Gastrointestinal Nutrient Infusion Site and Eating Behavior: Evidence for A Proximal to Distal Gradient within the Small Intestine?

Abstract
:1. Introduction
2. Site-Dependent Effects on GI Function
2.1. Motility
2.2. Small Intestinal Transit Time
2.3. Gastric Emptying Rate
3. Site-Dependent Effects on Food Intake and Satiety: Duodenal Brake
4. Site-Dependent Effects on Food Intake and Satiety: Jejunal Brake
5. Site-Dependent Effects on Food Intake and Satiety: Ileal Brake
6. The Effects of Length and Site of Intestinal Contact on Nutrient-Driven Satiety and Food Intake
7. The Effect of BMI, Age, and Gender on the Intestinal Brake Effect of Nutrients
8. Conclusions
Author Contributions
Conflicts of Interest
References
- WHO. Factsheet 311: Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 1 January 2015).
- Specchia, M.L.; Veneziano, M.A.; Cadeddu, C.; Ferriero, A.M.; Mancuso, A.; Ianuale, C.; Parente, P.; Capri, S.; Ricciardi, W. Economic impact of adult obesity on health systems: A systematic review. Eur. J. Public Health 2015, 25, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Read, N.W.; McFarlane, A.; Kinsman, R.I.; Bates, T.E.; Blackhall, N.W.; Farrar, G.B.; Hall, J.C.; Moss, G.; Morris, A.P.; O'Neill, B.; et al. Effect of infusion of nutrient solutions into the ileum on gastrointestinal transit and plasma levels of neurotensin and enteroglucagon. Gastroenterology 1984, 86, 274–280. [Google Scholar] [PubMed]
- Lin, H.C.; Zhao, X.T.; Wang, L. Intestinal transit is more potently inhibited by fat in the distal (ileal brake) than in the proximal (jejunal brake) gut. Dig. Dis. Sci. 1997, 42, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Layer, P.; Peschel, S.; Schlesinger, T.; Goebell, H. Human pancreatic secretion and intestinal motility: Effects of ileal nutrient perfusion. Am. J. Physiol. 1990, 258, G196–G201. [Google Scholar] [PubMed]
- Keller, J.; Holst, J.J.; Layer, P. Inhibition of human pancreatic and biliary output but not intestinal motility by physiological intraileal lipid loads. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G704–G709. [Google Scholar] [CrossRef] [PubMed]
- Layer, P.; Holst, J.J.; Grandt, D.; Goebell, H. Ileal release of glucagon-like peptide-1 (GLP-1). Association with inhibition of gastric acid secretion in humans. Dig. Dis. Sci. 1995, 40, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Welch, I.; Saunders, K.; Read, N.W. Effect of ileal and intravenous infusions of fat emulsions on feeding and satiety in human volunteers. Gastroenterology 1985, 89, 1293–1297. [Google Scholar] [PubMed]
- Maljaars, P.W. Intestinal Fat and Eating Behavior: Role of the Ileal Brake, Chapter 7: Both Intestinal Site and Timing of Fat Delivery Affect Appetite in Humans. Ph.D. Thesis, Maastricht University Medical Center, 22 October 2010; pp. 109–126. [Google Scholar]
- Maljaars, P.W.; Peters, H.P.; Mela, D.J.; Masclee, A.A. Ileal brake: A sensible food target for appetite control. A review. Physiol. Behav. 2008, 95, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Ingram, J.R.; McGill, A.T.; Poppitt, S.D. Lipids, chos, proteins: Can all macronutrients put a “brake” on eating? Physiol. Behav. 2013, 120, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Borgstrom, S.; Arborelius, M., Jr. Influence of a fatty acid on duodenal motility. Scand. J. Gastroenterol. 1975, 10, 599–601. [Google Scholar] [PubMed]
