High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys

 ,
,  ,
,
Abstract
1. Introduction
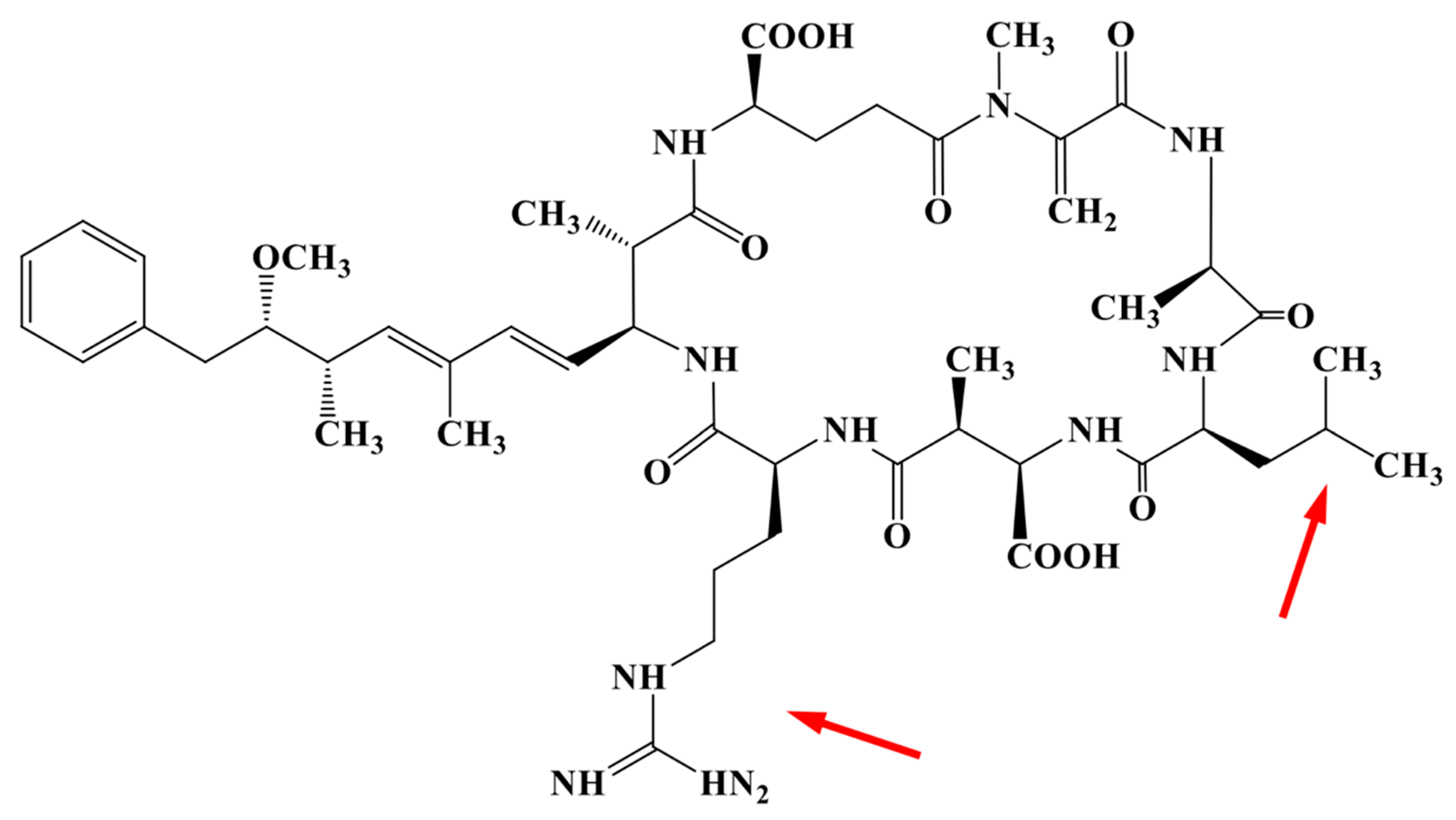
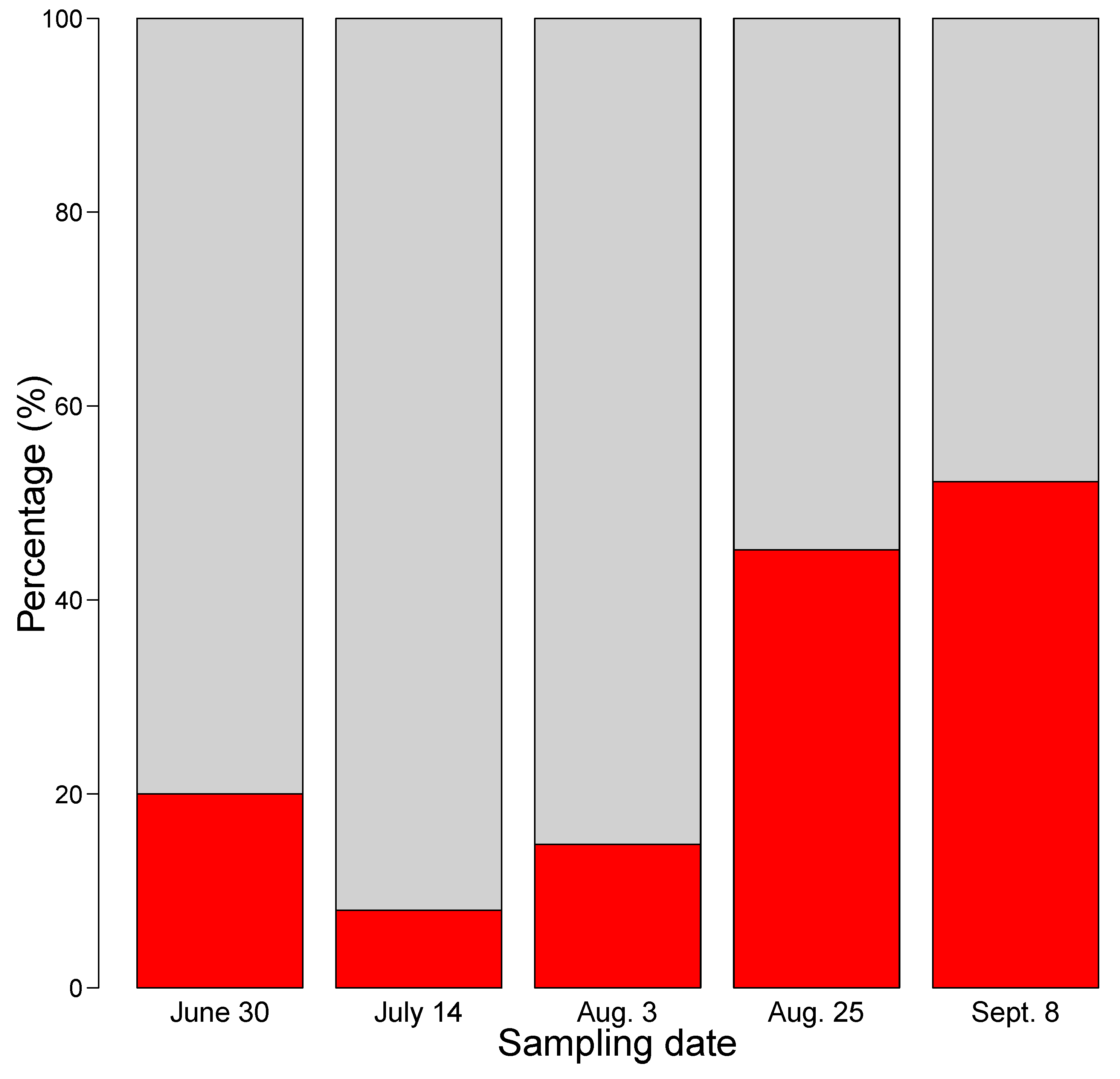
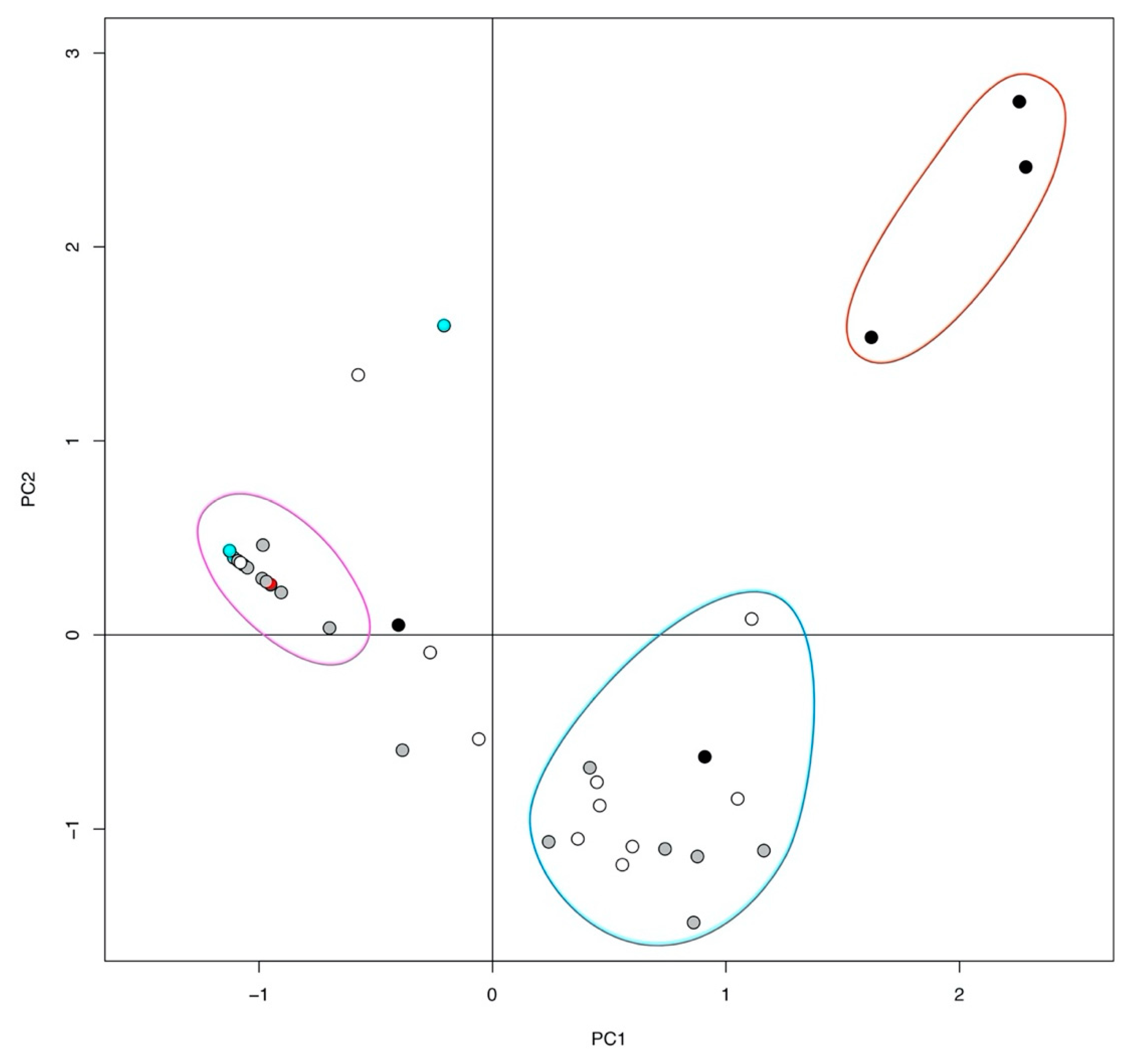
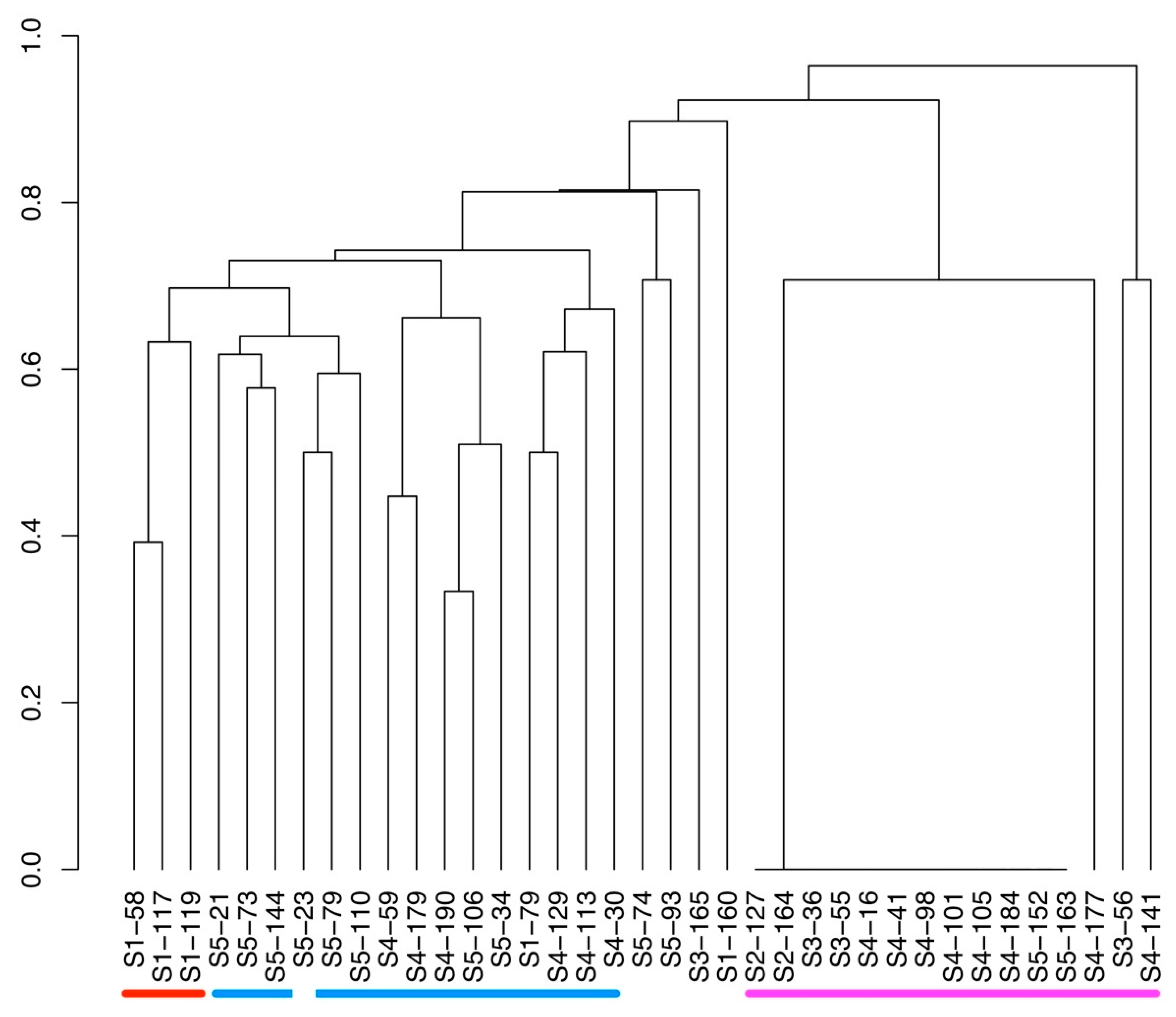
2. Results
3. Discussion
4. Methods
4.1. Sampling and Isolation of Microcystis Colonies
4.2. Harvest of Microcystis botrys Strains
4.3. Extraction and LC-MS/MS Analysis
4.4. Statistical Analysis
4.5. Water Chemistry
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Visser, P.M.; Verspagen, J.M.H.; Sandrini, G.; Stal, L.J.; Matthijs, H.C.P.; Davis, T.W.; Paerl, H.W.; Huisman, J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 2016, 54, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Blooms Like it Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; Donk, E.; Hansson, L.-A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Mantzouki, E.; Lurling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Wozniak, E.; Koreiviene, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krzton, W.; et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins 2018, 10, 76. [Google Scholar] [CrossRef]
- Berner, C.; Bertos-Fortis, M.; Pinhassi, J.; Legrand, C. Response of Microbial Communities to Changing Climate Conditions During Summer Cyanobacterial Blooms in the Baltic Sea. Front. Microbiol. 2018, 9, 1562. [Google Scholar] [CrossRef]
- Konst, H.; McKercher, P.D.; Gorham, P.R.; Robertson, A.; Howell, J. Symptoms and Pathology Produced By Toxic Microcystis aeruginosa NRC-1 In Laboratory and Domestic Animals. Can. J. Comp. Med. Vet. Sci. 1965, 29, 221–228. [Google Scholar]
