Protective Effect of N-Acetylcysteine against Oxidative Stress Induced by Zearalenone via Mitochondrial Apoptosis Pathway in SIEC02 Cells

 and
and
Abstract
1. Introduction
2. Results
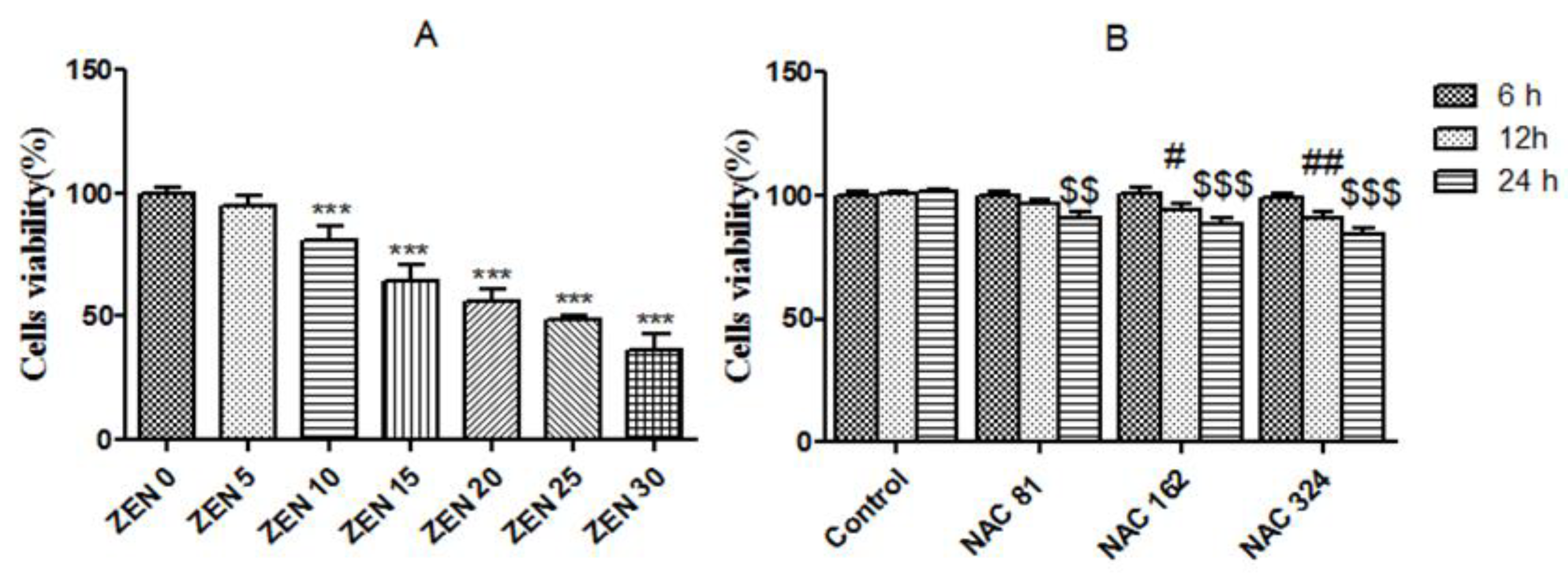
2.1. Effects of ZEN and NAC on Cell Viability
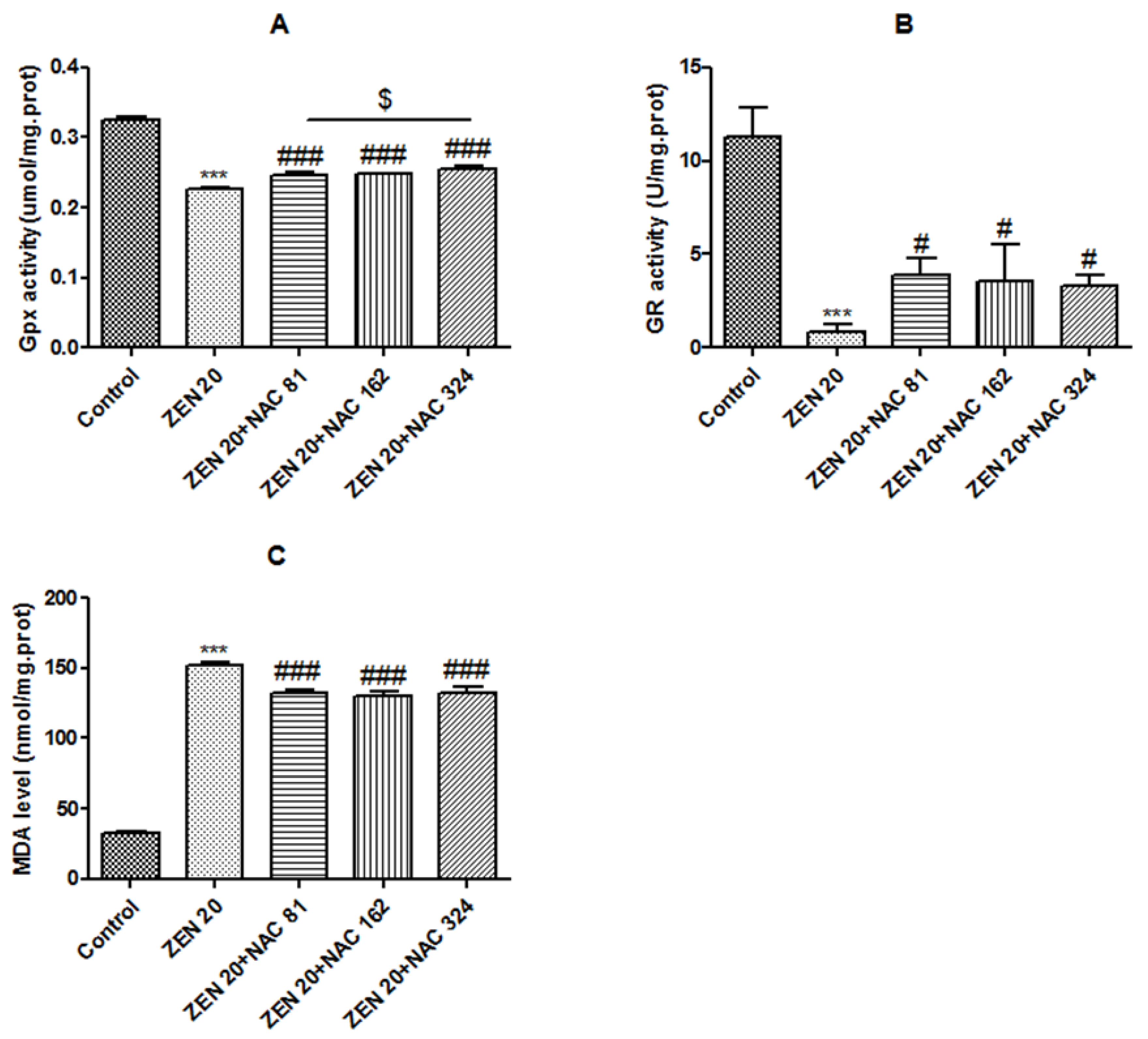
2.2. Effects of ZEN and NAC on Oxidative Stress
2.2.1. Glutathione peroxidase (Gpx) Activity
2.2.2. Glutathione reductase (GR) Activity
2.2.3. Malondialdehyde (MDA) Level
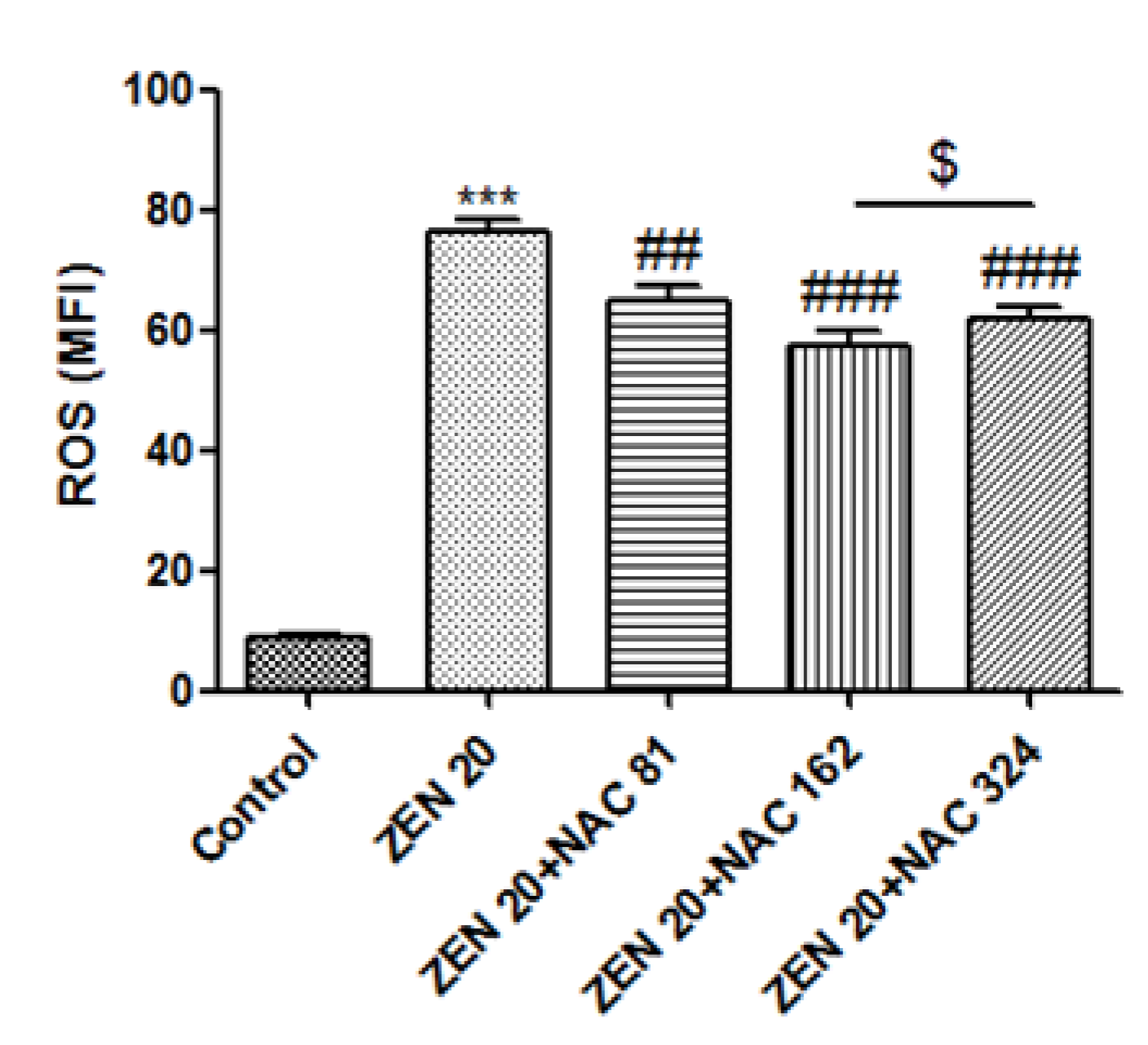
2.3. Effects of ZEN and NAC on Intracellular ROS Generation
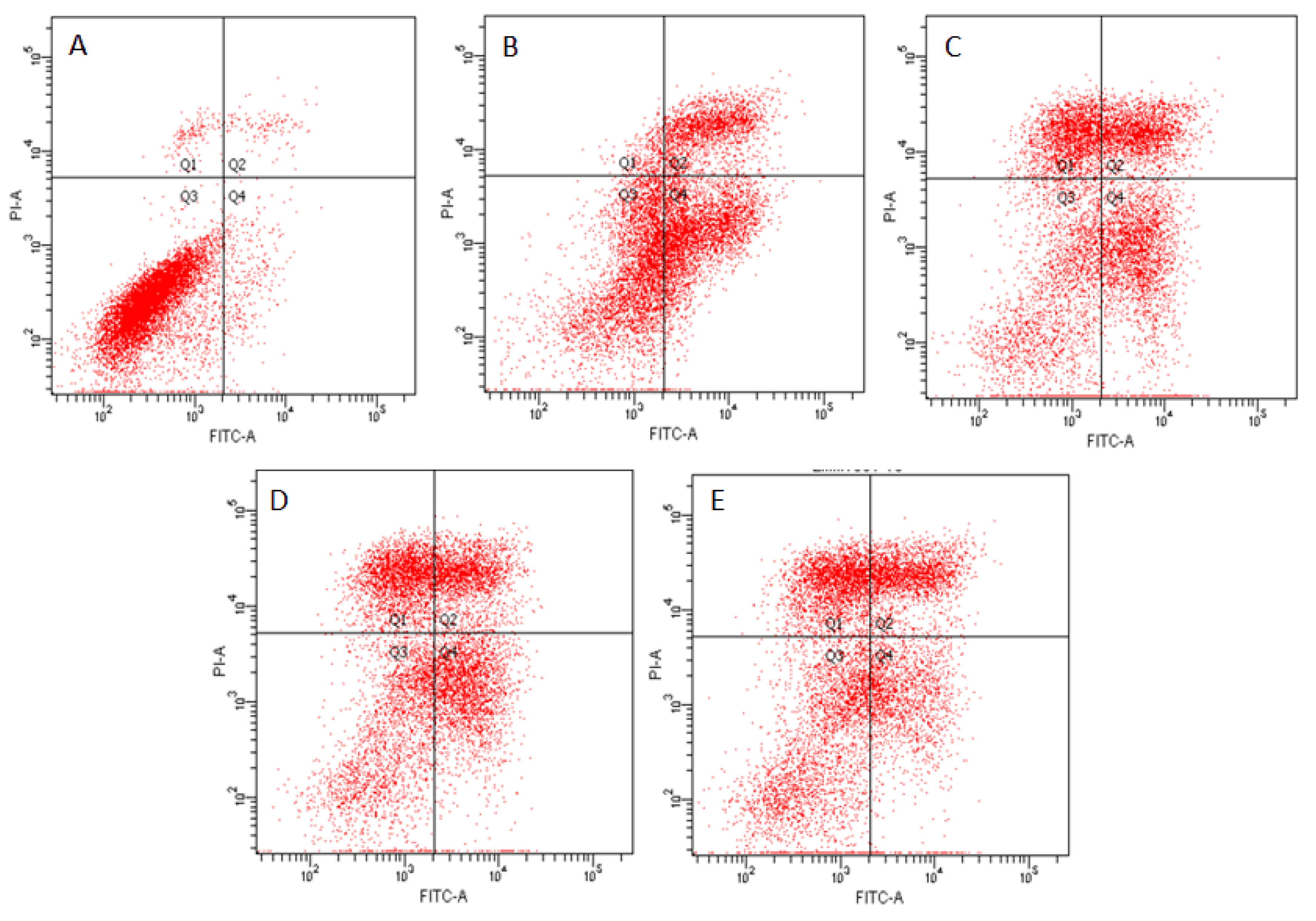
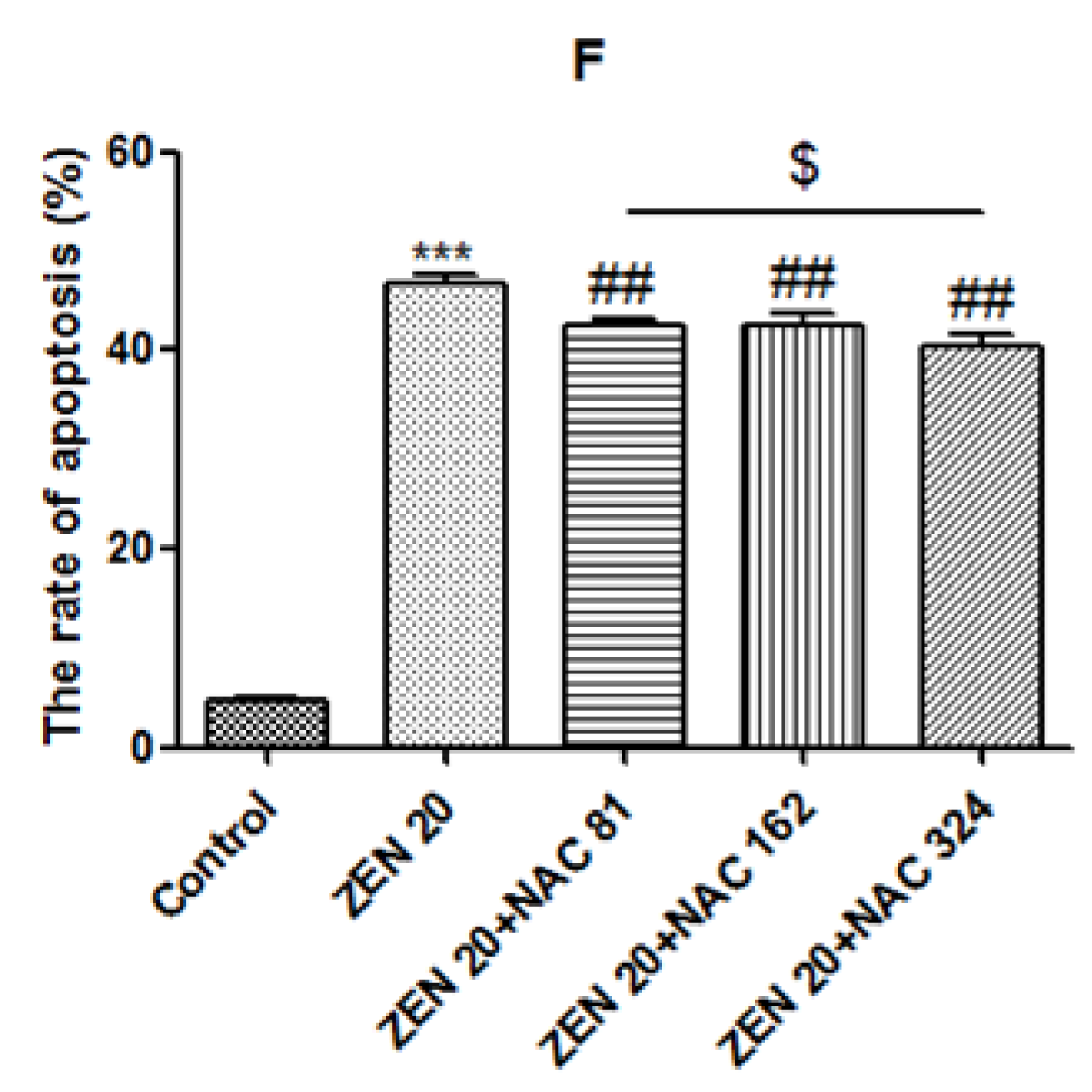
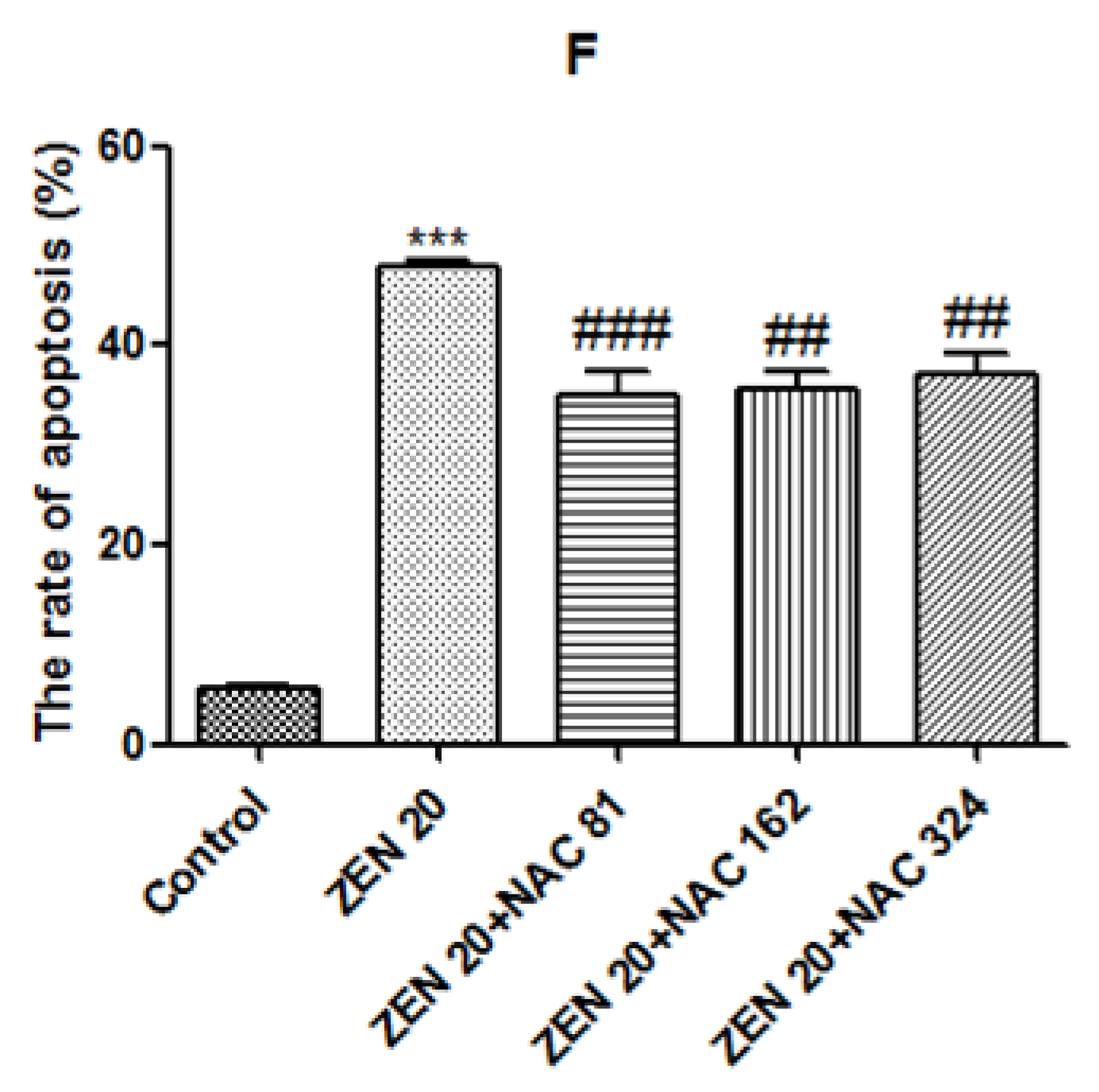
2.4. Effects of ZEN and NAC on Apoptosis
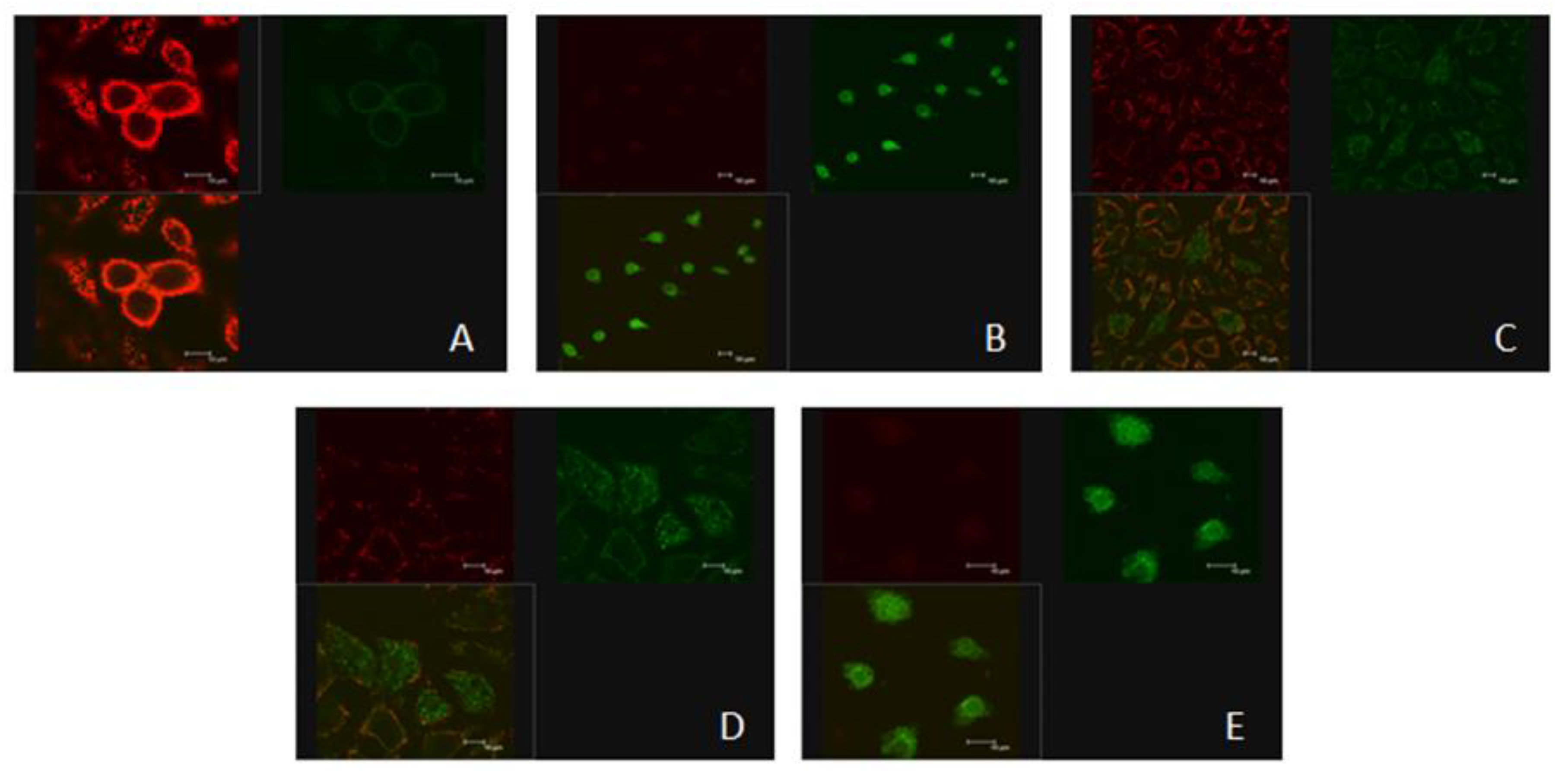
2.5. Effects of ZEN and NAC on the Change of ΔΨm
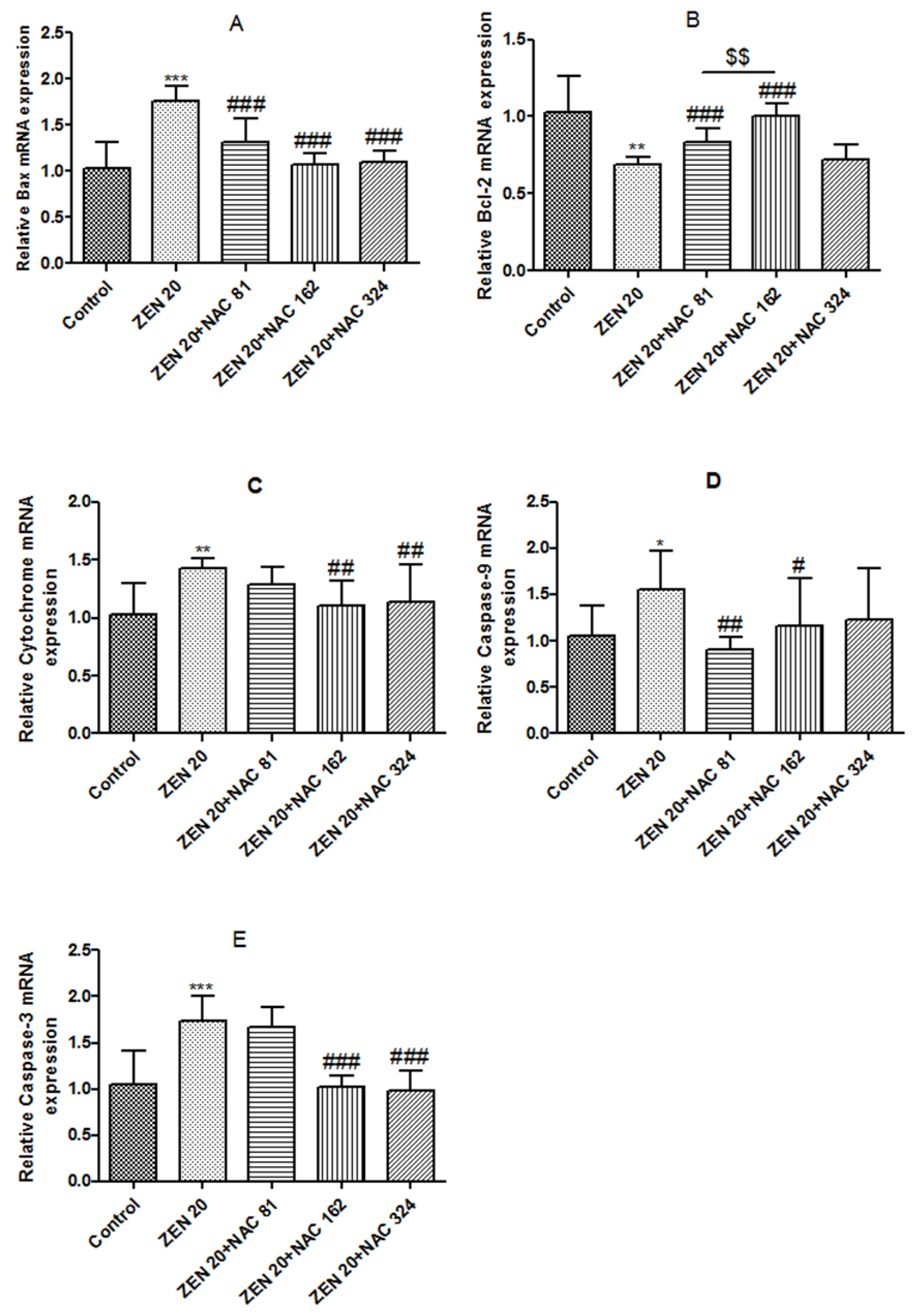
2.6. Effects of ZEN and NAC on Apoptosis-Related mRNA Expression
2.6.1. Bax
2.6.2. Bcl-2
2.6.3. Cytochrome c (Cyto c)
2.6.4. Caspase-9 and Caspase-3
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture and Conditions
5.3. Cell Viability Assay
5.4. Experiment Design
5.5. Determination of Antioxidant Enzyme Activity and Oxidative Products
5.6. Detection of ROS Generation
5.7. Apoptosis Detection
5.7.1. Annexin V-FITC/PI Double Staining
5.7.2. ΔΨm Assay
5.7.3. RNA Extraction and Quantitative real time polymerase chain reaction (qRT-PCR)
5.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Rui, G.; Meng, Q.; Li, J.; Min, L.; Zhang, Y.; Bi, C.; Shan, A. Modified halloysite nanotubes reduce the toxic effects of zearalenone in gestating sows on growth and muscle development of their offsprings. J. Anim. Sci. Biotechno. 2016, 7, 570–578. [Google Scholar]
- Gao, X.; Sun, L.; Zhang, N.; Li, C.; Zhang, J.; Xiao, Z.; Qi, D. Gestational zearalenone exposure causes reproductive and developmental toxicity in pregnant rats and female offspring. Toxins 2017, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, J.; Li, Y.; Li, T.; He, Q.; Tang, Y.; Liu, H.; Su, Y.; Yin, Y.; Liao, P. Aflatoxin b1, zearalenone and deoxynivalenol in feed ingredients and complete feed from different province in china. J. Anim. Sci. Biotechno. 2016, 7, 63. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Logrieco, A.F.; Susca, A. Mycotoxins: An underhand food problem. In Mycotoxigenic Fungi; Moretti, A., Susca, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1542, pp. 3–12. [Google Scholar]