- Spiller, R.C.; Trotman, I.F.; Higgins, B.E.; Ghatei, M.A.; Grimble, G.K.; Lee, Y.C.; Bloom, S.R.; Misiewicz, J.J.; Silk, D.B. The ileal brake—Inhibition of jejunal motility after ileal fat perfusion in man. Gut 1984, 25, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Spiller, R.C.; Trotman, I.F.; Adrian, T.E.; Bloom, S.R.; Misiewicz, J.J.; Silk, D.B. Further characterisation of the “ileal brake” reflex in man—Effect of ileal infusion of partial digests of fat, protein, and starch on jejunal motility and release of neurotensin, enteroglucagon, and peptide YY. Gut 1988, 29, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Heddle, R.; Collins, P.J.; Dent, J.; Horowitz, M.; Read, N.W.; Chatterton, B.; Houghton, L.A. Motor mechanisms associated with slowing of the gastric emptying of a solid meal by an intraduodenal lipid infusion. J. Gastroenterol. Hepatol. 1989, 4, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M.; Dent, J. Disordered gastric emptying: Mechanical basis, assessment and treatment. Baillieres Clin. Gastroenterol. 1991, 5, 371–407. [Google Scholar] [CrossRef]
- Cook, C.G.; Andrews, J.M.; Jones, K.L.; Wittert, G.A.; Chapman, I.M.; Morley, J.E.; Horowitz, M. Effects of small intestinal nutrient infusion on appetite and pyloric motility are modified by age. Am. J. Physiol. 1997, 273, R755–R761. [Google Scholar] [PubMed]
- Lin, H.C.; Doty, J.E.; Reedy, T.J.; Meyer, J.H. Inhibition of gastric emptying by sodium oleate depends on length of intestine exposed to nutrient. Am. J. Physiol. 1990, 259, G1031–G1036. [Google Scholar] [PubMed]
- Maljaars, J.; Haddeman, E.; Peters, H.; Masclee, A.A. Comparison of ileal and duodenal brake mechanisms on satiety and gastrointestinal transport. Gastroenterology 2007, 132, A207. [Google Scholar]
- Jain, N.K.; Boivin, M.; Zinsmeister, A.R.; DiMagno, E.P. The ileum and carbohydrate-mediated feedback regulation of postprandial pancreaticobiliary secretion in normal humans. Pancreas 1991, 6, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Siegle, M.L.; Schmid, H.R.; Ehrlein, H.J. Effects of ileal infusions of nutrients on motor patterns of canine small intestine. Am. J. Physiol. 1990, 259, G78–G85. [Google Scholar] [PubMed]
- Welch, I.M.; Cunningham, K.M.; Read, N.W. Regulation of gastric emptying by ileal nutrients in humans. Gastroenterology 1988, 94, 401–404. [Google Scholar] [PubMed]
- Lavin, J.H.; Wittert, G.; Sun, W.M.; Horowitz, M.; Morley, J.E.; Read, N.W. Appetite regulation by carbohydrate: Role of blood glucose and gastrointestinal hormones. Am. J. Physiol. 1996, 271, E209–E214. [Google Scholar] [PubMed]
- Mei, N. Vagal glucoreceptors in the small intestine of the cat. J. Physiol. 1978, 282, 485–506. [Google Scholar] [CrossRef] [PubMed]
- Lavin, J.H.; Wittert, G.A.; Andrews, J.; Yeap, B.; Wishart, J.M.; Morris, H.A.; Morley, J.E.; Horowitz, M.; Read, N.W. Interaction of insulin, glucagon-like peptide 1, gastric inhibitory polypeptide, and appetite in response to intraduodenal carbohydrate. Am. J. Clin. Nutr. 1998, 68, 591–598. [Google Scholar] [PubMed]
