Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium

Abstract
:1. Introduction
2. Experimental Section
2.1. Composition of Caprine Milk Oligosaccharide-Enriched Fraction
2.2. Stock Solutions of CMOF for Intestinal Epithelial Cell Based Assays
2.3. Cell Lines
2.4. Cell Culture
2.5. Measurement of Metabolic Activity of Intestinal Epithelial Cell Cultures
2.6. Trans-Epithelial Electrical Resistance Assay
2.7. Quantification of Mucin Gene Expression of Intestinal Epithelial Cell Cultures
2.8. Mucin Protein Abundance
2.9. Statistical Analysis
3. Results
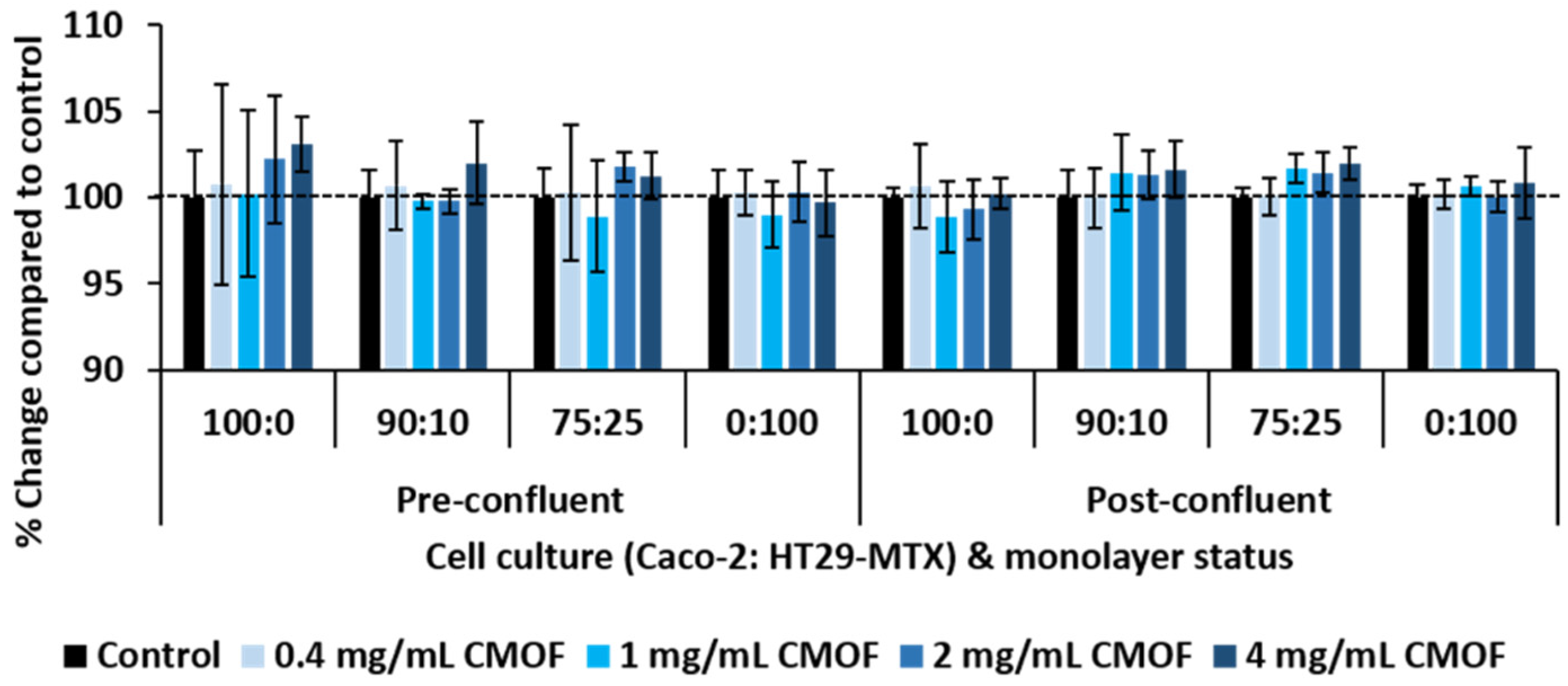
3.1. Metabolic Activity of Epithelial Cell Mono- and Co-Cultures in Response to CMOF
3.2. Impact of CMOF on TEER of Epithelial Cell Monolayers
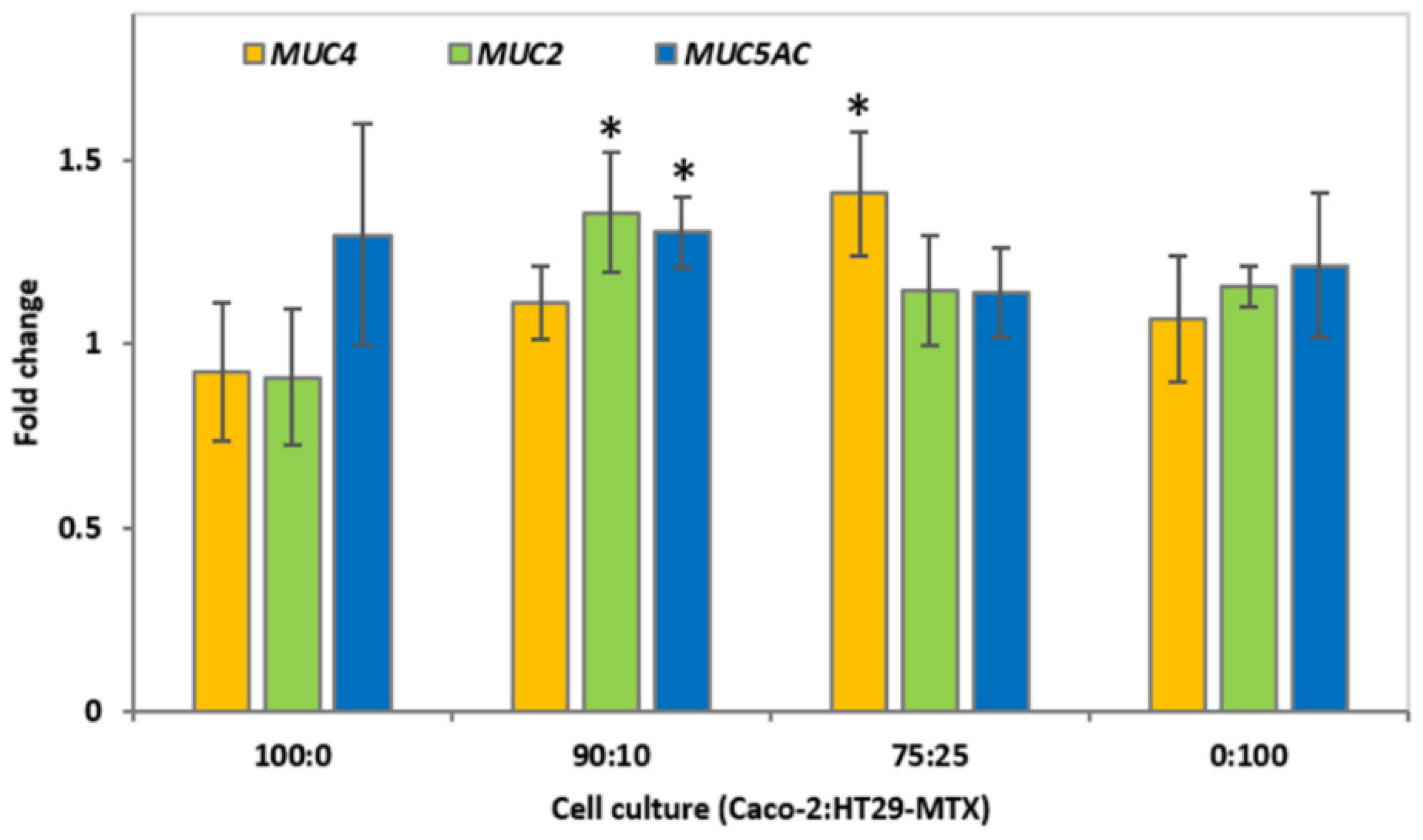
3.3. Mucin Gene Expression of Epithelial Cell Cultures in Response to CMOF
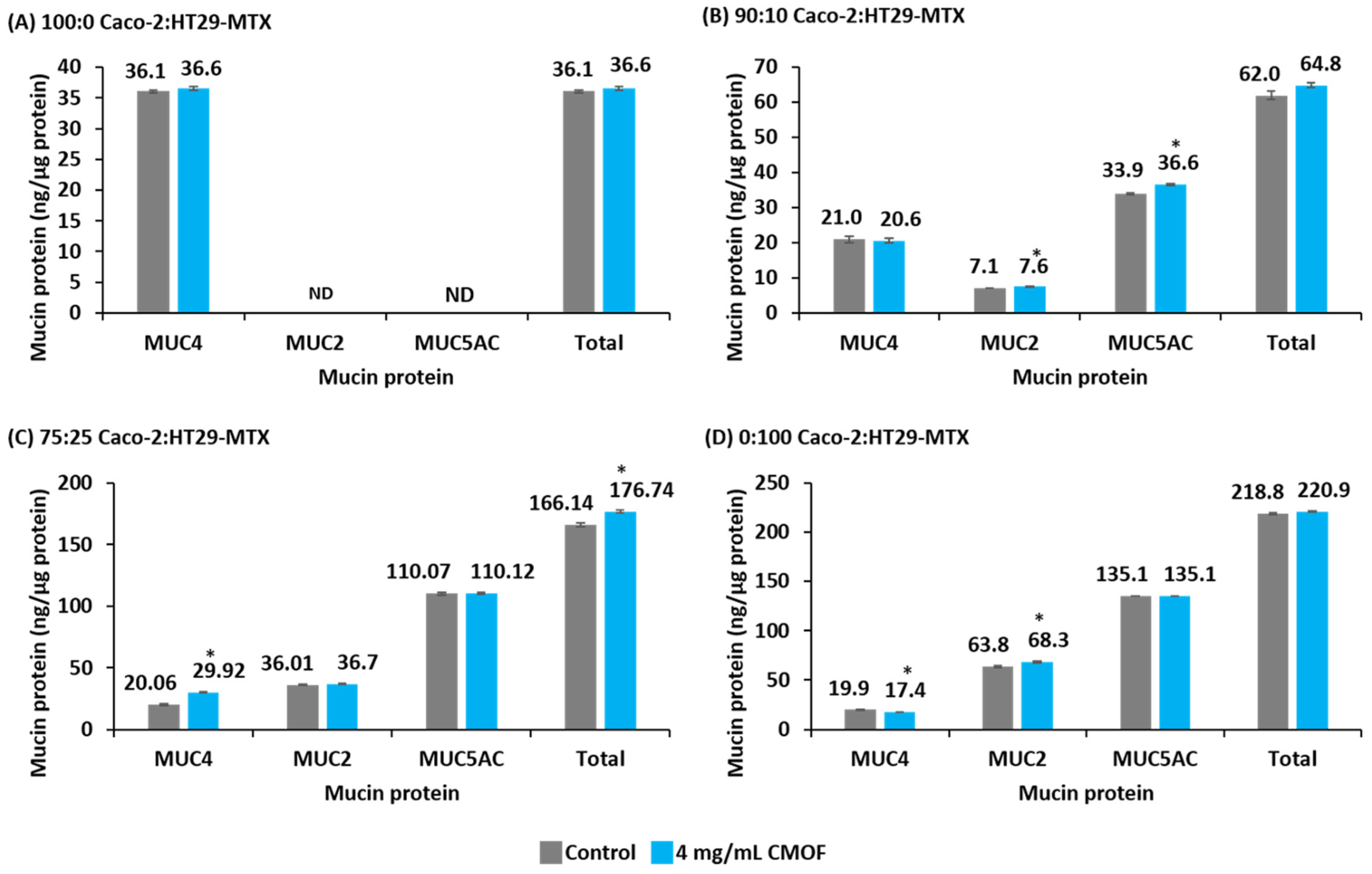
3.4. Mucin Protein Abundance of Epithelial Cell Cultures in Response to CMOF
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Smilowitz, J.T.; Lebrilla, C.B.; Mills, D.A.; German, J.B.; Freeman, S.L. Breast milk oligosaccharides: Structure-function relationships in the neonate. Annu. Rev. Nutr. 2014, 34, 143–169. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ferez, A.; Rudloff, S.; Guadix, A.; Henkel, C.A.; Pohlentz, G.; Boza, J.J.; Guadix, E.M.; Kunz, C. Goats’ milk as a natural source of lactose-derived oligosaccharides: Isolation by membrane technology. Int. Dairy J. 2006, 16, 173–181. [Google Scholar] [CrossRef]
- Lin, A.E.; Autran, C.A.; Espanola, S.D.; Bode, L.; Nizet, V. Human milk oligosaccharides protect bladder epithelial cells against uropathogenic Escherichia coli invasion and cytotoxicity. J. Infect. Dis. 2014, 209, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, S.; Rudloff, S.; Kunz, C. Oligosaccharides from human milk influence growth-related characteristics of intestinally transformed and non-transformed intestinal cells. Br. J. Nutr. 2008, 99, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.; Barnett, M.; McNabb, W.; Anderson, R.; Reynolds, G.; Roy, N. Post-weaning effects of milk and milk components on the intestinal mucosa in inflammation. Mutat. Res. Fundam. Mol. Mech. Mutag. 2010, 690, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.; Gabrielli, O.; Pierani, P.; Catassi, C.; Carlucci, A.; Giorgi, P.L. Changes in carbohydrate composition in human milk over 4 months of lactation. Pediatrics 1993, 91, 637–641. [Google Scholar] [PubMed]
