Abstract
Obesity is a global health concern with rising prevalence that increases the risk of developing other chronic diseases. A causal link connecting overnutrition, the development of obesity and obesity-associated co-morbidities is visceral adipose tissue (AT) dysfunction, characterized by changes in the cellularity of various immune cell populations, altered production of inflammatory adipokines that sustain a chronic state of low-grade inflammation and, ultimately, dysregulated AT metabolic function. Therefore, dietary intervention strategies aimed to halt the progression of obese AT dysfunction through any of the aforementioned processes represent an important active area of research. In this connection, fish oil-derived dietary long-chain n-3 polyunsaturated fatty acids (PUFA) in the form of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) have been demonstrated to attenuate obese AT dysfunction through multiple mechanisms, ultimately affecting AT immune cellularity and function, adipokine production, and metabolic signaling pathways, all of which will be discussed herein.
1. Introduction
The prevalence of overweight and obese individuals within the global population (consisting of both developing and developed countries) has steadily increased over the past 30 years [1,2]. As defined by body mass index (BMI), this increased prevalence of overweight (25–29.9 kg/m2) and obese (>30 kg/m2) individuals represents a global health epidemic [1] that is associated with the development of a myriad of other chronic conditions and diseases, including but not limited to insulin resistance (IR), hypertension, and dyslipidemia (collectively referred to as Metabolic Syndrome [3]), cardiovascular disease (CVD), type 2 diabetes (T2D), stroke, certain cancer types, liver and gall bladder disease, osteoarthritis, sleep apnea, and gynecological conditions [1,4]. Furthermore, the consequences of obesity extend beyond these physical comorbidities to include an increased risk of various psychological and psychosocial conditions [5,6,7,8].
In obesity, visceral adipose tissue (AT) exhibits a dysfunctional phenotype in comparison to that derived from lean individuals [9,10,11,12,13]. Functionally, AT is the primary storage site for excess energy in the form of triacylglycerol (TAG), but also functions as an endocrine organ that secretes signaling proteins, collectively termed adipokines (or cytokines, when not of AT origin), which influence systemic metabolism and immune function [11,14,15,16]. AT is composed of adipocytes and a non-adipocyte stromal vascular fraction (SVF), comprised of innate and adaptive immune cells, which are collectively engaged in the maintenance of AT insulin sensitivity (reviewed in [15,17,18,19,20]). However, overnutrition induces changes in the number and activity of AT immune cell populations [12,13,21,22,23,24,25] and the consequent paracrine interactions, or cross-talk, between those AT-infiltrated immune cells and resident adipocytes leads to increased production of inflammatory adipokines (e.g., leptin, monocyte chemoattractant protein (MCP)-1/chemokine (C-C motif) ligand 2 (CCL2), interleukin (IL)-6, tumor necrosis factor (TNF)-α and IL-1β) and decreased production of anti-inflammatory and insulin-sensitizing adipokines (e.g., adiponectin, IL-10) (reviewed here and in [19]) (shown in Figure 1). Accordingly, obese AT is characterized by chronic low-grade inflammation, which disrupts its normal functions and thus contributes to the development of Metabolic Syndrome [14]. As such, it is suggested that AT inflammation mechanistically links obesity to the development of CVD [26] and T2D [27].
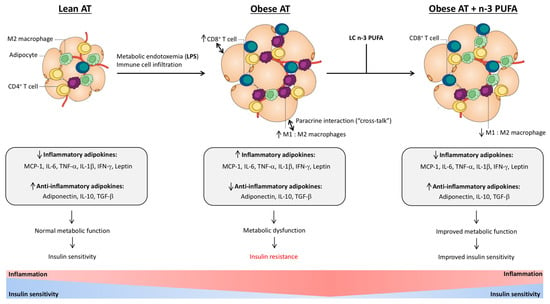
Figure 1.
Lean adipose tissue (AT) maintains an anti-inflammatory and insulin-sensitive tissue microenvironment through the secretion of anti-inflammatory adipokines (adiponectin, interleukin (IL)-10 and transforming growth factor (TGF)-β) and is populated by immune cells including M2-polarized macrophages and cluster of differentiation (CD)4+ regulatory T (Treg) cells. However, during the development of obesity, the population of AT immune cells shifts, demonstrated by an increase in M1 macrophage accumulation, CD4+ T helper 1 cells, and CD8+ T cells, which secrete (along with adipocytes) inflammatory adipokines, monocyte chemoattractant protein (MCP)-1, interleukin (IL)-6, tumor necrosis factor (TNF)-α, IL-1β, interferon (IFN)-γ and leptin, thus promoting an inflammatory AT microenvironment. Increased circulating lipopolysaccharide (LPS; i.e., metabolic endotoxemia) further promotes the secretion of inflammatory adipokines from adipocytes and immune cells in obese AT, promoting the paracrine interactions (“cross-talk”) between obese AT-infiltrated immune cells and resident adipocytes, leading to the development of metabolic dysfunction and insulin resistance (IR). Supplementation with long-chain (LC) n-3 polyunsaturated fatty acids (PUFA), eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), shifts the obese AT immune cell population towards a less inflammatory phenotype as suggested by the reduced M1:M2 macrophage ratio and increased anti-inflammatory adipokine production (adiponectin, IL-10, TGF-β), which improves metabolic function and insulin sensitivity.
Nutritional strategies for the prevention and treatment of AT dysfunction are of increasing importance given the prevalence of obesity [1,2]. As discussed throughout this review, dietary fatty acids, namely the fish oil (FO)-derived long-chain (LC) omega-3 (n-3) polyunsaturated fatty acids (PUFA), eicosapentaenoic acid (20:5n-3, EPA) and docosahexaenoic acid (22:6n-3, DHA), have been extensively studied by our research group and others as anti-inflammatory nutrients (the broader clinical implications of which have been reviewed in [28,29]) (shown in Figure 2). Investigation into the mechanisms underlying the n-3 PUFA-mediated attenuation of aspects of the obese inflammatory phenotype and/or associated metabolic dysfunction rely on rodent high-fat diet (HFD)-induced and genetic obesity models, whereas such evidence still requires confirmation in humans. Furthermore, despite efforts by our research group [30,31] and others [28,32,33], determinations of the minimum effective dosage of n-3 PUFA to modulate critical aspects of the obese human phenotype, namely systemic inflammation, are limited by the diversity amongst study designs (e.g., participant characteristics) and n-3 PUFA interventions (e.g., source and duration). Nevertheless, evidence from in vitro and rodent in vivo studies suggests that EPA and DHA can decrease inflammatory adipokine production in adipocytes [34,35,36,37,38,39,40,41,42,43]. Further, in an in vitro co-culture model of obese AT, we have shown that LC n-3 PUFA reduce the inflammatory cross-talk between adipocytes and (i) macrophages [44,45]; (ii) cluster of differentiation (CD)8+ T cells [46,47,48] and (iii) (CD)4+ T cells [49] via, at least in part, downregulation of inflammatory adipokine synthesis and secretion. As discussed herein, multiple factors contribute to the development of AT dysfunction in obesity, but the focus of this review will include the changes in AT immune cellularity, dysregulation of adipokine secretion, and inflammatory signaling mechanisms in obese AT, while identifying potential targets for n-3 PUFA intervention to mitigate the ensuing metabolic consequences of obesity.
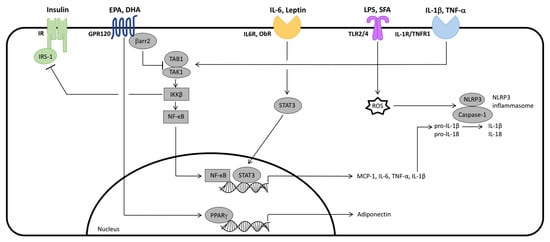
Figure 2.
Integration of selected cell signaling mechanisms regulated by long-chain (LC) n-3 polyunsaturated fatty acids, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), and inflammatory adipokines in obese adipose tissue (AT). EPA and DHA stimulate G-protein coupled receptor (GPR)120, which promotes the association between β-arrestin 2 (βarr2) and GPR120. This complex internalizes, allowing βarr2 to bind transforming growth factor-β activated kinase (TAK)1 binding protein (TAB)1, which inhibits TAK1/TAB1 binding and subsequent nuclear factor κ-light-chain-enhancer of activated B cells (NF-κΒ) activation. TAK1/TAB1 binding also leads to the inhibition of insulin signaling. Lipopolysaccharide (LPS) and saturated fatty acids (SFA) stimulate Toll-like receptor (TLR)2 and TLR4, and interleukin (IL)-1β and tumor necrosis factor (TNF)-α stimulate IL-1 Receptor (IL-1R) and TNF Receptor 1 (TNFR1), respectively, all of which promote TAK1/TAB1 binding and subsequent NF-κΒ activation. NF-κΒ (as well as IL-6 and leptin-induced signal transducer and activator of transcription (STAT)3) regulate inflammatory adipokine gene transcription. Among them, the immature protein form of IL-1β undergoes further processing to the mature form by the caspase-1 subunit of the nucleotide-binding oligomerization domain-like receptor, pyrin domain containing (NLRP)3 inflammasome, activated by TLR2/4-induced reactive oxygen species (ROS) accumulation. EPA and DHA signaling also promotes peroxisome proliferator-activated receptor (PPAR)γ activation and subsequent adiponectin gene expression.
2. Adipose Tissue Function and Obesity-Associated Dysfunction
2.1. Healthy Adipose Tissue Function
AT plays a fundamental role in the regulation of whole-body metabolic homeostasis, serving as both an energy storage depot and an active endocrine organ [15,16]. Adipocytes comprise approximately 90% of AT volume, but only 20–40% of the overall cellular content [21,22,23,50]. Hence, AT is a heterogeneous tissue composed of mature adipocytes and a non-adipocyte cells that comprise the SVF, which includes adipocyte progenitor cells and immune cell populations (e.g., macrophages, dendritic cells, natural killer cells, B cells and T cells (CD8+ and CD4+ T cell subsets) [15,20]. Functionally, within AT, adipocytes share several common features with immune cell populations, including expression of the innate pattern recognition receptors (PRR), nucleotide-binding oligomerization domain (NOD)-like receptors (NLR) and Toll-like receptors (TLR)2 and TLR4 [51,52,53]. In response to, for example, certain ligands (e.g., lipopolysaccharide (LPS) or adipokines), nutrient and/or oxygen status within the AT microenvironment, PRR signaling regulates the synthesis and secretion of a wide range of adipokines (shown in Figure 2), which in turn influence local and systemic immune function, and fatty acid and glucose homeostasis (reviewed in [54]).
Mature adipocytes are the primary energy storage cell type within AT, and as a dynamic tissue, AT undergoes remodeling to adjust its storage capacity to meet the energy storage demand (reviewed in [16,55,56]). During periods of excess energy intake (i.e., positive energy balance), insulin stimulates adipocytes to store free fatty acids (FFA) in the form of neutral TAG through their esterification to glycerol [16]. Likewise, adipocytes manage insulin-stimulated glucose uptake and utilization as a substrate for de novo lipogenesis; and finally mobilize these TAG stores via lipolysis for transport back into the circulation during energy deficit [16]. Intracellular TAG accumulation promotes adipocyte hypertrophy, which reduces the blood flow and delivery of oxygen per unit of adipocyte surface [55,56]. Hypoxic adipocytes become necrotic and secrete inflammatory adipokines that recruit immune cells, via chemotaxis, to phagocytose necrotic cell debris and stimulate local angiogenesis [55,56,57,58]. Dead adipocytes are eventually replaced with new, smaller adipocytes, ultimately increasing the capacity of AT to store excess energy [55,56].
2.2. Adipose Tissue Dysfunction in Obesity
AT is distributed throughout the body in subcutaneous and visceral depots; the former is generally considered to be a safer long-term energy storage depot in comparison to visceral AT depots, which increase in size in obesity and are associated with the development of metabolic complications [59,60,61,62]. To compensate for the continuous supply of FFA during overnutrition (positive energy balance), TAG accumulate within visceral AT adipocytes, thus inducing adipocyte hypertrophy [16]. Large adipocytes are characterized by decreased sensitivity to insulin and its anti-lipolytic effects, as well as dysregulated adipokine synthesis and secretion [60,63]. Eventually, adipocyte dysfunction leads to and is exacerbated by a ‘spillover’ of FFA, primarily of the saturated fatty acid (SFA) class (e.g., palmitic acid (16:0, PA)) [16], which may act in an autocrine or paracrine manner as ligands for inflammatory TLR2/4 signaling (shown in Figure 2). Specifically, SFA-induced TLR2/4 signaling induces a network of intracellular responses that further contribute to adipokine dysregulation and sustained chronic low-grade inflammation, including activation of the nuclear factor κ-light-chain-enhancer of activated B cells (NF-κΒ) transcription factor and the NLR, pyrin domain containing (NLRP)3 inflammasome [54,64,65,66,67,68,69,70,71,72,73,74,75] (discussed in Section 6). Ultimately, the autocrine and paracrine feedforward consequences of adipocyte dysfunction (i.e., adipokine dysregulation, FFA release) lead to whole AT dysfunction (reviewed here and in [16]). Specifically, dysfunctional AT cannot meet the ongoing demand for increased energy storage capacity during overnutrition and, coupled with its increased lipolytic activity, dysfunctional AT gives rise to chronically elevated circulating FFA levels [9,10] as reported in obese and T2D patients [76,77,78]. The spillover of FFA is delivered to ectopic tissues, including the liver and skeletal muscle, wherein the accumulation of lipid intermediates (e.g., diacylglycerol and ceramides) together with the abundance of inflammatory stimuli (e.g., inflammatory adipokines) ultimately impairs insulin signaling (reviewed in [15]), thereby causally linking AT dysfunction to systemic IR.
2.3. Adipokine Dysregulation in Obese Adipose Tissue
Adipokine synthesis and secretion from adipocytes and SVF cells is essential for normal AT function as a central regulator of systemic immunity and metabolism (reviewed in [15,16,79]). However, the constant demand to increase energy storage capacity during overnutrition leads to AT dysfunction, increased FFA release [63], and dysregulation of adipokine synthesis and secretion [11,60,80]; a process that we and others have shown is exacerbated by cross-talk between adipocytes and various immune cell populations [44,45,46,47,48,65,81]. The adipokine profiles of obese subcutaneous and visceral AT depots differ wherein visceral AT is associated with the metabolic complications of obesity [60,80]. Ultimately, obese visceral AT is characterized by a state of chronic low-grade inflammation owing, in part, to increased secretion of inflammatory adipokines and decreased secretion of anti-inflammatory and insulin-sensitizing adipokines (reviewed in [15,16,79]) (shown in Figure 1; discussed in Section 5). While an increasing number of adipokines are implicated in the development of the obese phenotype, the focus of this review will include MCP-1, IL-6, TNF-α, IL-1β, leptin and adiponectin as the key mediators of AT inflammation and dysfunction in obesity.
2.4. Metabolic Endotoxemia Drives Adipose Tissue Dysfunction in Obesity
Evidence from rodent models suggests that, in obesity, AT dysfunction is driven, in part, by increased circulating bacterial components (e.g., LPS, peptidoglycan, flagellin) and metabolites (e.g., secondary bile acids) (as reviewed in [82]). Among them, LPS, a component of Gram-negative bacteria cell walls [83,84,85,86,87,88], is the most commonly studied bacteria-derived inflammatory stimulus considered in investigations into the mechanistic link between the gut microbiota and obesity-associated inflammation. Studies assessing the effect of n-3 PUFA supplementation on either the lean or obese microbiome are limited [89,90], and therefore are beyond the scope of this review. Obesity is a gut-associated disease wherein the intestinal microenvironment of lean versus obese individuals differs dramatically including a dysbiotic microbial community structure and activity [91,92,93,94,95,96,97,98,99] and impaired intestinal epithelial barrier function [100,101]. The impaired obese intestinal microenvironment contributes to critical aspects of the obese pathology, in part by increasing energy harvested from non-digestible food components and initiating intestinal barrier dysfunction leading to enhanced barrier permeability, microbial invasion, and AT and systemic inflammation and metabolic dysfunction [83,101,102,103]. Moreover, these obesity-associated changes within the human intestinal microenvironment can be recapitulated in animal HFD-induced obesity models [83,104,105,106]. Studies performed in germ-free (microbiota-free) mice colonized with the cecal content obtained from obese mice have demonstrated that the disease phenotype (inflammation and/or metabolic dysfunction) can be transferred to the germ-free recipients through microbial colonization, thereby suggesting that gut microbes can drive metabolic alterations within the host tissues that ultimately produce the disease phenotype [107,108]. An important feature of the impaired obese intestinal microenvironment that contributes to the obese phenotype is reduced intestinal epithelial barrier integrity (i.e., “leaky gut”), which promotes bacterial translocation across the epithelial barrier leading to the development of increased systemic inflammation (metabolic endotoxemia), driven by bacterial-derived LPS signaling [91,103]. LPS is a potent systemic inflammatory stimulus that is significantly elevated in the blood of obese individuals [101,109] and rodents [83,110], and increased circulating LPS levels are attributable to a decrease in epithelial barrier function and integrity [83,91,111]. Intestinal bacterial overgrowth during obesity increases LPS levels within the enteric cavity, leading to greater epithelial mucosal barrier damage, increased bacterial translocation into the host tissues and, ultimately, metabolic endotoxemia [112]. LPS is a ligand for TLR2/TLR4 (shown in Figure 2; discussed in Section 6.1) expressed on the surface of adipocytes and immune cells [52], which signals to initiate NF-κB activation and adipokine secretion, thereby perpetuating the obese low-grade inflammatory state and subsequent metabolic dysfunction, including AT and systemic IR [66,83,85,113,114,115,116] (shown in Figure 1). For instance, in healthy humans, acute LPS administration increased AT production and circulating levels of MCP-1, IL-6 and TNF-α prior to the development of systemic IR [85]. Similarly, metabolic endotoxemia induced in mice via continuous infusion of LPS increased visceral AT mass, immune cell (i.e., macrophage) infiltration, and inflammation, as well as fasting glycaemia and insulinemia to a similar extent as observed in HFD-fed mice [83]. Importantly, the metabolic consequences of both LPS-infusion (i.e., metabolic endotoxemia) and HFD feeding were blunted in TLR4-deficient mice [83,117]. Thus, LPS is an important factor in obesity-induced AT dysfunction and warrants inclusion in studies investigating strategies to modulate adipokine synthesis and secretion.
3. Dietary n-3 Polyunsaturated Fatty Acids as a Strategy to Modulate Adipose Tissue Dysfunction in Obesity
Strategies to modulate adipokine synthesis and secretion from AT are warranted to attenuate the chronic low-grade inflammation that causally links obesity to pathologies such as systemic IR, CVD and T2D [14,26,27,60,85,118]. Since AT is adept at responding to nutritional stimuli, dietary n-3 PUFA may provide such a strategy by regulating the activity of adipokine and immune cell receptors that are intrinsic to adipokine modulation in the obese state.
n-3 PUFA can act as stimuli for specific cell membrane-bound (e.g., G protein-coupled receptor (GPR)120, discussed in Section 6.2) or intracellular (e.g., peroxisome proliferator-activated receptor (PPAR)-γ) receptors to directly influence inflammatory adipokine production by adipocytes and immune cells within AT (reviewed herein and in [119,120]). Further, LC n-3 PUFA membrane enrichment influences membrane fluidity, formation of lipid rafts and subsequent signal transduction efficiency (discussed here and in [121,122,123]), or can serve as substrates for the synthesis of bioactive lipid mediators (i.e., eicosanoids), which can influence inflammatory signaling (reviewed in [29]).
4. Obese Adipose Tissue Immune Cells and Modulation by n-3 Polyunsaturated Fatty Acids
4.1. Altered Immune Cell Composition in Obese Adipose Tissue
In lean AT, various immune cells, such as M2-polarized macrophages (F4/80+, CD11b+, CD11c−) and CD4+ regulatory T (Treg) cells (CD4+, forkhead box P3 (FOXP3)+), are engaged in the maintenance of insulin sensitivity, partly through their secretion of anti-inflammatory adipokines (reviewed in [15,17,18,19,20]). However, overnutrition induces changes in the number and activity of visceral AT immune cell populations, which collectively direct development of the obese phenotype [12,13,21,22,23,24,25] (shown in Figure 1). For instance, obese AT is characterized by increased macrophage accumulation, a greater proportion of which are polarized to the M1 inflammatory phenotype (F4/80+ CD11b+ CD11c+; discussed in Section 4.2) [12,21,22,124,125,126,127]. Further, Treg cell abundance is significantly reduced in obese AT [22,23,128,129,130], as are CD4+ T helper (Th)2 cells [13,23]. Conversely, the proportion of infiltrating dendritic cells [131,132], B cells [133,134], NK cells [135,136,137], CD4+ Th1 cells [13,22,23,138,139], and CD8+ T cells [13,22,23] are reported to increase in obese AT.
As obesity progresses, AT-derived FFA and inflammatory adipokines act in a controlled autocrine, paracrine and endocrine manner to ultimately recruit and activate immune cells in an attempt to repair the AT dysfunction (reviewed in [15,17,18,19,20]). However, as the metabolic consequences of overnutrition persist, immune cell infiltration into obese AT becomes dysregulated [22,23,24,124,133,137,140]. Specifically, FFA and LPS serve as ligands for adipocyte and immune cell TLR2/4 signaling [52,66,68,83,141,142] (discussed in Section 6.1), which promote the synthesis and secretion of inflammatory and chemotactic adipokines via, in part, NF-κΒ and NLRP3 inflammasome activation [73,74,143,144] (discussed in Section 6.1 and Section 6.3, respectively). Further, our research group and others have shown that the gene expression and/or secretion of inflammatory adipokines increases with LPS stimulation in adipocytes [43,116,145,146,147] and immune cells alone [38,147,148,149], as well as in co-culture [45,46,47,48,49,81], suggesting that the cross-talk between obese AT-infiltrated immune cells and resident adipocytes yields a vicious cycle that leads to a local and systemic state of chronic low-grade inflammation and metabolic dysfunction [20]. In summary, though many other immune cell populations including dendritic cells [131,132], B cells [133,134], NK cells [135,136,137], neutrophils [150], eosinophils [151], and mast cells [152] play key roles in the maintenance of AT homeostasis and development of the obese AT phenotype, perhaps through direct or indirect modulation of resident AT macrophage responses (reviewed in [15,17,18,19,20]), this review will focus on adipokine-mediated cross-talk between adipocytes, macrophages and T cells (CD4+ and CD8+ subsets).
