Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins


 , , ,
, , ,  ,
, 
 , , , , ,
, , , , ,  , ,
, ,  , ,
, ,  , , ,
, , ,  , ,
, , 
 ,
,  , , ,
, , ,  , , ,
, , ,  , ,
, ,  ,
,  ,
,  , , , ,
, , , ,  ,
, 
 , , ,
, , , 
 ,
,  ,
,  , ,
, ,  , , , , ,
, , , , ,  , ,
, ,  ,
, 

 , ,
, ,  , ,
, ,  , , , , , , , , and
, , , , , , , , and
Abstract
1. Introduction
2. Results
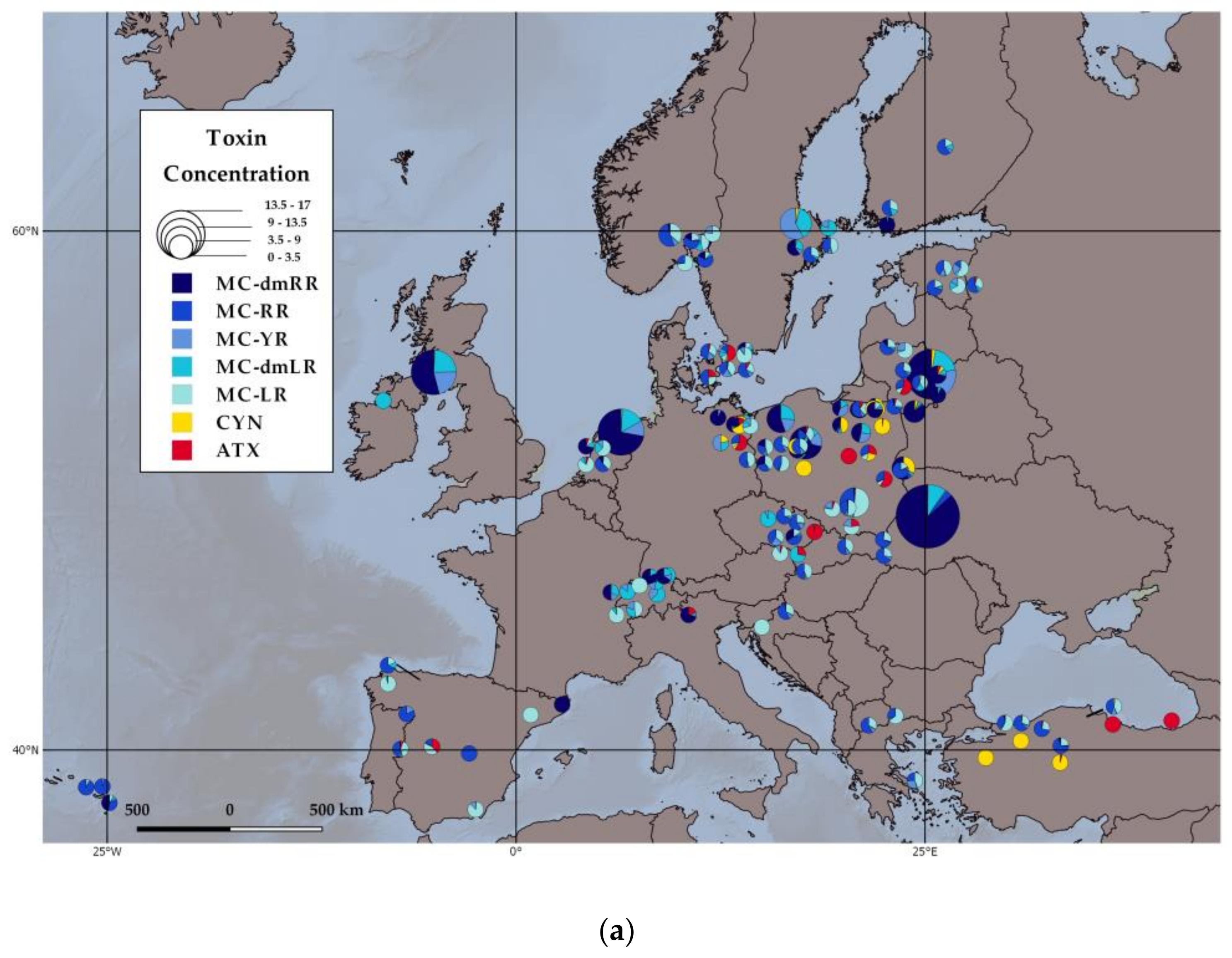
2.1. Toxin Distribution on a Continental Scale
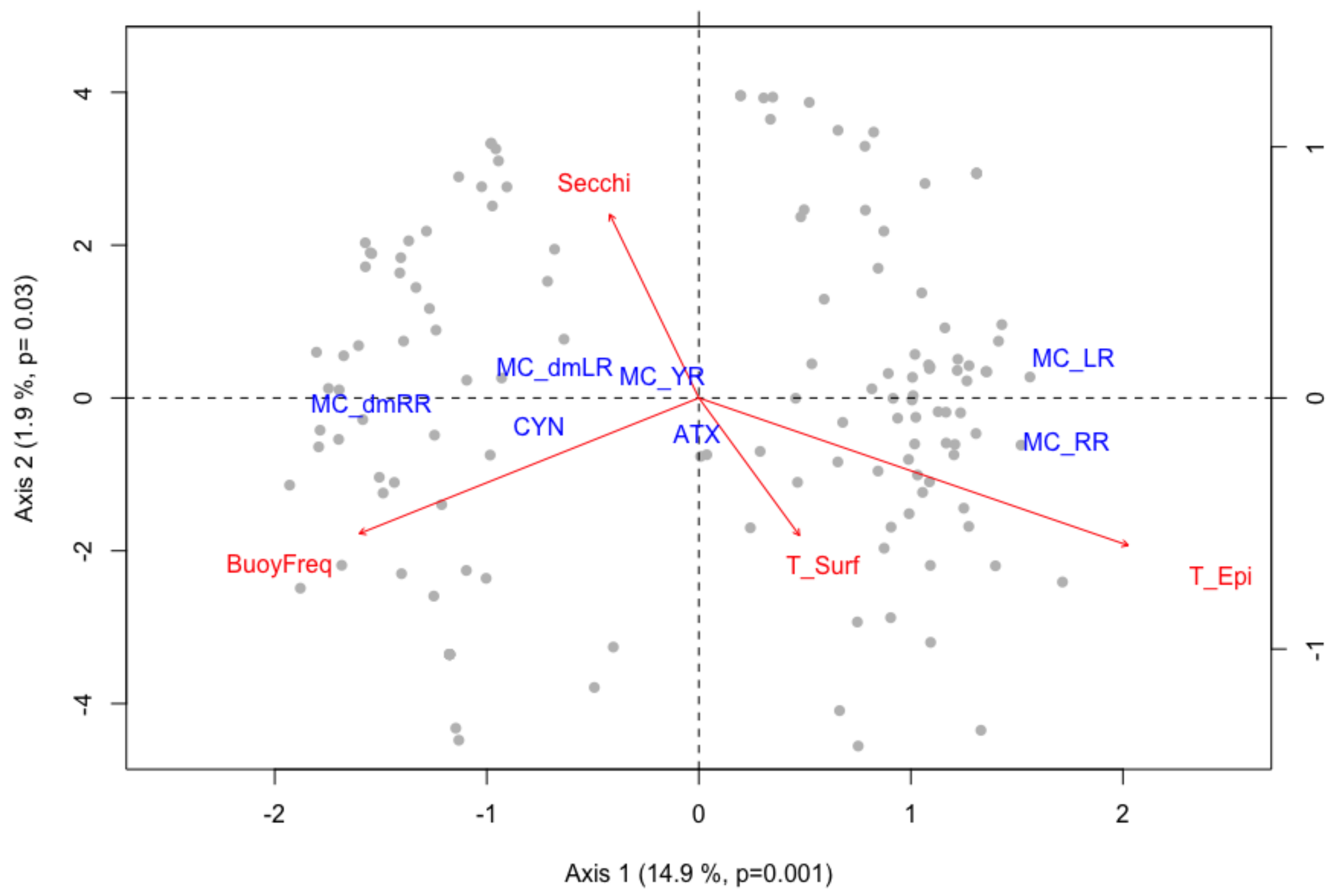
2.2. Multivariate Multiple Regression Analysis
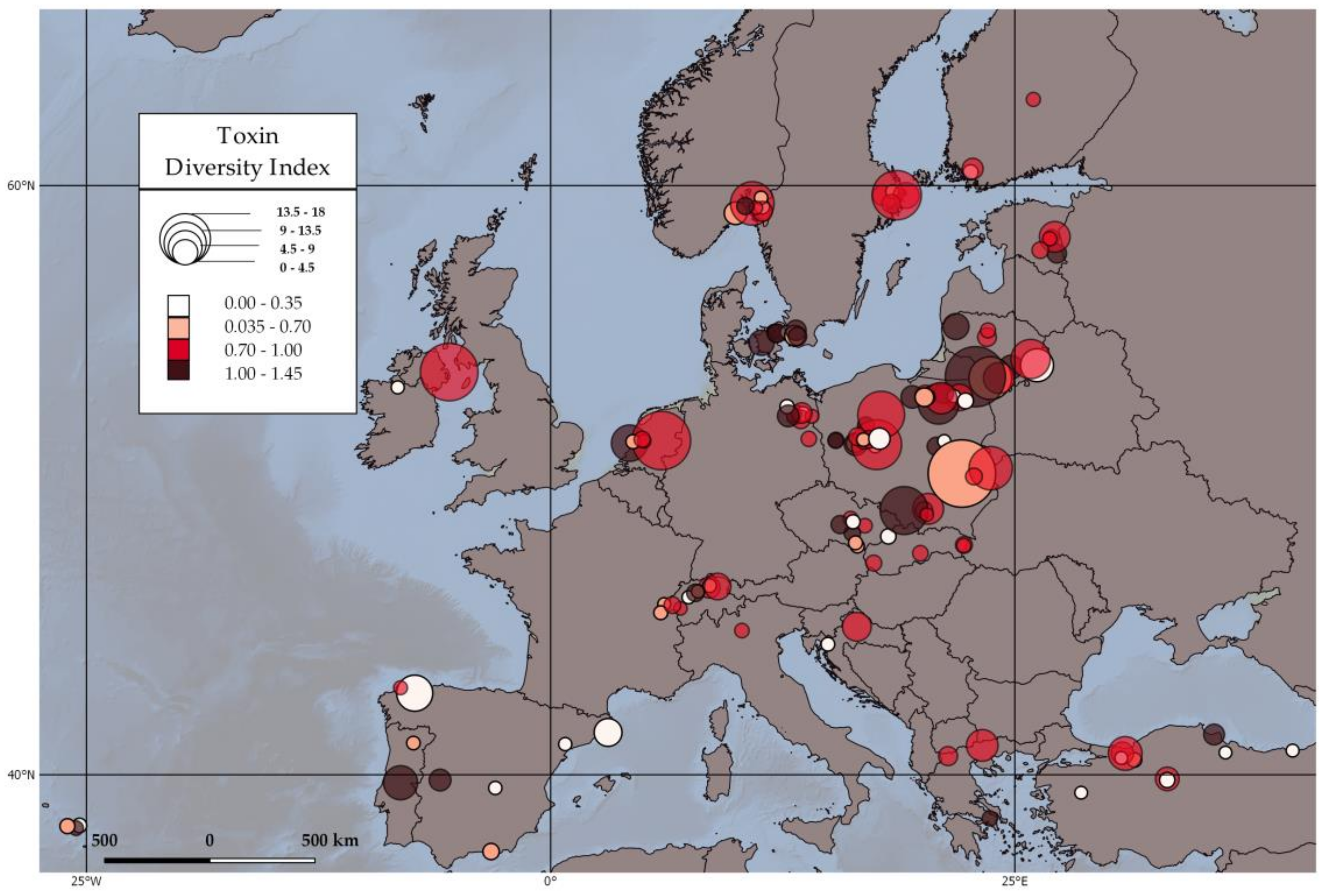
2.3. Toxin Diversity Index and Environmental Parameters
3. Discussion
4. Materials and Methods
4.1. Sampling Survey
4.2. Cyanotoxin Analysis
4.2.1. Microcystins (MCs) and Nodularin (NOD) Analysis
4.2.2. Cylindrospermopsin (CYN) and Anatoxin (ATX) Analysis
4.3. Nutrient Analysis
4.4. Pigment Analysis
4.5. Response Variables and Environmental Parameters
4.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Codd, G.A.; Meriluoto, J.; Metcalf, J.S. Introduction: Cyanobacteria, Cyanotoxins, Their Human Impact, and Risk Management. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Huisman, J.M.; Matthijs, H.C.P.; Visser, P.M. Harmful Cyanobacteria. Springer Aquatic Ecology Series 3; Springer: Dordrecht, The Netherlands, 2005; p. 243. [Google Scholar]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and Phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H.W. Spatiotemporal patterns and ecophysiology of toxigenic Microcystis blooms in Lake Taihu, China: Implications for water quality management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- United Nations Environment Programme (UNEP). Loss and Damage: The role of Ecosystem Services. United Nations Environment Programme; UNEP: Nairobi, Kenya, 2016. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., White, L.L., Eds.; IPCC: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.; De Meester, L.; et al. Allied attack: Climate change and eutrophication. Inland Waters 2011, 1, 101–105. [Google Scholar] [CrossRef]
