Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation

Abstract
1. Introduction
2. Results
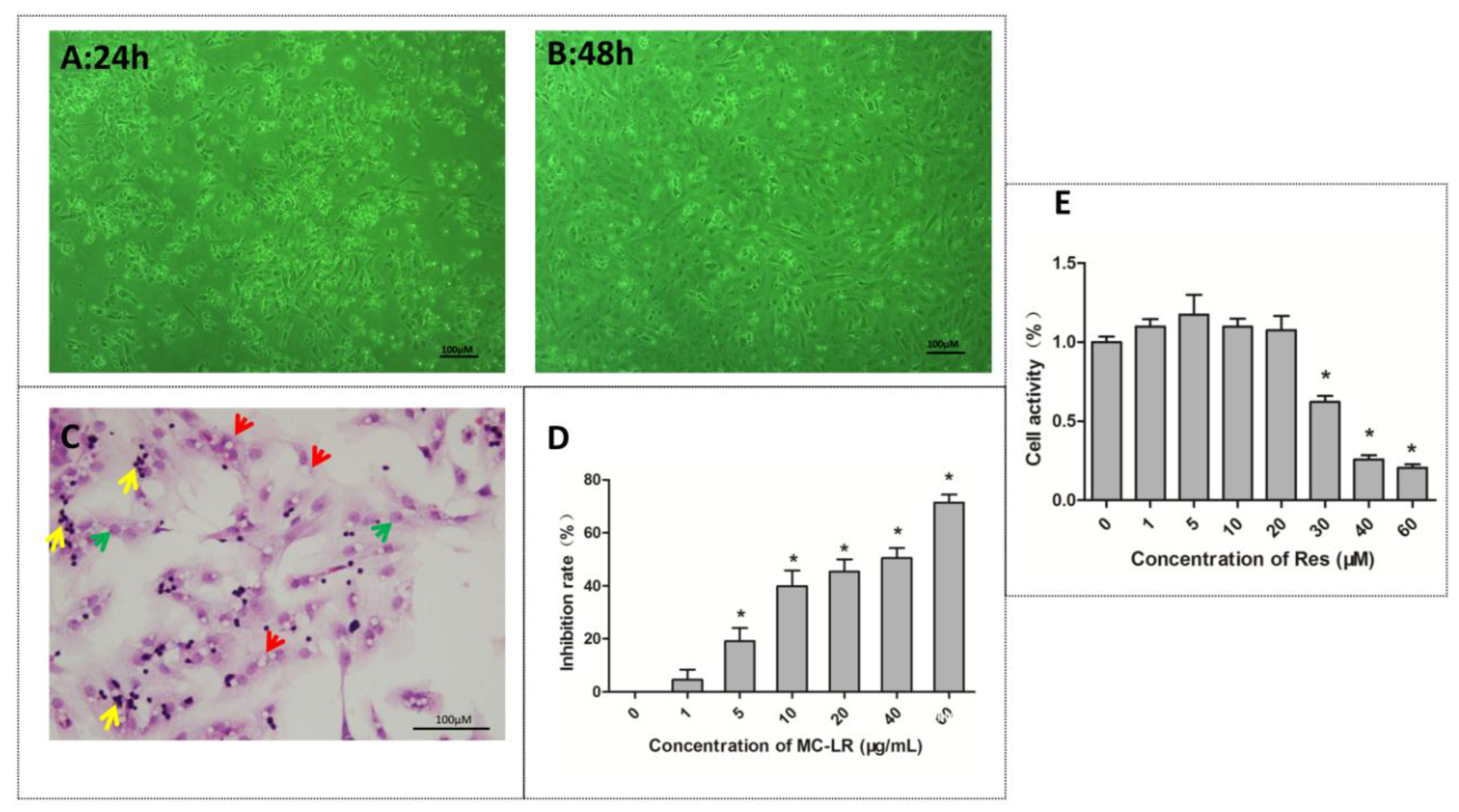
2.1. The Identification and Viability of Co-Cultured Sertoli–Germ Cells
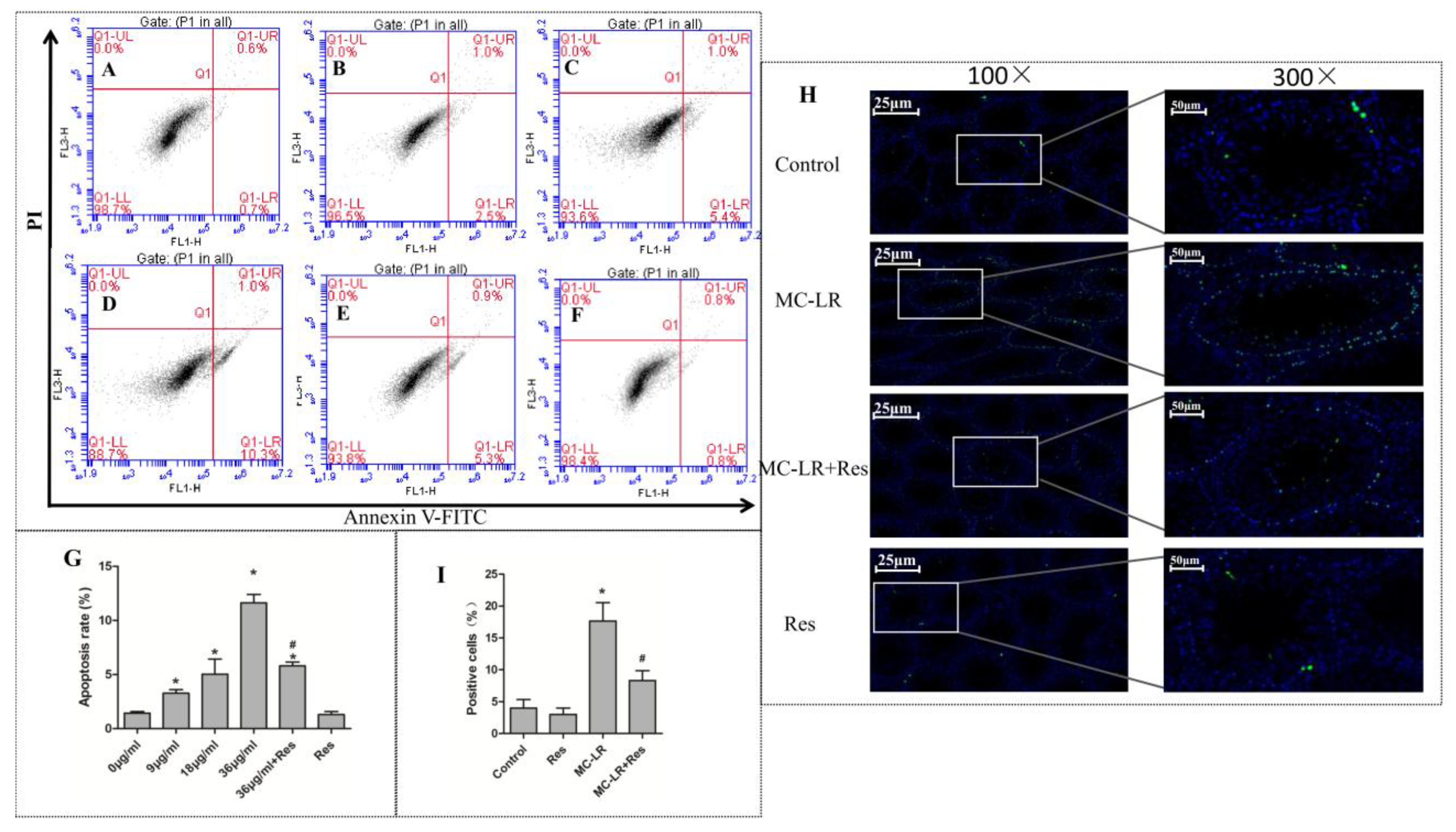
2.2. Protective Effect of RES on MC-LR-Induced Testicular Cell Apoptosis
2.3. Effect of RES on MC-LR-Induced Mitochondrial Membrane Collapse
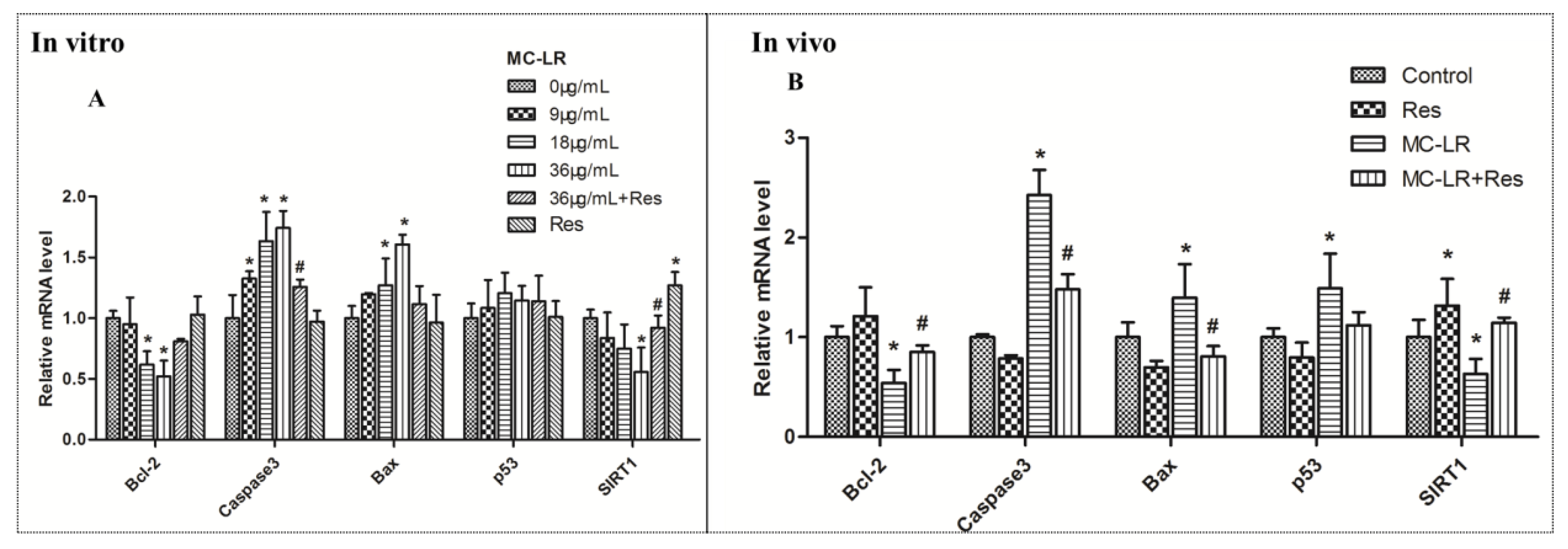
2.4. mRNA Levels of SIRT1/p53 Pathway Markers
2.5. RES Increased SIRT1 Protein Expression and Mediated p53-Related Apoptotic Protein Expression
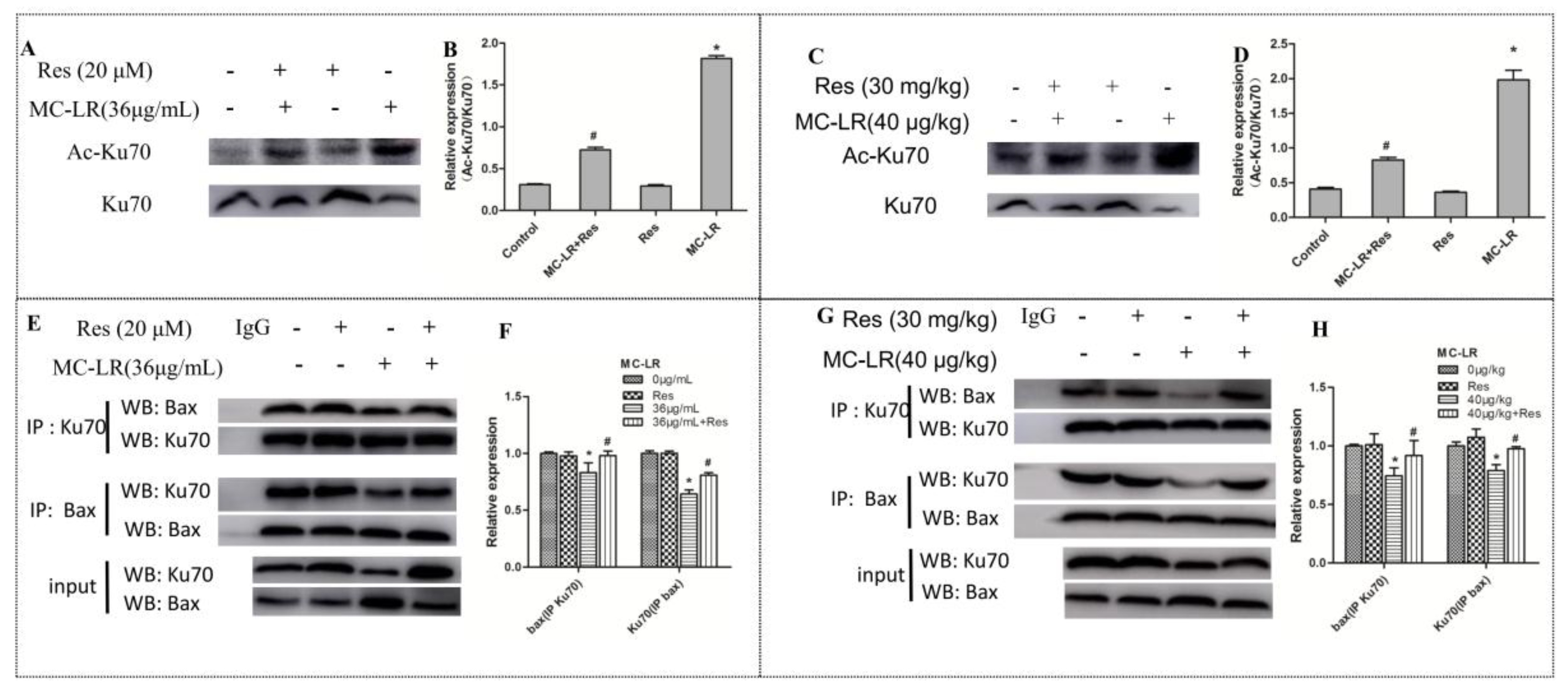
2.6. The Level of Acetylated Ku70 and Ku70–Bax Binding in the Co-Culture of Sertoli–Germ Cells and Testicular Tissues
2.7. Effect of RES on MC-LR-Induced Pathological Change
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Isolation and Identification of Sertoli–Germ Cells
4.3. Cell Viability Assay
4.4. Apoptosis Assay
4.5. Measurement of Mitochondrial Membrane Potential
4.6. Western Blot
4.7. Real-Time PCR
4.8. Animal Treatment
4.9. Hematoxylin-Eosin Staining
4.10. TUNEL Assay
4.11. Histology and Immunohistochemistry
4.12. Co-Immunoprecipitation Analysis
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from microcystis cawbg11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Hoeger, S.J.; Hitzfeld, B.C.; Dietrich, D.R. Occurrence and elimination of cyanobacterial toxins in drinking water treatment plants. Toxicol. Appl. Pharmacol. 2005, 203, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Giesy, J.P.; Xie, P. The dose makes the poison. Sci. Total Environ. 