- Dawson, R.M. The Toxicology of Microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Bartram, J.; Carmichael, W.W.; Chorus, I.; Jones, G.; Skulberg, O.M. Chapter 1: Introduction. In Toxic Cyanobacteria in Water: A Guide to their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; CRC Press: Boca Raton, FL, USA, 1999; p. 400. [Google Scholar]
- Codd, G.A.; Lindsay, J.; Young, F.M.; Morrison, L.F.; Metcalf, J.S. Harmful Cyanobacteria: From mass mortalities to management measures. In Harmful Cyanobacteria, 1st ed.; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 3, pp. 1–23. [Google Scholar]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017. [Google Scholar]
- Humbert, J.-F.; Fastner, J. Ecology of Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017. [Google Scholar]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Goncalves, F.J.; Pereira, M.J. Microcystin-producing blooms-a serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Sano, T.; Kubota, R.; Kobayashi, N.; Tahara, M.; Obama, T.; Sugimoto, N.; Nishimura, T.; Ikarashi, Y. Effects of the amino acid constituents of microcystin variants on cytotoxicity to primary cultured rat hepatocytes. Toxins 2014, 6, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Chapter 3: Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Welker, M.; von Döhren, H. Cyanobacterial peptides-nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Beasley, V.; Bunner, D.L.; Eloff, J.N.; Falconer, I.; Gorham, P.; Harada, K.-I.; Krishnamurthy, T.; Min-Juan, Y.; Moore, R.E.; et al. Naming of Cyclic Heptapeptide Toxins of Cyanobacteria (Blue-Green Algae). Toxicon 1988, 26, 971–973. [Google Scholar] [CrossRef]