- Lurling, M.; van Oosterhout, F.; Faassen, E. Eutrophication and Warming Boost Cyanobacterial Biomass and Microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef]
- Schopf, J.W. The Fossil Record: Tracing the Roots of the Cyanobacterial Lineage. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 13–35. [Google Scholar]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef] [PubMed]
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the key ecological traits of cyanobacteria as a basis for their management and control in changing lakes. Aquat. Ecol. 2015, 50, 333–350. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Mur, L.R.; Kinsman, R.; Walsby, A.E. Microcystis changes its buoyancy in response to the average irradiance in the surface mixed layer. Archiv für Hydrobiologie 1991, 120, 385–401. [Google Scholar]
- Granf, G.G.; Oliver, R.L. Vertical separation of light and available nutrients as a factor causing replacement of green-algae by blue-green algae in the plankton of a stratified lake. J. Ecol. 1982, 70, 829–844. [Google Scholar] [CrossRef]
- Arnaud, C.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Hubert, J.F.; Fastner, J. Ecology of Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Kokociński, M.; Cameán, A.M.; Carmeli, S.; Guzmán-Guillén, R.; Jos, Á.; Mankiewicz-Boczek, J.; Metcalf, J.S.; Moreno, I.M.; Prieto, A.I.; et al. Cylindrospermopsin and Congeners. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, L.S.J., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Nogueira, I.C.; Saker, M.L.; Pflugmacher, S.; Wiegand, C.; Vasconcelos, V.M. Toxicity of the cyanobacterium Cylindrospermopsis raciborskii to Daphnia magna. Environ. Toxicol. 2004, 19, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.J. Effect of food availability on the response of planktonic rotifers to a toxic strain of the cyanobacterium Anabaena flos-aquae. Limnol. Oceanogr. 1996, 41, 1565–1572. [Google Scholar] [CrossRef]