- Stanciu, O.; Banc, R.; Cozma, A.; Filip, L.; Miere, D.; Mañes, J.; Loghin, F. Occurence of fusarium mycotoxins in wheat from europe—a review. Acta. Universitatis. Cibiniensis. 2015, 19, 35–60. [Google Scholar] [CrossRef]
- Venkataramana, M.; Nayaka, S.C.; Anand, T.; Rajesh, R.; Aiyaz, M.; Divakara, S.T.; Murali, H.S.; Prakash, H.S.; Rao, P.V.L. Zearalenone induced toxicity in shsy-5y cells: The role of oxidative stress evidenced by n-acetyl cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Noser, J.; Wettstein, F.E.; Burkhardt-Holm, P. Unraveling the mechanisms involved in zearalenone-mediated toxicity in permanent fish cell cultures. Toxicon 2014, 88, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.D.; Su, P.; Shan, H. Mycotoxin contamination of maize in china. Compr. Rev. Food Sci. F. 2017, 16, 835–849. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar]
- Pinton, P.; Nougayrede, J.P.; Del Rio, J.C. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharm. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Font, G.; Ruiz, M.J. Cytotoxic effects of zearalenone and its metabolites and antioxidant cell defense in cho-k1 cells. Food Chem. Toxicol. 2016, 96, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Pistol, G.; Stancu, M. Effects of zearalenone and its metabolites on the swine epithelial intestinal cell line: Ipec 1. Proc. Nutr. Soc. 2013, 72, 85–89. [Google Scholar] [CrossRef]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Caloni, F.; Schreiber, N.B.; Spicer, L.J. Effects of fumonisin b1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology. 2014, 81, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Hassen, W.; Ayedboussema, I.; Oscoz, A.A.; Lopez, A.C.; Bacha, H. The role of oxidative stress in zearalenone-mediated toxicity in hep g2 cells: Oxidative DNA damage, gluthatione depletion and stress proteins induction. Toxicology 2007, 232, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yuan, H.; Guo, C.; Lu, Y.; Deng, S.; Yang, Y.; Wei, Q.; Wen, L.; He, Z. Zearalenone induces apoptosis and necrosis in porcine granulosa cells via a caspase-3- and caspase-9-dependent mitochondrial signaling pathway. J. Cell. Physiol. 