Bacillus thuringiensis Maize Expressing a Fusion Gene Cry1Ab/Cry1AcZM Does Not Harm Valued Pollen Feeders


Abstract
1. Introduction
2. Result
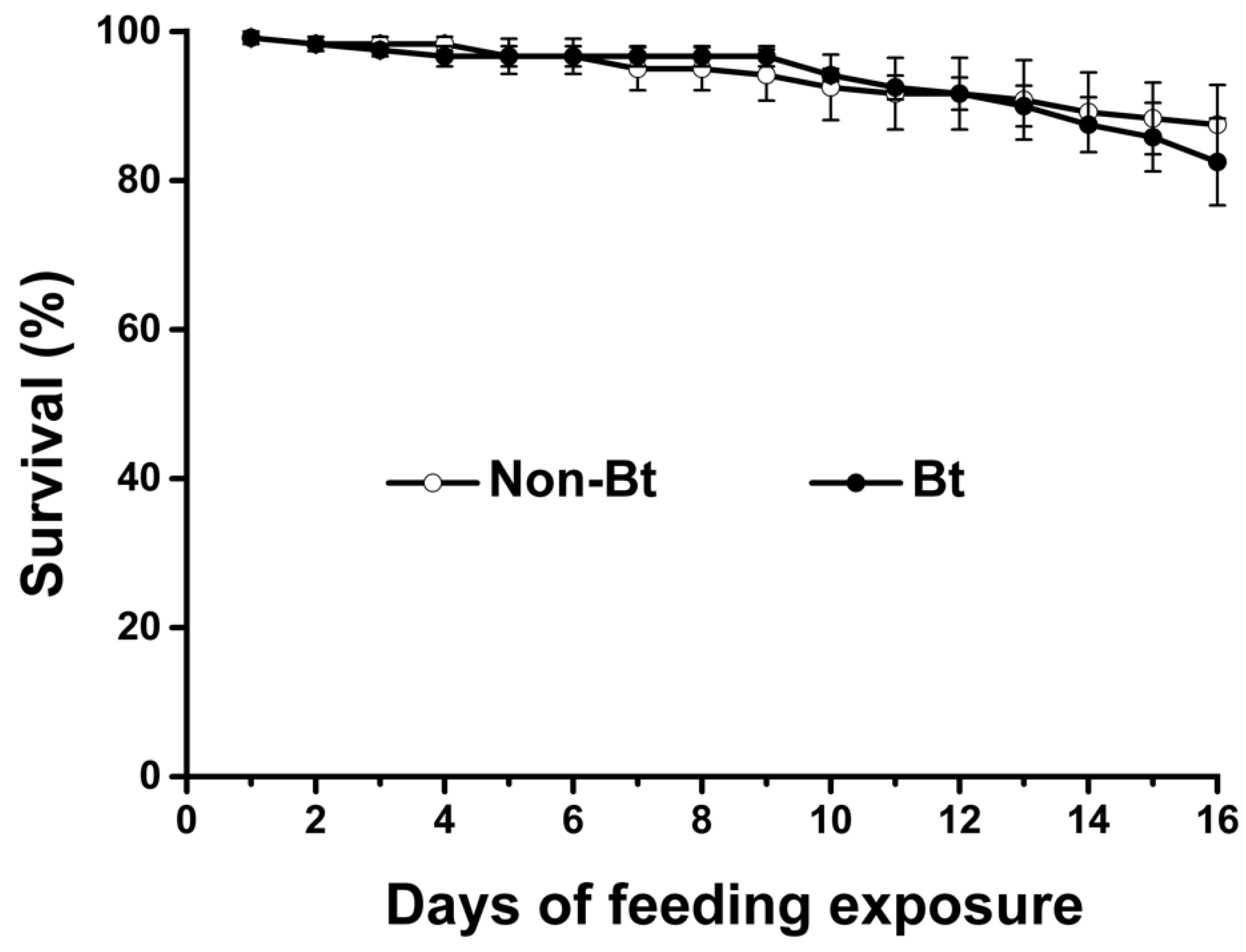
2.1. Effects on Life-Table Parameters
2.2. Uptake of Cry Protein by Test Species
2.3. Stability of Cry Proteins in Maize Pollen during the Feeding Exposure
2.4. Bioactivity of Cry1Ab/Cry1Ac in Maize Pollen during the Feeding Exposure
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Maize Plants and Pollen Collection
4.3. Feeding Experiment with P. japonica
4.4. Feeding Experiment with C. nipponensis
4.5. Feeding Experiment with A. mellifera
4.6. Stability of Cry Protein in Maize Pollen during the Feeding Exposure
4.7. ELISA Measurements
4.8. Bioactivity of Cry1Ab/Cry1Ac Protein in Bt Maize Pollen
4.9. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Hellmich, R.L.; Albajes, R.; Bergvinson, D.; Prasifka, J.R.; Wang, Z.Y.; Weiss, M.J. The present and future role of insect-resistant genetically modified maize in IPM. In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs; Romeis, J., Kennedy, G.G., Shelton, A.M., Eds.; Springer Science + Business Media B.V.: New York, NY, USA, 2008; pp. 119–158. [Google Scholar]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2017: Biotech Crop Adoption Surges as Economic Benefits Accumulate in 22 Years; ISAAA Brief No. 53; ISAAA: Ithaca, NY, USA, 2017. [Google Scholar]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alonso, M.; Jacobs, E.; Raybould, A.; Nickson, T.E.; Sowig, P.; Willekens, H.; Van der Kouwe, P.; Layton, R.; Amijee, F.; Fuentes, A.M.; et al. A tiered system for assessing the risk of genetically modified plants to non-target organisms. Environ. Biosaf. Res. 2007, 5, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.; Hallerman, E.M.; Wu, K. Biosafety management and commercial use of genetically modified crops in China. Plant Cell Rep. 2014, 33, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Bartsch, D.; Bigler, F.; Candolfi, M.P.; Gielkens, M.M.C.; Hartley, S.E.; Hellmich, R.L.; Huesing, J.E.; Jepson, P.C.; Layton, R.; et al. Assessment of risk of insect-resistant transgenic crops to nontarget arthropods. Nat. Biotechnol. 2008, 26, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Sanvido, O.; Romeis, J.; Gathmann, A.; Gielkens, M.; Raybould, A.; Bigler, F. Evaluating environmental risks of genetically modified crops: Ecological harm criteria for regulatory decision-making. Environ. Sci. Policy 2012, 15, 82–91. [Google Scholar] [CrossRef]