4.2. Obesity-Associated Changes in Adipose Tissue Macrophages
Among AT-infiltrating immune cell populations, macrophages have taken center stage as a hallmark of the obese AT phenotype since their degree of infiltration is associated with the progression of IR (reviewed in [75]). Specifically, the percentage of AT macrophages within the SVF increases by 20–30% in obese versus lean AT, in both humans and rodent obesity models [12,21,22,124,125,126,127]. AT macrophages cluster around necrotic adipocytes, forming crown-like structures (CLS) [153], which are observed more frequently in obese versus lean visceral AT [60].
There are two well-defined macrophage phenotypes that can be differentiated from circulating monocytes that infiltrate AT; however, macrophage phenotype is highly plastic in response to signals within the surrounding microenvironment [154,155], and thus these polarization states are not absolute and can vary along the M1–M2 spectrum [154]. These polarized macrophage subsets are referred to as M1, or ‘classically activated’, and M2, or ‘alternatively activated’, macrophages [154,156]. In obese AT, the macrophage population undergoes a phenotypic switch from the M2 phenotype (the dominant macrophage phenotype in lean AT) to the M1 phenotype, which functionally exacerbates AT inflammatory mediator production [12,21,22,124,125,126,127] (shown in Figure 1). The M1 macrophage phenotypic shift has been shown to occur when lipids are repartitioned from hypertrophic adipocytes to macrophages during obesity progression, wherein M1 macrophages form in response to lipotoxicity and resemble TAG droplet laden foam-cells [157].
Specially, M1 macrophages can be activated by the Th1 cytokine, interferon (IFN)-γ, or by TLR2/4 ligands such as gut-derived LPS or FFAs released from dysfunctional adipocytes [158,159,160,161]. The downstream effect of TLR2/4 signaling results in inflammatory adipokine secretion from M1 macrophages, yielding a paracrine communication loop with adipocytes and other immune cell populations that contribute to local AT, and ultimately systemic metabolic dysfunction [65,68]. Post-activation, M1 macrophages undergo a respiratory burst followed by production of reactive oxygen (ROS) and reactive nitrogen species (RNS) that promote microbicidal responses, antigen presentation via the major histocompatibility complex (MHC)II, and the secretion of adipokines that promote Th1 and/or Th17 cell-mediated responses [155,156].
Overactivation of M1 macrophages promote chronic inflammation [154], as these cells are characterized by increased lipid content, increased expression of anti-microbicidal inducible nitric oxide synthase (iNOS, observed in murine obese AT only), and secretion of inflammatory adipokines, namely IL-6, TNF-α and IL-1β, which impair adipocyte insulin signaling and promote lipolysis [12,69,75,124,126,154,157,162,163]. In this connection, M1 macrophages localize to CLS and macrophage-derived adipokines have been shown to perturb adipocyte insulin sensitivity [164,165] leading first to local AT IR and ultimately systemic IR [12,14,126,153,162,166,167]. M1 macrophages can be identified by increased cellular surface expression of integrin/complement receptors 3 (CD11b) and 4 (CD11c) [164] and this AT macrophage subset (F4/80+, CD11b+, CD11c+) has been shown to exhibit an enhanced inflammatory response when exposed to FFAs (i.e., TLR2/4 ligands), which is abrogated by TLR-4 antagonism [66]. Further, CD11c+ expressing macrophages appear to be a crucial contributor to obese AT dysfunction, as ablation of these cells results in reduced systemic inflammation and normalization of insulin sensitivity [168,169].
Conversely, M2 macrophages are characterized by the expression of scavenging receptors, mannose receptor (CD206) and macrophage galactose-type C-type lectin 1 (MGL1), and MHCII and co-stimulatory molecules [155]. Maturation of M2 macrophages, which exhibit anti-inflammatory and regulatory (i.e., tolerogenic) immune functions, is promoted by anti-inflammatory mediators such as IL-10, transforming growth factor (TGF)-β and glucocorticoids [156], and the ingestion of apoptotic cells [170]. Interestingly, macrophage polarization is not absolute and may shift throughout the development of obesity, or with weight loss interventions [171]. Therefore, macrophage populations exhibiting a mixed M1/M2 phenotype have been observed in obese AT in mice [171,172] and humans [12,167,173,174,175], wherein these cells express moderate levels of M1 (CD11c) and M2 (MGL1 or CD206) cell surface markers.
4.3. Influence of n-3 Polyunsaturated Fatty Acids on Obese Adipose Tissue Macrophages
Cross-talk between adipocytes and macrophages in obese AT represents a significant contributor to the obese AT inflammatory adipokine profile [65,69]. Multiple studies have demonstrated the effect of LC n-3 PUFA supplementation in HFD-induced rodent models of obesity, including reduced visceral AT M1 macrophage accumulation in favour of M2 macrophage polarization [42,176,177,178,179,180,181,182], which is associated with decreased inflammatory mediator production and increased systemic insulin sensitivity [176,177,179] (shown in Figure 1). Interestingly, these effects are similar to those reported in the HFD-fed Fat-1 mouse, which is capable of synthesizing LC n-3 PUFA de novo [183]. In randomized controlled human interventions, n-3 PUFA supplementation has been shown to reduce the formation of CLS observed in abdominal AT biopsies, which was associated with reduced circulating MCP-1 levels [184]; whereas macrophage number and CLS abundance in subcutaneous abdominal AT biopsies were unaffected by n-3 PUFA supplementation [185], thereby highlighting the differential effects between subcutaneous and visceral AT depots.
Mechanistically, cell culture studies from our group and others’ have demonstrated the ability of LC n-3 PUFA to reduce macrophage M1 polarization while promoting M2 polarization status, and to inhibit inflammatory adipokine expression and/or secretion [35,38,44,165,184,186,187,188]. Furthermore, these effects have been shown to be dependent, at least in part, on various mediators and signaling pathways, including adiponectin [187] and PPARγ activation [186], although further study is required. Interestingly, n-3 PUFA-derived lipid mediators, namely resolvins and protectins, which exert anti-inflammatory and pro-resolving physiological functions [189,190], have also been shown to influence macrophage function and inflammatory mediator production by stimulating phagocytic activity, decreasing infiltration into CLS, and promoting their polarization towards the M2 phenotype [179,191,192,193,194]. Ultimately, decreased secretion of inflammatory adipokines from obese AT could serve to decrease the ongoing recruitment of macrophages and ensuing inflammatory adipocyte-macrophage cross-talk [35,38,44,165,184,186,187] that exacerbates chronic inflammation in obesity.
4.4. Obesity-Associated Changes in Adipose Tissue T Cells
Despite the macrophage-centric focus of AT immune cells, several studies have demonstrated the significant contribution of T cells in the development of the obese phenotype, in both humans [13,25,140,195,196] and rodent models [22,23,24,25,50,138,195,196,197,198,199] (shown in Figure 1). In fact, T cells have been shown to localize to CLS [138,195,196,197], suggesting a role in the cross-talk between adipocytes and macrophages. Further, although controversy exists [168] and human data is less clear, changes in T cell AT infiltration (increased by 0.5–5% of the SVF), activation and/or effector status has been shown to precede significant AT macrophage accumulation in diet-induced obese rodent models [22,23,24,196]. In humans, a positive correlation between AT T cell accumulation and the degree of adiposity has been reported, with greater accumulation in visceral AT depots compared to subcutaneous [13,140], and the proportion of activated (CD25+, CD69+) AT T cells is consistently higher in obese individuals compared to lean [25]. Importantly, depletion of visceral AT T cells in diet-induced obese mice improved AT inflammation and systemic insulin sensitivity in young but not adult mice [50], suggesting an early window of time during which T cell-mediated immune function may be controlled in obesity.
Controversy exists over which T cell subset is the first to exhibit changes in cellular abundance or activation in obese AT; CD4+ T cells [23,196], CD8+ T cells [197,200,201], or both [13,24,138,177,202,203]. Such discrepancies have yet to be explained, but one reason could be the different methods used to express and normalize T cell number within AT, such as percentage of total SVF cells, number of cells/g of AT and number of cells/AT depot [17].
4.5. Obesity-Associated Changes in Adipose Tissue CD4+ T Helper Cell Subsets
CD4+ T cells are divided into Th1, Th2, Th17 and Treg subtypes as defined by the expression and/or production of signature transcription factors and cytokines (reviewed in [204]). Accordingly, obese AT is characterized by an increased cellular abundance of the inflammatory Th1 cell subtype [23,139,199,205] and a decreased abundance of the non-inflammatory Treg subtype [23,201,206,207,208]. However, controversy exits with respect to the AT abundance and function of Th17 and Th2 cell subsets in lean and obese AT [13,23].
Th1 cells can be identified by T-box transcription factor (T-bet) expression and the secretion of inflammatory cytokines such as IFN-γ, whereas Th2 cells are identified by GATA-binding protein (GATA)3 transcription factor expression and secrete anti-inflammatory IL-4 and IL-13 (reviewed in [18,204]). Although Th1 and Th2 cells are present in equal proportions in the AT of lean rodent models, diet-induced obesity has been shown to induce a dramatic increase in the Th1 cellular number, with no change in Th2 cells [23,139]. Likewise, human AT T cells have been reported to exhibit a Th1 (IFN-γ-secreting) profile [13,140], whereas the number of AT Th2 cells inversely correlates with IR and circulating liver-derived C-reactive protein (CRP) [13], a clinical inflammatory biomarker [209,210]. Accordingly, adipocyte-derived leptin and FFA, both of which are increased in obesity, have been shown to enhance Th1 proliferation and/or IFN-γ production [211,212].
In general, Th1 cells participate in obese AT inflammation and promote IR, primarily owing to their secretion of IFN-γ (reviewed in [18]). IFN-γ has been shown to stimulate adipocytes to express T cell and macrophage chemoattractants, whereas systemic depletion of IFN-γ reduces AT macrophage accumulation, local inflammation, and improved systemic insulin sensitivity [138]. In contrast, however, the depletion of all mature lymphocytes in the recombination activating gene (Rag)1-deficient mouse did not protect against HFD-induced obese AT inflammation and systemic IR [23]. Interestingly, adoptive CD4+ T cell transfer reversed these aspects of the obese phenotype, but predominantly through the actions of Th2 cells as adoptive transfer of T cells from signal transducer and activator of transcription (STAT)6-deficient rodents, which have normal Th1 but impaired Th2 development, did not [23]. Altogether, these data suggest the significant and opposing roles of Th1 and Th2 cells in the development of the obese phenotype.
Th17 cells are identified by retinoic acid receptor-related orphan receptor (ROR)γτ expression and IL-17 secretion (reviewed in [18,204]), and have been shown to promote inflammatory responses in autoimmune diseases; however, their role in obese AT requires further investigation. Th17 cells are detectable in AT, but their cellular abundance is minimal and remained unchanged in obese mice versus lean [23]; although, controversy exists since a higher proportion of Th17 cells has been observed in the peripheral blood and/or subcutaneous AT of obese humans [212,213,214,215]. Interestingly, the major cellular source of AT IL-17 has been reported to be γδT cells rather than αβT cells, and IL-17-deficient mice are more susceptible to HFD-induced obesity, but remain insulin sensitive [216]. Accordingly, IL-17 has been shown to impair adipogenesis and adipocyte expression of genes involved in glucose and lipid metabolism [216], in addition to increasing the pre-adipocyte, adipocyte and whole AT expression of inflammatory adipokines [217]. Likewise, IL-17 production has been reported to correlate with the severity of T2D in humans [218].
Tregs, identified by FOXP3 expression [204], play an important role in self-tolerance and reduce inflammation by suppressing autoreactive T cells and M1 macrophages (reviewed in [219]). Accordingly, in AT, Tregs are found dispersed between adipocytes, but also in CLS in close contact with macrophages and CD8+ T cells [128]. Lean AT is highly enriched with Tregs, comprising 30–40% of all AT CD4+ T cells; a proportion that is significantly higher compared to both lymphoid and other non-lymphoid tissues [23,128,220,221]. However, the Treg cellular proportion of all AT CD4+ expressing T cells decreases during the development of obesity [22,23,128,206], yet controversy exists [222]. Nonetheless, Treg depletion experiments have demonstrated their significance in limiting the development of the obese phenotype, wherein rodent models of genetic and HFD-induced obesity exhibited increased AT inflammation and decreased systemic insulin sensitivity [128,223]. Likewise, in similar rodent models of obesity, AT Treg accumulation produced opposing effects, consistent with a reduced AT M1 macrophage content and increased IL-10-producing M2 macrophages [23,128,224].
4.6. Influence of n-3 Polyunsaturated Fatty Acids on Obese Adipose Tissue CD4+ T Helper Cell Subsets
To our knowledge, there is only one report on the effect of n-3 PUFA on CD4+ T cells in obese AT wherein n-3 PUFA supplementation did not affect mesenteric AT expression of CD4 or FOXP3 in HFD-induced obese mice [177]. However, dietary plant-derived n-3 PUFA (α-linolenic acid (18:3n-3, ALA)-enriched flaxseed) has been shown to reduce perirenal AT total CD3+ T cell abundance in obese leptin receptor-defective fa/fa Zucker rats [225]. The ability of n-3 PUFA to influence effector subset polarization and function of CD4+ T cells has been described in other experimental conditions or disease models, thereby providing the proof of concept that T cell effector subset polarization and function may be influenced within obese AT by n-3 PUFA, although further study is required.
Antigen-driven CD4+ T cell activation involves the formation of an immunological synapse and assembly of the signalsome protein complex, which is stabilized by the actin cytoskeleton [226,227,228]. Formation of the immunological synapse involves reorganization of nanoscale lipid rafts and signaling proteins [227,229,230] and T cell activation is suppressed with lipid raft disruption [231,232]. Importantly, n-3 PUFA have been shown to alter the stability and/or size of lipid rafts in CD4+ T cells [122,233,234,235,236] and suppress downstream cellular activation via mechanisms including (i) displacement of T cell activation-associated signaling proteins from detergent-resistant membrane fractions [122,235,237,238,239], (ii) IL-2 secretion [122,240,241,242], (iii) lymphoproliferation [235,243,244], and (iv) mitochondrial translocation [245]. With respect to specific CD4+ T cell subsets and Th1/Th2 balance, n-3 PUFA have been shown to influence both polarization and signature cytokine secretion in favor of the Th2 subset while concomitantly decreasing the activation of the Th1 subset [246,247,248]. Additionally, n-3 PUFA have been shown to reduce CD4+ T cell polarization of Th17 cells, in part through lipid raft-mediated disruption of IL-6 signaling [249,250], whereas there was no effect of n-3 PUFA on Treg polarization [249]. Interestingly, in a model of concurrent HFD-induced obesity and colitis, dietary supplementation of n-3 PUFA reduced visceral AT gene expression of the Th1 subset signature cytokine, IFN-γ, and the Th17 transcription factor, RORγτ, and signature cytokine, IL-17, with no effect on FOXP3 expression [178].
4.7. Obesity-Associated Changes in Adipose Tissue CD8+ T Cells
The primary function of CD8+ T cells, also known as cytotoxic T cells, is to kill infected cells by producing perforin, granzymes and inflammatory cytokines, namely IFN-γ (reviewed in [251]). The degree of splenic CD8+ T cell activation in vitro was demonstrated to be markedly increased when co-cultured with obese versus lean AT [22], which complements in vivo reports in humans and rodents wherein the proportion of activated CD8+ T cells is increased in the SVF of obese AT versus lean [22,23,24,25,50,196,198,252].
Functionally, the obese AT microenvironment does not influence the development or maintenance of memory CD8+ T cell responses that are primed either before or after obesity is established [253]. Specifically, obesity did not impact the maintenance or function of pre-existing memory CD8+ T cells (i.e., cell surface phenotypic markers, cytokine production and secondary expansion), nor the differentiation and maintenance of newly activated memory CD8+ T cell responses [253]. Conversely, CD8+ T cells have been shown to accumulate in AT and localize to CLS in advance of macrophage accumulation in obese humans and rodents [22,25,50,197]. Specifically, in HFD-fed mice, CD8+ T cells infiltrated visceral AT within two weeks and reached their peak cellular abundance of 10% of the SVF by 11 weeks. Conversely, however, macrophage AT infiltration was not observed until six weeks of HFD feeding, but continued to increase thereafter [22].
The significance of CD8+ T cells in macrophage-mediated metabolic dysfunction in obesity has been elegantly demonstrated by Nishimura and colleagues, both in vitro and in vivo [22]. Specifically, conditioned media collected from activated CD8+ T cells contained several chemotactic and inflammatory cytokines (e.g., MCP-1) that induced macrophage migration and activation. Interestingly, macrophage differentiation and proliferation were dependent upon the cross-talk between CD8+ T cells and obese AT, as neither CD8+ T cells nor AT alone exerted the same effect as co-culture. Further, CD8+ T cell depletion protected mice against HFD-induced obese visceral AT gene expression of inflammatory and macrophage chemotactic adipokines, which coincided with reduced M1 macrophage infiltration and CLS frequency without affecting the M2 macrophage fraction, as well as with improved systemic insulin sensitivity. However, adoptive transfer of CD8+ T cells into HFD-fed CD8+ T cell-deficient mice reversed this protection [22].
4.8. Influence of n-3 Polyunsaturated Fatty Acids on Obese Adipose Tissue CD8+ T Cells
Although mesenteric AT gene expression of CD8 was shown to be reduced with n-3 PUFA supplementation in HFD-induced obese mice [177], the effect of n-3 PUFA on obese AT CD8+ T cell abundance/tissue infiltration and function, namely cellular function and influence on the development of critical components of the obese AT phenotype, has not been determined in vivo. Interestingly, our group has shown the potential of LC n-3 PUFA to modulate obese AT function utilizing a co-culture model comprised of 3T3-L1 murine adipocytes and primary splenic CD8+ T cells purified from mice consuming a FO (i.e., LC n-3 PUFA-enriched) diet. The obese AT microenvironment was recapitulated using a co-culture cellular ratio of 10% CD8+ T cells to adipocytes, as described in obese AT by Nishimura and colleagues [22], which was stimulated with LPS at a concentration that mimics in vivo circulating levels in obesity [83,109,254]. LC n-3 PUFA-enriched co-cultures exhibited both an anti-inflammatory and anti-chemotactic secretory profile that consisted of reduced activation of inflammatory transcription factors (NF-κB and STAT3) and reduced secretion of both inflammatory and macrophage chemotactic adipokines, which was functionally confirmed by reduced macrophage chemotaxis [46] and polarization towards the M1 phenotype [47]. Furthermore, these findings were confirmed in separate studies utilizing the same co-culture model comprised of adipocytes and plant-derived n-3 PUFA (i.e., ALA)-enriched CD8+ T cells [255], and well as FO (i.e., LC n-3 PUFA)-enriched CD8+ T cells purified from obese mice [47]. Interestingly, in the latter model, we showed that LPS-stimulated adipocyte-CD8+ T cell inflammatory cross-talk and ensuing M1 macrophage polarization and adipocyte dysfunction are attenuated by TNF-α neutralization [47,48].
5. Adipose Tissue Inflammation in Obesity and Modulation by n-3 PUFA
5.1. Monocyte Chemoattractant Protein-1
MCP-1, also known as CCL2 in humans, is a potent chemoattractant that recruits circulating monocytes and macrophages to the site of inflammation (i.e., obese AT) via binding the cell membrane CCL2 receptor (CC2R) (reviewed in [256]). AT and circulating levels of MCP-1 are increased in obese humans [11,257], and in rodent models of HFD-induced and genetic obesity [70,258]. Visceral AT has been shown to secrete more MCP-1 than subcutaneous AT and, while controversy exists, adipocytes are reported to be the main cellular source [81,257,259] with increased MCP-1 production reported in adipocytes isolated from obese versus lean humans [11,53]. Likewise, our group has observed increased MCP-1 gene expression and secretion from 3T3-L1 murine adipocytes stimulated with low-dose LPS [43] to mimic in vivo circulating levels in obesity [83,109,254]. Further, the cross-talk between co-cultured murine adipocytes and splenic immune cells, representative of cells of the SVF within AT, has been shown to increase MCP-1 secretion [81], which our group has confirmed in unstimulated co-cultures of murine 3T3-L1 adipocytes and RAW 264.7 macrophages [44], as well as in LPS-stimulated co-cultures of murine 3T3-L1 adipocytes with either splenic CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49]. Thus, adipocyte-immune cell cross-talk may play a crucial role in the recruitment of macrophages to obese AT and the development of AT and systemic inflammation and IR [14,75,258].
