, Washington, DC, USA, 17–23 September 2005; pp. 1817–1822. [Google Scholar]
- Faust, M.A. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely different sites including two new species. J. Phycol. 1995, 31, 996–1003. [Google Scholar] [CrossRef]
- Díaz-Asencio, L.; Vandersea, M.W.; Chomérat, N.; Fraga, S.; Clausing, R.J.; Litaker, R.W.; Chamero-Lago, D.; Gómez-Batista, M.; Moreira-González, A.; Tester, P.; et al. Morphology, toxicity and molecular characterization of Gambierdiscus spp. towards risk assessment of ciguatera in south central Cuba. Harmful Algae 2019, 86, 119–127. [Google Scholar] [CrossRef]
- Tester, P.A.; Kibler, S.R.; Holland, W.C.; Usup, G.; Vandersea, M.W.; Leaw, C.P.; Teen, L.P.; Larsen, J.; Mohammad-Noor, N.; Faust, M.A.; et al. Sampling harmful benthic dinoflagellates: Comparison of artificial and natural substrate methods. Harmful Algae 2014, 39, 8–25. [Google Scholar] [CrossRef]
- Yong, H.L.; Mustapa, N.I.; Lee, L.K.; Lim, Z.F.; Tan, T.H.; Usup, G.; Gu, H.; Litaker, R.W.; Tester, P.A.; Lim, P.T.; et al. Habitat complexity affects benthic harmful dinoflagellate assemblages in the fringing reef of Rawa Island, Malaysia. Harmful Algae 2018, 78, 56–68. [Google Scholar] [CrossRef]
- Tan, T.-H.; Lim, P.-T.; Mujahid, A.; Usup, G.; Leaw, C.-P. Benthic harmful dinoflagellates assemblages in a fringing reef of Sampadi Island, Sarawak, Malaysia. Mar. Res. Indo. 2013, 38, 77–87. [Google Scholar] [CrossRef][Green Version]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Fouc, M.T.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Gaboriau, M.; Ponton, D.; Darius, H.T.; Chinain, M. Ciguatera fish toxicity in French Polynesia: Size does not always matter. Toxicon 2014, 84, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological investigations on the sea urchin Tripneustes gratilla (Toxopneustidae, Echinoid) from Anaho Bay (Nuku Hiva, French Polynesia): Evidence for the presence of Pacific ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.I.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of Ciguatera Poisoning: Detection of Pacific ciguatoxins in toxic samples from Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 2. [Google Scholar] [CrossRef]
- Gatti, C.M.I.; Lonati, D.; Darius, H.T.; Zancan, A.; Roue, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of Ciguatera Poisoning: Clinical characterization and follow-up of a mass poisoning event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef]
- Clausing, R.J.; Chinain, M.; Bottein, M.-Y.D. Practical sampling guidance for determination of ciguatoxin in fish. In Guide for Designing and Implementing a Plant to Monitor Toxin-Producing Microalgae, 2nd ed.; Reguera, B., Alonso, R., Moreira, A., Mendez, S., Bottein, M.-Y.D., Eds.; Intergovernmental Oceanographic Commission (IOC) of UNESCO: Paris, France; International Atomic Energy Agency (IAEA): Vienna, Austria, 2016; pp. 51–63. [Google Scholar]
- Poli, M.A.; Lewis, R.J.; Dickey, R.W.; Musser, S.M.; Buckner, C.A.; Carpenter, L.G. Identification of Caribbean ciguatoxins as the cause of an outbreak of fish poisoning among U.S. soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar] [CrossRef]
- Olsen, D.A.; Nellis, D.W.; Wood, R.S. Ciguatera in the Eastern Caribbean. Mar. Fish. Rev. 1984, 46, 13–18. [Google Scholar]