- Duan, J.J.; Marvier, M.; Huesing, J.; Dively, G.; Huang, Z.Y. A meta-analysis of effects of Bt crops on honey bees (Hymenoptera: Apidae). PLoS ONE 2008, 3, e1415. [Google Scholar] [CrossRef] [PubMed]
- Icoz, I.; Stotzky, G. Fate and effects of insect-resistant Bt crops in soil ecosystems. Soil Biol. Biochem. 2008, 40, 559–586. [Google Scholar] [CrossRef]
- Li, Y.H.; Romeis, J.; Wu, K.M.; Peng, Y.F. Tier-1 assays for assessing the toxicity of insecticidal proteins produced by genetically engineered plants to non-target arthropods. Insect Sci. 2014, 21, 125–134. [Google Scholar] [CrossRef]
- Marvier, M.; Mccreedy, C.; Regetz, J.; Kareiva, P. A meta-analysis of effects of Bt cotton and maize on nontarget invertebrates. Science 2007, 316, 1475–1477. [Google Scholar] [CrossRef]
- Naranjo, S.E. Impacts of Bt crops on non-target invertebrates and insecticide use patterns. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2009, 4, 1–23. [Google Scholar] [CrossRef]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Contr. 2018. [Google Scholar] [CrossRef]
- Romeis, J.; Meissle, M.; Bigler, F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat. Biotechnol. 2006, 24, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Wolfenbarger, L.L.; Naranjo, S.E.; Lundgren, J.G.; Bitzer, R.J.; Watrud, L.S. Bt crop effects on functional guilds of non-target arthropods: A meta-analysis. PLoS ONE 2008, 3, e2118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Lang, Z.H.; Zhang, J.; He, K.L.; Song, F.P.; Huang, D.F. Ubi1 intron-mediated enhancement of the expression of Bt cry1Ah gene in transgenic maize (Zea mays L.). Chin. Sci. Bull. 2008, 53, 3185–3190. [Google Scholar] [CrossRef]
- Lv, X.; Wang, H.; Zeng, X.; Yang, X.; Weng, J.; Di, H.; Guo, Y.; Wang, Z.; Li, X. Research and application of transgenic Bt corn for insect resistance. Crops 2013, 2, 7–12. [Google Scholar]
- Devos, Y.; Romeis, J.; Luttik, R.; Maggiore, A.; Perry, J.N.; Schoonjans, R.; Streissl, F.; Tarazona, J.V.; Brock, T.C. Optimising environmental risk assessments: Accounting for ecosystem services helps to translate broad policy protection goals into specific operational ones for environmental risk assessments. EMBO Rep. 2015, 16, 1060–1063. [Google Scholar] [CrossRef]
- Romeis, J.; Raybould, A.; Bigler, F.; Candolfi, M.P.; Hellmich, R.L.; Huesing, J.E.; Shelton, A.M. Deriving criteria to select arthropod species for laboratory tests to assess the ecological risks from cultivating arthropod-resistant genetically engineered crops. Chemosphere 2013, 90, 901–909. [Google Scholar] [CrossRef]