- MacIntosh, C.G.; Horowitz, M.; Verhagen, M.A.; Smout, A.J.; Wishart, J.; Morris, H.; Goble, E.; Morley, J.E.; Chapman, I.M. Effect of small intestinal nutrient infusion on appetite, gastrointestinal hormone release, and gastric myoelectrical activity in young and older men. Am. J. Gastroenterol. 2001, 96, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Rayner, C.K.; Park, H.S.; Wishart, J.M.; Kong, M.; Doran, S.M.; Horowitz, M. Effects of intraduodenal glucose and fructose on antropyloric motility and appetite in healthy humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R360–R366. [Google Scholar] [PubMed]
- Ryan, A.T.; Feinle-Bisset, C.; Kallas, A.; Wishart, J.M.; Clifton, P.M.; Horowitz, M.; Luscombe-Marsh, N.D. Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2012, 96, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Soenen, S.; Giezenaar, C.; Hutchison, A.T.; Horowitz, M.; Chapman, I.; Luscombe-Marsh, N.D. Effects of intraduodenal protein on appetite, energy intake, and antropyloroduodenal motility in healthy older compared with young men in a randomized trial. Am. J. Clin. Nutr. 2014, 100, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.T.; Luscombe-Marsh, N.D.; Saies, A.A.; Little, T.J.; Standfield, S.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal lipid and protein on gut motility and hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2013, 98, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Luscombe-Marsh, N.D.; Little, T.J.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of l-tryptophan on ad libitum eating, antropyloroduodenal motility, glycemia, insulinemia, and gut peptide secretion in healthy men. J. Clin. Endocrinol. Metab. 2014, 99, 3275–3284. [Google Scholar] [CrossRef] [PubMed]
- Chaikomin, R.; Wu, K.L.; Doran, S.; Meyer, J.H.; Jones, K.L.; Feinle-Bisset, C.; Horowitz, M.; Rayner, C.K. Effects of mid-jejunal compared to duodenal glucose infusion on peptide hormone release and appetite in healthy men. Regul. Pept. 2008, 150, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Pilichiewicz, A.N.; Chaikomin, R.; Brennan, I.M.; Wishart, J.M.; Rayner, C.K.; Jones, K.L.; Smout, A.J.; Horowitz, M.; Feinle-Bisset, C. Load-dependent effects of duodenal glucose on glycemia, gastrointestinal hormones, antropyloroduodenal motility, and energy intake in healthy men. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E743–E753. [Google Scholar] [CrossRef] [PubMed]
- Chapman, I.M.; Goble, E.A.; Wittert, G.A.; Horowitz, M. Effects of small-intestinal fat and carbohydrate infusions on appetite and food intake in obese and nonobese men. Am. J. Clin. Nutr. 1999, 69, 6–12. [Google Scholar] [PubMed]
- Pilichiewicz, A.N.; Papadopoulos, P.; Brennan, I.M.; Little, T.J.; Meyer, J.H.; Wishart, J.M.; Horowitz, M.; Feinle-Bisset, C. Load-dependent effects of duodenal lipid on antropyloroduodenal motility, plasma CCK and PYY, and energy intake in healthy men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2170–R2178. [Google Scholar] [CrossRef] [PubMed]