- Nishizawa, T.; Asayama, M.; Fujii, K.; Harada, K.-i.; Shirai, M. Genetic Analysis of the Peptide Synthetase Genes for a Cyclic Heptapeptide Microcystin in Microcystis spp. J. Biochem. 1999, 126, 520–529. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Nishizawa, T.; Ueda, A.; Asayama, M.; Fujii, K.; Harada, K.-i.; Ochi, K.; Shirai, M. Polyketide Synthase Gene Coupled to the Peptide Synthetase Module Involved in the Biosynthesis of the Cyclic Heptapeptide Microcystin. J. Biochem. 2000, 127, 779–789. [Google Scholar] [CrossRef]
- Christiansen, G.; Fastner, J.; Erhard, M.; Börner, T.; Dittmann, E. Microcystin Biosynthesis in Planktothrix: Genes, Evolution, and Manipulation. J. Bacteriol. 2003, 185, 564–572. [Google Scholar] [CrossRef]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes coding for hepatotoxic heptapeptides (microcystins) in the cyanobacterium Anabaena strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3 Tables of Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017. [Google Scholar]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Dohren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Park, B.S.; Li, Z.; Kang, Y.H.; Shin, H.H.; Joo, J.H.; Han, M.S. Distinct Bloom Dynamics of Toxic and Non-toxic Microcystis (Cyanobacteria) Subpopulations in Hoedong Reservoir (Korea). Microb. Ecol. 2018, 75, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef]
- Holland, A.; Kinnear, S. Interpreting the possible ecological role(s) of cyanotoxins: Compounds for competitive advantage and/or physiological aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef]
- Ger, K.A.; Faassen, E.J.; Pennino, M.G.; Lurling, M. Effect of the toxin (microcystin) content of Microcystis on copepod grazing. Harmful Algae 2016, 52, 34–45. [Google Scholar] [CrossRef]