- Romeis, J.; Meissle, M.; Álvarez-Alfageme, F.; Bigler, F.; Bohan, D.A.; Devos, Y.; Malone, L.A.; Pons, X.; Rauschen, S. Potential use of an arthropod database to support the non-target risk assessment and monitoring of transgenic plants. Transgenic Res. 2014, 23, 995–1013. [Google Scholar] [CrossRef]
- Bai, Y.Y.; Jiang, M.X.; Cheng, J.A. Effects of transgenic cry1Ab rice pollen on fitness of Propylea japonica (Thunberg). J. Pest Sci. 2005, 78, 123–128. [Google Scholar] [CrossRef]
- Zhou, K.J.; Xiang, J.B. Observations on the efficacy of spiders and ladybirds against aphids in the seedling stage of cotton in the cotton fields. Nat. Enemies Insects 1987, 9, 17–20. [Google Scholar]
- Zhang, S.Y.; Li, D.M.; Cui, J.; Xie, B.Y. Effects of Bt-toxin Cry1Ac on Propylaea japonica Thunberg (Col., Coccinellidae) by feeding on Bt-treated Bt-resistant Helicoverpa armigera (Hubner) (Lep., Noctuidae) larvae. J. Appl. Entomol. 2006, 130, 206–212. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, Q.L.; Liu, Q.S.; Meissle, M.; Yang, Y.; Wang, Y.N.; Hua, H.X.; Chen, X.P.; Peng, Y.F.; Romeis, J. Bt rice in China—Focusing the nontarget risk assessment. Plant Biotechnol. J. 2017, 15, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meissle, M.; Romeis, J. Use of maize pollen by adult Chrysoperla carnea (Neuroptera: Chrysopidae) and fate of Cry proteins in Bt -transgenic varieties. J. Insect Physiol. 2010, 56, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Meissle, M.; Naranjo, S.; Li, Y.; Bigler, F. The end of a myth—Bt (Cry1Ab) maize does not harm green lacewings. Front. Plant Sci. 2014, 5, 391. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Niu, C.Y.; Biondi, A.; Desneux, N. Does transgenic Cry1Ac+CpTI cotton pollen affect hypopharyngeal gland development and midgut proteolytic enzyme activity in the honey bee Apis mellifera L. (Hymenoptera, Apidae)? Ecotoxicology 2012, 21, 2214–2221. [Google Scholar] [CrossRef]
- Hendriksma, H.P.; Kuting, M.; Hartel, S.; Nather, A.; Dohrmann, A.B.; Steffan-Dewenter, I.; Tebbe, C.C. Effect of stacked insecticidal Cry proteins from maize pollen on nurse bees (Apis mellifera carnica) and their gut bacteria. PLoS ONE 2013, 8, e59589. [Google Scholar] [CrossRef]
- Rose, R.; Dively, G.P.; Pettis, J. Effects of Bt corn pollen on honey bees: Emphasis on protocol development. Apidologie 2007, 38, 368–377. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, P.; Chen, X.; Romeis, J.; Shi, J.; Peng, Y.; Li, Y. Ingestion of Bt rice pollen does not reduce the survival or hypopharyngeal gland development of Apis mellifera adults. Environ. Toxicol. Chem. 2016, 36, 1243–1248. [Google Scholar] [CrossRef]