- Kunz, C.; Rudloff, S. Biological functions of oligosaccharides in human milk. Acta Paediatr. 1993, 82, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Hasnain, S.Z.; Thornton, D.J.; Grencis, R.K. Changes in the mucosal barrier during acute and chronic Trichuris muris infection. Parasite Immunol. 2011, 33, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Gouyer, V.; Wiede, A.; Buisine, M.-P.; Dekeyser, S.; Moreau, O.; Lesuffleur, T.; Hoffmann, W.; Huet, G. Specific secretion of gel-forming mucins and TFF peptides in HT-29 cells of mucin-secreting phenotype. Biochim. Biophys. Acta Mol. Cell Res. 2001, 1539, 71–84. [Google Scholar] [CrossRef]
- McGuckin, M.A.; Eri, R.; Simms, L.A.; Florin, T.H.; Radford-Smith, G. Intestinal barrier dysfunction in inflammatory bowel diseases. Inflamm. Bowel Dis. 2009, 15, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ho, S. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Mahida, Y.R. Epithelial cell responses. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Hilgendorf, C.; Spahn-Langguth, H.; Regårdh, C.G.; Lipka, E.; Amidon Gordon, L.; Langguth, P. Caco-2 versus caco-2/HT29-MTX co-cultured cell lines: Permeabilities via diffusion, inside- and outside-directed carrier-mediated transport. J. Pharm. Sci. 2000, 89, 63–75. [Google Scholar] [CrossRef]
- Brittan, M.; Wright, N.A. The gastrointestinal stem cell. Cell Prolif. 2004, 37, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van de Wetering, M.; Clevers, H. The intestinal stem cell. Genes Dev. 2008, 22, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; Young, W.; McNabb, W.C.; van Baarlen, P.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Live Faecalibacterium prausnitzii in an apical anaerobic model of the intestinal epithelial barrier. Cell. Microbiol. 2015, 17, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two MUC2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Holmén Larsson, J.M.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 4659–4665. [Google Scholar] [CrossRef] [PubMed]
- Pelaseyed, T.; Bergstrom, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schutte, A.; van der Post, S.; Svensson, F.; Rodriguez-Pineiro, A.M.; Nystrom, E.E.L.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Prabhu, P.N.; Benefiel, A.C.; Miller, M.J.; Chow, J.; Davis, S.R.; Gaskins, H.R. Galacto-oligosaccharides may directly enhance intestinal barrier function through the modulation of goblet cells. Mol. Nutr. Food Res. 2015, 59, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Jakaitis, B.M.; Denning, P.W. Human breast milk and the gastrointestinal innate immune system. Clin. Perinatol. 