2017, 621, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Woolbright, B.L.; Williams, C.D.; Ni, H.; Kumer, S.C.; Schmitt, T.; Kane, B.; Jaeschke, H. Microcystin-LR induced liver injury in mice and in primary human hepatocytes is caused by oncotic necrosis. Toxicon 2017, 125, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Milutinovic, A.; Zorc-Pleskovic, R.; Petrovic, D.; Zorc, M.; Suput, D. Microcystin-LR induces alterations in heart muscle. Folia Biol. 2006, 52, 116–118. [Google Scholar]
- Chen, C.; Liu, W.; Wang, L.; Li, J.; Chen, Y.; Jin, J.; Kawan, A.; Zhang, X. Pathological damage and immunomodulatory effects of zebrafish exposed to microcystin-LR. Toxicon 2016, 118, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Xu, L.; Zhang, X.; Wang, Y.; Meng, X.; Miao, A.; Yang, L. Endoplasmic reticulum stress in murine liver and kidney exposed to microcystin-LR. Toxicon 2010, 56, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shao, S.; Zhou, F.; Wen, S.; Chen, F.; Han, X. Reproductive toxicity on female mice induced by microcystin-LR. Environ. Toxicol. Pharmacol. 2014, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yuan, J.; Wu, J.; Han, X. The toxic effects of microcystin-LR on rat spermatogonia in vitro. Toxicol. Lett. 2012, 212, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xiang, Z.; Li, D.; Han, X. Regulation of microcystin-LR-induced toxicity in mouse spermatogonia by mir-96. Environ. Sci. Technol. 2014, 48, 6383–6390. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Y.; Wang, J.; Wang, L.; Xiang, Z.; Li, D.; Han, X. Microcystin-leucine arginine causes cytotoxic effects in sertoli cells resulting in reproductive dysfunction in male mice. Sci. Rep. 2016, 6, 39238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, C.; Li, Y.; Imam, M.U.; Huang, H.; Liu, H.; Xin, Y.; Zhang, H. Novel role of ER stress and autophagy in microcystin-LR induced apoptosis in Chinese hamster ovary cells. Front. Phys. 2016, 7, 527. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Luo, J. Sirt1 and p53, effect on cancer, senescence and beyond. Biochim. Biophys. Acta 2010, 1804, 1684–1689. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, I.; Boussabbeh, M.; Pires Da Silva, J.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Sirt1 protects cardiac cells against apoptosis induced by zearalenone or its metabolites alpha- and beta-zearalenol through an autophagy-dependent pathway. Toxicol. Appl. Pharmacol. 2017, 314, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Wu, L.Y.; Zhao, L.; Sun, L.N.; Liu, H.Y.; Liu, G.; Guan, G.J. Sirt1 activator ameliorates the renal tubular injury induced by hyperglycemia in vivo and in vitro via inhibiting apoptosis. Biomed. Pharmacother. 2016, 83, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, H.; Dessain, S.K.; Ng Eaton, E.; Imai, S.I.; Frye, R.A.; Pandita, T.K.; Guarente, L.; Weinberg, R.A. Hsir2(sirt1) functions as an nad-dependent p53 deacetylase. Cell 2001, 107, 149–159. [Google Scholar] [CrossRef]
- Reed, S.M.; Quelle, D.E. P53 acetylation: Regulation and consequences. Cancers 2014, 7, 30–69. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Amariglio, N.; Rechavi, G.; Jakob-Hirsch, J.; Kela, I.; Kaminski, N.; Getz, G.; Domany, E.; Givol, D. DNA microarrays identification of primary and secondary target genes regulated by p53. Oncogene 2001, 20, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Krajewski, S.; Krajewska, M.; Wang, H.G.; Lin, H.K.; Liebermann, D.A.; Hoffman, B.; Reed, J.C. Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo. Oncogene 1994, 9, 1799–1805. [Google Scholar] [PubMed]