- Pilichiewicz, A.N.; Little, T.J.; Brennan, I.M.; Meyer, J.H.; Wishart, J.M.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of load, and duration, of duodenal lipid on antropyloroduodenal motility, plasma CCK and PYY, and energy intake in healthy men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R668–R677. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, K.E.; Read, N.W.; French, S.J. Food intake responses to upper gastrointestinal lipid infusions in humans. Physiol. Behav. 1998, 64, 141–145. [Google Scholar] [CrossRef]
- Seimon, R.V.; Feltrin, K.L.; Meyer, J.H.; Brennan, I.M.; Wishart, J.M.; Horowitz, M.; Feinle-Bisset, C. Effects of varying combinations of intraduodenal lipid and carbohydrate on antropyloroduodenal motility, hormone release, and appetite in healthy males. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R912–R920. [Google Scholar] [CrossRef] [PubMed]
- Welch, I.M.; Sepple, C.P.; Read, N.W. Comparisons of the effects on satiety and eating behaviour of infusion of lipid into the different regions of the small intestine. Gut 1988, 29, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Feltrin, K.L.; Little, T.J.; Meyer, J.H.; Horowitz, M.; Rades, T.; Wishart, J.; Feinle-Bisset, C. Effects of lauric acid on upper gut motility, plasma cholecystokinin and peptide YY, and energy intake are load, but not concentration, dependent in humans. J. Physiol. 2007, 581, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Feltrin, K.L.; Little, T.J.; Meyer, J.H.; Horowitz, M.; Rades, T.; Wishart, J.; Feinle-Bisset, C. Comparative effects of intraduodenal infusions of lauric and oleic acids on antropyloroduodenal motility, plasma cholecystokinin and peptide YY, appetite, and energy intake in healthy men. Am. J. Clin. Nutr. 2008, 87, 1181–1187. [Google Scholar] [PubMed]
- Van Avesaat, M.; Troost, F.J.; Ripken, D.; Hendriks, H.F.; Masclee, A.A. Ileal brake activation: Macronutrient-specific effects on eating behavior? Int. J. Obes. 2015, 39, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Maljaars, P.W.; Symersky, T.; Kee, B.C.; Haddeman, E.; Peters, H.P.; Masclee, A.A. Effect of ileal fat perfusion on satiety and hormone release in healthy volunteers. Int. J. Obes. 2008, 32, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Maljaars, P.W.; Peters, H.P.; Kodde, A.; Geraedts, M.; Troost, F.J.; Haddeman, E.; Masclee, A.A. Length and site of the small intestine exposed to fat influences hunger and food intake. Br. J. Nutr. 2011, 106, 1609–1615. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Poppitt, S.D.; McCormack, D.; Buffenstein, R. Short-term effects of macronutrient preloads on appetite and energy intake in lean women. Physiol. Behav. 1998, 64, 279–285. [Google Scholar] [CrossRef]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [PubMed]
- Geraedts, M.C.; Troost, F.J.; Munsters, M.J.; Stegen, J.H.; de Ridder, R.J.; Conchillo, J.M.; Kruimel, J.W.; Masclee, A.A.; Saris, W.H. Intraduodenal administration of intact pea protein effectively reduces food intake in both lean and obese male subjects. PLoS ONE 2011, 6, e24878. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Meyer-Gerspach, A.C.; Beglinger, C. The role of the stomach in the control of appetite and the secretion of satiation peptides. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E666–E673. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.H.; Tecimer, S.N.; Shah, D.; Zafar, T.A. Protein source, quantity, and time of consumption determine the effect of proteins on short-term food intake in young men. J. Nutr. 2004, 134, 3011–3015. [Google Scholar] [PubMed]