2014, 41, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Heazlewood, C.K.; Cook, M.C.; Eri, R.; Price, G.R.; Tauro, S.B.; Taupin, D.; Thornton, D.J.; Png, C.W.; Crockford, T.L.; Cornall, R.J.; et al. Aberrant mucin assembly in mice causes endoplasmic reticulum stress and spontaneous inflammation resembling ulcerative colitis. PLoS Med. 2008, 5, e54. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Dalziel, J.E.; Gopal, P.K.; Bassett, S.; Ellis, A.; Roy, N.C. The role of intestinal barrier function in early life in the development of colitis. In Colitis; Fukata, D., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Cereijido, M.; Contreras, R.G.; Flores-Benítez, D.; Flores-Maldonado, C.; Larre, I.; Ruiz, A.; Shoshani, L. New diseases derived or associated with the tight junction. Arch. Med. Res. 2007, 38, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Lara-Villostada, F.; Debras, E.; Nieto, A.; Concha, A.; Galvez, J.; Lopez-Huertas, E.; Boza, J.; Obled, C.; Xaus, J. Oligosaccharides isolated from goat milk reduce intestinal inflammation in a rat model of dextran sodium sulfate-induced colitis. Clin. Nutr. 2006, 25, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Augustin, O.; Puerta, V.; Martinez-Ferez, A.; Baro, L.; Lopez-Huertas, E.; Suarez, M. Goat’s milk oligosaccharides modulate mucin and trefoil factors production in the mucus producing intestinal cell line HT29/MTX. Clin. Nutr. 2003, 22, S42. [Google Scholar] [CrossRef]
- Goehring, K.C.; Kennedy, A.D.; Prieto, P.A.; Buck, R.H. Direct evidence for the presence of human milk oligosaccharides in the circulation of breastfed infants. PLoS ONE 2014, 9, e101692. [Google Scholar] [CrossRef] [PubMed]
- Gnoth, M.J.; Rudloff, S.; Kunz, C.; Kinne, R.K.H. Investigations of the in vitro transport of human milk oligosaccharides by a Caco-2 monolayer using a novel high performance liquid chromatography-mass spectrometry technique. J. Biol. Chem. 2001, 276, 34363–34370. [Google Scholar] [CrossRef] [PubMed]
- Audie, J.P.; Janin, A.; Porchet, N.; Copin, M.C.; Gosselin, B.; Aubert, J.P. Expression of human mucin genes in respiratory, digestive, and reproductive tracts ascertained by in situ hybridization. J. Histochem. Cytochem. 1993, 41, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, T.; Longman, R.J.; Corfield, A.P.; Probert, C.S.J. Mucins and inflammatory bowel disease. Postgrad. Med. J. 2000, 76, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.; Harris, A. Developmental expression of mucin genes in the human gastrointestinal system. Gut 1998, 42, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Thum, C.; Cookson, A.; McNabb, W.C.; Roy, N.C.; Otter, D. Composition and enrichment of caprine milk oligosaccharides from New Zealand Saanen goat cheese whey. J. Food Compos. Anal. 2015, 42, 30–37. [Google Scholar] [CrossRef]