- Mak, Y.L.; Wai, T.-C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific ciguatoxins in food web components of coral reef systems in the Republic of Kiribati. Environ. Sci. Technol. 2013, 47, 14070–14079. [Google Scholar] [CrossRef]
- Soliño, L.; Costa, P.R. Differential toxin profiles of ciguatoxins in marine organisms: Chemistry, fate and global distribution. Toxicon 2018, 150, 124–143. [Google Scholar] [CrossRef]
- Ledreux, A.; Brand, H.; Chinain, M.; Bottein, M.-Y.D.; Ramsdell, J.S. Dynamics of ciguatoxins from Gambierdiscus polynesiensis in the benthic herbivore Mugil cephalus: Trophic transfer implications. Harmful Algae 2014, 39, 165–174. [Google Scholar] [CrossRef]
- Yang, Z.; Luo, Q.; Liang, Y.; Mazumder, A. Processes and pathways of ciguatoxin in aquatic food webs and fish poisoning of seafood consumers. Environ. Rev. 2016, 24, 144–150. [Google Scholar] [CrossRef]
- Tosteson, T.R.; Ballantine, D.L.; Durst, H.D. Seasonal frequency of ciguatoxic barracuda in southwest Puerto Rico. Toxicon 1988, 26, 795–801. [Google Scholar] [CrossRef]
- Clausing, R.J.; Losen, B.; Oberhaensli, F.R.; Darius, H.T.; Sibat, M.; Hess, P.; Swarzenski, P.W.; Chinain, M.; Marie-Yasmine, D.B. Experimental evidence of dietary ciguatoxin accumulation in an herbivorous coral reef fish. Aquat. Toxicol. 2018, 200, 257–265. [Google Scholar] [CrossRef]
- FDA. Natural Toxins. In Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; U.S. Department of Health and Human Services: Washington, DC, USA, 2011; pp. 99–112. Available online: https://tamug-ir.tdl.org/bitstream/handle/1969.3/29148/UCM251970%2004242012.pdf?sequence=1 (accessed on 14 November 2019).
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.F.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef]
- Inoue, M.; Hirama, M.; Satake, M.; Sugiyama, K.; Yasumoto, T. Inhibition of brevetoxin binding to the voltage-gated sodium channel by gambierol and gambieric acid-A. Toxicon 2003, 41, 469–474. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; del Río, V.; de la Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef]
- Tester, P.A.; Vandersea, M.W.; Buckel, C.A.; Kibler, S.R.; Holland, W.C.; Davenport, E.D.; Clark, R.D.; Edwards, K.F.; Taylor, J.C.; Pluym, J.L.V.; et al. Gambierdiscus (Dinophyceae) species diversity in the Flower Garden Banks National Marine Sanctuary, Northern Gulf of Mexico, USA. Harmful Algae 2013, 29, 1–9. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase, Version 06/2018; World Wide Web Electronic Publication. Available online: https://fishbase.org/ (assessed on 14 November 2019).
- FAO. The living marine resources of the Western Central Atlantic. Volume 2: Bony fishes part 1 (Acipenseridae to Grammatidae). In FAO Species Identification Guide for Fishery Purposes; Carpenter, K.E., Ed.; FAO: Rome, Italy, 2002; pp. 601–1374. [Google Scholar]
- Vandersea, M.W.; Kibler, S.R.; Holland, W.C.; Tester, P.A.; Schultz, T.F.; Faust, M.A.; Holmes, M.J.; Chinain, M.; Litaker, R.W. Development of semi-quantitative PCR assays for the detection and enumeration of Gambierdiscus species (Gonyaulacales, Dinophyceae). J. Phycol. 2012, 48, 902–915. [Google Scholar] [CrossRef]