- Orr, P.T.; Jones, G.J. Relationship between microcystin production and cell division rates in nitrogen-limited Microcystis aeruginosa cultures. Limnol. Ocanogr. 1998, 43, 1604–1614. [Google Scholar] [CrossRef]
- Kardinaal, W.E.A.; Janse, I.; Kamst-van Agterveld, M.; Meima, M.; Snoek, J.; Mur, L.R.; Huisman, J.; Zwart, G.; Visser, P.M. Microcystis genotype succession in relation to microcystin concentrations in freshwater lakes. Aquat. Microb. Ecol. 2007, 48, 1–12. [Google Scholar] [CrossRef]
- Van der Merwe, D. Cyanobacterial (Blue-Green Algae) Toxins. In Handbook of Toxicology of Chemical Warfare Agents; Elsevier: Kidlington, UK, 2015; pp. 421–429. [Google Scholar] [CrossRef]
- Chiu, Y.-T.; Chen, Y.-H.; Wang, T.-S.; Yen, H.-K.; Lin, T.-F. A qPCR-Based Tool to Diagnose the Presence of Harmful Cyanobacteria and Cyanotoxins in Drinking Water Sources. Int. J. Environ. Res. Public Health 2017, 14, 547. [Google Scholar] [CrossRef] [PubMed]
- Alström, T.; Holmström, K.; Holmström, C.; Davidsson, T.; Björklund, H. Vombsjön Faktasammanställning 2017 (Lake Vomb, A Compilation of Information); Kävlingeåns vattenråd: Landskrona, Sweden, 2017; p. 55. [Google Scholar]
- Ekvall, M.K.; de la Calle Martin, J.; Faassen, E.J.; Gustafsson, S.; Lürling, M.; Hansson, L.-A. Synergistic and species-specific effects of climate change and water colour on cyanobacterial toxicity and bloom formation. Freshw. Biol. 2013, 58, 2414–2422. [Google Scholar] [CrossRef]
- Urrutia-Cordero, P.; Ekvall, M.K.; Hansson, L.A. Controlling Harmful Cyanobacteria: Taxa-Specific Responses of Cyanobacteria to Grazing by Large-Bodied Daphnia in a Biomanipulation Scenario. PLoS ONE 2016, 11, e0153032. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Yepremian, C.; Humbert, J.F.; Quiblier, C. Competition between microcystin- and non-microcystin-producing Planktothrix agardhii (cyanobacteria) strains under different environmental conditions. Environ. Microbiol. 2008, 10, 3337–3348. [Google Scholar] [CrossRef]
- Briand, E.; Escoffier, N.; Straub, C.; Sabart, M.; Quiblier, C.; Humbert, J.-F. Spatiotemporal changes in the genetic diversity of a bloom-forming Microcystis aeruginosa (cyanobacteria) population. ISME J. 2009, 3, 419–429. [Google Scholar] [CrossRef]
- Sabart, M.; Pobel, D.; Briand, E.; Combourieu, B.; Salencon, M.J.; Humbert, J.F.; Latour, D. Spatiotemporal Variations in Microcystin Concentrations and in the Proportions of Microcystin-Producing Cells in Several Microcystis aeruginosa Populations. Appl. Environ. Microb. 2010, 76, 4750–4759. [Google Scholar] [CrossRef]
- Joung, S.-H.; Oh, H.-M.; You, K.-A. Dynamic variation of toxic and non-toxic Microcystis proportion in the eutrophic Daechung Reservoir in Korea. J. Microbiol. 2016, 54, 543–550. [Google Scholar] [CrossRef]
- Kurmayer, R.; Kutzenberger, T. Application of Real-Time PCR for Quantification of Microcystin Genotypes in a Population of the Toxic Cyanobacterium Microcystis sp. Appl. Environ. Microb. 2003, 69, 6723–6730. [Google Scholar] [CrossRef]
- Hansson, L.-A.; Gustafsson, S.; Rengefors, K.; Bomark, L. Cyanobacterial chemical warfare affects zooplankton community composition. Freshw. Biol. 2007, 52, 1290–1301. [Google Scholar] [CrossRef]
- Tonk, L.; Welker, M.; Huisman, J.; Visser, P.M. Production of cyanopeptolins, anabaenopeptins, and microcystins by the harmful cyanobacteria Anabaena 90 and Microcystis PCC 7806. Harmful Algae 2009, 8, 219–224. [Google Scholar] [CrossRef]