- DeMott, W.R.; Zhang, Q.X.; Carmichael, W.W. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Oceanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef]
- Holland, A.; Kinnear, S. Interpreting the possible ecological role(s) of cyanotoxins: Compounds for competitive advantage and/or physiological aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef] [PubMed]
- Capelli, C.; Ballot, A.; Cerasino, L.; Papini, A.; Salmaso, N. Biogeography of bloom-forming microcystin producing and non-toxigenic populations of Dolichospermum lemmermannii (Cyanobacteria). Harmful Algae 2017, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Meriluoto, J.L.; Spoof, L.; Codd, G. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Hoboken, NJ, USA, 2017. [Google Scholar]
- Bortoli, S.; Oliveira-Silva, D.; Krüger, T.; Dörr, F.A.; Copelicolo, P.; Volmer, D.A.; Pinto, E. Growth and microcystin production of a Brazilian Microcystis aeruginosa strain (LTPNA 02) under different nutrient conditions. Revista Brasileira de Farmacognosia 2014, 24, 389–398. [Google Scholar] [CrossRef]
- Cerasino, L.; Capelli, C.; Salmaso, N. A comparative study of the metabolic profiles of common nuisance cyanobacteria in southern perialpine lakes. Adv. Oceanogr. Limnol. 2017, 8. [Google Scholar] [CrossRef]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K. Effects of light, temperature, nitrate, orthophosphate and bacteria on growth of hepatotoxin production by Oscillatoria agardhii strains. Appl. Environ. Microbiol. 1990, 56, 2658–2666. [Google Scholar] [PubMed]
- Long, B.M.; Jones, G.J.; Orr, P.T. Cellular microcystin content in N-Limited Microcystis aeruginosa can be predicted from growth rate. Appl. Environ. Microbiol. 2001, 67, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Sivonen, K.; Luukkainen, R.; Niemelä, S.I. Anatoxin-a concentration in Anabaena and Aphanizomenon under different environmental conditions and comparison of growth by toxic and non-toxic Anabaena-strains: A laboratory study. J. Appl. Phycol. 1993, 5, 581–591. [Google Scholar] [CrossRef]