- Briske-Anderson, M.J.; Finley, J.W.; Newman, S.M. The influence of culture time and passage number on the morphological and physiological development of Caco-2 cells. Exp. Biol. Med. 1997, 214, 248–257. [Google Scholar] [CrossRef]
- Lesuffleur, T.; Barbat, A.; Dussaulx, E.; Zweibaum, A. Growth adaptation to methotrexate of HT-29 human colon carcinoma cells is associated with their ability to differentiate into columnar absorptive and mucus-secreting cells. Cancer Res. 1990, 50, 6334–6343. [Google Scholar] [PubMed]
- Lesuffleur, T.; Barbat, A.; Luccioni, C.; Beaumatin, J.; Clair, M.; Kornowski, A.; Dussaulx, E.; Dutrillaux, B.; Zweibaum, A. Dihydrofolate reductase gene amplification-associated shift of differentiation in methotrexate-adapted HT-29 cells. J. Cell Biol. 1991, 115, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Tajima, A.; Iwase, T.; Shinji, H.; Seki, K.; Mizunoe, Y. Inhibition of endothelial interleukin-8 production and neutrophil transmigration by Staphylococcus aureus β-hemolysin. Infect. Immunity 2009, 77, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Konsoula, R.; Barile, F.A. Correlation of in vitro cytotoxicity with paracellular permeability in Caco-2 cells. Toxicol. Vitro 2005, 19, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Maqueda, D.; Miralles, B.; De Pascual-Teresa, S.; Reverón, I.; Muñoz, R.; Recio, I. Food-derived peptides stimulate mucin secretion and gene expression in intestinal cells. J. Agric. Food Chem. 2012, 60, 8600–8605. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum DSM 2648 is a potential probiotic that enhances intestinal barrier function. FEMS Microbiol. Lett. 2010, 309, 184–192. [Google Scholar] [CrossRef] [PubMed]
- De Kok, J.B.; Roelofs, R.W.; Giesendorf, B.A.; Pennings, J.L.; Waas, E.T.; Feuth, T.; Swinkels, D.W.; Span, P.N. Normalization of gene expression measurements in tumor tissues: Comparison of 13 endogenous control genes. Lab. Investig. 2004, 85, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Lebrero-Fernández, C.; Wenzel, U.A.; Akeus, P.; Wang, Y.; Strid, H.; Simrén, M.; Gustavsson, B.; Börjesson, L.G.; Cardell, S.L.; Öhman, L.; et al. Altered expression of Butyrophilin (BTN) and BTN-like (BTNL) genes in intestinal inflammation and colon cancer. Immunity Inflamm. Dis. 2016. [Google Scholar] [CrossRef]