- Li, Y.; Meissle, M.; Romeis, J. Consumption of Bt maize pollen expressing Cry1Ab or Cry3Bb1 does not harm adult green Lacewings, Chrysoperla carnea (Neuroptera: Chrysopidae). PLoS ONE 2008, 3, e2909. [Google Scholar] [CrossRef]
- Malone, L.A. Potential effects of GM crops on honey bee health. Bee World 2004, 85, 29–36. [Google Scholar] [CrossRef]
- Romeis, J.; Hellmich, R.L.; Candolfi, M.P.; Carstens, K.; De Schrijver, A.; Gatehouse, A.M.R.; Herman, R.A.; Huesing, J.E.; Mclean, M.A.; Raybould, A.; et al. Recommendations for the design of laboratory studies on non-target arthropods for risk assessment of genetically engineered plants. Transgenic Res. 2011, 20, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Romeis, J.; Chen, X.; Zhang, J.; Chen, H.; Peng, Y. Consumption of Bt rice pollen expressing Cry2Aa does not cause adverse effects on adult Chrysoperla sinica Tjeder (Neuroptera: Chrysopidae). Biol. Contr. 2012, 61, 246–251. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Chen, X.; Romeis, J.; Yin, X.; Peng, Y. Consumption of Bt rice pollen containing Cry1C or Cry2A does not pose a risk to Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Sci. Rep. 2015, 5, 7679. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Yin, X.; Romeis, J.; Song, X.; Chen, X.; Geng, L.; Peng, Y.; Li, Y. Consumption of Bt maize pollen containing Cry1Ie does not negatively affect Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Toxins 2017, 9, 108. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Wang, Y.; Chen, X.; Song, X.; Romeis, J.; Li, Y.; Peng, Y. Ingestion of Bt corn pollen containing Cry1Ab/2Aj or Cry1Ac does not harm Propylea japonica larvae. Sci. Rep. 2016, 6, 23507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Li, Y.H.; Romeis, J.; Yin, X.M.; Wu, K.M.; Peng, Y.F. Use of a pollen-based diet to expose the ladybird beetle Propylea japonica to insecticidal proteins. PLoS ONE 2014, 9, e85395. [Google Scholar] [CrossRef]
- Meissle, M.; Zünd, J.; Waldburger, M.; Romeis, J. Development of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) on pollen from Bt-transgenic and conventional maize. Sci. Rep. 2014, 4, 5900. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Vojtech, E. The effects of pollen consumption of transgenic Bt maize on the common swallowtail, Papilio machaon L. (Lepidoptera, Papilionidae). Basic Appl. Ecol. 2006, 7, 296–306. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Romeis, J.; Liu, Q.; Lin, K.; Chen, X.; Peng, Y. Bt rice expressing Cry2Aa does not cause direct detrimental effects on larvae of Chrysoperla sinica. Ecotoxicology 2013, 22, 1413–1421. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Hu, L.; Romeis, J.; Peng, Y. Bt rice producing Cry1C protein does not have direct detrimental effects on the green lacewing Chrysoperla sinica (Tjeder). Environ. Toxicol. Chem. 2014, 33, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.C.; Wang, X.P.; Long, L.P.; Romeis, J.; Naranjo, S.E.; Hellmich, R.L.; Wang, P.; Earle, E.D.; Shelton, A.M. Bt crops producing Cry1Ac, Cry2Ab and Cry1F do not harm the green lacewing, Chrysoperla rufilabris. PLoS ONE 2013, 8, e60125. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J.; Warren, J.F.; du Plessis, H. The potential effect of Bt maize on Chrysoperla pudica (Neuroptera: Chrysopidae). Environ. Entomol. 2017, 46, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Hilbeck, A.; Moar, W.J.; Pusztai-Carey, M.; Filippin, A.; Bigler, F. Toxicity of Bacillus thuringiensis Cry1Ab toxin to the predator Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol. 1998, 27, 1255–1263. [Google Scholar] [CrossRef]
- Mason, C.E.; Sheldon, J.K.; Pesekadke, G.; Slabaugh, B. Assessment of Chrysoperla plorabunda longevity, fecundity, and egg viability when adults are fed transgenic Bt corn pollen. J. Agric. Urban Entomol. 2008, 25, 265–278. [Google Scholar] [CrossRef]
- Babendreier, D.; Kalberer, N.M.; Romeis, J.; Fluri, P.; Mulligan, E.; Bigler, F. Influence of Bt-transgenic pollen, Bt-toxin and protease inhibitor (SBTI) ingestion on development of the hypopharyngeal glands in honeybees. Apidologie 2005, 36, 585–594. [Google Scholar] [CrossRef]
- Malone, L.A.; Todd, J.H.; Burgess, E.P.J.; Christeller, J.T. Development of hypopharyngeal glands in adult honey bees fed with a Bt-toxin, a biotin-binding protein and a protease inhibitor. Apidologie 2004, 35, 655–664. [Google Scholar] [CrossRef]
- Sagili, R.R.; Pankiw, T.; Zhusalzman, K. Effects of soybean trypsin inhibitor on hypopharyngeal gland protein content, total midgut protease activity and survival of the honey bee (Apis mellifera L.). J. Insect Physiol. 2005, 51, 953–957. [Google Scholar] [CrossRef]
- Han, L.Z.; Li, S.B.; Peng, Y.F.; Hou, M.L. New artificial diet for continuous rearing of Chilo suppressalis. Ann. Entomol. Soc. Am. 2012, 105, 253–258. [Google Scholar] [CrossRef]
- Liu, B.L.; Tan, C.; Yang, Q.Q.; Xu, J.T.; Wen, Q.; Qiu, L.; Ma, C.L.; Zhang, W.G. Gene for Encoding Bacillus thuringiensis Crystal Proteins and Application Thereof. PCT/CN2016/090978. 22 July 2016. [Google Scholar]
- Xu, Y.J. Diapause Mechanism and Application of Chrysoperla sinica (Tjeder). Ph.D. Dissertation, Zhejiang University, Zhejiang, China, 2001. (In Chinese with English Abstract). [Google Scholar]