2012, 227, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Ayed-Boussema, I.; Bouaziz, C.; Rjiba, K.; Valenti, K.; Laporte, F.; Bacha, H.; Hassen, W. The mycotoxin zearalenone induces apoptosis in human hepatocytes (hepg2) via p53-dependent mitochondrial signaling pathway. Toxicol. in Vitro 2008, 22, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R., Jr.; Santos, C.F.; Clarimundo, V.; Dalmaz, C.; Elisabetsky, E.; Gomez, R. N-acetylcysteine prevents behavioral and biochemical changes induced by alcohol cessation in rats. Alcohol 2015, 49, 259–263. [Google Scholar]
- Wang, W.; Dan, L.; Ding, X.; Zhao, Q.; Chen, J.; Tian, K.; Yang, Q.; Lu, L. N-acetylcysteine protects inner ear hair cells and spiral ganglion neurons from manganese exposure by regulating ros levels. Toxicol. Lett. 2017, 279, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Halliwell, B.; Hoey, B.M.; Butler, J. The antioxidant action of n-acetylcysteine: Its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radical Biol. Med. 1989, 6, 593–597. [Google Scholar] [CrossRef]
- Moreira, M.A.; Irigoyen, M.C.; Saad, K.R.; Saad, P.F.; Koike, M.K.; Montero, E.F.; Martins, J.L. N-acetylcysteine reduces the renal oxidative stress and apoptosis induced by hemorrhagic shock. J. Surg. Res. 2016, 203, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Halasi, M.; Wang, M.; Chavan, T.S.; Gaponenko, V.; Hay, N.; Gartel, A.L. Ros inhibitor n-acetyl-l-cysteine antagonizes the activity of proteasome inhibitors. Biochem. J. 2013, 454, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M.; Calzetta, L.; Facciolo, F.; Rogliani, P.; Matera, M.G. Pharmacological investigation on the anti-oxidant and anti-inflammatory activity of n-acetylcysteine in an ex vivo model of copd exacerbation. Resp. Res. 2017, 18, 26. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Liu, W.; Wu, J.; Yang, X.; Xu, H. Chemoprotective effect of n-acetylcysteine (nac) on cellular oxidative damages and apoptosis induced by nano titanium dioxide under uva irradiation. Toxicol. in Vitro 2011, 25, 110. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kaur, T.; Singla, S.K. Role of mitochondria and nadph oxidase derived reactive oxygen species in hyperoxaluria induced nephrolithiasis: Therapeutic intervention with combinatorial therapy of n-acetyl cysteine and apocynin. Mitochondrion 2016, 27, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Kondakçı, G.; Aydın, A.F.; Doğru-Abbasoğlu, S.; Uysal, M. The effect of n-acetylcysteine supplementation on serum homocysteine levels and hepatic and renal oxidative stress in homocysteine thiolactone-treated rats. Arch. Physiol. Biochem. 