- McCann, M.J.; Rowland, I.R.; Roy, N.C. Anti-proliferative effects of physiological concentrations of enterolactone in models of prostate tumourigenesis. Mol. Nutr. Food Res. 2013, 57, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Gaj, S.; Eijssen, L.; Mensink, R.P.; Evelo, C.T.A. Validating nutrient-related gene expression changes from microarrays using RT2 PCR-arrays. Genes Nutr. 2008, 3, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Balimane, P.V.; Chong, S. Cell culture-based models for intestinal permeability: A critique. Drug Discov. Today 2005, 10, 335–343. [Google Scholar] [CrossRef]
- Sambuy, Y. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, M.; Vázquez, M.; Devesa, V.; Vélez, D. In vitro study of intestinal transport of inorganic and methylated arsenic species by Caco-2/HT29-MTX cocultures. Chem. Res. Toxicol. 2012, 25, 2654–2662. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, E.; Casselbrant, A.; Lundberg, S.; Fandriks, L. In vitro assessment of epithelial electrical resistance in human esophageal and jejunal mucosae and in Caco-2 cell layers. Scand. J. Gastroenterol. 2012, 47, 1321–1333. [Google Scholar] [CrossRef] [PubMed]
- Fleisher, D. Biological transport phenomena in the gastrointestinal tract. In Transport Processes in Pharmaceutical Systems; Informa Healthcare: New York, NY, USA, 1999; pp. 147–184. [Google Scholar]
- Soler, A.P.; Miller, R.D.; Laughlin, K.V.; Carp, N.Z.; Klurfeld, D.M.; Mullin, J.M. Increased tight junctional permeability is associated with the development of colon cancer. Carcinogenesis 1999, 20, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Wikman-Larhed, A.; Artursson, P. Co-cultures of human intestinal goblet (HT29-H) and absorptive (Caco-2) cells for studies of drug and peptide absorption. Eur. J. Pharm. Sci. 1995, 3, 171–183. [Google Scholar] [CrossRef]
- Fernandez Miyakawa, M.E.; Pistone Creydt, V.; Uzal, F.A.; McClane, B.A.; Ibarra, C. Clostridium perfringens enterotoxin damages the human intestine in vitro. Infect. Immunity 2005, 73, 8407–8410. [Google Scholar] [CrossRef] [PubMed]
- Rubas, W.; Cromwell, M.E.M.; Shahrokh, Z.; Villagran, J.; Nguyen, T.N.; Wellton, M.; Nguyen, T.H.; Mrsny, R.J. Flux measurements across Caco-2 monolayers may predict transport in human large intestinal tissue. J. Pharm. Sci. 1996, 85, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Various nondigestible saccharides open a paracellular calcium transport pathway with the induction of intracellular calcium signaling in human intestinal Caco-2 cells. J. Nutr. 2004, 134, 1935–1941. [Google Scholar] [PubMed]