AT production of MCP-1 is induced by stimuli that are reported to be elevated in the obese state, including LPS and FFA [147]. For instance, in healthy humans acutely administered LPS to mimic metabolic endotoxemia, AT production and circulating levels of MCP-1 increased prior to the development of systemic IR [85]. Further, MCP-1 synthesis and secretion was upregulated in PA- and LPS-treated human and 3T3-L1 murine adipocytes in vitro [116,147,260], but blunted by antagonizing TLR4 signaling or NF-κΒ activity [52,71] (discussed in Section 6.1). MCP-1 is also an insulin-responsive gene that remains sensitive in an insulin resistant state, as demonstrated in vitro in 3T3-L1 adipocytes induced to be insulin resistant, and in vivo in ob/ob mice [261]. In turn, MCP-1 further contributes to the development of IR as MCP-1 treatment was shown to impair 3T3-L1 adipocyte insulin-stimulated glucose uptake and the expression of several adipogenic genes, including PPARγ [261].
The significance of MCP-1 in the pathology of obesity-induced metabolic dysfunctions has been further demonstrated in HFD-fed MCP-1−/− and CCR2−/− mice. In brief, both knockout models were partially protected from the HFD-induced increase in adiposity and exhibited reduced AT macrophage accumulation and inflammatory adipokine production [258,262]. Further, MCP-1/CCR2-deficiency increased adiponectin expression, and improved systemic glucose homeostasis and insulin sensitivity; an effect that was mirrored by acute CCR2 antagonism in mice with established HFD-induced obesity [262]. Taken together, the metabolic endotoxemia and hyperinsulinemia that are characteristic of the obese phenotype may contribute to AT inflammation through, in part, the macrophage chemotactic function of adipocyte-derived MCP-1.
5.2. Interleukin-6
Circulating levels of the IL-6 are positively correlated with adiposity, circulating FFA, and IR in humans [263,264], and accordingly, increased circulating IL-6 is predictive of the development of T2D [265]. Approximately 15–35% of systemic IL-6 is secreted by AT [266], and an in vitro comparison suggested that obese visceral AT secretes more IL-6 than subcutaneous [263]. Further, IL-6 concentrations in the interstitial fluid of AT were reported to be 100-fold higher compared to circulating levels in the same participants [267], highlighting the significance of AT IL-6 secretion and its local action within obese AT.
LPS stimulates AT-derived IL-6 production [83,85] as demonstrated by the dose-dependent increase in IL-6 gene expression and secretion from human [116,145] and 3T3-L1 murine adipocytes in vitro [52,260]. Further, our group confirmed these findings in 3T3-L1 adipocytes using a low dose of LPS [43] to mimic in vivo circulating levels in obesity [83,109,254]. However, adipocytes secrete approximately 10% of total AT-derived IL-6 [263]; thus, the cells of the SVF, including M1-polarized macrophages, are considered to be the primary cellular source [11,126,268]. Nonetheless, adipocytes express the IL-6 receptor (IL-6R) [263,267], suggesting a role for IL-6 in the cross-talk between adipocytes and immune cells within AT, as confirmed in co-cultured adipocytes and a mixed population of SVF cells [81], and in co-cultured adipocytes with either CD11b+ macrophages [44,45], CD8+ T cells [46,47], or CD4+ T cells [49].
Upon binding IL-6, adipocyte IL-6R activates Janus kinase (JAK) family members, leading to the activation of transcription factors of the STAT family (reviewed in [269]) (shown in Figure 2). Adipocyte IL-6-signaling, in general, leads to dysregulated adipokine production and impaired insulin action. For instance, in vitro treatment with IL-6 increased 3T3-L1 murine adipocyte IL-6 synthesis and secretion in a feedforward manner [270], and decreased expression of the insulin-sensitizing adipokine, adiponectin [267]. IL-6 also induces its negative regulator, suppressor of cytokine signaling (SOCS)3, which has been shown to interfere with insulin receptor substrate (IRS)-1 and IRS-2 in 3T3-L1 murine adipocytes [271]. Accordingly, AT IL-6 was inversely correlated with insulin-stimulated adipocyte glucose uptake in vitro [264], which coincides with reduced IRS-1, glucose transporter 4, and PPARγ gene expression in cultured adipocytes treated with IL-6 [267,270,272,273]. Thus, IL-6-mediated adipocyte dysfunction and adipokine secretion may potentiate the cross-talk between adipocytes and immune cells within AT to contribute to the maintenance of chronic low-grade inflammation in obesity.
5.3. Tumor Necrosis Factor-α
TNF-α is a potent inflammatory adipokine that is overexpressed in human [274,275] and rodent [22,37,276] obese AT and, likewise, circulating levels of TNF-α are increased in obese humans [277] and correlate with markers of IR [278]. TNF-α is synthesized as a 26-kDa transmembrane monomer that undergoes proteolytic cleavage to yield a 17-kDa soluble TNF-α molecule; both of these are biologically active and are increased in human obese versus lean AT [279]. Characteristics of the obese phenotype, including metabolic endotoxemia, induce AT synthesis and secretion of TNF-α as evidenced by acute LPS administration in healthy humans [85]. Similarly, acute TNF-α infusion in healthy humans has been shown to induce systemic IR [280]. The significance of TNF-α in the pathology of obesity-induced metabolic dysfunctions has been further demonstrated in genetic and HFD-induced rodent models of obesity wherein TNF-α knockout or neutralization attenuated the development of systemic IR [276,281].
TNF-α expression is higher in obese visceral versus subcutaneous AT, and in cells of the SVF compared to adipocytes [21,65,275]. While TNF-α gene expression is increased in adipocytes isolated from obese versus lean humans [53], and in cultured human [116] and 3T3-L1 murine adipocytes treated with LPS [146], there are conflicting reports [145]. Additionally, M1-polarized macrophages within AT are suggested to be the primary cellular source compared to adipocytes, although further study is needed [21,65]. Nonetheless, adipocytes express TNF-α receptor (TNFR)1 and TNFR2 [275], suggesting a role for TNF-α in the cross-talk between adipocytes and immune cells within AT. The ligand-binding extracellular domains of TNFR1/2 are highly homologous, unlike the intracellular domains, which activate different signaling pathways (reviewed in [282]). The majority of evidence suggests that TNFR1 mediates the effects of TNF-α on AT dysfunction [275,283,284,285]. Indeed, neutralization of TNFR1 but not TNFR2 down-regulated expression of inflammatory adipokines in human adipocytes cultured in SVF-conditioned media [275]. Nonetheless, TNFR2 is suggested to cooperate with TNFR1 to regulate TNF-α signaling in chronic inflammatory conditions [282] such as obesity, and accordingly, only TNFR2 gene expression is reported to increase in obese versus lean human adipocytes [275,285].
Adipocyte TNF-α signaling is reported to induce inflammatory adipokine production and lipolysis via, in part, an NF-κΒ-dependent mechanism [65,69] (shown in Figure 2) and, further, TNF-α impairs insulin action by inhibiting the normal tyrosine phosphorylation of IRS-1 [284,286]. We and others have demonstrated that TNF-α secretion increases in co-cultured murine 3T3-L1 adipocytes and RAW 264.7 macrophages [44], coinciding with increased adipocyte gene expression of MCP-1, IL-6 and TNF-α, as well as increased FFA release in a similar model [65]. In turn, the adipocyte-derived SFA, PA, stimulated macrophage TNF-α gene expression [65], creating a vicious cycle that promotes AT dysfunction. Further, these inflammatory effects of adipocyte-macrophage cross-talk were determined to be independent of cell-cell contact [44,65]. Importantly, the increase in adipocyte inflammatory adipokine production and lipolysis were attenuated to a similar degree by both a TNF-α neutralizing antibody and a NF-κΒ inhibitor [65,69], which is consistent with findings in human adipocytes cultured in SVF-conditioned media [275]. Related to this, our group has shown that TNF-α gene expression and protein secretion increase in LPS-stimulated murine 3T3-L1 adipocytes co-cultured with either splenic CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49]. Interestingly, we have also shown that LPS-stimulated adipocyte-CD8+ T cell inflammatory cross-talk and ensuing M1 macrophage polarization and adipocyte dysfunction are attenuated by TNF-α neutralization [47,48]. In summary, TNF-α induces adipocyte lipolysis and regulates NF-κΒ activity to initiate a vicious cycle between adipocytes and immune cells that is central to the development of AT inflammation in obesity.
5.4. Interleukin-1β
IL-1β is a potent inflammatory adipokine that is overexpressed in human [11,53,207] and rodent [53,207,287] obese AT, and increased circulating levels are predictive of the development of T2D [265]. Compared to subcutaneous AT, obese visceral AT expresses more IL-1β, as well as the IL-1 cell membrane receptor (IL-1R), which reportedly coincides with increased AT accumulation of M1-polarized macrophage and CD8+ T cells [207,287,288]. Accordingly, cells of the SVF, particularly M1-polarized macrophages, are suggested to be the primary source of IL-1β within AT, although human and murine adipocyte IL-1β synthesis and secretion is markedly increased in obese versus lean AT [53,207,288]. Indeed, our group has demonstrated increased IL-1β gene expression in 3T3-L1 murine adipocytes stimulated with low-dose LPS [43] to mimic in vivo circulating levels in obesity [83,109,254]. Regardless of the cellular source, AT production of IL-1β is dependent upon the activation of NF-кB [289]; hence, IL-1β gene expression is increased in cultured human and 3T3-L1 murine adipocytes and macrophages stimulated with LPS, PA and TNF-α [53,73,116,290]. Subsequently, IL-1β is synthesized as an inert pro-protein and is cleaved by caspase-1, a cysteine protease domain of the NLRP3 inflammasome, to yield the mature, bioactive form of IL-1β in response to various obesity-induced intracellular stressors, such as ROS accumulation (reviewed in [291,292]) (shown in Figure 2; discussed in Section 6.3).
AT-derived IL-1β acts in an autocrine or paracrine fashion to contribute to obese AT inflammation and dysfunction, and accordingly, ablation of IL-1β signaling in IL-1R knockout mice attenuated the HFD-induced AT and systemic IR [287]. As a ligand for the IL-1R, which also mediates NF-кB activity [293], IL-1β has been shown to stimulate inflammatory adipokine synthesis, impair insulin-stimulated glucose uptake, and induce lipolysis in cultured human and 3T3-L1 murine adipocytes [294,295]. Thus, it is conceivable that IL-1β contributes to the inflammatory cross-talk between adipocytes and immune cells that ultimately impairs AT function. Indeed, our group has shown that LPS-stimulated 3T3-L1 murine adipocyte-specific IL-1β expression increases in co-culture with splenic CD11b+ macrophages [45]. Further, administration of an IL-1β neutralizing antibody blunted the expression and secretion of inflammatory adipokines, FFA release, and markers of IR in human adipocytes cultured in macrophage-conditioned media [163]. Also, our group has demonstrated that increased IL-1β gene expression coincides with increased secretion of other inflammatory adipokines in LPS-stimulated, co-cultured murine 3T3-L1 adipocytes with either splenic CD8+ T cells [46,47] or CD4+ T cells [49]. Interestingly, IL-1β and TNF-α have been shown to synergistically increase AT NF-кB activity and inflammatory adipokine production ex vivo [287]. Accordingly, TNF-α markedly increased 3T3-L1 murine adipocyte IL-1β gene expression and protein secretion [290] yet, in turn, TNF-α secretion was reduced in 3T3-L1 adipocytes co-cultured with macrophages derived from IL-1R knockout mice, which coincided with improved adipocyte insulin-stimulated glucose uptake [287]. Taken together, IL-1β contributes to the chronic low-grade inflammatory state that is characteristic of obese AT by impairing adipocyte function and regulating NF-кB activity to exacerbate adipokine dysregulation.
5.5. Inflammatory Adipokine Modulation by n-3 Polyunsaturated Fatty Acids in Obesity
In vitro investigations have demonstrated the anti-inflammatory effects of LC n-3 PUFA (EPA and DHA) on adipokine secretion (shown in Figure 2), however, some studies have shown that DHA is more potent than EPA in both the absence and presence of LPS [35,44,165]. The ability of dietary LC n-3 PUFA to modulate adipokines in the presence of LPS is significant in light of the contribution of metabolic endotoxemia to AT inflammation in obesity. Both LC n-3 PUFA (EPA and DHA) have been shown to attenuate both adipocyte (unstimulated and LPS-stimulated) [34,36,37,39,40,43,296,297] and macrophage [35,37,38,42] inflammatory adipokine secretion. Furthermore, our group has demonstrated reduced inflammatory adipokine secretion from co-cultured murine 3T3-L1 adipocytes and RAW264.7 macrophages treated with EPA or DHA or both [44], as well as from LPS-stimulated co-cultures of murine 3T3-L1 adipocytes with FO (i.e., LC n-3 PUFA)-enriched splenic CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49]. Likewise, LC n-3 PUFA supplementation has consistently been shown to reduce visceral AT production of inflammatory adipokines in rodent models of both genetic and diet-induced obesity [41,176,177,179,180,183,194,298].
In humans, LC n-3 PUFA have been shown to improve several metabolic risk factors, namely blood lipid levels [299]; however, the effects on circulating adipokines (i.e., cytokines) are less clear as circulating levels are not always reflective of the local AT concentrations. Instead, circulating CRP has emerged as a leading clinical inflammatory biomarker because of its association with habitual inflammatory status [209,210]. However, the effects of LC n-3 PUFA on any inflammatory markers in overweight/obese individuals are unclear; thus, determination of the minimum effective dosage of LC n-3 PUFA to modulate such critical aspects of the obese phenotype are limited, perhaps owing to differences between study designs, the inflammatory biomarkers analyzed, the duration of LC n-3 PUFA intervention, as well as the source (i.e., dietary sources versus supplements) and dose of LC n-3 PUFA interventions. In this connection, a range of LC n-3 PUFA intake levels (for example, 0.6–6.0 g/day EPA + DHA) from dietary sources and supplements has been shown to result in a range of outcomes in circulating inflammatory mediators in overweight/obese individuals, as reviewed by our research group [30,31] and others [28,32,33]. For instance, in randomized controlled interventions, dietary intake of LC n-3 PUFA reduced circulating levels of CRP and IL-6 in obese men [300], and improved circulating levels of CRP and IL-6 but not TNF-α in overweight women [301]. Further, purified DHA supplementation reduced circulating CRP, IL-6 and TNF-α levels in obese men and women, whereas EPA supplementation only reduced IL-6 [302]. Despite these findings, other studies report no association between LC n-3 PUFA intake and circulating inflammatory cytokines [303,304,305,306,307,308]. Thus, given the controversy, circulating levels of the aforementioned inflammatory cytokines may not be the optimal or reproducible primary endpoint in human studies to assess the efficacy of LC n-3 PUFA supplementation in improving aspects of the obese phenotype.
5.6. Adiponectin and Leptin
Adiponectin and leptin are two AT-derived adipokines whose functions tend to oppose one another in respecitvely attenuating or promoting obese AT inflammatory dysfunction (reviewed in [309]). In obesity, AT synthesis and circulating levels of adiponectin are reduced [310,311,312,313] such that adiponectin concentrations are inversely correlated with adiposity [312]. Adiponectin circulates in different oligomeric forms of trimeric, hexameric, or high molecular weight (HMW) [314], wherein the levels of the HMW isoform correlates most closely with systemic insulin sensitivity [315]. Thus, adiponectin exerts insulin-sensitizing effects and improves lipid metabolism in adipocytes and in peripheral tissues such as liver and skeletal muscle (reviewed in [316,317]). Accordingly, adiponectin deficiency was shown to induce IR in a rodent model, whereas its overexpression improved insulin sensitivity and glucose tolerance [318]. Similarly, in a rodent model of genetic obesity, overexpression of adiponectin reversed many characteristic components of the obese phenotype, resulting in improved glucose and lipid metabolic parameters, decreased circulating inflammatory adipokine levels, and reduced macrophage AT infiltration compared to obese littermates [313].
Adipocyte and/or visceral AT adiponectin expression and secretion is downregulated by inflammatory adipokines whose expression is increased within obese AT [272,294,319,320]. Specifically, the suppressive effects of TNF-α on adiponectin secretion from adipocytes can be partially recovered by a JNK inhibitor [320]. Conversely, adiponectin stimulation has been shown to suppress LPS-induced inflammatory adipokine production in adipocytes [321] via inhibition of NF-κB [322] and upregulation of PPARγ expression [323]; beneficial anti-inflammatory effects that are attenuated in obesity. Adiponectin antagonizes inflammatory adipokine expression by inhibiting NF-κB activation [324,325,326,327], and by stimulating anti-inflammatory IL-10 secretion from macrophages [328,329,330,331], which express the adiponectin receptor (AdipoR1/R2) and are responsive to adiponectin signaling [332]. Functionally, adiponectin has been shown to suppress M1 macrophage activation and production of inflammatory adipokines, in favour of promoting the polarization of M2 macrophages [323,331,332,333]. Additionally, adiponectin has been shown to influence dendritic cell function by decreasing the expression of co-stimulatory molecules (CD80/CD86) that resulted in reduced CD4+ T cell proliferation and increased FOXP3+ Treg expansion [334]; effects that could be beneficial in obese AT given the reduced tissue abundance of Tregs [22,23,128,206], although further study is required.
Obesity is also characterized by sustained elevated circulating levels of the adipokine leptin, which is positively associated with body fat mass [335,336,337]. While many cell types express the leptin receptor (Ob-Rb), including adipocytes, myocytes and hepatocytes, it is also expressed by immune cells such as T cells and macrophages, suggesting an immunomodulatory role for leptin in obese AT [211,338]. In this connection, leptin exerts inflammatory effects by promoting the secretion of inflammatory adipokines, namely TNF-α and IL-6 [338,339]. In turn, inflammatory stimuli such as LPS and other inflammatory adipokines stimulate leptin expression in AT, thereby creating a feedforward loop that perpetuates the low-grade chronic inflammatory phenotype that characterizes obese AT [340,341,342,343]. In this connection, leptin has also been shown to promote macrophage activation, proliferation, and enhanced phagocytic activity and secretion of inflammatory adipokines [338,339,344,345]. Leptin also induces T cell proliferative responses by polarizing CD4+ T cells towards the Th1 IFN-γ-secreting subset [211,346] and inhibiting Treg proliferation [347,348], whose cellular abundance in obese AT declines relative to AT mass [22,23,128,129,130]. Taken together, leptin contributes to the inflammatory microenvironment of obese AT.
5.7. Adiponectin and Leptin Modulation by n-3 Polyunsaturated Fatty Acids in Obesity
Dietary LC n-3 PUFA have been shown to increase circulating levels of adiponectin in obesity in both humans [305,349,350,351,352] and rodent models [179,180,351,353,354], thereby attenuating the obesity-associated reduction in adiponectin [310,311,312,313]. Our group has shown that EPA and DHA reduces the gene expression of M1 macrophage markers via an adiponectin-dependent mechanism [44,45]. Furthermore, LC n-3 PUFA function as activators of PPARγ [355] and, as such, LC n-3 PUFA have been shown to upregulate adiponectin production in adipocytes via a PPARγ-dependent mechanism [356,357,358,359]. Furthermore, PPARγ antagonizes NF-κB nuclear activity through a trans-repression mechanism, thereby decreasing the expression of NF-κB responsive genes [360], including inflammatory adipokines, as elegantly demonstrated in vivo by macrophage-specific deletion of PPARγ, which highlighted the critical role of PPARγ in the regulation of macrophage polarization (promoting the M2 phenotype and reducing M1), as well as AT and systemic inflammation and metabolic dysfunction [361].
Dietary LC n-3 PUFA have also been shown to reduce AT gene expression and/or circulating levels of leptin in overweight and obese humans [362,363] and rodents [178,364,365], wherein EPA has been shown to be more effective versus DHA [366]. Although the specific mechanisms through which LC n-3 PUFA exert these effects are undetermined, the ability to reduce leptin production within dysfunctional obese AT likely contributes to the LC n-3 PUFA-mediated reduction in inflammatory adipokine secretion and changes in AT macrophage and T cell function. Collectively, these data demonstrate the critical effect of LC n-3 PUFA on AT adipokine production that ultimately attenuates both the metabolic and inflammatory dysfunction that characterizes obese AT.
6. Cell Signaling Mechanisms Regulating Adipokine Production in Obese Adipose Tissue
6.1. TLR2/4 and NF-κB Regulation of Adipokines
Within AT, adipocytes and immune cells both express several signaling receptors including PRR, TLR2 and TLR4 [51,52,115], which respond to pathogen-associated molecular patterns (PAMPs) and contribute to the development of AT inflammation and IR in the obese state (reviewed in [54]). LPS is a well-established ligand for TLR4 within AT [52,83] and SFA (e.g., PA) are reported to be ligands for both TLR2 and TLR4 [66,68,69,117], although controversy exists [40,71]. A consequence of TLR2/4 stimulation is the activation of the NF-κB transcription factor complex [52] (shown in Figure 2). Specifically, the stimulation of TLR2/4 induces myeloid differentiation primary-response protein (MyD)88/interleukin-1 receptor-associated kinase (IRAK)1 signaling, which leads to the phosphorylation and activation of transforming growth factor-β activated kinase (TAK)1 by promoting its association with the TAK1 binding protein (TAB)1 [367]. Active TAK1 promotes NF-κB activation by phosphorylating and activating the inhibitor of κB (IκB) kinase (IKK) complex to downregulate IκB and allow NF-κB to translocate to the nucleus [367]. In turn, NF-κB regulates the expression of inflammatory and chemotactic adipokines [52,143,368], and thus, plays a pivotal role in the innate and adaptive immune responses within obese AT. Accordingly, the expression of TLR2 and TLR4, as well as the activity of NF-κB, are increased in obese and type 2 diabetic humans [109,369,370], and in rodent models of HFD-induced obesity [66,142,371,372].