- Agha, R.; Cires, S.; Wormer, L.; Dominguez, J.A.; Quesada, A. Multi-scale strategies for the monitoring of freshwater cyanobacteria: Reducing the sources of uncertainty. Water Res 2012, 46, 3043–3053. [Google Scholar] [CrossRef] [PubMed]
- Drugă, B.; Welker, M.; Sesărman, A.; Hegedus, A.; Coman, C.; Sicora, C.; Dragoş, N. Molecular characterization of microcystin-producing cyanobacteria from Romanian fresh waters. Eur. J. Phycol. 2013, 48, 287–294. [Google Scholar] [CrossRef]
- Welker, M.; Marsalek, B.; Sejnohova, L.; von Dohren, H. Detection and identification of oligopeptides in Microcystis (cyanobacteria) colonies: Toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar] [CrossRef]
- Welker, M.; Šejnohová, L.; Némethová, D.; von Döhren, H.; Jarkovský, J.; Maršálek, B. Seasonal shifts in chemotype composition of Microcystis sp. communities in the pelagial and the sediment of a shallow reservoir. Limnol. Ocanogr. 2007, 52, 609–619. [Google Scholar] [CrossRef]
- Agha, R.; Quesada, A. Oligopeptides as Biomarkers of Cyanobacterial Subpopulations. Toward an Understanding of Their Biological Role. Toxins 2014, 6, 1929–1950. [Google Scholar] [CrossRef]
- Spoof, L.; Blaszczyk, A.; Meriluoto, J.; Ceglowska, M.; Mazur-Marzec, H. Structures and Activity of New Anabaenopeptins Produced by Baltic Sea Cyanobacteria. Mar. Drugs 2015, 14, 8. [Google Scholar] [CrossRef]
- Kurmayer, R.; Deng, L.; Entfellner, E. Role of toxic and bioactive secondary metabolites in colonization and bloom formation by filamentous cyanobacteria Planktothrix. Harmful Algae 2016, 54, 69–86. [Google Scholar] [CrossRef]
- Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; John Wiley & Sons, Ltd.: London, UK, 2017.