2017, 123, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. Beauvericin-induced cytotoxicity via ros production and mitochondrial damage in caco-2 cells. Toxicol. Lett. 2013, 222, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jing, L.; Yuan, H.; Peng, S.-q. T-2 toxin induces apoptosis in ovarian granulosa cells of rats through reactive oxygen species-mediated mitochondrial pathway. Toxicol. Lett. 2011, 202, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Espín, S.; Garcíafernández, A.J.; Ruiz, M.J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. in Vitro 2017, 45, 334. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; Sharaf el Dein, O.; Hajlaoui, M.R.; Lemaire, C.; Bacha, H. In vitro investigation of toxicological interactions between the fusariotoxins deoxynivalenol and zearalenone. Toxicon 2014, 84, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cornelia, B.; Sonia, S.; Roxana, C.P.; Raduly, L.; Ovidiu, B.; Ionelia, T.; Eliza, M.D.; Monica, M.; Ancuta, J.; Patriciu, A.C. Evaluation of cellular and molecular impact of zearalenone andescherichia colico-exposure on ipec-1 cells using microarray technology. BMC Genomics 2016, 17, 576. [Google Scholar]
- Wan, L.Y.; Turner, P.C.; Elnezami, H. Individual and combined cytotoxic effects of fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins b1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, E.; Juangarcía, A.; Font, G.; Ruiz, M.J. Reactive oxygen species induced by beauvericin, patulin and zearalenone in cho-k1 cells. Toxicol. in Vitro 2009, 23, 1504–1509. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Lee, H.; Yoo, Y.S.; Hah, D.Y.; Kim, C.H.; Kim, E.; Kim, J.S. Evaluation of oxidative DNA damage using an alkaline single cell gel electrophoresis (scge) comet assay, and the protective effects of n-acetylcysteine amide on zearalenone-induced cytotoxicity in chang liver cells. Toxicol. Res. 2013, 29, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Pundir, M.; Arora, S.; Kaur, T.; Singh, R.; Singh, A.P. Effect of modulating the allosteric sites of n -methyl-d-aspartate receptors in ischemia-reperfusion induced acute kidney injury. J. Surg. Res. 2013, 183, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.H.; Wang, J.H.; Zhu, L.S.; Chen, A.M.; Wang, J. Thiamethoxam induces oxidative stress and antioxidant response in zebrafish (danio rerio) livers. Environ. Toxicol. 2016, 31, 2006–2015. [Google Scholar] [CrossRef] [PubMed]