- Andrews, J.M.; Doran, S.; Hebbard, G.S.; Rassias, G.; Sun, W.M.; Horowitz, M. Effect of glucose supplementation on appetite and the pyloric motor response to intraduodenal glucose and lipid. Am. J. Physiol. 1998, 274, G645–G652. [Google Scholar] [PubMed]
- Hunt, J.N.; Smith, J.L.; Jiang, C.L. Effect of meal volume and energy density on the gastric emptying of carbohydrates. Gastroenterology 1985, 89, 1326–1330. [Google Scholar] [PubMed]
- Wen, J.; Phillips, S.F.; Sarr, M.G.; Kost, L.J.; Holst, J.J. PYY and GLP-1 contribute to feedback inhibition from the canine ileum and colon. Am. J. Physiol. 1995, 269, G945–G952. [Google Scholar] [PubMed]
- Izukura, M.; Hashimoto, T.; Gomez, G.; Uchida, T.; Greeley, G.H., Jr.; Thompson, J.C. Intracolonic infusion of bile salt stimulates release of peptide YY and inhibits cholecystokinin-stimulated pancreatic exocrine secretion in conscious dogs. Pancreas 1991, 6, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, H.S.; Ferri, G.L.; Sarson, D.L.; Polak, J.M.; Bloom, S.R. The effects of ileal transposition and jejunoileal bypass on food intake and GI hormone levels in rats. Physiol. Behav. 1984, 33, 601–609. [Google Scholar] [CrossRef]
- Chen, D.C.; Stern, J.S.; Atkinson, R.L. Effects of ileal transposition on food intake, dietary preference, and weight gain in zucker obese rats. Am. J. Physiol. 1990, 258, R269–R273. [Google Scholar] [PubMed]
- Payne, J.H.; DeWind, L.T. Surgical treatment of obesity. Am. J. Surg. 1969, 118, 141–147. [Google Scholar] [CrossRef]
- Pilkington, T.R.; Gazet, J.C.; Ang, L.; Kalucy, R.S.; Crisp, A.H.; Day, S. Explanations for weight loss after ileojejunal bypass in gross obesity. Br. Med. J. 1976, 1, 1504–1505. [Google Scholar] [CrossRef] [PubMed]
- Griffen, W.O., Jr.; Bivins, B.A.; Bell, R.M. The decline and fall of the jejunoileal bypass. Surg. Gynecol. Obstet. 1983, 157, 301–308. [Google Scholar] [PubMed]
- Layer, P.; Groger, G. Fate of pancreatic enzymes in the human intestinal lumen in health and pancreatic insufficiency. Digestion 1993, 54, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Feinle, C.; O'Donovan, D.; Doran, S.; Andrews, J.M.; Wishart, J.; Chapman, I.; Horowitz, M. Effects of fat digestion on appetite, apd motility, and gut hormones in response to duodenal fat infusion in humans. Am J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G798–G807. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.H.; Tabrizi, Y.; DiMaso, N.; Hlinka, M.; Raybould, H.E. Length of intestinal contact on nutrient-driven satiety. Am. J. Physiol. 1998, 275, R1308–R1319. [Google Scholar] [PubMed]
- Maljaars, J. Overeating makes the gut grow fonder; new insights in gastrointestinal satiety signaling in obesity. Curr. Opin. Gastroenterol. 2013, 29, 177–183. [Google Scholar] [CrossRef] [PubMed]
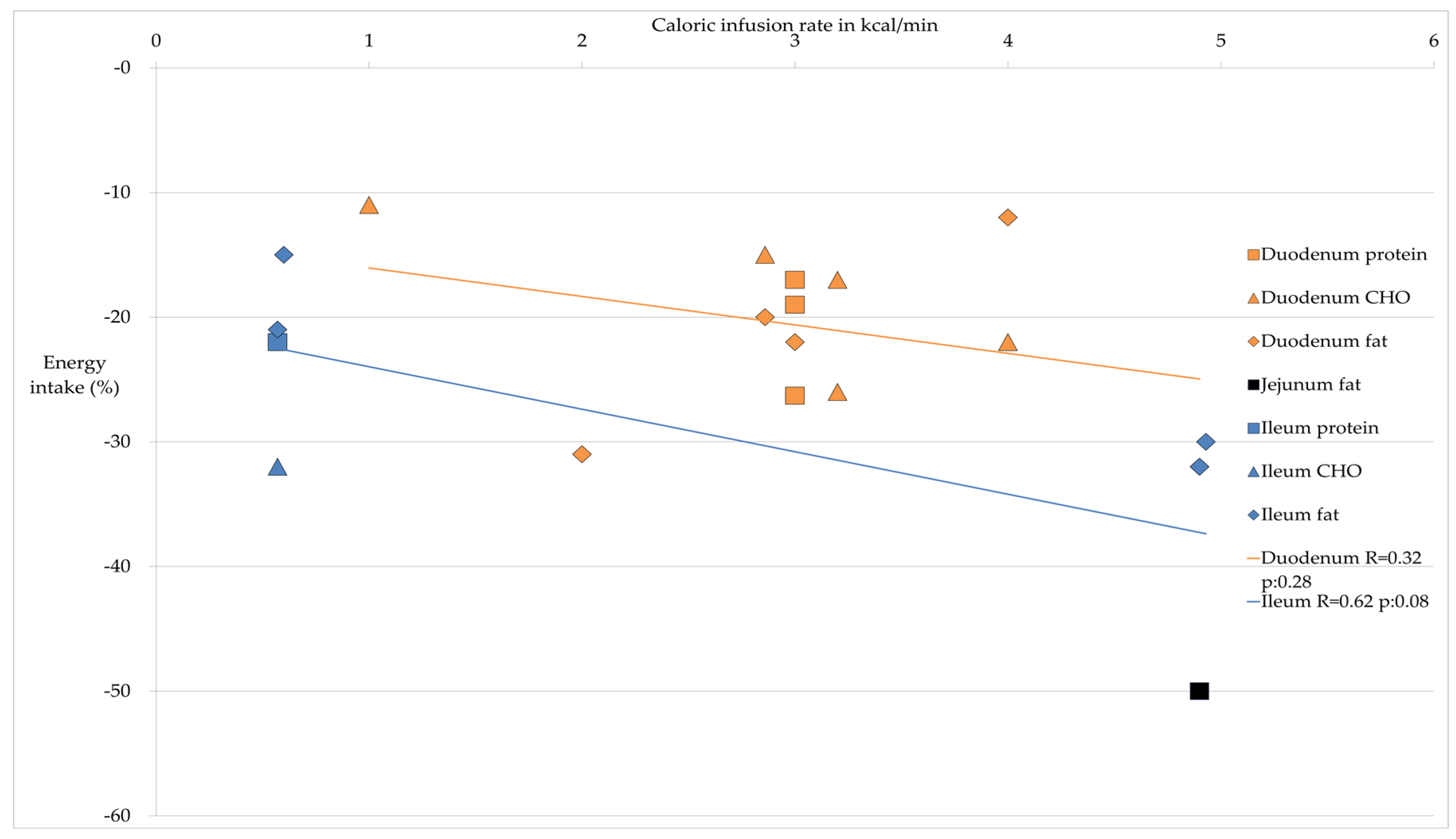

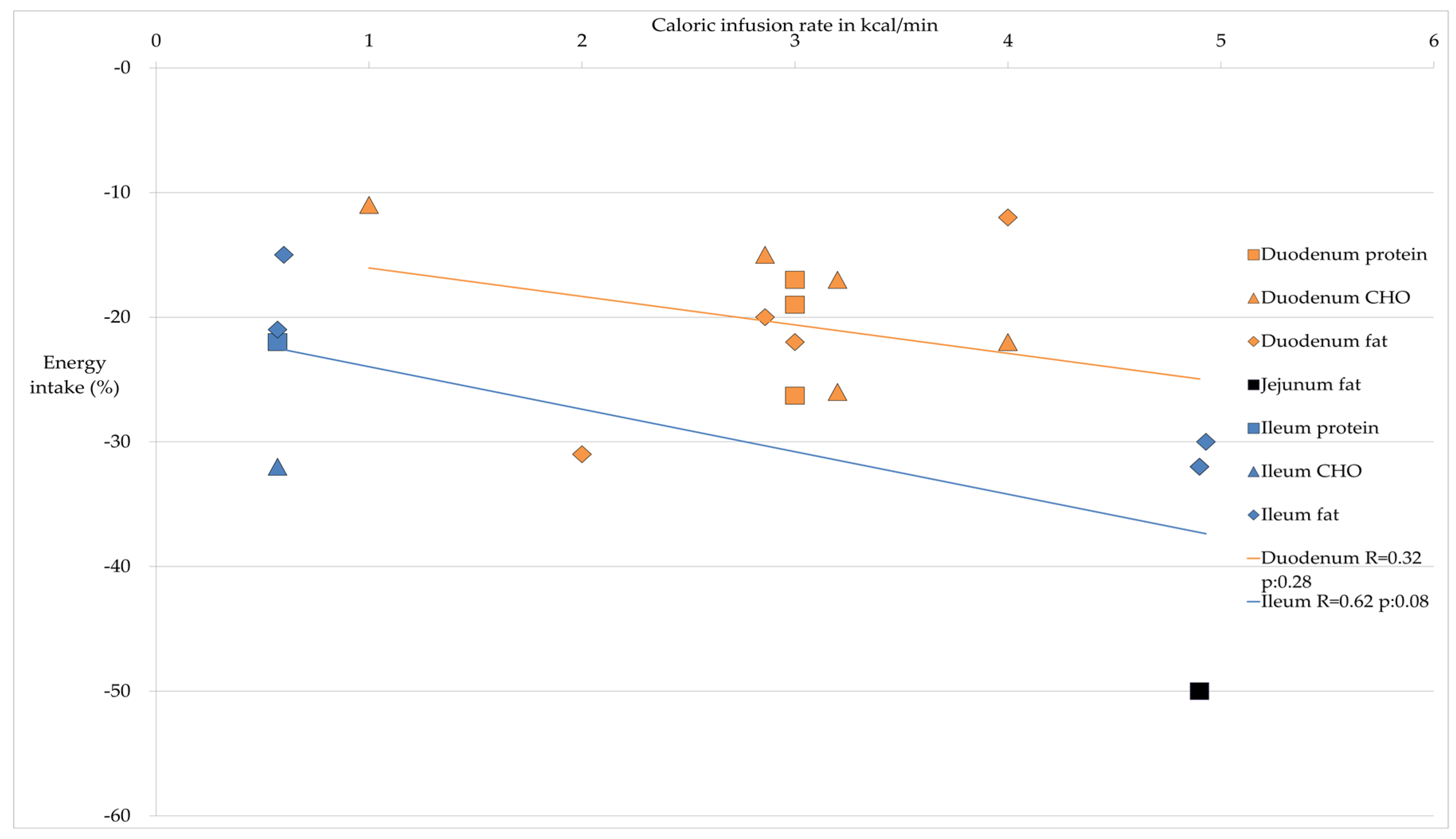{kind=link}
| Site Nutrient | Caloric Load, kcal/min; Total Amount of Calories, kcal; Nutrient Specifications | Effect on Satiety, Appetite, Hunger, Fullness, and Desire to Eat | Energy Intake, kcal (Mean, SEM) | Energy Intake (± %) | Reference |