- Litaker, R.W.; Tester, P.A.; Vandersea, M.W. Species-specific PCR assays for Gambierdiscus excentricus and Gambierdiscus silvae (Gonyaulacales, Dinophyceae). J. Phycol. 2019, 55, 730–732. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar] [CrossRef]
- Chomérat, N.; Couté, A. Protoperidinium bolmonense sp. nov. (Peridiniales, Dinophyceae), a small dinoflagellate from brackish hypereutrophic lagoon (South of France). Phycologia 2008, 47, 392–403. [Google Scholar] [CrossRef]
- Besada, E.G.; Loeblich, L.A.; Loeblich, A.R., III. Observations on tropical, benthic dinoflagellates form ciguatera-endemic areas: Coolia, Gambierdiscus, and Ostreopsis. Bull. Mar. Sci. 1982, 32, 723–735. [Google Scholar]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Asencio, L.; Clausing, R.J.; Rañada, M.L.; Alonso-Hernández, C.M.; Bottein, M.-Y.D. A radioligand receptor binding assay for ciguatoxin monitoring in environmental samples: Method development and determination of quality control criteria. J. Environ. Radioact. 2018, 192, 289–294. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- IAEA-TECDOC-1729. Detection of Harmful Algal Toxins Using the Radioligand Receptor Binding Assay A Manual of Methods; IAEA: Vienna, Austria, 2013. [Google Scholar]
- Bottein, M.-Y.D.; Clausing, R.J. Receptor-Binding Assay for the Analysis of Marine Toxins: Detection and Mode of Action. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; Volume 78, pp. 277–295. [Google Scholar]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogène, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. 2012, 29, 1000–1010. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Local Name | Family | Weight (#) |
|---|---|---|---|---|
| Mycteroperca bonaci | Black grouper | Aguají o bonací arará | Serranidae | (1) |
| Mycteroperca venenosa | Yellowfin grouper | Arigua o bonací cardenal | Serranidae | (2) |
| Mycteroperca tigris | Tiger grouper | Bonací gato | Serranidae | (3) |
| Carangoides bartholomaei | Yellowjack | Cibí amarillo | Carangidae | (4) |
| Seriola rivoliana | Longfin yellowtail | Coronado | Carangidae | (3) |
| Seriola zonata | Banded rudderfish | Coronado de bandas | Carangidae | (3) |
| Seriola dumerili | Greater amberjack | Coronado de ley | Carangidae | (3) |
| Lutjanus cyanopterus | Cubera snapper | Cubera | Lutjanidae | (5) |
| Caranx latus | Horse-eyejack | Gallego o jurel | Carangidae | (6) |
| Chilomycterus reticulatus | Spotfin burrfish | Guanábana | Diodontidae | (3) |
| Rypticus saponaceus | Greater soapfish | Jaboncillo o jabón | Grammistidae | (3) |
| Lutjanus jocu | Dog snapper | Pargo jocú | Lutjanidae | (4) |
| Gymnothorax funebris | Green moray | Morena verde | Muraenidae | (3) |
| Ogcocephalus vespertilio | Seadevil | Pez diablo | Ogcocephalidae | (3) |
| Diodon holocanthus | Longspined porcupinefish | Pez erizo | Diodontidae | (3) |
| Sphyraena barracuda | Great barracuda | Picúa o Picuda | Sphyraenidae | (3) |
| Diodon hystrix | Spot-fin porcupinefish | Puerco espín | Diodontidae | (3) |
| Lagocephalus laevigatus | Smooth puffer | Tamboril gigante | Tetrodontidae | (3) |
| Sphoeroides testudineus | Checkered puffer | Tamboril rayado | Tetrodontidae | (3) |
| Caranx lugubris | Black jack | Tiñosa o Tiñosa prieta | Carangidae | (3) |
| Species | Family | Trophic Level | Weight (range) g | Total Length (range) cm | N | RBA+ | RBA− |
|---|---|---|---|---|---|---|---|
| Invertebrates | |||||||