- Abid, E.S.; Ee, B.C.B. Comparative study of toxic effects of zearalenone and its two major metabolites alpha-zearalenol and beta-zearalenol on cultured human caco-2 cells. J. Biochem. Mol. Toxicol. 2009, 23, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Shen, T.; Ding, Q.; Lv, Y.; Li, L.; Huang, K.; Yan, L.; Song, S. Zearalenone induces ros-mediated mitochondrial damage in porcine ipec-j2 cells. J. Biochem. Mol. Toxicol. 2017, 31, e21944. [Google Scholar] [CrossRef] [PubMed]
- Dennog, C.; Radermacher, P.; Barnett, Y.A.; Speit, G. Antioxidant status in humans after exposure to hyperbaric oxygen. Mutat. Res. 1999, 428, 83–89. [Google Scholar] [CrossRef]
- Phamhuy, L.A.; He, H.; Phamhuy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Assady, M.; Farahnak, A.; Golestani, A.; Esharghian, M. Superoxide dismutase (sod) enzyme activity assay in fasciola spp. Parasites and liver tissue extract. Iran. J. Parasitol. 2011, 6, 17. [Google Scholar] [PubMed]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489. [Google Scholar] [CrossRef] [PubMed]
- Ashrafzadeh, T.H.; Saeed, H.; Foad, R.; Ashrafzadeh, T.M.; Hadi, H. Effects of n-acetylcysteine and pentoxifylline on remote lung injury in a rat model of hind-limb ischemia/reperfusion injury. J. Bras. Pneumol. 2016, 42, 9. [Google Scholar]
- Nazıroğlu, M.; Şenol, N.; Ghazizadeh, V.; Yürüker, V. Neuroprotection induced by n-acetylcysteine and selenium against traumatic brain injury-induced apoptosis and calcium entry in hippocampus of rat. Cell Mol. Neurobiol. 2014, 34, 895. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Zhang, S.; Zhang, M.; Yang, L.; Cheng, B.; Li, J.; Shan, A. Individual and combined effects of fusarium toxins on apoptosis in pk15 cells and the protective role of n -acetylcysteine. Food Chem. Toxicol. 2017, 111, 27–43. [Google Scholar]