|---|---|---|---|---|---|
| Duodenum | |||||
| Protein | 0.5 kcal/min; 30 kcal; 18.1% hydrolyzed whey protein | No decreased hunger | 1191 ± 113 kcal (control 1237 ± 111 kcal) ¤ | −2.7% | [28] |
| 0.5 kcal/min; 30 kcal; 18.1% hydrolyzed whey protein | No decreased hunger No increased fullness | 1153 ± 151 kcal (control 1270 ± 150 kcal) ¤ | −9% | [29] | |
| 1.5 kcal/min; 90 kcal; 18.1% hydrolyzed whey protein | No decreased hunger | 1077 ± 125 kcal (control 1237 ± 111 kcal) ¤ | −13% | [28] | |
| 1.5 kcal/min; 90 kcal; 18.1% hydrolyzed whey protein | No decreased hunger No increased fullness | 1118 ± 163 kcal (control 1270 ± 150 kcal) ¤ | −12% | [29] | |
| 3.0 kcal/min; 270 kcal; 18.5% Whey protein isolate | No decreased hunger | 1032 ± 83 kcal (control 1241 ± 80 kcal) * | −17% | [30] | |
| 3.0 kcal/min; 180 kcal; 18.1% hydrolyzed whey protein | No decreased hunger No increased fullness | 1031 ± 153 kcal (control 1270 ± 150 kcal) * | −19% | [29] | |
| 3.0 kcal/min; 180 kcal/min; 18.1% hydrolyzed whey protein | No decreased hunger | 912 ± 120 kcal (control 1237 ± 111 kcal) * | −26.3% | [28] | |
| Amino acid | 0.075 kcal/min; 6.75 kcal; l-tryptophan | Increased fullness * | 1155 ± 109 kcal (control 1215 ± 107 kcal) ¤ | −4.9% | [31] |
| 0.15 kcal/min; 13.5 kcal; l-tryptophan | Increased fullness * Increased desire to eat * | 996 ± 122 kcal(control 1215 ± 107 kcal) ** | −19% | [31] | |
| CHO | 1 kcal/min; 90 kcal Glucose 5% | Decreased hunger (compared to jejunal infusion) * | 1252 ± 97 kcal (compared to mid-jejunal CHO 1413 ± 62 kcal) ¥ | −11% | [32] |
| 1 kcal/min; 120 kcal Glucose 25% | No objective | 1195 ± 61 kcal(control 1062 ± 118 kcal) | +12.5% | [33] | |
| 2 kcal/min; 240 kcal; Glucose 25% | No objective | 1200 ± 87 kcal(control 1062 ± 118 kcal) | +13% | [33] | |
| 2 kcal/min; 180 kcal; Glucose 25% | Suppression desire to eat * | No significant decrease food intake | Kcal energy intake not mentioned | [27] | |
| 2 kcal/min; 180 kcal; Fructose 25% | No suppression desire to eat | Significant decrease food intake * | Kcal energy intake not mentioned | [27] | |
| 2.86 kcal/min; 343.2 kcal; Glucose 25% | Increased fullness ** | 896.9 ± 94.6 kcal (control 960.2 ± 85.5 kcal) ¤ | −3.6% | [34] | |
| 2.86 kcal/min; 343.2 kcal; Glucose 25% | Decreased hunger *** | 821.0 ± 61.1 kcal (control 964.0 ± 56.1 kcal) ** | −15% | [26] | |
| 2.9 kcal/min; 348 kcal; Glucose 25% | Increased fullness * | No significant decrease in energy intake compared to fat | Kcal energy intake not mentioned | [17] | |
| 3.2 kcal/min; 288 kcal; Glucose 20% | Decreased hunger ** Increased fullness ** | 1136.2 kcal (control 1536.4 kcal) ** | −26% | [25] | |
| 3.2 kcal/min; 288 kcal; Glucose 20% | Decreased hunger * Increased fullness * Increased satiety * | 907 ± 150 kcal (control 1093 ± 152 kcal) * | −17% | [23] | |
| 4 kcal/min; 480 kcal; Glucose 25% | No objective | 939.9 ± 114.6 kcal (control 1061.4 ± 11 [33] 7.5 kcal) *** | −22% | [33] | |
| Fat | 0.25 kcal/min; 25 kcal; 10% Intralipid | Decreased hunger* | 1282 ± 44 kcal (control 1289 ± 62 kcal) ¤ | −0.5% | [35] |