- DeMott, W.R.; Moxter, F. Foraging Cyanobacteria by Copepods: Responses to Chemical Defense and Resource Abundance. Ecology 1991, 72, 1820–1834. [Google Scholar] [CrossRef]
- DeMott, W.R.; Zhang, Q.-x.; Carmichael, W.W. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Ocanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef]
- Harke, M.J.; Jankowiak, J.G.; Morrell, B.K.; Gobler, C.J. Transcriptomic Responses in the Bloom-Forming Cyanobacterium Microcystis Induced During Exposure to Zooplankton. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Cordero, P.; Agha, R.; Cires, S.; Lezcano, M.A.; Sanchez-Contreras, M.; Waara, K.O.; Utkilen, H.; Quesada, A. Effects of harmful cyanobacteria on the freshwater pathogenic free-living amoeba Acanthamoeba castellanii. Aquat. Toxicol. 2013, 130–131, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Meissner, S.; Steinhauser, D.; Dittmann, E. Metabolomic analysis indicates a pivotal role of the hepatotoxin microcystin in high light adaptation of Microcystis. Environ. Microbiol. 2015, 17, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Jacinavicius, F.R.; Pacheco, A.B.F.; Chow, F.; Verissimo da Costa, G.C.; Kalume, D.E.; Rigonato, J.; Schmidt, E.C.; Sant’Anna, C.L. Different ecophysiological and structural strategies of toxic and non-toxic Microcystis aeruginosa (cyanobacteria) strains assessed under culture conditions. Algal Res. 2019, 41. [Google Scholar] [CrossRef]
- Briand, E.; Bormans, M.; Quiblier, C.; Salencon, M.J.; Humbert, J.F. Evidence of the cost of the production of microcystins by Microcystis aeruginosa under differing light and nitrate environmental conditions. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Blossom, H.E.; Markussen, B.; Daugbjerg, N.; Krock, B.; Norlin, A.; Hansen, P.J. The Cost of Toxicity in Microalgae: Direct Evidence from the Dinoflagellate Alexandrium. Front. Microbiol. 2019, 10, 1065. [Google Scholar] [CrossRef]
- Kardinaal, W.E.; Tonk, L.; Janse, I.; Hol, S.; Slot, P.; Huisman, J.; Visser, P.M. Competition for light between toxic and nontoxic strains of the harmful cyanobacterium Microcystis. Appl. Environ. Microbiol. 2007, 73, 2939–2946. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- Leblanc Renaud, S.; Pick, F.R.; Fortin, N. Effect of light intensity on the relative dominance of toxigenic and nontoxigenic strains of Microcystis aeruginosa. Appl. Environ. Microbiol. 2011, 77, 7016–7022. [Google Scholar] [CrossRef] [PubMed]
- Suominen, S.; Brauer, V.S.; Rantala-Ylinen, A.; Sivonen, K.; Hiltunen, T. Competition between a toxic and a non-toxic Microcystis strain under constant and pulsed nitrogen and phosphorus supply. Aquat. Ecol. 2017, 51, 117–130. [Google Scholar] [CrossRef]
- Harris, T.D.; Smith, V.H.; Graham, J.L.; Van de Waal, D.B.; Tedesco, L.P.; Clercin, N. Combined effects of nitrogen to phosphorus and nitrate to ammonia ratios on cyanobacterial metabolite concentrations in eutrophic Midwestern USA reservoirs. Inland Waters 2016, 6, 199–210. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Davis, T.W.; Smith, D.J.; Baer, M.M.; Dick, G.J. Interactions between nitrogen form, loading rate, and light intensity on Microcystis and Planktothrix growth and microcystin production. Harmful Algae 2018, 73, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Newell, S.E.; Davis, T.W.; Johengen, T.H.; Gossiaux, D.; Burtner, A.; Palladino, D.; McCarthy, M.J. Reduced forms of nitrogen are a driver of non-nitrogen-fixing harmful cyanobacterial blooms and toxicity in Lake Erie. Harmful Algae 2019, 81, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Lorenzen, C.J. Yellow-green algae with chlorophyllide C1,2. J. Phycol. 1972, 10–14. [Google Scholar] [CrossRef]
- Staub, R. Ernährungsphysiologisch-autökologische Untersuchungen an der planktischen Blaualge Oscillatoria rubescens DC. Schweiz. Z. Hydrol. 1961, 23. [Google Scholar] [CrossRef]
- IBM Corp. SPSS Statistics for Macintosh; Version 25.0, Release 25.0.0.1; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- RStudio; 1.1.453; R Core Team: Vienna, Austria, 2018; Available online: https://rstudio.com/products/rstudio/ (accessed on 10 October 2019).