- De Vos, M.; Huygelen, V.; Van Raemdonck, G.; Willemen, S.; Fransen, E.; Van Ostade, X.; Casteleyn, C.; Van Cruchten, S.; Van Ginneken, C. Supplementing formula-fed piglets with a low molecular weight fraction of bovine colostrum whey results in an improved intestinal barrier. J. Anim. Sci. 2014, 92, 3491–3501. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, S.; Kunz, C.; Rudloff, S. Oligosaccharides from human milk induce growth arrest via G2/M by influencing growth-related cell cycle genes in intestinal epithelial cells. Br. J. Nutr. 2009, 101, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Seth, A.; Elias, B.; Naren Anjaparavanda, P.; Rao, R. MAPK interacts with occludin and mediates EGF-induced prevention of tight junction disruption by hydrogen peroxide. Biochem. J. 2006, 393, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Flores-Benitez, D.; Rincon-Heredia, R.; Razgado, L.F.; Larre, I.; Cereijido, M.; Contreras, R.G. Control of tight junctional sealing: Roles of epidermal growth factor and prostaglandin E2. Am. J. Physiol. Cell Physiol. 2009, 297, C611–C620. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.M.; Singh, C.; Lang, J.D.; Strotman, L.N.; Alarid, E.T.; Beebe, D.J. Streamlining gene expression analysis: Integration of co-culture and mRNA purification. Integr. Biol. 2014, 6, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Bouwens, M.; Afman, L.A.; Müller, M. Fasting induces changes in peripheral blood mononuclear cell gene expression profiles related to increases in fatty acid β-oxidation: Functional role of peroxisome proliferator-activated receptor α in human peripheral blood mononuclear cells. Am. J. Clin. Nutr. 2007, 86, 1515–1523. [Google Scholar] [PubMed]
- Rodenburg, W.; Heidema, A.G.; Boer, J.M.A.; Bovee-Oudenhoven, I.M.J.; Feskens, E.J.M.; Mariman, E.C.M.; Keijer, J. A framework to identify physiological responses in microarray-based gene expression studies: Selection and interpretation of biologically relevant genes. Physiol. Genomics 2008, 33, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Perrais, M.; Pigny, P.; Copin, M.C.; Aubert, J.P.; Van Seuningen, I. Induction of MUC2 and MUC5AC mucins by factors of the epidermal growth factor (EGF) family is mediated by EGF receptor/Ras/Raf/Extracellular signal-regulated kinase cascade and Sp1*. J. Biol. Chem. 2002, 277, 32258–32267. [Google Scholar] [CrossRef] [PubMed]