| 0.9 kcal/min; 54 kcal; 10% high oleic rapeseed oil in water | No significant effect on satiety | No significant decrease energy intake compared to ileal infusion | Kcal energy intake not mentioned | [9] | |
| 1.33 kcal/min; 66.5 kcal; 10% Intralipid | No significant difference in appetite scores | 1346 ± 109 kcal (control 1346 ± 90 kcal) ¤ | 0% | [36] | |
| 1.33 kcal/min; 199.5 kcal; 10% Intralipid | No significant difference in appetite scores | 1242 ± 84 kcal (control 1346 ± 90 kcal) ¤ | −7.7% | [36] | |
| 1.5 kcal/min; 75 kcal; 10% Intralipid | Decreased hunger * | 1235 ± 71 kcal (control 1289 ± 62 kcal) ¤ | −4.2% | [35] | |
| 2.0 kcal/min; 189 kcal; 20% Intralipid | No significant effect on satiety | 359 ± 51 kcal (control 521 ± 80 kcal) ** | −31% | [37] | |
| 2.86 kcal/min; 343.2 kcal; 10% Intralipid | Increased fullness *** | 742.6 ± 120.9 kcal (control 960.2 ± 85.5 kcal) ¤ | −22% | [34] | |
| 2.86 kcal/min; 343.2 kcal; 10% Intralipid | No significant difference in appetite scores | 775.4 ± 80.3 kcal (control 964.0 ± 56.1 kcal) ** | −20% | [26] | |
| 2.9 kcal/min; 348 kcal; 10% Intralipid | Decreased hunger ** Increased fullness * | Significant decrease in food intake compared to duodenal glucose * | Kcal energy intake not mentioned | [17] | |
| 3.0 kcal/min; 270 kcal; 20% Intralipid | No decrease in hunger | 973 ± 103 kcal (control 1241 ± 80 kcal) * | −22% | [30] | |
| 3.0 kcal/min; 270 kcal; 20% Intralipid | Suppression of desire to eat * | 1012.7 ± 107.8 kcal (compared to L2: 1263.5 ± 110.1 kcal and L1: 1303.9 ± 78.8 kcal) * | −22% | [38] | |
| 4 kcal/min; 200 kcal; 10% Intralipid | Decreased hunger * | 1139 ± 65 kcal (control 1289 ± 62 kcal) * | −12% | [35] | |
| 4 kcal/min; 200 kcal; 10% Intralipid | No significant difference in appetite scores | 1358 ± 132 kcal (control 1346 ± 90 kcal) ¤ | −1% | [36] | |
| 4.9 kcal/min; 367.5 kcal; 50% corn oil emulsion | Increased fullness * | 1377 ± 79 kcal (control 2026 ± 194 kcal) * | −32% | [39] | |
| Fatty acid | 0.2 kcal/min; 18 kcal; Lauric acid (C12) | No significant difference in appetite scores | 1341 ± 130 kcal | No significant difference compared to 0.3 or 0.4 kcal/min | [40] |
| 0.3 kcal/min; 27 kcal Lauric acid (C12) | No significant difference in appetite scores | 1312 ± 65 kcal | No significant difference compared to 0.2 kcal/min | [40] | |
| 0.4 kcal/min; 36 kcal; Lauric acid (C12) | No significant difference in appetite scores | 1077.2 ± 112.3 kcal (1339.9 ± 130.2 kcal) * | −20% (compared to 0.2 kcal/min) | [40] | |
| 0.4 kcal/min; 24 kcal; Lauric acid (C12) | No significant difference in appetite scores | 1134 ± 80 kcal (control 1265 ± 92 kcal) * | −10% | [41] | |
| 0.4 kcal/min; 24 kcal; Oleic acid (C18) | No significant difference in appetite scores | 1249 ± 72 kcal (control 1265 ± 92 kcal) ¤ | −1% | [41] | |
| Jejunum | |||||
| Protein | No data | No data | No data | No data | |
| CHO | 1 kcal/min; 90 kcal; Glucose 5% | Increased hunger and desire to eat (compared to duodenal infusion) * | 1413 ± 62 kcal (compared to duodenal infusion 1252 ± 97 kcal) ¥ | −11% (compared to duodenal infusion) | [32] |
| Fat | 4.9 kcal/min; 367.5 kcal; 50% corn oil emulsion | Decreased hunger * | 1076 ± 202 kcal (control 2157 ± 196 kcal) ** | −50% | [39] |
| Ileum | |||||