- Mazur-Marzec, H.; Bertos-Fortis, M.; Torunska-Sitarz, A.; Fidor, A.; Legrand, C. Chemical and Genetic Diversity of Nodularia spumigena from the Baltic Sea. Mar. Drugs 2016, 14, 209. [Google Scholar] [CrossRef]
- Synergy Software KaleidaGraph; Version 4.5. Available online: http://www.synergy.com/wordpress_650164087/ (accessed on 10 October 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Prefix | Sampling Date | Water Temp at 0.5 m Depth (°C) | Colonies Isolated | Cultures Established | Cultures Analyzed | Number of MC Producing Strains |
|---|---|---|---|---|---|---|
| S1 | 2014-06-30 | 19 | 143 | 35 | 25 | 5 |
| S2 | 2014-07-14 | 20 | 192 | 33 | 25 | 2 |
| S3 | 2014-08-03 | 26 | 192 | 35 | 27 | 4 |
| S4 | 2014-08-25 | 18 | 148 | 35 | 30 | 14 |
| S5 | 2014-09-08 | 19 | 168 | 30 | 23 | 12 |
| Sampling Date (2014) | Toxigenic Strain ID | Microcystin Variants | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MC-WR | [Asp3]MC-ThTyrR | MC-RY | MC-RR | MC? | [Asp3] MC-RY | MC-FR | [Asp3] MC-RR | [Dha7] MC-RR | MC-HilR | [Ser1] MC-VR | MC-LR | [Asp3Dhb7] MC-LR | MC? | MC? | MC? | MC? | MC? | ||
| m/z [M+H]+ | 1035 | 861 | 528 | 1031 | 509 | 502 | |||||||||||||
| June 30 | S1-58 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||
| S1-79 | ● | ● | ● | ● | ● | ● | ● | ||||||||||||
| S1-117 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||
| S1-119 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||
| S1-160 | ● | ● | ● | ||||||||||||||||
| July 14 | S2-127 | ● | |||||||||||||||||
| S2-164 | ● | ||||||||||||||||||
| August 3 | S3-36 | ● | |||||||||||||||||
| S3-55 | ● | ||||||||||||||||||
| S3-56 | ● | ||||||||||||||||||
| S3-165 | ● | ● | ● | ● | |||||||||||||||
| August 25 | S4-16 | ● | |||||||||||||||||
| S4-30 | ● | ● | ● | ● | |||||||||||||||
| S4-41 | ● | ||||||||||||||||||
| S4-59 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||
| S4-98 | ● | ||||||||||||||||||
| S4-101 | ● | ||||||||||||||||||
| S4-105 | ● | ||||||||||||||||||
| S4-113 | ● | ● | ● | ● | ● | ||||||||||||||
| S4-129 | ● | ● | ● | ● | ● | ● | ● | ||||||||||||
| S4-141 | ● | ● | |||||||||||||||||
| S4-177 | ● | ● | |||||||||||||||||
| S4-179 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||
| S4-184 | ● | ||||||||||||||||||
| S4-190 | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||
| September 8 | S5-21 | ● | ● | ● | ● | ● | ● | ● | |||||||||||
| S5-23 | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||
| S5-34 | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||
| S5-73 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||
| S5-74 | ● | ● | ● | ● | ● | ||||||||||||||
| S5-79 | ● | ● | ● | ● | ● | ● | |||||||||||||
| S5-93 | ● | ● | ● | ● | |||||||||||||||
| S5-106 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||
| S5-110 | ● | ● | ● | ● | ● | ● | ● | ||||||||||||
| S5-144 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||
| S5-152 | ● | ||||||||||||||||||
| S5-163 | ● | ||||||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansson, E.; Legrand, C.; Björnerås, C.; Godhe, A.; Mazur-Marzec, H.; Säll, T.; Rengefors, K. High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys. Toxins 2019, 11, 698. https://doi.org/10.3390/toxins11120698
Johansson E, Legrand C, Björnerås C, Godhe A, Mazur-Marzec H, Säll T, Rengefors K. High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys. Toxins. 2019; 11(12):698. https://doi.org/10.3390/toxins11120698
Chicago/Turabian StyleJohansson, Emma, Catherine Legrand, Caroline Björnerås, Anna Godhe, Hanna Mazur-Marzec, Torbjörn Säll, and Karin Rengefors. 2019. "High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys" Toxins 11, no. 12: 698. https://doi.org/10.3390/toxins11120698
APA StyleJohansson, E., Legrand, C., Björnerås, C., Godhe, A., Mazur-Marzec, H., Säll, T., & Rengefors, K. (2019). High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys. Toxins, 11(12), 698. https://doi.org/10.3390/toxins11120698