- Yeo, E.H.; Goh, W.L.; Chow, S.C. The aminopeptidase inhibitor, z-l-cmk is toxic and induced cell death in jurkat t cells through oxidative stress. Toxicol. Mech. Methods. 2017, 28, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Kyotani, Y.; Itoh, S.; Nakayama, H.; Isosaki, M.; Yoshizumi, M. Big mitogen-activated protein kinase 1 protects cultured rat aortic smooth muscle cells from oxidative damage. J. Pharmacol. Sci. 2011, 116, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Strasser, A.; McDunn, J.E.; Swanson, P.E. Cell death. N. Engl. J. Med. 2009, 361, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Borys, S.; Khozmi, R.; Kranc, W.; Bryja, A.; Dyszkiewiczkonwińska, M.; Jeseta, M.; Kempisty, B. Recent findings of the types of programmed cell death. Adv. Cell Biol. 2017, 5, 43–49. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.Y.; Xu, M.H.; He, W.; Wang, S.T.; Guo, Z.Y.; Xu, M.H.; He, W.; Wang, S.T. Effect of mitochondria permeability transition pore on h9c2 myocardial cell apoptosis induced by lipopolysaccharide. J. Shanghai Jiaotong Univ. 2017, 37, 942–949. [Google Scholar]
- Vaca, C.E.; Wilhelm, J.; Harms-Ringdahl, M. Interaction of lipid peroxidation products with DNA. A review. Mutat. Res. 1988, 195, 137–149. [Google Scholar] [CrossRef]
- Hüttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Bras, M.; Queenan, B.; Susin, S.A. Programmed cell death via mitochondria: Different modes of dying. Biochemistry (Moscow) 2005, 70, 231–239. [Google Scholar] [CrossRef]
- Leal, A.M.D.S.; Queiroz, J.D.F.D.; Lima, T.K.D.S.; Agnezlima, L.F. Violacein induces cell death by triggering mitochondrial membrane hyperpolarization in vitro. BMC Microbiol. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abbès, S.; Salahabbès, J.B.; Ouanes, Z.; Houas, Z.; Othman, O.; Bacha, H.; Abdelwahhab, M.A.; Oueslati, R. Preventive role of phyllosilicate clay on the immunological and biochemical toxicity of zearalenone in balb/c mice. Int. Immunopharmacol. 2006, 6, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, G.; Kanbur, M.; Aslan, Ö.; Karabacak, M. The antioxidant effects of pumpkin seed oil on subacute aflatoxin poisoning in mice. Environ. Toxicol. 2013, 28, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.L.; Dunning, K.R.; Yang, X.; Russell, D.L.; Norman, R.J.; Robker, R.L. In Oocytes exhibit lipid accumulation, endoplasmic reticulum stress, mitochondrial dysfunction, and apoptosis in response to high fat diet. Biol. Reprod. 2010, 83, 185. [Google Scholar] [CrossRef]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (δψ m) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Zong, M.; Xu, W.; Yang, Z.; Bo, W.; Yang, M.; Tao, L. Natural pyrethrins induces apoptosis in human hepatocyte cells via bax- and bcl-2-mediated mitochondrial pathway. Chem. Biol. Interact. 2017, 262, 38–45. [Google Scholar]