- Yan, S.; Wang, M.; Zhao, J.; Zhang, H.; Zhou, C.; Jin, L.; Zhang, Y.; Qiu, X.; Ma, B.; Fan, Q. Microrna-34a affects chondrocyte apoptosis and proliferation by targeting the sirt1/p53 signaling pathway during the pathogenesis of osteoarthritis. Int. J. Mol. Med. 2016, 38, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, M.; Wang, A.; Xu, M.; Wang, C.; Xu, G.; Zhang, B.; Zou, X.; Zhuge, Y. Tweak increases sirt1 expression and promotes p53 deacetylation affecting human hepatic stellate cell senescence. Cell Biol. Int. 2017, 41, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Zhu, X.; He, W.; Zhao, S.; Zhao, W.; Zhu, C. Resveratrol inhibits the hydrogen dioxide-induced apoptosis via sirt 1 activation in osteoblast cells. Biosci. Biotechnol. Biochem. 2015, 79, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Lieber, M.R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.Y.; Lavu, S.; Bitterman, K.J.; Hekking, B.; Imahiyerobo, T.A.; Miller, C.; Frye, R.; Ploegh, H.; Kessler, B.M.; Sinclair, D.A. Acetylation of the c terminus of ku70 by cbp and pcaf controls bax-mediated apoptosis. Mol. Cell 2004, 13, 627–638. [Google Scholar] [CrossRef]
- Hada, M.; Kwok, R.P. Regulation of ku70-bax complex in cells. J. Cell Death 2014, 7, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, C.; Opipari, A.W., Jr.; Bian, X.; Castle, V.P.; Kwok, R.P. Ku70 acetylation mediates neuroblastoma cell death induced by histone deacetylase inhibitors. Proc. Natl. Acad. Sci. USA 2005, 102, 4842–4847. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Juhn, K.; Lee, H.; Kim, S.H.; Min, B.H.; Lee, K.M.; Cho, M.H.; Park, G.H.; Lee, K.H. Sirt1 promotes DNA repair activity and deacetylation of ku70. Exp. Mol. Med. 2007, 39, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Oberdoerffer, P.; Michan, S.; McVay, M.; Mostoslavsky, R.; Vann, J.; Park, S.K.; Hartlerode, A.; Stegmuller, J.; Hafner, A.; Loerch, P.; et al. Sirt1 redistribution on chromatin promotes genomic stability but alters gene expression during aging. Cell 2008, 135, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Schoonjans, K.; Auwerx, J. Sirtuin functions in health and disease. Mol. Endocrinol. 