The significance of TLR2/4 signaling in the pathology of obesity-induced metabolic dysfunctions has been demonstrated in vitro and in knockout mouse models fed a HFD. For instance, a substantial amount of evidence supports that LPS induces NF-κB activity and the production of inflammatory adipokines, as well as impairs insulin signaling in cultured human [109,116,369] and 3T3-L1 murine adipocytes [43,65,71]; all of which was inhibited by co-treatment with an antibody against TLR4 [52]. TLR2 and TLR4-dependent NF-κB activity and inflammatory adipokine production have also been demonstrated in adipocytes and macrophages treated with the SFA, PA [64,65,66,67,68,69,70,71,72]. Interestingly, in vitro TLR4 signaling was demonstrated to increase TLR2 expression in adipocytes [66], and co-culture of adipocytes with TLR4-deficient macrophages significantly attenuated inflammatory adipokine gene expression and adipocyte lipolysis [69], suggesting that TLR2/4 signaling contributes to the inflammatory cross-talk between adipocytes and immune cells within obese AT. Indeed, our group has demonstrated that LPS increases inflammatory adipokine expression and secretion in 3T3-L1 murine adipocytes alone [43] and in co-culture with splenic CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49]. In vivo, TLR2 and TLR4 knockout rodent models were protected against the HFD-induced increase in visceral AT mass, NF-κB activity and gene expression of inflammatory adipokines, which coincided with reduced circulating levels of MCP-1 and ensuing AT accumulation of M1-polarized macrophages [66,117,142]. Further, the same rodent models were protected again HFD-induced IR [66,117,141]; an effect that was also demonstrated in TLR4 knockout mice infused with lipids [66] and infused with LPS to induce metabolic endotoxemia [83]. Taken together, TLR2 and TLR4 play a crucial role in modulating adipokines in response to circulating LPS and SFA, and therefore represent potential targets for dietary intervention during development of the obese phenotype.
6.2. TLR2/4 and NF-κB Modulation by n-3 Polyunsaturated Fatty Acids
During the progression of obesity, LC n-3 PUFA antagonize AT inflammation by antagonizing LPS- and SFA-induced TLR2/4 signaling in adipocytes and immune cells. Specifically, LC n-3 PUFA are suggested to block the ligand binding sites of TLR2 and TLR4, as demonstrated in cultured RAW 264.7 murine macrophages wherein the LPS- and SFA-induced activation of NF-κB was inhibited by EPA and DHA [64,372]. Likewise, our group has shown that EPA and DHA blunt LPS-induced inflammatory adipokine gene expression and specifically MCP-1 secretion [43].
The incorporation of LC n-3 PUFA (EPA and DHA) into cell membranes disrupts the formation of lipid rafts that are necessary for TLR4 signaling [122,123,373,374]. In cultured macrophages, DHA inhibited the LPS- and SFA-induced recruitment of TLR4 and MyD88 to lipid raft fractions, which coincided with reduced ROS accumulation and NF-κB activity [123]. Similarly, our group has shown that NF-κB activity and/or inflammatory adipokine production is blunted in LPS-stimulated co-cultures of murine 3T3-L1 adipocytes and FO (i.e., LC n-3 PUFA)-enriched murine splenic CD11b+ macrophages [45] or CD8+ T cells [46,47]. It is conceivable that the TLR4 response to LPS was perturbed in LC n-3 PUFA-enriched immune cell-adipocyte co-cultures in a lipid raft-dependent manner, but the underlying mechanisms remain unknown. TLR2 activation may also be dependent upon lipid raft formation to promote TLR2 dimerization with cell membrane-bound TLR1 or TLR6 [54], and accordingly, DHA was reported to inhibit TLR2 signaling and subsequent inflammatory adipokine production in adipocytes and macrophages in vitro [37,72,375].
6.3. GPR120-Dependent Regulation of Adipokines
GPR120 is highly expressed in adipocytes and macrophages and plays a pivotal role in the maintenance of metabolic homeostasis [37]. Although body weight and AT mass were unaffected, GPR120 knockout mice exhibit impaired insulin sensitivity compared to wildtype [37]. Likewise, in a HFD-fed obese rodent model, GPR120 knockout increased body weight and AT mass, as well as AT M1 macrophage accumulation, consistent with a greater degree of systemic IR compared to WT rodents [376]. In turn, macrophage-conditioned media has been shown to inhibit GPR120 expression in human adipocytes [377], as has the classically M1 macrophage-derived TNF-α and IL-1β [378]. Interestingly, GPR120 expression is increased in human obese versus lean visceral AT, yet its dysfunction is associated with the development of the obese phenotype [376].
GPR120 expression is dependent on functional PPARγ and vice versa [378,379], and both of these respond to n-3 PUFA [37,42,359,380,381]. The n-6 PUFA, arachidonic acid (AA), is reported to induce GPR120 signaling, though is a less potent ligand compared to EPA and DHA [382], and SFA exert no effect on GPR120 signaling [37]. Ligand-stimulated GPR120 promotes the association of GPR120 and β-arrestin2 (βarr2), an adaptor protein that mediates GPR120 internalization and signaling [37] (shown in Figure 2). The GPR120- βarr2 complex then interrupts TLR2/4 signaling by associating with TAB1 to block the association between TAB1 and TAK1 and, therefore, block TAK1 phosphorylation/activation and downstream NF-κB activation [37,42,383]. Thus, GPR120 may represent a link between dietary LC n-3 PUFA and the modulation of inflammatory adipokines in obesity.
6.4. GPR120 Modulation by n-3 Polyunsaturated Fatty Acids
The role of GPR120 in mediating the effects of dietary LC n-3 PUFA has primarily been interpreted from EPA and DHA-treated adipocytes and macrophages in vitro [37,42,378,380,381,383], and in GPR120 knockout rodent models fed a HFD enriched with n-3 PUFA [37,376]. For instance, EPA and DHA attenuated the LPS- and TNF-α-induced gene expression and secretion of inflammatory adipokines in cultured macrophages via a GPR120- and βarr2-dependent mechanism [37,42]. Specifically, the anti-inflammatory actions of EPA and DHA in vitro mimicked those induced by a selective GPR120 agonist, GW9508, and were lost in macrophages isolated from GPR120 and βarr2 knockout rodent models [37,42]. Similarly, our group has shown that the anti-inflammatory effects of DHA are mimicked by GW9508 in 3T3-L1 murine adipocytes [43] wherein other work has shown that GPR120-agonism mitigates NF-κB activation [383]. Ultimately, GPR120 is reported to regulate adipocyte function as DHA increased basal and insulin-stimulated glucose uptake in 3T3-L1 murine adipocytes, but the effect was abrogated by GPR120 knockdown [37].
In HFD-fed rodents, the insulin-sensitizing effects of EPA and DHA supplementation were lost as a result of GPR120 knockout, which coincided with increased M1 macrophage accumulation and inflammatory adipokine expression in obese visceral AT [37,376]. Importantly, the change in MCP-1 expression was specific to adipocytes, and both MCP-1 and adipocyte-conditioned media promoted macrophage chemotaxis in vitro [37]; an effect that was blunted by macrophage pre-treatment with DHA, but not in macrophages isolated from GPR120 knockout mice [37], suggesting the potential for GPR120 to mediate the anti-inflammatory action of LC n-3 PUFA on adipocyte-immune cell cross-talk within obese AT. Further, our group has shown that EPA and DHA attenuate the expression and secretion of inflammatory adipokines in co-cultured murine 3T3-L1 adipocytes and RAW264.7 macrophages [44], and likewise in LPS-stimulated co-cultures of murine 3T3-L1 adipocytes and FO (i.e., LC n-3 PUFA)-enriched splenic CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49]. While the underlying mechanisms remain unknown, it is conceivable that GPR120 signaling negatively regulated inflammatory signaling in these co-culture models, although further study is required.
6.5. NLRP3 Inflammasome-Dependent Regulation of Adipokines
During the progression of obesity, the NLRP3 inflammasome regulates the innate immune response within AT (reviewed in [291,292]). Inflammasomes are multi-protein complexes comprised of a danger-sensing intracellular PRR from the family of NLR, such as NLRP3; the cysteine protease, caspase-1; and the adaptor protein, apoptosis-associated speck-like protein containing a caspase-recruitment domain (PYCARD) (reviewed in [291,292]). The NLRP3 inflammasome assembles in response to many stimuli including endogenous obesity-induced metabolites (reviewed in [291,292]), such as the accumulation of ROS [384,385,386] (shown in Figure 2). Next, active caspase-1 cleaves pro-IL-1β and pro-IL-18 to produce their mature isoforms [291,292]. In turn, IL-1β, specifically, impairs AT insulin signaling [163,287,295] and, hence, increased circulating levels of IL-1β are predictive of the development of T2D [265].
In humans, adipocyte but not SVF expression of NLPR3 inflammasome components and activation of the caspase-1 domain correlate with adiposity [53], suggesting that hypertrophic and dysfunctional adipocytes dominate the NLRP3 inflammasome-mediated response in obesity. Accordingly, the expression and activation of the NLRP3 inflammasome is increased in obese versus lean [53,203,207,387], and visceral versus subcutaneous AT [288], which coincides with increased IL-1β secretion form obese visceral AT [207,288]. Further, NLRP3 inflammasome (i.e., caspase-1) activity correlates with increased M1 macrophage and CD8+ T cell accumulation in visceral AT of obese humans [207,288]. Related to this, our group has shown that the gene expression of NLRP3 inflammasome components, IL-1β production, and/or caspase-1 activity are increased in co-cultures of murine 3T3-L1 adipocytes and CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49], highlighting the significance of adipocyte-immune cell cross-talk in the activation of the NLRP3 inflammasome. Accordingly, NLRP3 knockout mice were protected against the HFD-induced immune cell (e.g., macrophages, CD8+ T cells) infiltration into visceral obese AT, which coincided with reduced AT inflammation and improved systemic insulin sensitivity [203,388].
NLRP3 inflammasome activity is regulated by two signals; an initial priming signal to induce the gene expression of NLRP3 and pro-IL-1β, and a second signal to activate the multi-protein complex to promote caspase-1 activity [291,292]. Priming of the NLRP3 inflammasome is potently induced by inflammatory signals that activate the NF-κB transcription factor to regulate NLRP3 and IL-1β expression, such as inflammatory adipokines or those transmitted via TLR2/4 (e.g., LPS or SFA) to induce MyD88/IRAK1 signaling [72,143,144,389]. In addition to priming, the MyD88/IRAK1 signaling axis directly links TLR2/4 signaling to activation of the NLRP3 inflammasome [390], perhaps, in part, via a ROS-dependent mechanism as SFA (e.g., PA) and LPS have been shown to prime and activate the NLRP3 inflammasome in cultured adipocytes and macrophages, yet the effects were blunted by antioxidant pre-treatment [39,72,73,74]. Thus, ROS accumulation may be a crucial intermediate in TLR2/4-mediated NLRP3 inflammasome activity and IL-1β secretion in obese AT. Importantly, secreted IL-1β can act in an autocrine or paracrine fashion as a ligand for IL-1R, which also induces MyD88/IRAK1 signaling [293] and has been demonstrated to prime the NLRP3 inflammasome in cultured adipocytes [53]. Therefore, the NLRP3 inflammasome mediates a vicious cycle within obese AT that contributes to adipokine dysregulation and systemic IR.
6.6. NLRP3 Inflammasome Modulation by n-3 Polyunsaturated Fatty Acids
Recent evidence suggests that dietary fatty acids can modulate NLRP3 inflammasome activity (reviewed in [391]), which is suppressed by LC n-3 PUFA in particular. For instance, DHA supplementation reduced AT NLRP3 inflammasome (i.e., caspase-1) activity and IL-1β production in a HFD-induced obese rodent model, which coincided with improved systemic insulin sensitivity to a similar degree that was observed in NLRP3 knockout rodents consuming the same diet [42]. Interestingly, the effects of DHA were lost in NLRP3 knockout rodents, suggesting that the anti-inflammatory and insulin-sensitizing effects of DHA during a HFD are dependent upon the inhibition of NLRP3 inflammasome activity [42]. Unfortunately though, to our knowledge, this is the only in vivo investigation into the ability of dietary LC n-3 PUFA to modulate the NLRP3 inflammasome in obesity. However, related to this, our group established an in vitro co-culture model of obese AT and showed that LC n-3 PUFA decrease NLRP3 inflammasome priming, IL-1β production, and/or caspase-1 activity in LPS-stimulated co-cultures of murine 3T3-L1 adipocytes and CD11b+ macrophages [45], CD8+ T cells [46,47], or CD4+ T cells [49].
Other in vitro evidence supporting LC n-3 PUFA-mediated inhibition of the NLRP3 inflammasome suggests mechanisms that overlap with their inhibition of TLR2/4 signaling [37,42]. For instance, in murine bone marrow-derived macrophages, pre-treatment with EPA, DHA and, to a lesser extent, ALA inhibited the LPS-induced priming of the NLRP3 inflammasome and activation of the caspase-1 domain, which coincided with reduced IL-1β secretion [42]. Interestingly, the ability of EPA and DHA to inhibit NLRP3 inflammasome activation was not dependent upon their enzymatic metabolism to lipid mediators (e.g., resolvins), but was, in part, dependent upon a GPR120-mediated interaction between βarr2 and NLRP3 [42]. However, since GPR120 signaling was only partially responsible for the LC n-3 PUFA-mediated inhibition of NLRP3 inflammasome activity [42], another mechanism must be involved. Accordingly, DHA has been shown to disrupt the PA-induced recruitment of MyD88 to lipid rafts, which inhibited TLR2/4 activation, downstream ROS accumulation, and IL-1β secretion from human macrophages [72].
7. Conclusions
Obese AT inflammation and dysfunction is sustained via multiple interrelated mechanisms that integrate the effects of altered immune cell AT infiltration and altered cellular abundance of inflammatory subsets [12,13,21,22,23,24,25], with cross-talk via paracrine interactions with adipocytes to stimulate the secretion of a panel of inflammatory adipokines that collectively influence local AT and systemic metabolic function [11,15,16,19,60,79,80]. Interestingly, we have outlined both the immune cell- and adipocyte-mediated mechanisms through which LC n-3 PUFA impact AT immune cell polarization and function via critical cell signaling pathways within AT to modulate inflammatory adipokine secretion as well as local (i.e., AT) and systemic metabolic function, thereby improving these critical aspects of the obese phenotype. Further research is required to explore these LC n-3 PUFA-mediated mechanisms to improve obese AT dysfunction and to determine how other dietary fatty acids can influence these cellular processes. Additionally, determining the effective dosage of LC n-3 PUFA intake (either through dietary or supplemental sources) as well as that of other dietary fatty acids, namely plant-derived n-3 PUFA (i.e., ALA) represent other potential future directions. Collectively, the AT response to dietary LC n-3 PUFA may provide a strategy to mitigate obesity-associated AT inflammation prior to the development of systemic IR and T2D.
Acknowledgments
Danyelle M. Liddle and Amber L. Hutchinson are supported by graduate scholarships from the Natural Sciences and Engineering Research Council of Canada (NSERC).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and obesity: Prevalence, consequences, and causes of a growing public health problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Witherspoon, D.; Latta, L.; Wang, Y.; Black, M.M. Do depression, self-esteem, body-esteem, and eating attitudes vary by BMI among African American adolescents? J. Pediatr. Psychol. 2013, 38, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Brewis, A.A. Stigma and the perpetuation of obesity. Soc. Sci. Med. 2014, 118, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, M.J.; Frontini, R.; Canavarro, M.C.; Moreira, H. Quality of life and psychological functioning in pediatric obesity: The role of body image dissatisfaction between girls and boys of different ages. Qual. Life Res. 2014, 23, 2629–2638. [Google Scholar] [CrossRef] [PubMed]
- Van Geel, M.; Vedder, P.; Tanilon, J. Are overweight and obese youths more often bullied by their peers? A meta-analysis on the correlation between weight status and bullying. Int. J. Obes. 2014, 38, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Landin, K.; Lonnroth, P.; Krotkiewski, M.; Holm, G.; Smith, U. Increased insulin resistance and fat cell lipolysis in obese but not lean women with a high waist/hip ratio. Eur. J. Clin. Investig. 1990, 20, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Nasr, A.; Tfayli, H.; Bacha, F.; Michaliszyn, S.F.; Arslanian, S. Increased lipolysis, diminished adipose tissue insulin sensitivity and impaired β-cell function relative to adipose tissue insulin sensitivity in obese youth with impaired glucose tolerance (IGT). Diabetes 2017, 66, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Maury, E.; Ehala-Aleksejev, K.; Guiot, Y.; Detry, R.; Vandenhooft, A.; Brichard, S.M. Adipokines oversecreted by omental adipose tissue in human obesity. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E656–E665. [Google Scholar] [CrossRef] [PubMed]
- Wentworth, J.M.; Naselli, G.; Brown, W.A.; Doyle, L.; Phipson, B.; Smyth, G.K.; Wabitsch, M.; O’Brien, P.E.; Harrison, L.C. Pro-inflammatory CD11c+CD206+ adipose tissue macrophages are associated with insulin resistance in human obesity. Diabetes 2010, 59, 1648–1656. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, T.; Liu, L.F.; Lamendola, C.; Shen, L.; Morton, J.; Rivas, H.; Winer, D.; Tolentino, L.; Choi, O.; Zhang, H.; et al. T-cell profile in adipose tissue is associated with insulin resistance and systemic inflammation in humans. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2637–2643. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- McArdle, M.A.; Finucane, O.M.; Connaughton, R.M.; McMorrow, A.M.; Roche, H.M. Mechanisms of obesity-induced inflammation and insulin resistance: Insights into the emerging role of nutritional strategies. Front. Endocrinol. 2013, 4, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M. Adipose tissue and adipocyte dysregulation. Diabetes Metab. 2014, 40, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Lolmèd, K.; Duffaut, C.; Zakaroff-Girard, A.; Bouloumié, A. Immune cells in adipose tissue: Key players in metabolic disorders. Diabetes Metab. 2011, 37, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Winer, S.; Winer, D.A. The adaptive immune system as a fundamental regulator of adipose tissue inflammation and insulin resistance. Immunol. Cell Biol. 2012, 90, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Park, Y.J.; Ham, M.; Kim, J.B. Crosstalk between adipocytes and immune cells in adipose tissue inflammation and metabolic dysregulation in obesity. Mol. Cells 2014, 37, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys. Acta 2014, 1842, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Winer, S.; Chan, Y.; Paltser, G.; Truong, D.; Tsui, H.; Bahrami, J.; Dorfman, R.; Wang, Y.; Zielenski, J.; Mastronardi, F.; et al. Normalization of obesity-associated insulin resistance through immunotherapy. Nat. Med. 2009, 15, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Duffaut, C.; Galitzky, J.; Lafontan, M.; Bouloumié, A. Unexpected trafficking of immune cells within the adipose tissue during the onset of obesity. Biochem. Biophys. Res. Commun. 2009, 384, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Travers, R.L.; Motta, A.C.; Betts, J.A.; Bouloumié, A.; Thompson, D. The impact of adiposity on adipose tissue-resident lymphocyte activation in humans. Int. J. Obes. 2014, 39, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S5–S78. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.E.; Buchholz, A.C.; Mazurak, V.C. Inflammation, obesity, and fatty acid metabolism: Influence of n-3 polyunsaturated fatty acids on factors contributing to metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 1008–1024. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.E.; Mazurak, V.C. n-3 polyunsaturated fatty acids: Relationship to inflammation in healthy adults and adults exhibiting features of metabolic syndrome. Lipids 2013, 48, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Mesa, M.D.; Gil, A. Omega-3 long-chain polyunsaturated fatty acids supplementation on inflammatory biomarkers: A systematic review of randomized clinical trials. Br. J. Nutr. 2012, 107 (Suppl. 2), S159–S170. [Google Scholar] [CrossRef] [PubMed]
- Khorson, R.; Crawford, C.; Ives, J.A.; Walter, A.R.; Jonas, W.B. The effect of omega-3 fatty acids on biomarkers of inflammation: A rapid evidence assessment of the literature. Mil. Med. 2014, 179, 2–60. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M.; Spurlock, M.E. Palmitate activates the NF-kappaB transcription factor and induces IL-6 and TNFalpha expression in 3T3-L1 adipocytes. J. Nutr. 2005, 135, 1841–1846. [Google Scholar] [PubMed]