- Niu, L.; Ma, Y.; Mannakkara, A.; Zhao, Y.; Ma, W.; Lei, C.; Chen, L. Impact of single and stacked insect-resistant Bt-cotton on the honey bee and silkworm. PLoS ONE 2013, 8, e72988. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | Non-Bt Pollen | Bt Pollen | Statistics |
|---|---|---|---|
| Larval survival rate (%) a | 91.43 (70) | 92.86 (70) | X2 = 0.10, P = 0.75 |
| Pupation rate (%) a | 75.71 (70) | 90.00 (70) | X2 = 5.03, P = 0.03 |
| Eclosion rate (%) a | 90.57 (53) | 95.24 (63) | X2 = 0.98, P = 0.32 |
| Days to pupa (days ± SE) b | 9.34 ± 0.20 (52) | 9.07 ± 0.14 (63) | U = 1485, P = 0.37 |
| Female fresh weight (mg ± SE) c | 6.07 ± 0.22 (23) | 6.26 ± 0.17 (34) | t = −0.68, P = 0.50 |
| Male fresh weight (mg ± SE) c | 4.60 ± 0.16 (25) | 5.21 ± 0.15 (26) | t = −2.74, P = 0.01 |
| Pre-oviposition period (days ± SE) b | 4.81 ± 0.61 (16) | 4.65 ± 0.42 (20) | U = 145, P = 0.39 |
| Total fecundity per pair over 21 d (eggs ± SE) c | 120.81 ± 10.68 (16) | 129.00 ± 9.23 (20) | t = −0.71, P = 0.48 |
| Parameters | Bt Pollen | Non-Bt Pollen | Statistics |
|---|---|---|---|
| Male survival (%) a | 90 (30) | 90 (30) | X2 = 0.48, P = 0.49 |
| Female survival (%) a | 79.31 (29) | 71.43 (28) | X2 = 0.48, P = 0.49 |
| Pre-oviposition period (days ± SE) b | 10.19 ± 0.55 (26) | 9.26 ± 0.56 (27) | U = 289, P = 0.26 |
| Total fecundity (eggs ± SE) c | 63.96 ± 7.56 (25) | 79.04 ± 10.25 (25) | t = −1.18, P = 0.24 |
| Male fresh weight (mg ± SE) c | 4.61 ± 0.18 (27) | 4.59 ± 0.15 (27) | t = 0.08, P = 0.93 |
| Female fresh weight (mg ± SE) c | 6.94 ± 0.32 (23) | 6.87 ± 0.26 (20) | t = 0.16, P = 0.87 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, X.; Cui, Z.; Wang, Y.; Wang, Y.; Cao, F.; Romeis, J.; Peng, Y.; Li, Y. Bacillus thuringiensis Maize Expressing a Fusion Gene Cry1Ab/Cry1AcZM Does Not Harm Valued Pollen Feeders. Toxins 2019, 11, 8. https://doi.org/10.3390/toxins11010008
Xie X, Cui Z, Wang Y, Wang Y, Cao F, Romeis J, Peng Y, Li Y. Bacillus thuringiensis Maize Expressing a Fusion Gene Cry1Ab/Cry1AcZM Does Not Harm Valued Pollen Feeders. Toxins. 2019; 11(1):8. https://doi.org/10.3390/toxins11010008
Chicago/Turabian StyleXie, Xiaowei, Zhifu Cui, Yanan Wang, Yuanyuan Wang, Fengqin Cao, Jörg Romeis, Yufa Peng, and Yunhe Li. 2019. "Bacillus thuringiensis Maize Expressing a Fusion Gene Cry1Ab/Cry1AcZM Does Not Harm Valued Pollen Feeders" Toxins 11, no. 1: 8. https://doi.org/10.3390/toxins11010008
APA StyleXie, X., Cui, Z., Wang, Y., Wang, Y., Cao, F., Romeis, J., Peng, Y., & Li, Y. (2019). Bacillus thuringiensis Maize Expressing a Fusion Gene Cry1Ab/Cry1AcZM Does Not Harm Valued Pollen Feeders. Toxins, 11(1), 8. https://doi.org/10.3390/toxins11010008