2007, 21, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Nikolaev, A.Y.; Imai, S.; Chen, D.; Su, F.; Shiloh, A.; Guarente, L.; Gu, W. Negative control of p53 by sir2alpha promotes cell survival under stress. Cell 2001, 107, 137–148. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Zhang, D.; Lei, H. In situ studies on the distribution patterns and dynamics of microcystins in a biomanipulation fish--bighead carp (aristichthys nobilis). Environ. Pollut. 2007, 147, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Carmichael, W.; Yu, M.; Li, S. Cyclic peptide hepatotoxins from freshwater cyanobacterial (biue-green algae) waterblooms collected in central China. Environ. Toxicol. Chem. 1991, 10, 313–321. [Google Scholar] [CrossRef]
- Zhang, D.; Xie, P.; Liu, Y.; Qiu, T. Transfer, distribution and bioaccumulation of microcystins in the aquatic food web in lake taihu, china, with potential risks to human health. Sci. Total Environ. 2009, 407, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, C.; Fu, X.; Zhang, S.; Xin, Y.; Li, Y.; Xue, L.; Cheng, X.; Zhang, H. Microcystin-LR induced apoptosis in rat sertoli cells via the mitochondrial caspase-dependent pathway: Role of reactive oxygen species. Front. Phys. 2016, 7, 397. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.; Adhikari, N.; Saxena, D.K. Effect of endosulfan on the enzymes of polyol pathway in rat sertoli-germ cell coculture. Bull. Environ. Contam. Toxicol. 