- Weldon, S.; Mullen, A.; Loscher, C.; Hurley, L.; Roche, H. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.L.; Fisher, F.M.; Maratos-Flier, E. Dietary fatty acids differentially regulate production of TNF-alpha and IL-10 by murine 3T3-L1 adipocytes. Obesity 2008, 16, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of EPA and DHA are dependent upon time and dose-response elements associated with LPS stimulation in THP-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Yeop Han, C.; Kargi, A.Y.; Omer, M.; Chan, C.; Wabitsch, M.; O’Brien, K.D.; Wight, T.N.; Chait, A. Differential effect of saturated and unsaturated free fatty acids on the generation of monocyte adhesion and chemotactic factors by adipocytes: Dissociation of adipocyte hypertrophy from inflammation. Diabetes 2010, 59, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Murumalla, R.K.; Gunasekaran, M.K.; Padhan, J.K.; Bencharif, K.; Gence, L.; Festy, F.; Césari, M.; Roche, R.; Hoareau, L. Fatty acids do not pay the toll: Effect of SFA and PUFA on human adipose tissue and mature adipocytes inflammation. Lipids Health Dis. 2012, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Kalupahana, N.S.; Fletcher, S.; Xin, W.; Claycombe, K.J.; Quignard-Boulange, A.; Zhao, L.; Saxton, A.M.; Moustaid-Moussa, N. n-3 and n-6 polyunsaturated fatty acids differentially regulate adipose angiotensinogen and other inflammatory adipokines in part via NF-κB-dependent mechanisms. J. Nutr. Biochem. 2012, 23, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Jiang, W.; Spinetti, T.; Tardivel, A.; Castillo, R.; Bourquin, C.; Guarda, G.; Tian, Z.; Tschopp, J.; Zhou, R. Omega-3 fatty acids prevent inflammation and metabolic disorder through inhibition of NLRP3 inflammasome activation. Immunity 2013, 38, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Cranmer-Byng, M.M.; Liddle, D.M.; De Boer, A.A.; Monk, J.M.; Robinson, L.E. Proinflammatory effects of arachidonic acid in a lipopolysaccharide-induced inflammatory microenvironment in 3T3-L1 adipocytes in vitro. Appl. Physiol. Nutr. Metab. 2015, 40, 142–154. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.A.; Monk, J.M.; Robinson, L.E. Docosahexaenoic acid decreases pro-inflammatory mediators in an in vitro murine adipocyte macrophage co-culture model. PLoS ONE 2014, 9, e85037. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.A.; Monk, J.M.; Liddle, D.M.; Hutchinson, A.L.; Power, K.A.; Ma, D.W.L.; Robinson, L.E. Fish-oil-derived n-3 polyunsaturated fatty acids reduce NLRP3 inflammasome activity and obesity-related inflammatory cross-talk between adipocytes and CD11b(+) macrophages. J. Nutr. Biochem. 2016, 34, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Liddle, D.M.; De Boer, A.A.; Brown, M.J.; Power, K.A.; Ma, D.W.; Robinson, L.E. Fish-oil-derived n-3 PUFAs reduce inflammatory and chemotactic adipokine-mediated cross-talk between co-cultured murine splenic CD8+ T cells and adipocytes. J. Nutr. 2015, 145, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Liddle, D.M.; Monk, J.M.; Hutchinson, A.L.; De Boer, A.A.; Power, K.A.; Robinson, L.E. Fish oil-derived n-3 polyunsaturated fatty acids mitigate lean and obese CD8+ T cell-adipocyte inflammatory cross-talk, in part through a TNFa-dependent mechanism. Appl. Physiol. Nutr. Metab. 2015, 40, S16. [Google Scholar] [CrossRef]
- Liddle, D.M.; Hutchinson, A.L.; Monk, J.M.; Ma, D.W.L.; Power, K.A.; Robinson, L.E. Adipocyte dysfunction induced by inflammatory cross-talk with CD8+ T cells is mitigated by fish oil-derived n-3 polyunsaturated fatty acids, in part through a TNFa-dependent mechanism. Appl. Physiol. Nutr. Metab. 2016, 41, S23–S24. [Google Scholar]
- Liddle, D.M.; Hutchinson, A.L.; Monk, J.M.; Ma, D.W.L.; Robinson, L.E. n-3 polyunsaturated fatty acids mitigate adipocyte-CD4+ T cell inflammatory cross-talk in co-culture models of obese adipose tissue. Appl. Physiol. Nutr. Metab. 2016, 41, S24. [Google Scholar]
- Yang, H.; Youm, Y.H.; Vandanmagsar, B.; Ravussin, A.; Gimble, J.M.; Greenway, F.; Stephens, J.M.; Mynatt, R.L.; Dixit, V.D. Obesity increases the production of proinflammatory mediators from adipose tissue T cells and compromises TCR repertoire diversity: Implications for systemic inflammation and insulin resistance. J. Immunol. 2010, 185, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Bès-Houtmann, S.; Roche, R.; Hoareau, L.; Gonthier, M.P.; Festy, F.; Caillens, H.; Gasque, P.; Lefebvre d’Hellencourt, C.; Cesari, M. Presence of functional TLR2 and TLR4 on human adipocytes. Histochem. Cell Biol. 2006, 127, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Kopp, A.; Buechler, C.; Neumeier, M.; Weigert, J.; Aslanidis, C.; Scholmerich, J.; Schaffler, A. Innate immunity and adipocyte function: Ligand-specific activation of multiple Toll-like receptors modulates cytokine, adipokine, and chemokine secretion in adipocytes. Obesity 2009, 17, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Deng, T.; Peterson, L.E.; Yu, R.; Lin, J.; Hamilton, D.J.; Reardon, P.R.; Sherman, V.; Winnier, G.E.; Zhan, M.; et al. Transcriptome analysis of human adipocytes implicates the NOD-like receptor pathway in obesity induced adipose inflammation. Mol. Cell. Endocrinol. 2014, 394, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Nagai, Y.; Takatsu, K. Activation and regulation of the pattern recognition receptors in obesity-induced adipose tissue inflammation and insulin resistance. Nutrients 2013, 5, 3757–3778. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose tissue remodeling in pathophysiology of obesity. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. 2016, 7, 1541. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wood, I.S.; Trayhurn, P. Dysregulation of the expression and secretion of inflammation-related adipokines by hypoxia in human adipocytes. Pflugers Arch. 2007, 455, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Wood, I.S.; Stezhka, T.; Trayhurn, P. Modulation of adipokine production, glucose uptake and lactate release in human adipocytes by small changes in oxygen tension. Pflugers Arch. 2011, 462, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Tchernof, A.; Despres, J.P. Pathophysiology of human visceral obesity: An update. Physiol. Rev. 2013, 93, 359–404. [Google Scholar] [CrossRef] [PubMed]
- Kranendonk, M.E.; van Herwaarden, J.A.; Stupkova, T.; de Jager, W.; Vink, A.; Moll, F.L.; Kalkhoven, E.; Visseren, F.L. Inflammatory characteristics of distinct abdominal adipose tissue depots relate differently to metabolic risk factors for cardiovascular disease: Distinct fat depots and vascular risk factors. Atherosclerosis 2015, 239, 419–427. [Google Scholar] [CrossRef] [PubMed]
- McQuaid, S.N.; Humphreys, S.M.; Hodson, L.; Fielding, B.A.; Karpe, F.; Frayn, K.N. Femoral adipose tissue may accumulate the fat that has been recycled as VLDL and nonesterified fatty acids. Diabetes 2010, 59, 2465–2473. [Google Scholar] [CrossRef] [PubMed]
- Manolopoulos, K.N.; Karpe, F.; Frayn, K.N. Gluteofemoral body fat as a determinant of metabolic health. Int. J. Obes. 2010, 34, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Laurencikiene, J.; Skurk, T.; Kulyté, A.; Hedén, P.; Åström, G.; Sjölin, E.; Rydén, M.; Hauner, H.; Arner, P. Regulation of lipolysis in small and large fat cells of the same subject. J. Clin. Endocrinol. Metab. 2011, 96, E2045–E2049. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Plakidas, A.; Lee, W.H.; Heikkinen, A.; Chanmugam, P.; Bray, G.; Hwang, D.H. Differential modulation of Toll-like receptors by fatty acids: Preferential inhibition by n-3 polyunsaturated fatty acids. J. Lipid Res. 2003, 44, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Nishida, J.; Ogawa, Y. A paracrine loop between adipocytes and macrophages aggravates inflammatory changes: Role of free fatty acids and tumor necrosis factor alpha. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid–induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Song, M.J.; Kim, K.H.; Yoon, J.M.; Kim, J.B. Activation of Toll-like receptor 4 is associated with insulin resistance in adipocytes. Biochem. Biophys. Res. Commun. 2006, 346, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Favelyukis, S.; Nguyen, A.K.; Reichart, D.; Scott, P.A.; Jenn, A.; Liu-Bryan, R.; Glass, C.K.; Neels, J.G.; Olefsky, J.M. A subpopulation of macrophages infiltrates hypertrophic adipose tissue and is activated by free fatty acids via toll-like receptors 2 and 4 and JNK-dependent pathways. J. Biol. Chem. 2007, 282, 35279–35292. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Chen, Q.; Shah, S.; Du, J.; Tao, B.; Tzameli, I.; Yan, W.; Xu, H. Obesity-related upregulation of monocyte chemotactic factors in adipocytes: Involvement of nuclear factor-kappaB and c-Jun NH2-terminal kinase pathways. Diabetes 2009, 58, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Schaeffler, A.; Gross, P.; Buettner, R.; Bollheimer, C.; Buechler, C.; Neumeier, M.; Kopp, A.; Schoelmerich, J.; Falk, W. Fatty acid-induced induction of toll-like receptor-4/nuclear factor-kappaB pathway in adipocytes links nutritional signalling with innate immunity. Immunology 2009, 126, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, R.G.; Huang, S.; Choi, I.W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-mediated secretion of IL-1 in human monocytes through TLR2 activation; modulation by dietary fatty acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.; Brickey, W.J.; Ting, J.P. Fatty acid–induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Joe, Y.; Jeong, S.O.; Zheng, M.; Back, S.H.; Park, S.W.; Ryter, S.W.; Chung, H.T. Endoplasmic reticulum stress is sufficient for the induction of IL-1β production via activation of the NF-κB and inflammasome pathways. Innate Immun. 2014, 20, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Osborn, O.; Olefsky, J.M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med. 2012, 18, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Labbé, S.M.; Croteau, E.; Grenier-Larouche, T.; Frisch, F.; Ouellet, R.; Langlois, R.; Guérin, B.; Turcotte, E.E.; Carpentier, A.C. Normal postprandial nonesterified fatty acid uptake in muscles despite increased circulating fatty acids in type 2 diabetes. Diabetes 2011, 60, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Grapov, D.; Adams, S.H.; Pedersen, T.L.; Garvey, W.T.; Newman, J.W. Type 2 diabetes associated changes in the plasma non-esterified fatty acids, oxylipins and endocannabinoids. PLoS ONE 2012, 7, e48852. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.J.; Skrtic, S.; Katsogiannos, P.; Abrahamsson, N.; Sidibeh, C.O.; Dahgam, S.; Mansson, M.; Risérus, U.; Kullberg, J.; Eriksson, J.W. Impaired adipose tissue lipid storage, but not altered lipolysis, contributes to elevated levels of NEFA in type 2 diabetes. Degree of hyperglycemia and adiposity are important factors. Metabolism 2016, 65, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Samaras, K.; Botelho, N.K.; Chisholm, D.J.; Lord, R.V. Subcutaneous and visceral adipose tissue gene expression of serum adipokines that predict type 2 diabetes. Obesity 2010, 18, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Nitta, C.F.; Orlando, R.A. Crosstalk between immune cells and adipocytes requires both paracrine factors and cell contact to modify cytokine secretion. PLoS ONE 2013, 8, e77306. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P.J. Impact of high-fat diet on the intestinal microbiota and small intestinal physiology before and after the onset of obesity. Biochimie 2017, 141, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.N.; McGillicuddy, F.C.; Anderson, P.D.; Hinkle, C.C.; Shah, R.; Pruscino, L.; Tabita-Martinez, J.; Sellers, K.F.; Rickels, M.R.; Reilly, M.P.; et al. Experimental endotoxemia induces adipose inflammation and insulin resistance in humans. Diabetes 2010, 59, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Backhed, F. Functional interactions between microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Luck, H.; Tsai, S.; Winer, S. The intestinal immune system in obesity and insulin resistance. Cell Metab. 2016, 23, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 fatty acids correlate with gut microbiome diversity and production of N-carbamylglutamate in middle aged and elderly women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef] [PubMed]
- Noriega, B.S.; Sanchez-Gonzalez, M.A.; Salyakina, D.; Coffman, J. Understanding the impact of omega-3 rich diet on the gut microbiota. Case Rep. Med. 2016, 2016, 3089303. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Cani, P.D. Interaction between obesity and the gut microbiota: Relevance in nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, A.; Collado, M.C.; Garcia-Valdes, L.; Segura, M.T.; Martin-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Wittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [PubMed]
- Rahat-Rozenbloom, S.; Fernandes, J.; Gloor, G.B.; Wolever, T.M. Evidence for greater production of colonic short-chain fatty acids in overweight than lean humans. Int. J. Obes. 2014, 38, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Ruiz, A.; Lanza, F.; Haange, S.B.; Oberbach, A.; Till, H.; Bargiela, R.; Campoy, C.; Segura, M.T.; Richter, M.; et al. Microbiota from the distal guts of lean and obese adolescents exhibit partial functional redundancy besides clear differences in community structure. Environ. Microbiol. 2013, 15, 211–226. [Google Scholar] [CrossRef]
- Shen, J.; Obin, M.S.; Zhao, L. The gut microbiota, obesity and insulin resistance. Mol. Aspects Med. 2013, 34, 39–58. [Google Scholar] [CrossRef] [PubMed]
- Salden, B.N.; Troost, F.J.; Wilms, E.; Truchado, P.; Vilchez-Vargas, R.; Pieper, D.H.; Jáuregui, R.; Marzorati, M.; van de Wiele, T.; Possemiers, S.; et al. Reinforcement of intestinal epithelial barrier by arabinoxylans in overweight and obese subjects: A randomized controlled trial: Arabinoxylans in gut barrier. Clin. Nutr. 2017, S026–S5614, 30048-1. [Google Scholar] [CrossRef]
- Troseid, M.; Nestvold, T.K.; Rudi, K.; Thoresen, H.; Nielsen, E.W.; Lappegard, K.T. Plasma lipopolysaccharide is closely associated with glycemic control and abdominal obesity: Evidence from bariatric surgery. Diabetes Care 2013, 36, 3627–3632. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.; Kassis, A.; Major, G.; Chou, C.J. Is the gut microbiota a new factor contributing to obesity and its metabolic disorders? J. Obes. 2012, 2012, 879151. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Crosstalk between the gut microbiota and the endocannabinoid system: Impact on the gut barrier function and the adipose tissue. Clin. Microbiol. Infect. 2012, 18 (Suppl. 4), 50–53. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.F.; Cotter, P.D.; Healy, S.; Marques, T.M.; O’Sullivan, O.; Fouhy, F.; Clarke, S.F.; O’Toole, P.W.; Quigley, E.M.; Stanton, C.; et al. Composition and energy harvesting capacity of the gut microbiota: Relationship to diet, obesity and time in mouse models. Gut 2010, 59, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Sanmiguel, C.; Gupta, A.; Mayer, E.A. Gut microbiome and obesity: A plausible explanation for obesity. Curr. Obes. Rep. 2015, 4, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.M.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Castagliuolo, I.; Leo, V.D.; Buda, A.; Pinzani, M.; Palù, G.; Martines, D. Increased intestinal permeability in obese mice: New evidence in the pathogenesis of nonalcoholic steatohepatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G518–G525. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Delzenne, N.M. Interplay between obesity and associated metabolic disorders: New insights into the gut microbiota. Curr. Opin. Pharmacol. 2009, 9, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, W.C.; He, C.Y.; Han, Z.; Jin, D.Y.; Wang, L. Change of intestinal mucosa barrier function in the progress of non-alcoholic steatohepatitis in rats. World J. Gastroenterol. 2008, 14, 3254–3258. [Google Scholar] [CrossRef] [PubMed]
- Caesar, R.; Tremaroli, V.; Kovatcheva-Datchary, P.; Cani, P.D.; Backhed, F. Crosstalk between gut microbiota and dietary lipids aggravates WAT inflammation through TLR signaling. Cell Metab. 2015, 22, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Muzio, M.; Polentarutti, N.; Bosisio, D.; Manoj Kumar, P.P.; Mantovani, A. Toll-like receptor family and signalling pathway. Biochem. Soc. Trans. 2000, 28, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lee, H.; Berg, A.H.; Lisanti, M.P.; Shapiro, L.; Scherer, P.E. The lipopolysaccharide-activated toll-like receptor (TLR)-4 induces synthesis of the closely related receptor TLR-2 in adipocytes. J. Biol. Chem. 2000, 275, 24255–24263. [Google Scholar] [CrossRef] [PubMed]
- Meijer, K.; de Vries, M.; Al-Lahham, S.; Bruinenberg, M.; Weening, D.; Dijkstra, M.; Kloosterhuis, N.; van der Leij, R.J.; van der Want, H.; Kroesen, B.J.; et al. Human primary adipocytes exhibit immune cell function: Adipocytes prime inflammation independent of macrophages. PLoS ONE 2011, 6, e17154. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Obesity 2008, 16, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Li, P.; Huh, J.Y.; Hwang, I.J.; Lu, M.; Kim, J.I.; Ham, M.; Talukdar, S.; Chen, A.; Lu, W.J.; et al. Inflammation is necessary for long-term but not short-term high-fat diet–induced insulin resistance. Diabetes 2011, 60, 2474–2483. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. The relationship between the fatty acid composition of immune cells and their function. Prostaglandins Leukot. Essent. Fatty Acids 2008, 79, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed]
- Turk, H.F.; Chapkin, R.S. Membrane lipid raft organization is uniquely modified by n-3 polyunsaturated fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Ly, L.H.; Barhoumi, R.; McMurray, D.N.; Chapkin, R.S. Dietary docosahexaenoic acid suppresses T cell protein kinase C theta lipid raft recruitment and IL-2 production. J. Immunol. 2004, 173, 6151–6160. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.W.; Kwon, M.J.; Choi, A.M.; Kim, H.P.; Nakahira, K.; Hwang, D.H. Fatty acids modulate toll-like receptor 4 activation through regulation of receptor dimerization and recruitment into lipid rafts in a reactive oxygen species-dependent manner. J. Biol. Chem. 2009, 284, 27384–27392. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Usui, I.; Bukhari, A.; Ikutani, M.; Oya, T.; Kanatani, Y.; Tsuneyama, K.; Nagai, Y.; Takatsu, K.; Urakaz, M.; et al. Regulatory mechanisms for adipose tissue M1 and M2 macrophages in diet-induced obese mice. Diabetes 2009, 58, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; DeYoung, S.M.; Bodzin, J.L.; Saltiel, A.R. Increased inflammatory properties of adipose tissue macrophages recruited during diet-induced obesity. Diabetes 2006, 56, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Koh, V.; Kua, L.F.; So, J.; Davide, L.; Lim, K.S.; Petersen, S.H.; Yong, W.P.; Shabbir, A.; Kono, K. Accumulation of CD11c+CD163+ adipose tissue macrophages through upregulation of intracellular 11β-HSD1 in human obesity. J. Immunol. 2016, 197, 3735–3745. [Google Scholar] [CrossRef] [PubMed]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A.; Lee, J.; Goldfine, A.B.; Benoist, C.; Shoelson, S.; Mathis, D. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Nunez Ruiz, A.; Cortes-Garcia, J.D.; Cortez-Espinosa, N.; Herrera-Rojas, P.I.; Ruiz-Rodriguez, V.M.; Salgado-Bustamante, M.; García-Hernández, M.H.; Reynaga-Hernández, E.; Martínez-Jimenez Vdel, C.; Portales-Pérez, D.P. Diminished levels of regulatory T cell subsets (CD8+Foxp3, CD4+Foxp3 and CD4+CD39+Foxp3) but increased Foxp3 expression in adipose tissue from overweight subjects. Nutrition 2016, 32, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Gyllenhammer, L.E.; Lam, J.; Alderete, T.L.; Allayee, H.; Akbari, O.; Katkhouda, N.; Goran, M.I. Lower omental T-regulatory cell count is associated with higher fasting glucose and lower β-cell function in adults with obesity. Obesity 2016, 24, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Ciucci, T.; Rousseau, D.; Bourlier, V.; Duffaut, C.; Bonnafous, S.; Blin-Wakkach, C.; Anty, R.; Iannelli, A.; Gugenheim, J.; et al. Identification of adipose tissue dendritic cells correlated with obesity-associated insulin-resistance and inducing Th17 responses in mice and patients. Diabetes 2012, 61, 2238–2247. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.W.; Zamarron, B.F.; Muir, L.A.; Singer, K.; Porsche, C.E.; DelProposto, J.B.; Geletka, L.; Meyer, K.A.; O’Rourke, R.W.; Lumeng, C.N. Adipose tissue dendritic cells are independent contributors to obesity-induced inflammation and insulin resistance. J. Immunol. 2016, 197, 3650–3661. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N.; et al. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat. Med. 2011, 17, 610–617. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.E.; Ganley-Leal, L.M.; Mehta, A.; Bigornia, S.J.; Mott, M.; Rehman, Q.; Farb, M.G.; Hess, D.T.; Joseph, L.; Gokce, N.; et al. B lymphocytes in human subcutaneous adipose crown-like structures. Obesity 2012, 20, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, K.; Ishimori, N.; Ohmura, Y.; Tokuhara, S.; Nozawa, A.; Horii, S.; Andoh, Y.; Fujii, S.; Iwabuchi, K.; Onoé, K.; et al. Natural killer T cells are involved in adipose tissues inflammation and glucose intolerance in diet-induced obese mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 193–199. [Google Scholar] [CrossRef] [PubMed]