- Rapala, J.; Sivonen, K. Assessment of environmental conditions that favor hepatotoxic and neurotoxic Anabaena spp. strains cultured under light limitation and different temperatures. Microb. Ecol. 2008, 36, 181–192. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Backerb, L.C.W.; Kardinaa, E.A.; Chorus, I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae 2014, 40, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R.; Choice, A.; Hosja, W. Toxicity of edible mussels (Mytilus edulis) growing naturally in an estuary during a water bloom of the blue-green alga Nodularia spumigena. Environmental Toxicology and Water Quality. Environ. Toxicol. 1992, 7, 119–123. [Google Scholar]
- Fawell, J. Toxins from Blue-Green Algae: Toxicological Assessment of Microcystin-LR; W.R. Centre: Medmenham, UK, 1993; pp. 1–259. [Google Scholar]
- Wolf, H.U.; Frank, C. Toxicity assessment of cyanobacterial toxin mixtures. Environ. Toxicol. 2002, 17, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Lurling, M. Occurrence of the microcystins MC-LW and MC-LF in Dutch surface waters and their contribution to total microcystin toxicity. Mar. Drugs 2013, 11, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water—A Guide to Their Public Health Consequences, Monitoring and Management; Spon Press: London, UK, 1999. [Google Scholar]
- Ibelings, B.; Havens, K. Cyanobacterial toxins: A qualitative meta-analysis of concentrations, dosage and effects in freshwater, estuarine and marine biota. Adv. Exp. Med. Biol. 2008, 619, 675–732. [Google Scholar] [PubMed]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.; Lehmann, S.; Meyer, M.T.; Dietze, J.E.; Griffith, C. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I. Current Approaches to Cyanotoxin Risk Assessment, Risk Management and Regulations in Different Countries; Chorus, I., Ed.; Federal Environmental Agency (Umweltbundesamt): Dessau-Roßlau, Germany, 2005. [Google Scholar]