- Guo, L.; Peng, Y.; Yao, J.; Sui, L.; Gu, A.; Wang, J. Anticancer activity and molecular mechanism of resveratrol-bovine serum albumin nanoparticles on subcutaneously implanted human primary ovarian carcinoma cells in nude mice. Cancer Biother. Radiopharm. 2010, 25, 471. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Yang, S.H.; Chien, C.M.; Lu, M.C.; Lo, C.S.; Lin, Y.H.; Hu, X.W.; Lin, S.R. Mechanisms of cardiotoxin iii-induced apoptosis in human colorectal cancer colo205 cells. Clin. Exp. Pharmacol. Physiol. 2006, 33, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Ding, D.; Su, J.; Manohar, S.; Salvi, R. Salicylate selectively kills cochlear spiral ganglion neurons by paradoxically up-regulating superoxide. Neurotox. Res. 2013, 24, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Hua, P.; Liu, J.; Tao, J.; Liu, J.; Yang, S. Influence of caspase-3 silencing on the proliferation and apoptosis of rat bone marrow mesenchymal stem cells under hypoxia. Int. J. Clin. Exp. Med. 2015, 8, 1624–1633. [Google Scholar] [PubMed]
- Wang, X.; Diao, Y.; Liu, Y.; Gao, N.; Gao, D.; Wan, Y.; Zhong, J.; Jin, G. Synergistic apoptosis-inducing effect of aspirin and isosorbide mononitrate on human colon cancer cells. Mol. Med. Rep. 2015, 12, 4750. [Google Scholar] [CrossRef] [PubMed]
- Vacca, R.A.; Valenti, D.; Bobba, A.; Merafina, R.S.; Passarella, S.; Marra, E. Cytochrome c is released in a reactive oxygen species-dependent manner and is degraded via caspase-like proteases in tobacco bright-yellow 2 cells en route to heat shock-induced cell death. Plant Physiol. 2006, 141, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the bcl-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell. Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Gao, X.; Li, C.; Krumm, C.S.; Qi, D.S. Individual and combined cytotoxic effects of aflatoxin b1, zearalenone, deoxynivalenol and fumonisin b1 on brl 3a rat liver cells. Toxicon 2015, 95, 6. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, G.; Lin, Z.; He, L.; Xu, L.; Zhang, Y. Characteristic and functional analysis of a newly established porcine small intestinal epithelial cell line. PLoS ONE 2014, 9, e110916. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, L.; Pruvost, A.; Guyot, A.; Farinotti, R.; Mabondzo, A. Combination of tenofovir and emtricitabine plus efavirenz: In vitro modulation of abc transporter and intracellular drug accumulation. Antimicrob. Agents Chemother. 2009, 53, 896. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Orientation | Sequences (5′→3′) | Fragments Size (bp) | Tm (°C) |
|---|---|---|---|---|---|
| β-actin | AY550069 | Forward | ATGCTTCTAGGCGGACTGT | 211 | 58.2 |
| Reverse | CCATCCAACCGACTGCT | ||||
| Bcl-2 | AB271960.1 | Forward | GCGACTTTGCCGAGATGT | 116 | 55.9 |
| Reverse | CACAATCCTCCCCCAGTTC | ||||
| Bax | XM_ 003127290.3 | Forward | TTTGCTTCAGGGTTTCATCC | 113 | 54.4 |
| Reverse | GACACTCGCTCAACTTCTTGG | ||||
| Cyto c | NM_ 001129970.1 | Forward | CTCTTACACAGATGCCAACAA | 139 | 56.1 |
| Reverse | TTCCCTTTCTCCCTTCTTCT | ||||
| Caspase-9 | XM_ 013998997.1 | Forward | GGACATTGGTTCTGGAGGATT | 116 | 52.3 |
| Reverse | TGTTGATGATGAGGCAGTGG | ||||
| Caspase-3 | NM_ 214131.1 | Forward | GACACTCGCTCAACTTCTTGG | 121 | 54.5 |
| Reverse | TTGGACTGTGGGATTGAGAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, M.; Zhang, W.; Gu, A.; Dong, J.; Li, J.; Shan, A. Protective Effect of N-Acetylcysteine against Oxidative Stress Induced by Zearalenone via Mitochondrial Apoptosis Pathway in SIEC02 Cells. Toxins 2018, 10, 407. https://doi.org/10.3390/toxins10100407
Wang J, Li M, Zhang W, Gu A, Dong J, Li J, Shan A. Protective Effect of N-Acetylcysteine against Oxidative Stress Induced by Zearalenone via Mitochondrial Apoptosis Pathway in SIEC02 Cells. Toxins. 2018; 10(10):407. https://doi.org/10.3390/toxins10100407
Chicago/Turabian StyleWang, Jingjing, Mengmeng Li, Wei Zhang, Aixin Gu, Jiawen Dong, Jianping Li, and Anshan Shan. 2018. "Protective Effect of N-Acetylcysteine against Oxidative Stress Induced by Zearalenone via Mitochondrial Apoptosis Pathway in SIEC02 Cells" Toxins 10, no. 10: 407. https://doi.org/10.3390/toxins10100407
APA StyleWang, J., Li, M., Zhang, W., Gu, A., Dong, J., Li, J., & Shan, A. (2018). Protective Effect of N-Acetylcysteine against Oxidative Stress Induced by Zearalenone via Mitochondrial Apoptosis Pathway in SIEC02 Cells. Toxins, 10(10), 407. https://doi.org/10.3390/toxins10100407