- Homsi, M.E.; Ducroc, R.; Claustre, J.; Jourdan, G.; Gertler, A.; Estienne, M.; Bado, A.; Scoazec, J.-Y.; Plaisancie, P. Leptin modulates the expression of secreted and membrane-associated mucins in colonic epithelial cells by targeting PKC, PI3K, and MAPK pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G365–G373. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, M.G.; Guo, L.; Birchall, J.P.; Pearson, J.P. LPS up-regulates mucin and cytokine mRNA expression and stimulates mucin and cytokine secretion in goblet cells. Cell. Immunol. 2003, 221, 42–49. [Google Scholar] [CrossRef]
- McCool, D.J.; Forstner, J.F.; Forstner, G.G. Regulated and unregulated pathways for MUC2 mucin secretion in human colonic LS180 adenocarcinoma cells are distinct. Biochem. J. 1995, 312, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.-Y.M.; Allen, K.J.; Turner, P.C.; El-Nezami, H. Modulation of mucin mRNA (MUC5AC and MUC5B) expression and protein production and secretion in Caco-2/HT29-MTX co-cultures following exposure to individual and combined fusarium mycotoxins. Toxicol. Sci. 2014, 139, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Nollet, S.; Forgue-Lafitte, M.E.; Kirkham, P.; Bara, J. Mapping of two new epitopes on the apomucin encoded by MUC5AC gene: Expression in normal GI tract and colon tumors. Int. J. Cancer 2002, 99, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Legrier, M.E.; de Pinieux, G.; Boye, K.; Arvelo, F.; Judde, J.G.; Fontaine, J.J.; Bara, J.; Poupon, M.F. Mucinous differentiation features associated with hormonal escape in a human prostate cancer xenograft. Br. J. Cancer 2004, 90, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 236–252. [Google Scholar] [CrossRef] [PubMed]
- Pino, V.; Ramsauer, V.P.; Salas, P.; Carraway, C.A.C.; Carraway, K.L. Membrane mucin Muc4 induces density-dependent changes in ERK activation in mammary epithelial and tumor cells: Role in reversal of contact inhibition. J. Biol. Chem. 2006, 281, 29411–29420. [Google Scholar] [CrossRef] [PubMed]
- Andrianifahanana, M.; Moniaux, N.; Batra, S.K. Regulation of mucin expression: Mechanistic aspects and implications for cancer and inflammatory diseases. Biochim. Biophys. Acta Rev. Cancer 2006, 1765, 189–222. [Google Scholar] [CrossRef] [PubMed]