- Beniston, M.; Stephenson, D.B.; Christensen, O.B.; Ferro, C.A.T.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Chang. 2007, 81, 71–95. [Google Scholar] [CrossRef]
- Vautard, R.; Gobiet, A.; Sobolowski, S.; Kjellström, E.; Stegehuis, A.; Watkiss, P.; Mendlik, T.; Landgren, O.; Nikulin, G.; Teichmann, C.; et al. The European climate under a 2 °C global warming. Environ. Res. Lett. 2014, 9, 034006. [Google Scholar] [CrossRef]
- Hoy, A.; Hänsel, S.; Skalak, P.; Ustrnul, Z.; Bochnicek, O. The extreme European summer of 2015 in a secular perspective. Int. J. Climatol. 2016, 37, 1–34. [Google Scholar]
- Messineo, V.; Melchiorre, S.; Di Corcia, A.; Gallo, P.; Bruno, M. Seasonal succession of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum blooms with cylindrospermopsin occurrence in the volcanic Lake Albano, Central Italy. Environ. Toxicol. 2010, 25, 18–27. [Google Scholar] [PubMed]
- Salmaso, N.; Cerasino, L.; Boscaini, A.; Capelli, C. Planktic Tychonema (Cyanobacteria) in the large lakes south of the Alps: Phylogenetic assessment and toxigenic potential. FEMS Microbiol. Ecol. 2016, 92, fiw155. [Google Scholar] [CrossRef] [PubMed]
- Pawlik-Skowrońska, B.; Kalinowska, R.; Skowroński, T. Cyanotoxin diversity and food web bioaccumulation in a reservoir with decreasing phosphorus concentrations and perennial cyanobacterial blooms. Harmful Algae 2013, 28, 118–125. [Google Scholar] [CrossRef]
- Straile, D.; Jochimsen, M.C.; Kümmerlin, R. The use of long-term monitoring data for studies of planktonic diversity: A cautionary tale from Swiss lakes. Freshw. Biol. 2013, 58, 1292–1301. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A. Trait-Based Community Ecology of Phytoplankton. Ann. Rev. Ecol. Evolut. Syst. 2008, 39, 615–639. [Google Scholar] [CrossRef]
- Loftin, K.A.; Dietze, J.E.; Meyer, M.T.; Graham, J.L.; Maksimowicz, M.M.; Toyne, K.D. Total Cylindrospermopsins, Microcystins/Nodularins, and Saxitoxins Data for the 2007 United States Envrionmnetal Protection Agency National Lake Assessment; Kansas Water Science Center, U.S. Geological Survey: Lawrence, KS, USA, 2016.