| Diadema antillarum | Diadematidae | 2 | 2 | 0 | |||
| Eucidaris tribuloides* | Cidaridae | 1 | 0 | 1 | |||
| Holothuria mexicana | Holothuriidae | 1 | 0 | 1 | |||
| Lobatus gigas | Strombidae | 5 | 0 | 5 | |||
| Panulirus argus | Palinuridae | 1 | 1 | 0 | |||
| Herbivorous fishes | |||||||
| Acanthurus chirurgus | Acanthuridae | 2 | 86–220 | 17–21 | 6 | 1 | 5 (1) |
| Acanthurus coeruleus | Acanthuridae | 2 | 150–310 | 19–23 | 4 | 3 | 1 |
| Sparisoma aurofrenatum | Scaridae | 2 | 147–165 | 20–21 | 3 | 3 | 0 |
| Sparisoma chrysopterum | Scaridae | 2 | 180 | 21 | 1 | 0 | 1 (1) |
| Stegastes diencaeus | Pomacentridae | 2 | 22 | 11 | 1 | 1 | 0 |
| Carnivorous fishes | |||||||
| Apsilus dentatus | Lutjanidae | 4.08 | 950–1615 | 40–48 | 2 | 2 | 0 |
| Caranx latus | Carangidae | 4.16 | 1355–2200 | 49–59 | 10 | 10 | 0 |
| Caranx lugubris | Carangidae | 4.0 | 3195–3505 | 66–68 | 3 | 2 | 1 |
| Chaetodon capistratus | Chaetodontidae | 3.43 | 13 | 8 | 1 | 1 | 0 |
| Epinephelus guttattus | Serranidae | 4.15 | 287 | 27 | 1 | 1 | 0 |
| Heteropriacanthus cruentatus | Priacanthidae | 3.7 | 113 | 19 | 1 | 1 | 0 |
| Holocentrus rufus | Holocentridae | 3.49 | 69 | 21 | 1 | 1 | 0 |
| Hypoplectrus unicolor | Serranidae | 3.97 | 12 | 9 | 1 | 1 | 0 |
| Lutjanus buccanella | Lutjanidae | 3.9 | 1025–1170 | 41–43 | 2 | 2 | 0 |
| Lutjanus jocu | Lutjanidae | 3.92 | 3350–5250 | 58–62 | 2 | 2 | 0 |
| Mycteroperca bonaci | Serranidae | 4.5 | 2000 | 55 | 1 | 1 (1) | 0 |
| Mycteroperca venenosa | Serranidae | 4.5 | 753–9400 | 37–89 | 6 | 6 (2) | 0 |
| Pterois volitans | Scorpaenidae | 3.44 | 234 | 25 | 1 | 0 | 1 |
| Rhomboplites aurorubens | Lutjanidae | 3.8 | 685 | 37 | 1 | 1 | 0 |
| Scomberomorus regalis | Scombridae | 4.38 | 636 | 51 | 1 | 1 (1) | 0 |
| Seriola rivoliana | Carangidae | 4.5 | 1795–13650 | 54–111 | 3 | 3 (2) | 0 |
| Seriola dumerili | Carangidae | 4.5 | 14000 | 110 | 1 | 1 | 0 |
| Sphyraena barracuda | Sphyraenidae | 4.5 | 785–6400 | 56–112 | 12 | 12 (1) | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Asencio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gómez-Batista, M.; Hernández-Albernas, J.I.; Chomérat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications. Toxins 2019, 11, 722. https://doi.org/10.3390/toxins11120722
Díaz-Asencio L, Clausing RJ, Vandersea M, Chamero-Lago D, Gómez-Batista M, Hernández-Albernas JI, Chomérat N, Rojas-Abrahantes G, Litaker RW, Tester P, et al. Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications. Toxins. 2019; 11(12):722. https://doi.org/10.3390/toxins11120722
Chicago/Turabian StyleDíaz-Asencio, Lisbet, Rachel J. Clausing, Mark Vandersea, Donaida Chamero-Lago, Miguel Gómez-Batista, Joan I. Hernández-Albernas, Nicolas Chomérat, Gabriel Rojas-Abrahantes, R. Wayne Litaker, Patricia Tester, and et al. 2019. "Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications" Toxins 11, no. 12: 722. https://doi.org/10.3390/toxins11120722
APA StyleDíaz-Asencio, L., Clausing, R. J., Vandersea, M., Chamero-Lago, D., Gómez-Batista, M., Hernández-Albernas, J. I., Chomérat, N., Rojas-Abrahantes, G., Litaker, R. W., Tester, P., Diogène, J., Alonso-Hernández, C. M., & Dechraoui Bottein, M.-Y. (2019). Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications. Toxins, 11(12), 722. https://doi.org/10.3390/toxins11120722