2001, 67, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Saxena, D.K.; Das, M. Effect of argemone oil and argemone alkaloid, sanguinarine on sertoli-germ cell coculture. Toxicol. Lett. 2009, 186, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Danz, E.D.; Skramsted, J.; Henry, N.; Bennett, J.A.; Keller, R.S. Resveratrol prevents doxorubicin cardiotoxicity through mitochondrial stabilization and the sirt1 pathway. Free Radic. Boil. Med. 2009, 46, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, X.; Geng, Z.; Zhou, Y.; Chen, Y.; Wu, J.; Han, X. Distribution of microcystin-LR to testis of male sprague-dawley rats. Ecotoxicology 2013, 22, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Huh, Y.J.; Cho, H.J.; Lee, B.; Park, J.; Hwang, D.Y.; Kim, D.W. Sirt1 enhances the survival of human embryonic stem cells by promoting DNA repair. Stem Cell Rep. 2017, 9, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Coussens, M.; Maresh, J.G.; Yanagimachi, R.; Maeda, G.; Allsopp, R. Sirt1 deficiency attenuates spermatogenesis and germ cell function. PLoS ONE 2008, 3, e1571. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting camp phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Abdelali, A.; Al-Bader, M.; Kilarkaje, N. Effects of trans-resveratrol on hyperglycemia-induced abnormal spermatogenesis, DNA damage and alterations in poly (adp-ribose) polymerase signaling in rat testis. Toxicol. Appl. Pharmacol. 2016, 311, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Anekonda, T.S.; Adamus, G. Resveratrol prevents antibody-induced apoptotic death of retinal cells through upregulation of sirt1 and ku70. BMC Res. Notes 2008, 1, 122. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, J.J.; Gu, Q.H.; An, J.; Cao, L.M.; Yang, H.P.; Hu, C.P. Egcg induces lung cancer a549 cell apoptosis by regulating ku70 acetylation. Oncol. Rep. 2016, 35, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, D.; Narayan, R.; Saxena, D.K.; Chowdhuri, D.K. Endosulfan induced cell death in sertoli-germ cells of male wistar rat follows intrinsic mode of cell death. Chemosphere 2014, 94, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Uguralp, S.; Usta, U.; Mizrak, B. Resveratrol may reduce apoptosis of rat testicular germ cells after experimental testicular torsion. Eur. J. Pediatr. Surg. 2005, 15, 333–336. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| SIRT1 | 5′-TCATTCTGACTGTGATGACGA-3′ | 5′-CTGCCACAGTGTCATATCCAA-3′ |
| P53 | 5′-CCCCTGAAGACTGGATAACTGTC-3′ | 5′-AACTCTGCAACATCCTGGGG-3′ |
| Bax | 5′-GAACCATCATGGGCTGGACA-3′ | 5′-GTGAGTGAGGCAGTGAGGAC-3′ |
| Bcl-2 | 5′-CTGAACCGGCATCTGCACAC-3′ | 5′-GCAGGTCTGCTGACCTCACT-3′ |
| Caspase-3 | 5′-GACTGCGGTATTGAGACAGA-3′ | 5′-CGAGTGAGGATGTGCATGAA-3′ |
| GAPDH | 5′-GGCACAGTCAAGGCTGAGAATG-3′ | 5′-ATGGTGGTGAAGACGCCAGTA-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Zhang, S.; Liu, C.; Wu, J.; Wang, Y.; Yuan, L.; Du, X.; Wang, R.; Marwa, P.W.; Zhuang, D.; et al. Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation. Toxins 2018, 10, 235. https://doi.org/10.3390/toxins10060235
Liu H, Zhang S, Liu C, Wu J, Wang Y, Yuan L, Du X, Wang R, Marwa PW, Zhuang D, et al. Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation. Toxins. 2018; 10(6):235. https://doi.org/10.3390/toxins10060235
Chicago/Turabian StyleLiu, Haohao, Shenshen Zhang, Chuanrui Liu, Jinxia Wu, Yueqin Wang, Le Yuan, Xingde Du, Rui Wang, Phelisters Wegesa Marwa, Donggang Zhuang, and et al. 2018. "Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation" Toxins 10, no. 6: 235. https://doi.org/10.3390/toxins10060235
APA StyleLiu, H., Zhang, S., Liu, C., Wu, J., Wang, Y., Yuan, L., Du, X., Wang, R., Marwa, P. W., Zhuang, D., Cheng, X., & Zhang, H. (2018). Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation. Toxins, 10(6), 235. https://doi.org/10.3390/toxins10060235