- Kurmayer, R.; Sivonen, K.; Salmaso, N. Introduction. In Molecular Tools for the Detection and Quantification of Toxigenic Cyanobacteria; Kurmayer, R., Sivonen, K., Wilmotte, A., Salmaso, N., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. PNAS 2003, 101, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Climate 2008, 320, 57–58. [Google Scholar]
- Visser, P.M.; Ibelings, B.W.; Mur, L.R.; Walsby, A.E. The ecophysiology of the harmful cyanobacterium Microcystis. In Harmful Cyanobacteria; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Jang, M.H.; Jung, J.M.; Takamura, N. Changes in microcystin production in cyanobacteria exposed to zooplankton at different population densities and infochemical concentrations. Limnol. Oceanogr. 2007, 52, 1454–1466. [Google Scholar] [CrossRef]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of Light on the Microcystin Content of Microcystis Strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Maier, M.Y.; Puddick, J.; Pochon, X.; Zaiko, A.; Dietrich, D.R.; Hamilton, D.P. Trophic state and geographic gradients influence planktonic cyanobacterial diversity and distribution in New Zealand lakes. FEMS Microbiol. Ecol. 2016, 93, fiw234. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bertram, J., Eds.; E & FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Horst, G.P.; Sarnelle, O.; White, J.D.; Hamilton, S.K.; Kaul, R.B.; Bressie, J.D. Nitrogen availability increases the toxin quota of a harmful cyanobacterium, Microcystis aeruginosa. Water Res. 2014, 54, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Nimptsch, J.; Woelfl, S.; Osorio, S.; Valenzuela, J.; Moreira, C.; Ramos, V.; Castelo-Branco, R.; Leão, P.N.; Vasconcelos, V. First record of toxins associated with cyanobacterial blooms in oligotrophic North Patagonian lakes of Chile- a genomic approach. Int. Rev. Hydrobiol. 2016, 101, 57–68. [Google Scholar] [CrossRef]
- Mischke, U. Cyanobacteria associations in shallow polytrophic lakes: Influence of environmental factors. Acta Oecol. 2003, 24, S11–S23. [Google Scholar] [CrossRef]
- Anneville, O.; Souissi, S.; Ibanez, F.; Ginot, V.; Druant, J.C.; Angeli, N. Temporal mapping of phytoplankton assemblages in Lake Geneva: Annual and interannual changes in their patterns of succession. Limnol. Oceanogr. 2002, 47, 1355–1366. [Google Scholar] [CrossRef]
- Anneville, O.; Souissi, S.; Gammeter, S.; Straile, D. Seasonal and inter-annual scales of variability in phytoplankton assemblages: Comparison of phytoplankton dynamics in three peri-alpine lakes over a period of 28 years. Freshw. Biol. 2004, 49, 98–115. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Peleato, M.L.; Fillat, M.F. Microcystin-LR synthesis as response to nitrogen: Transcriptional analysis of the mcyD gene in Microcystis aeruginosa PCC7806. Ecotoxicology 2010, 19, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Mitigating harmful cyanobacterial blooms in a human- and climatically-impacted world. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.K.; Litchman, E. Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Cerasino, L.; Salmaso, N. Diversity and distribution of cyanobacterial toxins in the Italian subalpine lacustrine district. Oceanol. Hydrobiol. Stud. 2012, 41, 54–63. [Google Scholar] [CrossRef]