- O'Rourke, R.W.; White, A.E.; Metcalf, M.D.; Winters, B.R.; Diggs, B.S.; Zhu, X.; Marks, D.L. Systemic inflammation and insulin sensitivity in obese IFN-γ knockout mice. Metabolism 2012, 61, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Wensveen, F.M.; Jelenčić, V.; Valentić, S.; Šestan, M.; Wensveen, T.T.; Theurich, S.; Glasner, A.; Mendrila, D.; Stimac, D.; Wunderlich, F.T.; et al. NK cells link obesity-induced adipose stress to inflammation and insulin resistance. Nat. Immunol. 2015, 16, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Rocha, V.Z.; Folco, E.J.; Sukhova, G.; Shimizu, K.; Gotsman, I.; Vernon, A.H.; Libby, P. Interferon-gamma, a Th1 cytokine, regulates fat inflammation: A role for adaptive immunity in obesity. Circ. Res. 2008, 103, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Strissel, K.J.; DeFuria, J.; Shaul, M.E.; Bennett, G.; Greenberg, A.S.; Obin, M.S. T-cell recruitment and Th1 polarization in adipose tissue during diet-induced obesity in C57BL6 mice. Obesity 2010, 18, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Duffaut, C.; Zakaroff-Girard, A.; Bourlier, V.; Decaunes, P.; Maumus, M.; Chiotasso, P.; Sengenès, C.; Ladontan, M.; Galitzky, J.; Bouloumié, A. Interplay between human adipocytes and T lymphocytes in obesity: CCL20 as an adipochemokine and T lymphocytes as lipogenic modulators. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Himes, R.W.; Smith, C.W. Tlr2 is critical for diet-induced metabolic syndrome in a murine model. FASEB J. 2010, 24, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.E.; Braucher, D.R.; Walker-Daniels, J.; Spurlock, M.E. Absence of Tlr2 protects against high-fat diet-induced inflammation and results in greater insulin-stimulated glucose transport in cultured adipocytes. J. Nutr. Biochem. 2011, 22, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting Edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, P.; Qi, J.; Zhang, L.; Gao, C. TLR-induced NF-κB activation regulates NLRP3 expression in murine macrophages. FEBS Lett. 2012, 586, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Hoch, M.; Eberle, A.N.; Peterli, R.; Peters, T.; Seboek, D.; Keller, U.; Muller, B.; Linscheid, P. LPS induces interleukin-6 and interleukin-8 but not tumor necrosis factor-alpha in human adipocytes. Cytokine 2008, 41, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S.; Franceschetti, G.; Zoico, E.; Bambace, C.; Cominacini, L.; Zamboni, M. LPS response pattern of inflammatory adipokines in an in vitro 3T3-L1 murine adipocyte model. Inflamm. Res. 2014, 63, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Cullberg, K.B.; Larsen, J.O.; Pedersen, S.B.; Richelsen, B. Effects of LPS and dietary free fatty acids on MCP-1 in 3T3-L1 adipocytes and macrophages in vitro. Nutr. Diabetes 2014, 4, e113–e115. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Joshi, N.S.; Liu, Y.; Meng, H.; Kleinstein, S.H.; Kaech, S.M. TLR4 ligands lipopolysaccharide and monophosphoryl lipid A differentially regulate effector and memory CD8 T cell differentiation. J. Immunol. 2014, 192, 4221–4232. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Liddle, D.M.; Cohen, D.J.; Tsang, D.H.; Hillyer, L.M.; Abdelmagid, S.A.; Nakamura, M.T.; Power, K.A.; Ma, D.W.; Robinson, L.E. The delta 6 desaturase knock out mouse reveals that immunomodulatory effects of essential n-6 and n-3 polyunsaturated fatty acids are both independent of and dependent upon conversion. J. Nutr. Biochem. 2016, 32, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Elgazar-Carmon, V.; Rudich, A.; Hadad, N.; Levy, R. Neutrophils transiently infiltrate intra-abdominal fat early in the course of high-fat feeding. J. Lipid Res. 2008, 49, 1894–1903. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Molofsky, A.B.; Liang, H.E.; Ricardo-Gonzalez, R.R.; Jouihan, H.A.; Bando, J.K.; Chawla, A.; Locksley, R.M. Eosinophils sustain adipose alternatively activated macrophages associated with glucose homeostasis. Science 2011, 332, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Divoux, A.; Sun, J.; Zhang, J.; Clement, K.; Glickman, J.N.; Sukhova, G.K.; Wolters, P.J.; Du, J.; Gorgun, C.Z.; et al. Genetic deficiency and pharmacological stabilization of mast cells reduce diet-induced obesity and diabetes in mice. Nat. Med. 2009, 15, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A. Control of macrophage activation and function by PPARs. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Prieur, X.; Mok, C.; Velagapudi, V.R.; Núñez, V.; Fuentes, L.; Montaner, D.; Ishikawa, K.; Camacho, A.; Barbarroja, N.; O’Rahilly, S.; et al. Differential lipid partitioning between adipocytes and tissue macrophages modulates macrophage lipotoxicity and M2/M1 polarization in obese mice. Diabetes 2011, 60, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.A.; Underhill, D.M.; Sweet, M.J.; Ozinsky, A.O.; Liew, F.Y.; Aderem, A. Macrophages exposed continuously to lipopolysaccharide and other agonists that act via toll-like receptors exhibit a sustained and additive activation state. BMC Immunol. 2001, 2, 1–12. [Google Scholar] [CrossRef]
- Jones, E.; Adcock, I.M.; Ahmed, B.Y.; Punchard, N.A. Modulation of LPS stimulated NF-kappaB mediated nitric oxide production by PKCε and JAK2 in RAW macrophages. J. Inflamm. 2007, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, C.; Razzak, A.; Babcock, T.A.; Helton, W.S.; Espat, N.J. Lipopolysaccharide-stimulated RAW 264.7 macrophage inducible nitric oxide synthase and nitric oxide production is decreased by an omega-3 fatty acid lipid emulsion. J. Surg. Res. 2008, 149, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Madi, M.; Ding, C.; Fok, M.; Steele, T.; Ford, C.; Hunter, L.; Bing, C. Interleukin-1 mediates macrophage-induced impairment of insulin signaling in human primary adipocytes. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E289–E304. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; DeYoung, S.M.; Saltiel, A.R. Macrophages block insulin action in adipocytes by altering expression of signaling and glucose transport proteins. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E166–E174. [Google Scholar] [CrossRef] [PubMed]
- Oliver, E.; McGillicuddy, F.C.; Harford, K.A.; Reynolds, C.M.; Phillips, C.M.; Ferguson, J.F.; Roche, H.M. Docosahexaenoic acid attenuates macrophage-induced inflammation and improves insulin sensitivity in adipocytes-specific differential effects between LC n-3 PUFA. J. Nutr. Biochem. 2012, 23, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Young-Bum, K.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.; Yao-Borengasser, A.; Unal, R.; Rasouli, N.; Gurley, C.M.; Zhu, B.; Peterson, C.A.; Kern, P.A. Adipose tissue macrophages in insulin-resistant subjects are associated with collagen VI and fibrosis and demonstrate alternative activation. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E1016–E1027. [Google Scholar] [CrossRef] [PubMed]
- Patsouris, D.; Li, P.P.; Thapar, D.; Chapman, J.; Olefsky, J.M.; Neels, J.G. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab. 2008, 8, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Perrard, X.D.; Wang, Q.; Perrard, J.L.; Polsani, V.R.; Jones, P.H.; Smith, C.W.; Ballantyne, C.M. CD11c expression in adipose tissue and blood and its role in diet-induced obesity. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Ariel, A.; Serhan, C.N. New lives given by cell death: Macrophage differentiation following their encounter with apoptotic leukocytes during the resolution of inflammation. Front. Immunol. 2012, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lu, M.; Nguyen, M.T.; Bae, E.J.; Chapman, J.; Feng, D.; Hawkins, M.; Pessin, J.E.; Sears, D.D.; Nguyen, A.K.; et al. Functional heterogeneity of CD11c-positive adipose tissue macrophages in diet-induced obese mice. J. Biol. Chem. 2010, 285, 15333–15345. [Google Scholar] [CrossRef] [PubMed]
- Shaul, M.E.; Bennett, G.; Strissel, K.J.; Greenberg, A.S.; Obin, M.S. Dynamic, M2-like remodeling phenotypes of CD11c+ adipose tissue macrophages during high-fat diet-induced obesity in mice. Diabetes 2010, 59, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Tordjman, J.; Poitou, C.; Darakhshan, F.; Hugol, D.; Basdevant, A.; Aissat, A.; Guerre-Millo, M.; Clément, K. Human adipose tissue macrophages: M1 and M2 cell surface markers in subcutaneous and omental depots and after weight loss. J. Clin. Endocrinol. Metab. 2009, 94, 4619–4623. [Google Scholar] [CrossRef] [PubMed]
- Bourlier, V.; Zakaroff-Girard, A.; Miranville, A.; De Barros, S.; Maumus, M.; Sengenes, C.; Galitzky, J.; Ladontan, M.; Karpe, F.; Frayn, K.N.; et al. Remodeling phenotype of human subcutaneous adipose tissue macrophages. Circulation 2008, 117, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Farmer, D.; Todoric, J.; Aszmann, O.; Speiser, M.; Györi, G.; Zlabinger, G.J.; Stulnig, T.M. Human adipose tissue macrophages are of an anti-inflammatory phenotype but capable of excessive pro-inflammatory mediator production. Int. J. Obes. 2007, 31, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Kalupahana, N.S.; Claycombe, K.; Newman, S.J.; Stewart, T.; Siriwardhana, N.; Matthan, N.; Lichtenstein, A.H.; Moustaid-Moussa, N. Eicosapentaenoic acid prevents and reverses insulin resistance in high-fat diet-induced obese mice via modulation of adipose tissue inflammation. J. Nutr. 2010, 140, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, T.; Worsch, S.; Heikenwalder, M.; Daniel, H.; Hauner, H.; Bader, B.L. Metabolic and immunomodulatory effects of n-3 fatty acids are different in mesenteric and epididymal adipose tissue of diet-induced obese mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1140–E1156. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Hou, T.Y.; Turk, H.F.; Weeks, B.; Wu, C.; McMurray, D.N.; Chapkin, R.S. Dietary n-3 polyunsaturated fatty acids (PUFA) decrease obesity-associated Th17 cell-mediated inflammation during colitis. PLoS ONE 2012, 7, e49739. [Google Scholar] [CrossRef] [PubMed]
- Titos, E.; Rius, B.; Gonzalez-Periz, A.; Lopez-Vicario, C.; Moran-Salvador, E.; Martinez-Clemente, M.; Arroyo, V.; Clària, J. Resolvin D1 and its precursor docosahexaenoic acid promote resolution of adipose tissue inflammation by eliciting macrophage polarization toward an M2-Like phenotype. J. Immunol. 2011, 187, 5408–5418. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Löffler, M.; Huber, J.; Bilban, M.; Reimers, M.; Kadl, A.; Zeyda, M.; Waldhäusl, W.; Stulnig, T.M. Adipose tissue inflammation induced by high-fat diet in obese diabetic mice is prevented by n−3 polyunsaturated fatty acids. Diabetologia 2006, 49, 2109–2119. [Google Scholar] [CrossRef] [PubMed]
- Talamonti, E.; Pauter, A.M.; Asadi, A.; Fischer, A.W.; Chiurchiù, V.; Jacobsson, A. Impairment of systemic DHA synthesis affects macrophage plasticity and polarization: Implications for DHA supplementation during in ammation. Cell. Mol. Life Sci. 2017, 74, 2815–2826. [Google Scholar] [CrossRef] [PubMed]
- Belchior, T.; Paschoal, V.A.; Magdalon, J.; Chimin, P.; Farias, T.M.; Chaves-Filho, A.B.; Gorjão, R.; St-Pierre, P.; Miyamoto, S.; Kang, J.X.; et al. Omega-3 fatty acids protect from diet-induced obesity, glucose intolerance, and adipose tissue inflammation through PPARγ-dependent and PPARγ-independent actions. Mol. Nutr. Food Res. 2015, 59, 957–967. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Arita, M.; Taguchi, R.; Kang, J.X.; Marette, A. Transgenic restoration of long-chain n-3 fatty acids in insulin target tissues improves resolution capacity and alleviates obesity-linked inflammation and insulin resistance in high-fat-fed mice. Diabetes 2010, 59, 3066–3073. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.; Finlin, B.S.; Unal, R.; Zhu, B.; Morris, A.J.; Shipp, L.R.; Lee, J.; Walton, R.G.; Adu, A.; Erfani, R.; et al. Omega-3 fatty acids reduce adipose tissue macrophages in human subjects with insulin resistance. Diabetes 2013, 62, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Hames, K.C.; Morgan-Bathke, M.; Harteneck, D.A.; Zhou, L.; Port, J.D.; Lanza, I.R.; Jensen, M.D. Very-long-chain ω-3 fatty acid supplements and adipose tissue functions: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 1552–1558. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Lee, H.N.; Kim, W.; Surh, Y.J. Docosahexaenoic acid induces M2 macrophage polarization through peroxisome proliferator-activated receptor γ activation. Life Sci. 2015, 120, 39–47. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.A.; Monk, J.M.; Liddle, D.M.; Power, K.A.; Ma, D.W.L.; Robinson, L.E. Fish oil-derived long-chain n-3 polyunsaturated fatty acids reduce expression of M1-associated macrophage markers in an ex vivo adipose tissue culture model, in part through adiponectin. Front. Nutr. 2015, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Rombaldova, M.; Janovska, P.; Kopecky, J.; Kuda, O. Omega-3 fatty acids promote fatty acid utilization and production of pro-resolving lipid mediators in alternatively activated adipose tissue macrophages. Biochem. Biophys. Res. Commun. 2017, 490, 1080–1085. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Bannenberg, G.; Serhan, C.N. Specialized pro-resolving lipid mediators in the inflammatory response: An update. Biochim. Biophys. Acta 2010, 1801, 1260–7123. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.; Tang, Y.; Kosuri, M.; Bhatnagar, A.; Spite, M. Resolvin D1 decreases adipose tissue macrophage accumulation and improves insulin sensitivity in obese-diabetic mice. FASEB J. 2011, 25, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Titos, E.; Rius, B.; Lopez-Vicario, C.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Lopategi, A.; Dalli, J.; Lozano, J.J.; Arroyo, V.; Delgado, S.; et al. Signaling and immunoresolving actions of resolvin D1 in inflamed human visceral adipose tissue. J. Immunol. 2016, 197, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Claria, J.; Dalli, J.; Yacoubian, S.; Gao, F.; Serhan, C.N. Resolvin D1 and Resolvin D2 govern local inflammatory tone in obese fat. J. Immunol. 2012, 189, 2597–2605. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, L.; González-Muniesa, P.; Laiglesia, L.M.; Sáinz, N.; Prieto-Hontoria, P.L.; Escoté, X.; Odriozola, L.; Corrales, F.J.; Arbones-Mainar, J.M.; Martínez, J.A.; et al. Maresin 1 improves insulin sensitivity and attenuates adipose tissue inflammation in ob/ob and diet-induced obese mice. FASEB J. 2017, 31, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ghosh, S.; Perrard, X.D.; Feng, L.; Garcia, G.E.; Perrard, J.L.; Sweeney, J.F.; Peterson, L.E.; Chan, L.; Smith, C.W.; et al. T-Cell accumulation and regulated on activation, normal T cell expressed and secreted upregulation in adipose tissue in obesity. Circulation 2007, 115, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Kintscher, U.; Hartge, M.; Hess, K.; Foryst-Ludwig, A.; Clemenz, M.; Wabitsch, M.; Fischer-Posovszky, P.; Barth, T.F.; Dragun, D.; Skurk, T.; et al. T-lymphocyte infiltration in visceral adipose tissue: A primary event in adipose tissue inflammation and the development of obesity-mediated insulin resistance. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Rausch, M.E.; Weisberg, S.; Vardhana, P.; Tortoriello, D.V. Obesity in C57BL/6J mice is characterized by adipose tissue hypoxia and cytotoxic T-cell infiltration. Int. J. Obes. 2008, 32, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Jiang, E.; Perrard, X.D.; Yang, D.; Khan, I.M.; Perrard, J.L.; Smith, C.W.; Ballantyne, C.M.; Wu, H. Essential role of CD11a in CD8+ T-cell accumulation and activation in adipose tissue. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.M.; Perrard, X.Y.; Perrard, J.L.; Mansoori, A.; Smith, C.W.; Wu, H.; Ballantyne, C.M. Attenuated adipose tissue and skeletal muscle inflammation in obese mice with combined CD4+ and CD8+ T cell deficiency. Atherosclerosis 2014, 233, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Seo, K.; Yamashita, H.; Hosoya, Y.; Ohsugi, M.; Tobe, K.; Kadowaki, T.; Nagai, R.; et al. In vivo imaging in mice reveals local cell dynamics and inflammation in obese adipose tissue. J. Clin. Investig. 2008, 118, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Poggi, M.; Engel, D.; Christ, A.; Beckers, L.; Wijnands, E.; Boon, L.; Driessen, A.; Cleutjens, J.; Weber, C.; Gerdes, N.; et al. CD40L deficiency ameliorates adipose tissue inflammation and metabolic manifestations of obesity in mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Cho, K.W.; DelProposto, J.L.; Oatmen, K.E.; Geletka, L.M.; Martinez-Santibanez, G.; Singer, K.; Lumeng, C.N. Adipose tissue macrophages function as antigen-presenting cells and regulate adipose tissue CD4+ T cells in mice. Diabetes 2013, 62, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Oestreich, K.J.; Weinmann, A.S. Master regulators or lineage-specifying? Changing views on CD4+ T cell transcription factors. Nat. Rev. Immunol. 2012, 12, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Deng, T.; Lyon, C.J.; Minze, L.J.; Lin, J.; Zou, J.; Liu, J.Z.; Ren, Y.; Yin, Z.; Hamilton, D.J.; Reardon, P.R.; et al. Class II major histocompatibility complex plays an essential role in obesity-induced adipose inflammation. Cell Metab. 2013, 17, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.; Shah, Z.; Shah, N.; Needleman, B.; Mikami, D.; Narula, V.; Perry, K.; Hazey, J.; Kampfrath, T.; Kollengode, M.; et al. Visceral adipose inflammation in obesity is associated with critical alterations in tregulatory cell numbers. PLoS ONE 2011, 6, e16376. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; L’homme, L.; De Roover, A.; Kohnen, L.; Scheen, A.J.; Moutschen, M.; Piette, J.; Legrand-Poels, S.; Paquot, N. Obesity phenotype is related to NLRP3 inflammasome activity and immunological profile of visceral adipose tissue. Diabetologia 2013, 56, 2487–2497. [Google Scholar] [CrossRef] [PubMed]
- Poggi, M.; Kara, I.; Brunel, J.M.; Landrier, J.F.; Govers, R.; Bonardo, B.; Fluhrer, R.; Haass, C.; Alessi, M.C.; Peiretti, F. Palmitoylation of TNF alpha is involved in the regulation of TNF receptor 1 signalling. Biochim. Biophys. Acta 2013, 1833, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Festa, A.; D’Agostino, R.; Tracy, R.P.; Haffner, S.M. Elevated levels of acute-phase proteins and plasminogen activator inhibitor-1 predict the development of type 2 diabetes: The insulin resistance atherosclerosis study. Diabetes 2002, 51, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.J.; Norrie, J.; Caslake, M.J.; Gaw, A.; Ford, I.; Lowe, G.D.; O’Reilly, D.S.; Packard, C.J.; Sattar, N. C-reactive protein is an independent predictor of risk for the development of diabetes in the West of Scotland Coronary Prevention Study. Diabetes 2002, 51, 1596–1600. [Google Scholar] [CrossRef] [PubMed]
- Lord, G.M.; Matarese, G.; Howard, J.K.; Baker, R.J.; Bloom, S.R.; Lechler, R.I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998, 394, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Cella, M.; McCartney, S.A.; Fuchs, A.; Abumrad, N.A.; Pietka, T.A.; Chen, Z.; Finck, B.N.; Han, D.H.; Magkos, F.; et al. Association between specific adipose tissue CD4+ T-cell populations and insulin resistance in obese individuals. Gastroenterology 2013, 145, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J.B.; Ferraro, A.A.; Sananez, I.; Gancedo, M.C.; Baz, P.; Billordo, L.A.; Fainboim, L.; Arruvito, L. ATP-induced inflammation drives tissue-resident Th17 cells in metabolically unhealthy obesity. J. Immunol. 2016, 196, 3287–3296. [Google Scholar] [CrossRef] [PubMed]