- Dharmani, P.; Srivastava, V.; Kissoon-Singh, V.; Chadee, K. Role of intestinal mucins in innate host defense mechanisms against pathogens. J. Innate Immunity 2009, 1, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Jarry, A.; Dorso, L.; Gratio, V.; Forgue-Lafitte, M.-E.; Laburthe, M.; Laboisse, C.L.; Darmoul, D. PAR-2 activation increases human intestinal mucin secretion through EGFR transactivation. Biochem. Biophys. Res. Commun. 2007, 364, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Davis, S.R.; Tappenden, K.A. Human milk oligosaccharides influence maturation of human Intestinal Caco-2Bbe and HT-29. Cell Lines J. Nutr. 2014, 144, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Hester, S.N.; Donovan, S.M. Individual and combined effects of nucleotides and human milk oligosaccharides on proliferation, apoptosis and necrosis in a human fetal intestinal cell line. Food Nutr. Sci. 2012, 3, 1567–1576. [Google Scholar] [CrossRef]
- Purup, S.; Vestergaard, M.; O Pedersen, L.; Sejrsen, K. Biological activity of bovine milk on proliferation of human intestinal cells. J. Dairy Res. 2007, 74, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Sakata, M.; Minekawa, R.; Yamamoto, T.; Hayashi, M.; Tasaka, K.; Murata, Y. Human milk induces fetal small intestinal cell proliferation involvement of a different tyrosine kinase signaling pathway from epidermal growth factor receptor. J. Endocrinol. 2004, 181, 449–457. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrate | Percentage (%) |
|---|---|
| Galacto-oligosaccharide | 0.4 |
| Lactose | 46.1 |
| Glucose | 12.0 |
| Galactose | 15.9 |
| Oligosaccharides | 25.6 |
| Oligosaccharide Common Name | Type | Percentage (%) |
|---|---|---|
| α-3′-Galactosyl-lactose or β-6′-Galactosyl-lactose | Neutral | 15 |
| 6′-N-Acetylglucosaminyl-lactose | Neutral | 3 |
| 6′-Sialyl-lactose or 3′-Sialyl-lactose | Acidic | 22 |
| 6′-Glycolyl-neuraminyl-lactose | Acidic | 42 |
| Disialyl-N-lactose | Acidic | 18 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnett, A.M.; Roy, N.C.; McNabb, W.C.; Cookson, A.L. Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium. Nutrients 2016, 8, 267. https://doi.org/10.3390/nu8050267
Barnett AM, Roy NC, McNabb WC, Cookson AL. Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium. Nutrients. 2016; 8(5):267. https://doi.org/10.3390/nu8050267
Chicago/Turabian StyleBarnett, Alicia M., Nicole C. Roy, Warren C. McNabb, and Adrian L. Cookson. 2016. "Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium" Nutrients 8, no. 5: 267. https://doi.org/10.3390/nu8050267
APA StyleBarnett, A. M., Roy, N. C., McNabb, W. C., & Cookson, A. L. (2016). Effect of a Semi-Purified Oligosaccharide-Enriched Fraction from Caprine Milk on Barrier Integrity and Mucin Production of Co-Culture Models of the Small and Large Intestinal Epithelium. Nutrients, 8(5), 267. https://doi.org/10.3390/nu8050267