| Protein | 0.19 kcal/min; 17.1 kcal; Casein | No significant difference in appetite scores | 528.4 ± 86.1 kcal (control 586.7 ± 70.2 kcal) ¤ | −9.9% | [42] |
| 0.57 kcal/min; 51.3 kcal; Casein | Decreased hunger *** | 458.0 ± 78.6 kcal (control 586.7 ± 70.2 kcal) *** | −22% | [42] | |
| CHO | 0.19 kcal/min; 17.1 kcal; Sucrose | No significant difference in appetite scores | 491.4 ± 77.5 kcal (control 586.7 ± 70.2 kcal) ¤ | −21% | [42] |
| 0.57 kcal/min; 51.3 kcal; Sucrose | No significant difference in appetite scores | 399.0 ± 57.0 kcal (control 586.7 ± 70.2 kcal) *** | −32% | [42] | |
| Fat | 0.57 kcal/min; 51.3 kcal; Safflower oil | No significant difference in appetite scores | 464.3 ± 90.7 kcal (control 586.7 ± 70.2 kcal) *** | −21% | [42] |
| 0.6 kcal/min; 27 kcal; Safflower oil 6% | Increased satiety * | No objective | No objective | [43] | |
| 0.6 kcal/min; 54 kcal; Oil 10% | Decreased hunger * | 422 kcal (control 499 kcal) ** | −15% | [44] | |
| 0.9 kcal/min; 54 kcal; 10% high oleic rapeseed oil in water | Decreased hunger * | No significant decrease energy intake compared to duodenal infusion | Kcal energy intake not mentioned | [9] | |
| 1.8 kcal/min; 60.75 kcal; Safflower oil 6% | Increased satiety * | No objective | No objective | [43] | |
| 4.9 kcal/min; 367.5 kcal; 50% corn oil emulsion | Increased fullness * | 1377 kcal (control 2026 kcal) * | −32% | [39] | |
| 4.93 kcal/min; 370 kcal; 50% corn oil | No significant difference in appetite scores | 1313 kcal (control 1883 kcal) | −30% | [8] |
| Site Nutrient | Average Decrease Energy Intake in %; Average Caloric Infusion Rate in kcal/min | Energy Intake | Reference |
|---|---|---|---|
| Duodenum | |||
| Protein | 20.8%; 3.0 kcal/min | ↓ | [28,29,30] |
| CHO | 20%; 2.85 kcal/min | ↓ | [23,25,26,32,33] |
| Fat | 21.5%; 3.69 kcal/min | ↓ | [26,30,35,37,39] |
| Jejunum | |||
| Protein | * | * | |
| CHO | 11%; 1 kcal/min | ↓↓ ° | [32] |
| Fat | 50%; 4.9 kcal/min | ↓↓ | [39] |
| Ileum | |||
| Protein | 22%; 0.57 kcal/min | ↓↓↓ | [42] |
| CHO | 32%; 0.57 kcal/min | ↓↓↓ | [42] |
| Fat | 21.7%; 2.02 kcal/min | ↓↓ | [39,42,44] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alleleyn, A.M.E.; Van Avesaat, M.; Troost, F.J.; Masclee, A.A.M. Gastrointestinal Nutrient Infusion Site and Eating Behavior: Evidence for A Proximal to Distal Gradient within the Small Intestine? Nutrients 2016, 8, 117. https://doi.org/10.3390/nu8030117
Alleleyn AME, Van Avesaat M, Troost FJ, Masclee AAM. Gastrointestinal Nutrient Infusion Site and Eating Behavior: Evidence for A Proximal to Distal Gradient within the Small Intestine? Nutrients. 2016; 8(3):117. https://doi.org/10.3390/nu8030117
Chicago/Turabian StyleAlleleyn, Annick M. E., Mark Van Avesaat, Freddy J. Troost, and Adrian A. M. Masclee. 2016. "Gastrointestinal Nutrient Infusion Site and Eating Behavior: Evidence for A Proximal to Distal Gradient within the Small Intestine?" Nutrients 8, no. 3: 117. https://doi.org/10.3390/nu8030117
APA StyleAlleleyn, A. M. E., Van Avesaat, M., Troost, F. J., & Masclee, A. A. M. (2016). Gastrointestinal Nutrient Infusion Site and Eating Behavior: Evidence for A Proximal to Distal Gradient within the Small Intestine? Nutrients, 8(3), 117. https://doi.org/10.3390/nu8030117