- Łuczyński, W.; Grubczak, K.; Moniuszko, M.; Głowińska-Olszewska, B.; Bossowski, A. Elevated levels of Th17 cells in children with central obesity. Scand. J. Clin. Lab. Invest. 2015, 75, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Sumarac-Dumanovic, M.; Stevanovic, D.; Ljubic, A.; Jorga, J.; Simic, M.; Stamenkovic-Pejkovic, D.; Starcevic, V.; Trajkovic, V.; Micic, D. Increased activity of interleukin-23/interleukin-17 proinflammatory axis in obese women. Int. J. Obes. 2008, 33, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, L.A.; Shen, W.J.; Joyce-Shaikh, B.; Pyatnova, E.A.; Richards, A.G.; Thom, C.; Andrade, S.M.; Cua, D.J.; Kraemer, F.B.; Butcher, E.C. IL-17 regulates adipogenesis, glucose homeostasis, and obesity. J. Immunol. 2010, 185, 6947–6959. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Zhang, Q.; Ma, S.; Liu, S.; Chen, Z.; Mo, Z.; You, Z. Interleukin-17A differentially induces inflammatory and metabolic gene expression in the adipose tissues of lean and obese mice. Int. J. Mol. Sci. 2016, 17, 522. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan-Bogdan, M.; McDonnell, M.E.; Shin, H.; Rehman, Q.; Hasturk, H.; Apovian, C.M.; Nikolajczyk, B.S. Elevated proinflammatory cytokine production by a skewed T cell compartment requires monocytes and promotes inflammation in type 2 diabetes. J. Immunol. 2011, 186, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Patterson, S.J.; Speck, M.; Ehses, J.A.; Levings, M.K. Insulin inhibits IL-10-mediated regulatory T cell function: Implications for obesity. J. Immunol. 2014, 192, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Wu, D.; Denroche, H.C.; Yao, Y.; Verchere, C.B.; Levings, M.K. IL-33 reverses an obesity-induced deficit in visceral adipose tissue ST2 +T regulatory cells and ameliorates adipose tissue inflammation and insulin resistance. J. Immunol. 2015, 194, 4777–4783. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Huber, J.; Prager, G.; Stulnig, T.M. Inflammation correlates with markers of T-cell subsets including regulatory T cells in adipose tissue from obese patients. Obesity 2011, 19, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Eller, K.; Kirsch, A.; Wolf, A.M.; Sopper, S.; Tagwerker, A.; Stanzl, U.; Wolf, D.; Patsch, W.; Rosenkranz, A.R.; Eller, P. Potential role of regulatory T cells in reversing obesity-linked insulin resistance and diabetic nephropathy. Diabetes 2011, 60, 2954–2962. [Google Scholar] [CrossRef] [PubMed]
- Ilan, Y.; Maron, R.; Tukpah, A.M.; Maioli, T.U.; Murugaiyan, G.; Yang, K.; Wu, H.Y.; Weiner, H.L. Induction of regulatory T cells decreases adipose inflammation and alleviates insulin resistance in ob/ob mice. Proc. Natl. Acad. Sci. USA 2010, 107, 9765–9770. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, M.; Enns, J.; Blewett, H.; Yakandawala, U.; Zahradka, P.; Taylor, C.G. Dietary flaxseed oil reduces adipocyte size, adipose monocyte chemoattractant protein-1 levels and T-cell infiltration in obese, insulin-resistant rats. Cytokine 2012, 59, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Burkhardt, J.K. T-cell-receptor-dependent actin regulatory mechanisms. J. Cell Sci. 2007, 120, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Meiri, K.F. Lipid rafts and regulation of the cytoskeleton during T cell activation. Philos. Trans. R. Soc. Lond. B 2005, 360, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Tybulewicz, V.L.; Henderson, R.B. Rho family GTPases and their regulators in lymphocytes. Nat. Rev. Immunol. 2009, 9, 630–644. [Google Scholar] [CrossRef] [PubMed]
- Jury, E.C.; Flores-Borja, F.; Kabouridis, P.S. Lipid rafts in T cell signalling and disease. 2007, 18, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Lillemeier, B.F.; Mortelmaier, M.A.; Forstner, M.B.; Huppa, J.B.; Groves, J.T.; Davis, M.M. TCR and Lat are expressed on separate protein islands on T cell membranes and concatenate during activation. Nat. Immunol. 2010, 11, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.; Brennan, T.; Li, Q.; McCormack, C.; Seed, B. Membrane compartmentation is required for efficient T cell activation. Immunity 1998, 8, 723–732. [Google Scholar] [CrossRef]
- Rentero, C.; Zech, T.; Quinn, C.M.; Engelhardt, K.; Williamson, D.; Grewal, T.; Jessup, W.; Harder, T.; Gaus, K. Functional implications of plasma membrane condensation for T cell activation. PLoS ONE 2008, 3, e2262. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; McMurray, D.N.; Ly, L.H.; Chapkin, R.S. Dietary (n-3) polyunsaturated fatty acids remodel mouse T-cell lipid rafts. J. Nutr. 2003, 133, 1913–1920. [Google Scholar] [PubMed]
- Kim, W.; Barhoumi, R.; McMurray, D.N.; Chapkin, R.S. Dietary fish oil and DHA down-regulate antigen-activated CD4+ T-cells while promoting the formation of liquid-ordered mesodomains. Br. J. Nutr. 2014, 111, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Fan, Y.Y.; Barhoumi, R.; Smith, R.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress the localization and activation of signaling proteins at the immunological synapse in murine CD4+ T cells by affecting lipid raft formation. J. Immunol. 2008, 181, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Zech, T.; Ejsing, C.S.; Gaus, K.; de Wet, B.; Shevchenko, A.; Simons, K.; Harder, T. Accumulation of raft lipids in T-cell plasma membrane domains engaged in TCR signalling. EMBO J. 2009, 28, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Stulnig, T.M.; Berger, M.; Sigmund, T.; Raederstorff, D.; Stockinger, H.; Waldhausl, W. Polyunsaturated fatty acids inhibit T cell signal transduction by modification of detergent-insoluble membrane domains. J. Cell Biol. 1998, 143, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Stulnig, T.M.; Huber, J.; Leitinger, N.; Imre, E.M.; Angelisová, P.; Nowotny, P.; Waldhausl, W. Polyunsaturated eicosapentaenoic acid displaces proteins from membrane rafts by altering raft lipid composition. J. Biol. Chem. 2001, 276, 37335–37340. [Google Scholar] [CrossRef] [PubMed]
- Saderson, P.; Calder, P.C. Dietary fish oil appears to prevent the activation of phospholipase C-gamma in lymphocytes. Biochim. Biophys. Acta 1998, 1392, 300–308. [Google Scholar] [CrossRef]
- Arrington, J.L.; McMurray, D.N.; Switzer, K.C.; Fan, Y.Y.; Chapkin, R.S. Docosahexaenoic acid suppresses function of the CD28 costimulatory membrane receptor in primary murine and Jurkat T cells. J. Nutr. 2001, 131, 1147–1153. [Google Scholar] [PubMed]
- Chapkin, R.S.; Arrington, J.L.; Apanasovich, T.V.; Carroll, R.J.; McMurray, D.N. Dietary n-3 PUFA affect TcR-mediated activation of purified murine T cells and accessory cell function in co-cultures. Clin. Exp. Immunol. 2002, 130, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Ly, L.H.; Smith, R.; Switzer, K.C.; Chapkin, R.S.; McMurray, D.N. Dietary eicosapentaenoic acid modulates CTLA-4 expression in murine CD4+ T-cells. Prostaglandins Leukot. Essent. Fatty Acids 2006, 74, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Kim, W.; Callaway, E.; Smith, R.; Jia, Q.; Zhou, L.; McMurray, D.N.; Chapkin, R.S. Fat-1 transgene expression prevents cell culture-induced loss of membrane n-3 fatty acids in activated CD4+ T-cells. Prostaglandins Leukot. Essent. Fatty Acids 2008, 79, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Y.; Barhoumi, R.; Fan, Y.Y.; Rivera, G.M.; Hannoush, R.N.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress CD4+ T cell proliferation by altering phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P2] organization. Biochim. Biophys. Acta 2016, 1858, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Yog, R.; Barhoumi, R.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress mitochondrial translocation to the immunologic synapse and modulate calcium signaling in T cells. J. Immunol. 2010, 184, 5865–5873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kim, W.; Zhou, L.; Wang, N.; Ly, L.H.; McMurray, D.N.; Chapkin, R.S. Dietary fish oil inhibits antigen-specific murine Th1 cell development by suppression of clonal expansion. J. Nutr. 2006, 136, 2391–2398. [Google Scholar] [PubMed]
- Zhang, P.; Smith, R.; Chapkin, R.S.; McMurray, D.N. Dietary (n-3) polyunsaturated fatty acids modulate murine Th1/Th2 balance toward the Th2 pole by suppression of Th1 development. J. Nutr. 2005, 135, 1745–1751. [Google Scholar] [PubMed]
- Petursdottir, D.H.; Hardardottir, I. Dietary fish oil decreases secretion of T helper (Th) 1-type cytokines by a direct effect on murine splenic T cells but enhances secretion of a Th2-type cytokine by an effect on accessory cells. Br. J. Nutr. 2008, 101, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Hou, T.Y.; Turk, H.F.; McMurray, D.N.; Chapkin, R.S. n3 PUFAs reduce mouse CD4+ T-cell ex vivo polarization into Th17 cells. J. Nutr. 2013, 143, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Fan, Y.Y.; Monk, J.M.; Hou, T.Y.; Barhoumi, R.; McMurray, D.N.; Chapkin, R.S. n-3 PUFAs reduce T-helper 17 cell differentiation by decreasing responsiveness to interleukin-6 in isolated mouse splenic CD4+ T cells. J. Nutr. 2014, 144, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Bevan, M.J. CD8+ T Cells: Foot soldiers of the immune system. Immunity 2011, 35, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zhang, Y.; Yang, L.; Liu, G.; Ye, J.; Wang, H. Effects of a high-fat diet on adipose tissue CD8+ T cells in young vs. adult mice. Inflammation 2017, 40, 1944–1958. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Hemann, E.A.; Legge, K.L.; Norian, L.A.; Badovinac, V.P. Diet-induced obesity does not impact the generation and maintenance of primary memory CD8 T cells. J. Immunol. 2014, 193, 5873–5882. [Google Scholar] [CrossRef] [PubMed]
- Laugerette, F.; Furet, J.P.; Debard, C.; Daira, P.; Loizon, E.; Geloen, A.; Soulage, C.O.; Simonet, C.; Lefils-Lacourtablaise, J.; Bernoud-Hubac, N.; et al. Oil composition of high-fat diet affects metabolic inflammation differently in connection with endotoxin receptors in mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E374–E386. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.M.; Liddle, D.M.; Brown, M.J.; Zarepoor, L.; De Boer, A.A.; Ma, D.W.; Power, K.A.; Robinson, L.E. Anti-inflammatory and anti-chemotactic effects of dietary flaxseed oil on CD8 +T cell/adipocyte-mediated cross-talk. Mol. Nutr. Food Res. 2015, 60, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Bruun, J.M.; Lihn, A.S.; Pedersen, S.B.; Richelsen, B. Monocyte chemoattractant protein-1 release is higher in visceral than subcutaneous human adipose tissue (AT): Implication of macrophages resident in the AT. J. Clin. Endocrinol. Metab. 2005, 90, 2282–2289. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.I.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Soga, Y.; Iwamoto, Y.; Asano, T.; Li, Y.; Abiko, Y.; Nishimura, F. DNA microarray analyses of genes expressed differentially in 3T3-L1 adipocytes co-cultured with murine macrophage cell line RAW264.7 in the presence of the toll-like receptor 4 ligand bacterial endotoxin. Int. J. Obes. 2008, 32, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.T.; Xia, D.S.; Yang, W.K.; Wang, X.G.; Chen, X.Z.; Dong, N.G. Secretion of adipocytes and macrophages under conditions of inflammation and/or insulin resistance and effect of adipocytes on preadipocytes under these conditions. Biochemistry 2014, 79, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Sartipy, P.; Loskutoff, D. Monocyte chemoattractant protein 1 in obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Hunter, D.; Huber, R.; Lemieux, J.; Slaymaker, S.; Vaddi, K.; Charo, I.; Leibel, R.L.; Ferrante, A.W. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J. Clin. Investig. 2006, 116, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Fried, S.K.; Bunkin, D.A.; Greenberg, A.S. Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: Depot difference and regulation by glucocorticoid. J. Clin. Endocrinol. Metab. 1998, 83, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Bastard, J.P.; Maachi, M.; Van Nhieu, J.T.; Jardel, C.; Bruckert, E.; Grimaldi, A.; Robert, J.J.; Capeau, J.; Hainque, B. Adipose tissue IL-6 content correlates with resistance to insulin activation of glucose uptake both in vivo and in vitro. J. Clin. Endocrinol. Metab. 2002, 87, 2084–2089. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Mohlig, M.; Hoffman, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Mohamed-Ali, V.; Goodrick, S.; Rawesh, A.; Katz, D.R.; Miles, J.M.; Yudkin, J.S.; Klein, S.; Coppack, S.W. Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J. Clin. Endocrinol. Metab. 1997, 82, 4196–4200. [Google Scholar] [CrossRef] [PubMed]
- Sopasakis, V.R.; Sandqvist, M.; Gustafson, B.; Hammarstedt, A.; Schmelz, M.; Yang, X.; Jansson, P.A.; Smith, U. High local concentrations and effects on differentiation implicate interleukin-6 as a paracrine regulator. Obesity 2004, 12, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Harkins, J.M.; Moustaid-Moussa, N.; Chung, Y.J.; Penner, K.M.; Pestka, J.J.; North, C.M.; Claycombe, K.J. Expression of interleukin-6 is greater in preadipocytes than in adipocytes of 3T3-L1 cells and C57BL/6J and ob/ob mice. J. Nutr. 2004, 134, 2673–2677. [Google Scholar] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Muller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Bastard, J.P.; Auclair, M.; Maachi, M.; Capeau, J.; Caron, M. Chronic interleukin-6 (IL-6) treatment increased IL-6 secretion and induced insulin resistance in adipocyte: Prevention by rosiglitazone. Biochem. Biophys. Res. Commun. 2003, 311, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Tzameli, I.; Bjorbaek, C.; Flier, J.S. Suppressor of cytokine signaling 3 is a physiological regulator of adipocyte insulin signaling. J. Biol. Chem. 2004, 279, 34733–34740. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Kralisch, S.; Klier, M.; Lossner, U.; Bluher, M.; Klein, J.; Paschke, R. Adiponectin gene expression and secretion is inhibited by interleukin-6 in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2003, 301, 1045–1050. [Google Scholar] [CrossRef]
- Sopasakis, V.R.; Nagaev, I.; Smith, U. Interleukin-6 (IL-6) induces insulin resistance in 3T3-L1 adipocytes and is, like IL-8 and tumor necrosis factor-alpha, overexpressed in human fat cells from insulin-resistant subjects. J. Biol. Chem. 2003, 278, 45777–45784. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Maury, E.; Noël, L.; Detry, R.; Brichard, S.M. In vitro hyperresponsiveness to tumor necrosis factor-alpha contributes to adipokine dysregulation in omental adipocytes of obese subjects. J. Clin. Endocrinol. Metab. 2009, 94, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Weinstock, R.; Thusu, K.; Abdel-Rahman, E.; Aljada, A.; Wadden, T. Tumor necrosis factor-alpha in sera of obese patients: Fall with weight loss. J. Clin. Endocrinol. Metab. 1998, 83, 2907–2910. [Google Scholar] [CrossRef] [PubMed]
- Hivert, M.F.; Sullivan, L.M.; Fox, C.S.; Nathan, D.M.; D’Agostino, R.B.; Wilson, P.W.; Meigs, J.B. Associations of adiponectin, resistin, and tumor necrosis factor-alpha with insulin resistance. J. Clin. Endocrinol. Metab. 2008, 93, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Uysal, K.T.; Becherer, J.D.; Arner, P.; Hotamisligil, G.S. Altered tumor necrosis factor-alpha (TNF-alpha) processing in adipocytes and increased expression of transmembrane TNF-alpha in obesity. Diabetes 2002, 51, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
- Plomgaard, P.; Bouzakri, K.; Krogh-Madsen, R.; Mittendorfer, B.; Zierath, J.R.; Pedersen, B.K. Tumor necrosis factor-alpha induces skeletal muscle insulin resistance in healthy human subjects via inhibition of Akt substrate 160 phosphorylation. Diabetes 2005, 54, 2939–2945. [Google Scholar] [CrossRef] [PubMed]
- Uysal, T.K.; Wiesbrock, S.M.; Marino, M.W.; Hotamisligil, G.S. Protection from obesity-induced insulin resistancein mice lacking TNF-alpha function. Nature 1997, 389, 610–614. [Google Scholar] [CrossRef] [PubMed]
- MacEwan, D.J. TNF receptor subtype signalling: Differences and cellular consequences. Cell Signal. 2002, 14, 477–492. [Google Scholar] [CrossRef]
- Uysal, T.K.; Wiesbrock, S.M.; Hotamisligil, G.S. Functional analysis of tumor necrosis factor (TNF) receptors in TNF-alpha-mediated insulin resistance in genetic obesity. Endocrinology 1998, 139, 4832–4838. [Google Scholar] [CrossRef] [PubMed]
- Sethi, J.K.; Xu, H.; Uysal, K.T.; Wiesbrock, S.M.; Scheja, L.; Hotamisligil, G.S. Characterisation of receptor-specific TNFalpha functions in adipocyte cell lines lacking type 1 and 2 TNF receptors. FEBS Lett. 2000, 469, 77–82. [Google Scholar] [CrossRef]
- Good, M.; Newell, F.M.; Haupt, L.M.; Whitehead, J.P.; Hutley, L.J.; Prins, J.B. TNF and TNF receptor expression and insulin sensitivity in human omental and subcutaneous adipose tissue—Influence of BMI and adipose distribution. Diabetes Vasc. Dis. Res. 2006, 3, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Satoh, H.; Favelyukis, S.; Babendure, J.L.; Imamura, T.; Sbodio, J.I.; Zalevsky, J.; Dahiyat, B.I.; Chi, N.W.; Olefsky, J.M. JNK and tumor necrosis factor-alpha mediate free fatty acid-induced insulin resistance in 3T3-L1 adipocytes. J. Biol. Chem. 2005, 280, 35361–35371. [Google Scholar] [CrossRef] [PubMed]
- McGillicuddy, F.C.; Harford, K.A.; Reynolds, C.M.; Oliver, E.; Claessens, M.; Mills, K.H.; Roche, H.M. Lack of interleukin-1 receptor I (IL-1RI) protects mice from high-fat diet-induced adipose tissue inflammation coincident with improved glucose homeostasis. Diabetes 2011, 60, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Koenen, T.B.; Stienstra, R.; van Tits, L.J.; Joosten, L.A.; van Velzen, J.F.; Hijmans, A.; Pol, J.A.; Netea, M.G.; Tack, C.J.; Stalenhoef, A.F.; et al. The inflammasome and caspase-1 activation: A new mechanism underlying increased inflammatory activity in human visceral adipose tissue. Endocrinology 2011, 152, 3769–3778. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J.; Marois, J.; Garoufalis, J.; Daddario, M.; Roulston, A.; Kwan, I.; Pepin, N.; Lacoste, J.; Nguyen, H.; Bensi, G. Characterization of a functional NF-kappaB site in the human interleukin 1 beta promoter: Evidence for a positive autoregulatory loop. Mol. Cell. Biol. 1993, 13, 6231–6240. [Google Scholar] [CrossRef] [PubMed]
- Nov, O.; Kohl, A.; Lewis, E.C.; Bashan, N.; Dvir, I.; Ben-Shlomo, S.; Fishman, S.; Wueest, S.; Konrad, D.; Rudich, A. Interleukin-1beta may mediate insulin resistance in liver-derived cells in response to adipocyte inflammation. Endocrinology 2010, 151, 4247–4256. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; Tack, C.J.; Kanneganti, T.D.; Joosten, L.A.; Netea, M.G. The inflammasome puts obesity in the danger zone. Cell Metab. 2012, 15, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Yvan-Charvet, L.; Bastard, J.P.; Maachi, M.; Quignard-Boulangé, A.; Capeau, J.; Caron, M. Long-term treatment with interleukin-1beta induces insulin resistance in murine and human adipocytes. Diabetologia 2006, 49, 2162–2173. [Google Scholar] [CrossRef] [PubMed]
- Jager, J.; Grémeaux, T.; Cormont, M.; Le Marchand-Brustel, Y.; Tanti, J.F. Interleukin-1beta-induced insulin resistance in adipocytes through down-regulation of insulin receptor substrate-1 expression. Endocrinology 2007, 148, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, H.W.; Zhou, Z.; Whelan, J.; Allen, K.G.; Moustaid-Moussa, N.; Kim, H.; Claycombe, K.J. Stearidonic and eicosapentaenoic acids inhibit interleukin-6 expression in ob/ob mouse adipose stem cells via toll-like receptor-2-mediated pathways. J. Nutr. 2011, 141, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Manickam, E.; Sinclair, A.J.; Cameron-Smith, D. Suppressive actions of eicosapentaenoic acid on lipid droplet formation in 3T3-L1 adipocytes. Lipids Health Dis. 2010, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Echarri, N.; Pérez-Matute, P.; Marcos-Gómez, B.; Baena, M.J.; Marti, A.; Martínez, J.A.; Moreno-Aliaga, M.J. Differential inflammatory status in rats susceptible or resistant to diet-induced obesity: Effects of EPA ethyl ester treatment. Eur. J. Nutr. 2008, 47, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M.A.; Cohen, D.J.; Liddle, D.M.; Robinson, L.E.; Ma, D.W. A review of the effect of omega-3 polyunsaturated fatty acids on blood triacylglycerol levels in normolipidemic and borderline hyperlipidemic individuals. Lipids Health Dis. 2015, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- Ramel, A.; Martinez, J.A.; Kiely, M.; Bandarra, N.M.; Thorsdottir, I. Effects of weight loss and seafood consumption on inflammation parameters in young, overweight and obese European men and women during 8 weeks of energy restriction. Eur. J. Clin. Nutr. 2010, 64, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Browning, L.M.; Krebs, J.D.; Moore, C.S.; Mishra, G.D.; O’Connell, M.A.; Jebb, S.A. The impact of long chain n-3 polyunsaturated fatty acid supplementation on inflammation, insulin sensitivity and CVD risk in a group of overweight women with an inflammatory phenotype. Diabetes Obes. Metab. 2007, 9, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Couture, P.; Leclerc, M.; Charest, A.; Marin, J.; Lépine, M.C.; Talbot, D.; Tchernof, A.; Lamarche, B. A randomized, crossover, head-to-head comparison of eicosapentaenoic acid and docosahexaenoic acid supplementation to reduce inflammation markers in men and women: The Comparing EPA to DHA (ComparED) Study. Am. J. Clin. Nutr. 2016, 104, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Woodman, R.J.; Burke, V.; Puddey, I.B.; Croft, K.D.; Beilin, L.J. Effect of eicosapentaenoic acid and docosahexaenoic acid on oxidative stress and inflammatory markers in treated-hypertensive type 2 diabetic subjects. Free Radic. Biol. Med. 2003, 35, 772–781. [Google Scholar] [CrossRef]