- Pires, L.M.; Sarpe, D.; Brehm, M.; Ibelings, B.W. Potential synergistic effects of microcystins and bacterial lipopolysaccharides on life history traits of Daphnia galeata raised on low and high food levels. Aquat. Toxicol. 2011, 104, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Freitas, E.C.; Pinheiro, C.; Rocha, O.; Loureiro, S. Can mixtures of cyanotoxins represent a risk to the zooplankton? The case study of Daphnia magna Straus exposed to hepatotoxic and neurotoxic cyanobacterial extracts. Harmful Algae 2014, 31, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedzialek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J. Dog Poisonings associated with Microcystis aeruginosa Bloom in the Netherlands. Toxins 2013, 5, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Beulker, C.; Geiser, B.; Hoffmann, A.; Kröger, R.; Teske, K.; Hoppe, J.; Mundhenk, L.; Neurath, H.; Sagebiel, D.; et al. Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin. Toxins 2018, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Netherlands Normalization Institute (NEN). Water: Photometric Determination of the Content of Dissolved Orthophosphate and the Total Content of Phosphorous Compounds by Continuous Flow Analysis; NEN 6663; The Netherlands Normalization Institute: Delft, The Netherlands, 1986. (In Dutch) [Google Scholar]
- Netherlands Normalization Institute (NEN). Water: Photometric Determination of the Content of Ammonium Nitrogen and the Sum of the Contents of Ammoniacal and Organically Bound Nitrogen According to Kjeldahl by Continuous Flow Analysis; NEN-6646; Netherlands Normalization Institute: Delft, The Netherlands, 1990. [Google Scholar]
- Van der Staay, G.W.M.; Brouwer, A.; Baard, R.L.; van Mourik, F. Separation of photosystems I and II from the oxychlorobacterium (prochlorophyte) Prochlorothrix hollandica and association of Chlb binding antennae with PS II. Biochim. Biophys. Acta 1992, 1102, 220–228. [Google Scholar] [CrossRef]
- Leach, T.H.; Beisner, B.E.; Carey, C.C.; Pernica, P.; Rose, K.C.; Huot, Y.; Brentrup, J.A.; Domaizon, I.; Grossart, H.-P.; Ibelings, B.W.; et al. Patterns and drivers of deep chlorophyll maxima structure in 100 lakes: The relative importance of light and thermal stratification. Limnol. Oceanogr. 2017, 63, 628–646. [Google Scholar] [CrossRef]
- Winslow, L.A.; Read, J.S.; Woolway, R.I.; Brentrup, J.A.; Leach, T.H.; Zwart, J.A. rLakeAnalyzer: Lake Physics Tools. Zenodo. Available online: https://zenodo.org/record/1003169#.Ws82F9NubEY (accessed on 6 October 2017).
- Chen, C.T.A.; Millero, F.J. Thermodynamic properties for natural waters covering only the limnological range. Limnol. Oceanogr. 1986, 31, 657–662. [Google Scholar] [CrossRef]
- Scheffer, M.; Rinaldi, S.; Gragnani, A.; Mur, L.R.; van Nes, E.H. On the Dominance of Filamentous Cyanobacteria in Shallow, Turbid Lakes. Ecology 1997, 78, 272–282. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, H. The vegan package. Community Ecol. Package 2007, 10, 631–637. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-Plus, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Variant | Present (n Lakes) | Concentration Range (μg/L) | Limit of Quantification 1 (μg/L) | Mean | Stdv |
|---|---|---|---|---|---|
| MC-Tot | 127 | 0–17.18 | 1.20 | 2.70 | |
| MC-YR | 113 | 0–4.92 | 0.0050 | 0.14 | 0.56 |
| MC-dmLR | 108 | 0–3.16 | 0.0054 | 0.15 | 0.50 |
| MC-LR | 93 | 0–3.97 | 0.0086 | 0.20 | 0.55 |
| MC-RR | 67 | 0–3.31 | 0.0358 | 0.20 | 0.50 |
| ATX | 54 | 0–1.33 | 0.0004 | 0.03 | 0.12 |
| CYN | 53 | 0–2.01 | 0.0004 | 0.05 | 0.20 |
| MC-dmRR | 52 | 0–14.89 | 0.0489 | 0.52 | 1.83 |
| RDA | AdjR2 | Predictor | Variance | F | p |
|---|---|---|---|---|---|
| Toxin Concentrations | 0.14 | T_Epi | 0.05 | 13.22 | 0.001 |
| T_Surf | 0.02 | 4.93 | 0.002 | ||
| BuoyFreq | 0.01 | 3.17 | 0.01 | ||
| Secchi | 0.01 | 2.87 | 0.01 | ||
| Toxin Quota | 0.14 | T_Epi | 0.05 | 13.22 | 0.001 |
| T_Surf | 0.02 | 4.93 | 0.003 | ||
| BuoyFreq | 0.01 | 3.17 | 0.02 | ||
| Secchi | 0.01 | 2.87 | 0.02 |
| Index | GLM, Family = Negative Binomial | Predictor | Χ2 | p |
|---|---|---|---|---|
| TDIquota | −1.93 + 0.003 DMax + 0.03 Latitude ** + 0.03 T_Epi −24.2 BuoyFreq * − 0.06 Secchi | Latitude | 1.21 | 0.004 |
| BuoyFreq | 0.75 | 0.02 | ||
| DMax | 0.08 | 0.8 | ||
| T_Epi | 0.24 | 0.2 | ||
| Secchi | 0.30 | 0.15 | ||
| Richnessquota | −0.16 + 0.002 DMax + 0.02 Latitude ** + 0.04 T_Epi * −26.3 BuoyFreq *** − 0.09 Secchi ** | Latitude | 1.49 | 0.006 |
| BuoyFreq | 2.14 | 0.001 | ||
| DMax | 0.41 | 0.15 | ||
| T_Epi | 1.13 | 0.02 | ||
| Secchi | 1.40 | 0.007 |
| Toxin Quota | Response When TDI ↑ | Χ2 | p | Response When Richness ↑ | Χ2 | p |
|---|---|---|---|---|---|---|
| MC-YR | ↑ | 0.10 | 0.02 | ↑ | 0.10 | 0.01 |
| MC-dmLR | ↑ | 0.10 | 0.02 | ↑ | 0.06 | 0.06 |
| MC-LR | ↓ | 0.09 | 0.54 | ↑ | 0.3 | 0.2 |
| MC-RR | ↑ | 0.44 | 0.06 | ↑ ** | 0.90 | 0.003 |
| ATX | ↑ *** | 0.15 | 0.0002 | ↑ *** | 0.19 | <0.0001 |
| CYN | ↑ ** | 0.38 | 0.007 | ↑ *** | 0.56 | 0.0009 |
| MC-dmRR | ↑ | 0.2 | 0.85 | ↑ | 0.41 | 0.02 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantzouki, E.; Lürling, M.; Fastner, J.; De Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. https://doi.org/10.3390/toxins10040156
Mantzouki E, Lürling M, Fastner J, De Senerpont Domis L, Wilk-Woźniak E, Koreivienė J, Seelen L, Teurlincx S, Verstijnen Y, Krztoń W, et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins. 2018; 10(4):156. https://doi.org/10.3390/toxins10040156
Chicago/Turabian StyleMantzouki, Evanthia, Miquel Lürling, Jutta Fastner, Lisette De Senerpont Domis, Elżbieta Wilk-Woźniak, Judita Koreivienė, Laura Seelen, Sven Teurlincx, Yvon Verstijnen, Wojciech Krztoń, and et al. 2018. "Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins" Toxins 10, no. 4: 156. https://doi.org/10.3390/toxins10040156
APA StyleMantzouki, E., Lürling, M., Fastner, J., De Senerpont Domis, L., Wilk-Woźniak, E., Koreivienė, J., Seelen, L., Teurlincx, S., Verstijnen, Y., Krztoń, W., Walusiak, E., Karosienė, J., Kasperovičienė, J., Savadova, K., Vitonytė, I., Cillero-Castro, C., Budzyńska, A., Goldyn, R., Kozak, A., ... Ibelings, B. W. (2018). Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins, 10(4), 156. https://doi.org/10.3390/toxins10040156