- Jellema, A.; Plat, J.; Mensink, R.P. Weight reduction, but not a moderate intake of fish oil, lowers concentrations of inflammatory markers and PAI-1 antigen in obese men during the fasting and postprandial state. Eur. J. Clin. Investig. 2004, 34, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.D.; Browning, L.M.; McLean, N.K.; Rothwell, J.L.; Mishra, G.D.; Moore, C.S.; Jebb, S.A. Additive benefits of long-chain n-3 polyunsaturated fatty acids and weight-loss in the management of cardiovascular disease risk in overweight hyperinsulinaemic women. Int. J. Obes. 2006, 30, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.J.; Meyer, B.J.; Mori, T.A.; Burke, V.; Mansour, J.; Patch, C.S.; Tapsell, L.C.; Noakes, M.; Clifton, P.A.; Barden, A.; et al. Impact of foods enriched with n-3 long-chain polyunsaturated fatty acids on erythrocyte n-3 levels and cardiovascular risk factors. Br. J. Nutr. 2007, 97, 749. [Google Scholar] [CrossRef] [PubMed]
- Skulas-Ray, A.C.; Kris-Etherton, P.M.; Harris, W.S.; Vanden Heuvel, J.P.; Wagner, P.R.; West, S.G. Dose-response effects of omega-3 fatty acids on triglycerides, inflammation, and endothelial function in healthy persons with moderate hypertriglyceridemia. Am. J. Clin. Nutr. 2011, 93, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Dewell, A.; Marvasti, F.F.; Harris, W.S.; Tsao, P.; Gardner, C.D. Low- and high-dose plant and marine (n-3) fatty acids do not affect plasma inflammatory markers in adults with metabolic syndrome. J. Nutr. 2011, 141, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; La Rocca, C.; Matarese, G. Immunological functions of leptin and adiponectin. Biochimie 2012, 94, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.G.; Javorschi, S.; Hevener, A.L.; Kruszynska, Y.T.; Norman, R.A.; Sinha, M.; Olefsky, M. The effect of thiazolidinediones on plasma adiponectin levels in normal, obese, and type 2 diabetic subjects. Diabetes 2002, 51, 2968–2974. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, Y.; Murakami, K.; Tsuboyama-Kasaoka, N.; Ezaki, O.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Guenther, M.; James, R.; Marks, J.; Zhao, S.; Szabo, A.; Kidambi, S. Adiposity distribution influences circulating adiponectin levels. Transl. Res. 2014, 164, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; van de Wall, E.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G.; et al. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Investig. 2007, 117, 2621–2637. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Vasseur, F.; Froguel, P.; Kimura, S.; et al. Impaired multimerization of human adiponectin mutants associated with diabetes. J. Biol. Chem. 2003, 278, 40352–40363. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Horikoshi, M.; Yamauchi, T.; Yago, H.; Miyazaki, O.; Ebinuma, H.; Imai, Y.; Nagai, R.; Kadowaki, T. Measurement of the high-molecular weight form of adiponectin in plasma is useful for the prediction of insulin resistance and metabolic syndrome. Diabetes Care 2006, 29, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Shao, J. Adiponectin and energy homeostasis. Rev. Endocr. Metab. Disord. 2014, 15, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Tishinsky, J.M.; Robinson, L.E.; Dyck, D.J. Insulin-sensitizing properties of adiponectin. Biochimie 2012, 94, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y.; et al. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 2002, 8, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Hector, J.; Schwarzloh, B.; Goehring, J.; Strate, T.; Hess, U.; Deuretzbacher, G.; Hansen-Algenstaedt, N.; Beil, F.U.; Algenstaedt, P. TNF-alpha alters visfatin and adiponectin levels in human fat. Horm. Metab. Res. 2007, 39, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Kim, J.K.; Jeon, J.H.; Yoon, S.R.; Choi, I.; Yang, Y. c-Jun N-terminal kinase is involved in the suppression of adiponectin expression by TNF-alpha in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2005, 327, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Park, P.H.; Huang, H.; McMullen, M.R.; Mandal, P.; Sun, L.; Nagy, L.E. Suppression of lipopolysaccharide-stimulated tumor necrosis factor-alpha production by adiponectin is mediated by transcriptional and post-transcriptional mechanisms. J. Biol. Chem. 2008, 283, 26850–26858. [Google Scholar] [CrossRef] [PubMed]
- Lira, F.S.; Rosa, J.C.; Pimentel, G.D.; Seelaender, M.; Damaso, A.R.; Oyama, L.M.; do Nascimento, C.O. Both adiponectin and interleukin-10 inhibit LPS-induced activation of the NF-κB pathway in 3T3-L1 adipocytes. Cytokine 2012, 57, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M.; Spurlock, M.E. Adiponectin inhibits LPS-induced NF-kappaB activation and IL-6 production and increases PPARgamma2 expression in adipocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1220–R1225. [Google Scholar] [CrossRef] [PubMed]
- Folco, E.J.; Rocha, V.Z.; López-Ilasaca, M.; Libby, P. Adiponectin inhibits pro-inflammatory signaling in human macrophages independent of interleukin-10. J. Biol. Chem. 2009, 284, 25569–25575. [Google Scholar] [CrossRef] [PubMed]
- Lovren, F.; Pan, Y.; Quan, A.; Szmitko, P.E.; Singh, K.K.; Shukla, P.C.; Gupta, M.; Chan, L.; Al-Omran, M.; Teoh, H.; et al. Adiponectin primes human monocytes into alternative anti-inflammatory M2 macrophages. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H656–H663. [Google Scholar] [CrossRef] [PubMed]
- Wulster-Radcliffe, M.C.; Ajuwon, K.M.; Wang, J.; Christian, J.A.; Spurlock, M.E. Adiponectin differentially regulates cytokines in porcine macrophages. Biochem. Biophys. Res. Commun. 2004, 316, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Argueta, J.G.; Masuhiro, Y.; Kagishita, M.; Nonaka, K.; Saito, T.; Hanazawa, S.; Yamashita, Y. Adiponectin inhibits toll-like receptor family-induced signaling. FEBS Lett. 2005, 579, 6821–6826. [Google Scholar] [CrossRef] [PubMed]
- Kollias, A.; Tsiotra, P.C.; Ikonomidis, I.; Maratou, E.; Mitrou, P.; Boutati, E.; Lekakis, J.; Economopoulos, T.; Kremastinos, D.T.; Dimitriadis, G.; et al. Adiponectin levels and expression of adiponectinreceptors in isolated monocytes from overweightpatients with coronary artery disease. Cardiovasc. Diabetol. 2011, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumada, M.; Kihara, S.; Ouchi, N.; Kobayashi, H.; Okamoto, Y.; Ohashi, K.; Maeda, K.; Nagaretani, H.; Kishida, K.; Maeda, N.; et al. Adiponectin specifically increased tissue inhibitor of metalloproteinase-1 through interleukin-10 expression in human macrophages. Circulation 2004, 109, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Pratt, B.T.; Barnes, M.; McMullen, M.R.; Nagy, L.E. Molecular mechanism for adiponectin-dependent M2 macrophage polarization. J. Biol. Chem. 2011, 286, 13460–13469. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Wolf, D.; Rumpold, H.; Enrich, B.; Tilg, H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL-1RA in human leukocytes. Biochem. Biophys. Res. Commun. 2004, 323, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Roychowdhury, S.; Park, P.H.; Pratt, B.T.; Roger, T.; Nagy, L.E. Adiponectin and heme oxygenase-1 suppress TLR4/MyD88-independent signaling in rat kupffer cells and in mice after chronic ethanol exposure. J. Immunol. 2010, 185, 4928–4937. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.; Li, D.; Ho, D.; Peng, J.; Xu, A.; Lamb, J.; Chen, Y.; Tam, P.K. Novel immunomodulatory effecs of adiponectin on dendritic cell functions. Int. Immunopharmacol. 2011, 11, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.S.; Paglia, D.; Kwan, A.; Deitel, M. Increased obese mRNA expression in omental fat cells from massively obese humans. Nat. Med. 1995, 1, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Considine, R.V.; Sinha, M.K.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Nyce, M.R.; Ohannesian, J.P.; Marco, C.C.; McKee, L.J.; Bauer, T.L.; et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N. Engl. J. Med. 1996, 334, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Frederich, R.C.; Hamann, A.; Anderson, S.; Lollmann, B.; Lowell, B.B.; Flier, J.S. Leptin levels reflect body lipid content in mice: Evidence from diet-induced resistance to leptin action. Nat. Med. 1995, 1, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Santos-Alvarez, J.; Goberna, R.; Sánchez-Margalet, V. Human leptin stimulates proliferation and activation of human circulating monocytes. Cell. Immunol. 1999, 194, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Sakaida, I.; Uchida, K.; Terai, S.; Okita, K. Leptin enhances TNF-alpha production via p38 and JNK MAPK in LPS-stimulated Kupffer cells. Life Sci. 2005, 77, 1502–1515. [Google Scholar] [CrossRef] [PubMed]
- Landman, R.E.; Puder, J.J.; Xiao, E.; Freda, P.U.; Ferin, M.; Wardlaw, S.L. Endotoxin stimulates leptin in the human and nonhuman primate. J. Clin. Endocrinol. Metab. 2003, 88, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Faggioni, R.; Fantuzzi, G.; Fuller, J.; Dinarello, C.A.; Feingold, K.R.; Grunfeld, C. IL-1 beta mediates leptin induction during inflammation. Am. J. Physiol. 1998, 274, R204–R208. [Google Scholar] [PubMed]
- Surmi, B.K.; Hasty, A.H. Macrophage infiltration into adipose tissue: Initiation, propagation and remodeling. Future Lipidol. 2008, 3, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Kiguchi, N.; Maeda, T.; Kobayashi, Y.; Fukazawa, Y.; Kishioka, S. Leptin enhances CC-chemokine ligand expression in cultured murine macrophage. Biochem. Biophys. Res. Commun. 2009, 384, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Zarkesh-Esfahani, H.; Pockley, G.; Metcalfe, R.A.; Bidlingmaier, M.; Wu, Z.; Ajami, A.; Weetman, A.P.; Strasburger, C.J.; Ross, R.J. High-dose leptin activates human leukocytes via receptor expression on monocytes. J. Immunol. 2001, 167, 4593–4599. [Google Scholar] [CrossRef] [PubMed]
- Loffreda, S.; Yang, S.Q.; Lin, H.Z.; Karp, C.L.; Brengman, M.L.; Wang, D.J.; Klein, A.S.; Bulkley, G.B.; Bao, C.; Noble, P.W.; et al. Leptin regulates proinflammatory immune responses. FASEB J. 1998, 12, 57–65. [Google Scholar] [PubMed]
- Berg, A.H.; Scherer, P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005, 96, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jenkins, J.R.; Trayhurn, P. Expression and secretion of inflammation-related adipokines by human adipocytes differentiated in culture: Integrated response to TNF-alpha. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E731–E740. [Google Scholar] [CrossRef] [PubMed]
- Von Frankenberg, A.D.; Silva, F.M.; de Almeida, J.C.; Piccoli, V.; Nascimento, F.V.; Sost, M.M.; Leitão, C.B.; Remonti, L.L.; Umpierre, D.; Reis, A.F.; et al. Effect of dietary lipids on circulating adiponectin: A systematic review with meta-analysis of randomised controlled trials. Br. J. Nutr. 2014, 112, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Cahill, L.E.; Mozaffarian, D. Effect of fish oil on circulating adiponectin: A systematic review and meta-analysis of randomized controlled trials. J. Clin. Endocrinol. Metab. 2013, 98, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Itoh, M.; Suganami, T.; Satoh, N.; Tanimoto-Koyama, K.; Yuan, X.; Tanaka, M.; Kawano, H.; Yano, T.; Aoe, S.; Takeya, M.; et al. Increased adiponectin secretion by highly purified eicosapentaenoic acid in rodent models of obesity and human obese subjects. Aterioscler. Thromb. Vasc. Biol. 2007, 27, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Gammelmark, A.; Madsen, T.; Varming, K.; Lundbye-Christensen, S.; Schmidt, E.B. Low-dose fish oil supplementation increases serum adiponectin without affecting inflammatory markers in overweight subjects. Nutr. Res. 2012, 32, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Mohamed-Ali, V.; Horakova, O.; Rossmeisl, M.; Hosseinzadeh-Attar, M.J.; Hensler, M.; Ruzickova, J.; Kopecky, J. Polyunsaturated fatty acids of marine origin induce adiponectin in mice fed a high-fat diet. Diabetologia 2006, 49, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Tishinsky, J.M.; Gulli, R.A.; Mullen, K.L.; Dyck, D.J.; Robinson, L.E. Fish oil prevents high-saturated fat diet-induced impairments in adiponectin and insulin response in rodent soleus muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R598–R605. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Itoh, T.; Abe, D.; Shimizu, M.; Kanda, T.; Koyama, T.; Nishikawa, M.; Tamai, T.; Ooizumi, H.; Yamada, S. Identification of putative metabolites of docosahexaenoic acid as potent PPARgamma agonists and antidiabetic agents. Bioorg. Med. Chem. Lett. 2005, 15, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, M.; Suraamornkul, S.; Piper, P.; Hardies, L.J.; Glass, L.; Cersosimo, E.; Pratipanawatr, T.; Miyazaki, Y.; DeFronzo, R.A. Decreased plasma adiponectin concentrations are closely related to hepatic fat content and hepatic insulin resistance in pioglitazone-treated type 2 diabetic patients. J. Clin. Endocrinol. Metab. 2004, 89, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Neschen, S.; Morino, K.; Rossbacher, J.C.; Pongratz, R.L.; Cline, G.W.; Sono, S.; Gillum, M.; Shulman, G.I. Fish oil regulates adiponectin secretion by a peroxisome proliferator-activated receptor-gamma–dependent mechanism in mice. Diabetes 2006, 55, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Oster, R.T.; Tishinsky, J.M.; Yuan, Z.; Robinson, L.E. Docosahexaenoic acid increases cellular adiponectin mRNA and secreted adiponectin protein, as well as PPARγ mRNA, in 3T3-L1 adipocytes. Appl. Physiol. Nutr. Metab. 2010, 35, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Tishinsky, J.M.; Ma, D.W.; Robinson, L.E. Eicosapentaenoic acid and rosiglitazone increase adiponectin in an additive and PPARγ-dependent manner in human adipocytes. Obesity 2010, 19, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K. Nuclear receptor transrepressionpathways that regulate inflammation in macrophages and T cells. Nat. Rev. Immunol. 2010, 10, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Red Eagle, A.; Vats, D.; Brombacher, F.; Ferrante, A.W.; et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Winnicki, M.; Somers, V.K.; Accurso, V.; Phillips, B.G.; Puato, M.; Palatini, P.; Pauletto, P. Fish-rich diet, leptin, and body mass. Circulation 2002, 106, 289–291. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Shawb, J.E.; Beilin, L.J. Effect of fish diets and weight loss on serum leptin concentration in overweight, treated-hypertensive subjects. J. Hypertens. 2004, 22, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Ide, T. Dietary n-3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br. J. Nutr. 2000, 84, 175–184. [Google Scholar] [PubMed]
- Ruzickova, J.; Rossmeisl, M.; Prazak, T.; Flachs, P.; Sponarova, J.; Veck, M.; Tvrzicka, E.; Bryhn, M.; Kopecky, J. Omega-3 PUFA of marine origin limit diet-induced obesity in mice by reducing cellularity of adipose tissue. Lipids 2004, 39, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Pinel, A.; Pitois, E.; Rigaudiere, J.P.; Jouve, C.; De Saint-Vincent, S.; Laillet, B.; Montauier, C.; Huertas, A.; Morio, B.; Chapel, F. EPA prevents fat mass expansion and metabolic disturbances in mice fed with a Western diet. J. Lipid Res. 2016, 57, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, L.; Bekaert, T.; Chau, T.L.; Tavernier, J.; Chariot, A.; Beyaert, R. TLR-4, IL-1R and TNF-R signaling to NF-kappaB: Variations on a common theme. Cell. Mol. Life Sci. 2008, 65, 2964–2978. [Google Scholar] [CrossRef] [PubMed]
- Vitseva, O.I.; Tanriverdi, K.; Tchkonia, T.T.; Kirkland, J.L.; McDonnell, M.E.; Apovian, C.M.; Freedman, J.; Gokce, N. Inducible toll-like receptor and NF-kappaB regulatory pathway expression in human adipose tissue. Obesity 2008, 16, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Fusaru, A.M.; Stănciulescu, C.E.; Surlin, V.; Taisescu, C.; Bold, A.; Pop, O.T.; Banita, I.M.; Craitoiu, S.; Pisoschi, C.G. Role of innate immune receptors TLR2 and TLR4 as mediators of the inflammatory reaction in human visceral adipose tissue. Rom. J. Morphol. Embryol. 2012, 53, 693–701. [Google Scholar] [PubMed]
- Harte, A.L.; Tripathi, G.; Piya, M.K.; Barber, T.M.; Clapham, J.C.; Al-Daghri, N.; Al-Disi, D.; Kumsaiyai, W.; Saravanan, P.; Fowler, A.E.; et al. NFκB as a potent regulator of inflammation in human adipose tissue, influenced by depot, adiposity, T2DM status, and TNFα. Obesity 2013, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Bujo, H.; Unoki, H.; Saito, Y. High fat intake induces a population of adipocytes to co-express TLR2 and TNFalpha in mice with insulin resistance. Biochem. Biophys. Res. Commun. 2007, 354, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Akashi, S.; Miyake, K.; Petty, H.R. Lipopolysaccharide induces physical proximity between CD14 and toll-like receptor 4 (TLR4) prior to nuclear translocation of NF-kappa B. J. Immunol. 2000, 165, 3541–3544. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Triantafilou, K. Lipopolysaccharide recognition: CD14, TLRs and the LPS-activation cluster. Trends Immunol. 2002, 23, 301–304. [Google Scholar] [CrossRef]
- Lee, J.Y.; Zhao, L.; Youn, H.S.; Weatherill, A.R.; Tapping, R.; Feng, L.; Lee, W.H.; Fitzgerald, K.A.; Hwang, D.H. Saturated fatty acid activates but polyunsaturated fatty acid inhibits toll-like receptor 2 dimerized with toll-like receptor 6 or 1. J. Biol. Chem. 2004, 279, 16971–16979. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature 2012, 483, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Denyer, G. Mining microarray datasets in nutrition: Expression of the GPR120 (n-3 fatty acid receptor/sensor) gene is down-regulated in human adipocytes by macrophage secretions. J. Nutr. Sci. 2012, 1, e3. [Google Scholar] [CrossRef] [PubMed]
- Muredda, L.; Kepczynska, M.A.; Zaibi, M.S.; Alomar, S.Y.; Trayhurn, P. IL-1β inhibit GPR120 (FFAR4) and stimulate GPR84 (EX33) and GPR41 (FFAR3) fatty acid receptor expression in human adipocytes: Implications for the anti-inflammatory action of n-3 fatty acids. Arch. Physiol. Biochem. 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bae, I.S.; Park, P.J.; Lee, J.H.; Cho, E.G.; Lee, T.R.; Kim, S.H. PPARγ-mediated G-protein coupled receptor 120 signaling pathway promotes transcriptional activation of miR-143 in adipocytes. Gene 2017, 626, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Burns, R.N.; Moniri, N.H. Agonism with the omega-3 fatty acids alpha-linolenic acid and docosahexaenoic acid mediates phosphorylation of both the short and long isoforms of the human GPR120 receptor. Biochem. Biophys. Res. Commun. 2010, 396, 1030–1035. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.U.; Ohmori, K.; Konishi, K.; Igarashi, J.; Hashimoto, T.; Kamitori, K.; Yamaguchi, F.; Tsukamoto, I.; Uyama, T.; Ishihara, Y.; et al. Eicosapentarnoic acid upregulates VEGF-A through both GPR120 and PPARγ mediated pathways in 3T3-L1 adipocytes. Mol. Cell. Endocrinol. 2015, 406, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Mobraten, K.; Haug, T.M.; Kleiveland, C.R.; Lea, T. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health Dis. 2013, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.U.; Ohmori, K.; Hashimoto, T.; Kamitori, K.; Yamaguchi, F.; Noma, T.; Igarashi, J.; Tsuboi, K.; Tokuda, M.; Nishiyama, A.; et al. GPR120 in adipocytes has differential roles in the production of pro-inflammatory adipocytokines. Biochem. Biophys. Res. Commun. 2017, 486, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Pires, K.M.; Ilkun, O.; Valente, M.; Boudina, S. Treatment with a SOD mimetic reduces visceral adiposity, adipocyte death, and adipose tissue inflammation in high fat-fed mice. Obesity 2014, 22, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.; Khemka, V.K.; Chatterjee, G.; Ganguly, A.; Mukhopadhyay, S.; Chakrabarti, S. Enhanced ROS production and oxidative damage in subcutaneous white adipose tissue mitochondria in obese and type 2 diabetes subjects. Mol. Cell. Biol. Chem. 2015, 399, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Blaak, E.E.; Theunissen, R.; Duijvestijn, A.M.; Clement, K.; Tervaert, J.W.; Thewissen, M.M. Expression of NLRP3 inflammasome and T cell population markers in adipose tissue are associated with insulin resistance and impaired glucose metabolism in humans. Mol. Immunol. 2012, 50, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; van Diepen, J.A.; Tack, C.J.; Zaki, H.; van de Veerdonk, F.L.; Perera, D.; Neale, G.A.; Hooiveld, G.J.; Hijmans, A.; Vroegrijk, I.; et al. Inflammasome is a central player in the induction of obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 15324–15329. [Google Scholar] [CrossRef] [PubMed]
- Legrand-Poels, S.; Esser, N.; L’homme, L.; Scheen, A.; Paquot, N.; Piette, J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem. Pharmacol. 2014, 92, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.M.; Hu, W.; Troutman, T.D.; Jennings, M.; Brewer, T.; Li, X.; Nanda, S.; Cohen, P.; Thomas, J.A.; Pasare, C. IRAK-1 bypasses priming and directly links TLRs to rapid NLRP3 inflammasome activation. Proc. Natl. Acad. Sci. USA 2014, 111, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Ralston, J.C.; Lyons, C.L.; Kennedy, E.B.; Kirwan, A.M.; Roche, H.M. Fatty acids and NLRP3 inflammasome–mediated inflammation in metabolic tissues. Annu. Rev. Nutr. 2017, 37, 77–102. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).