Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Diabetes Mellitus
2.3. Physical Training
2.4. Administration of Beta-Glucans
2.5. Collection of Biological Material and Assessment of the Atherogenic Index of Plasma
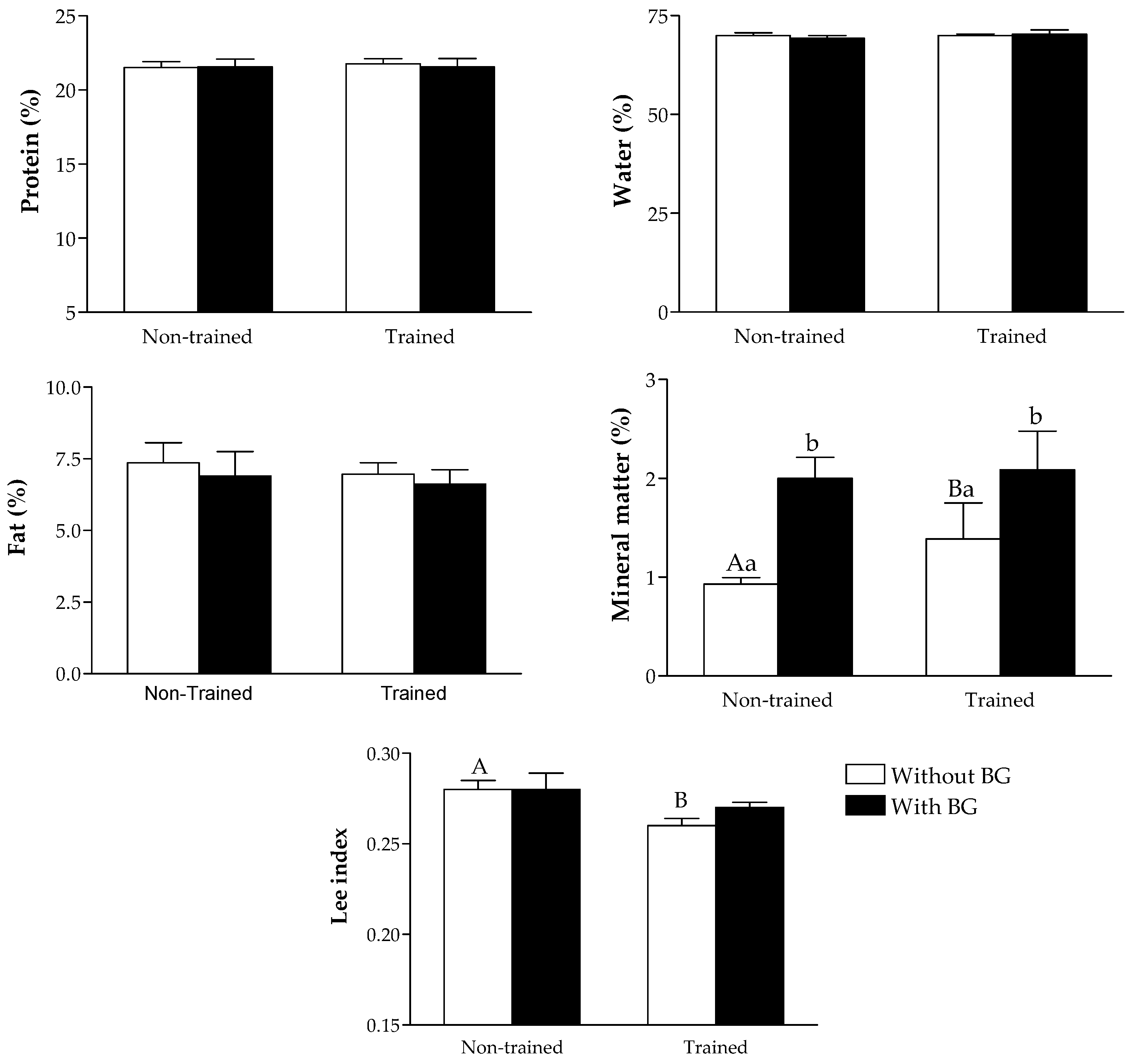
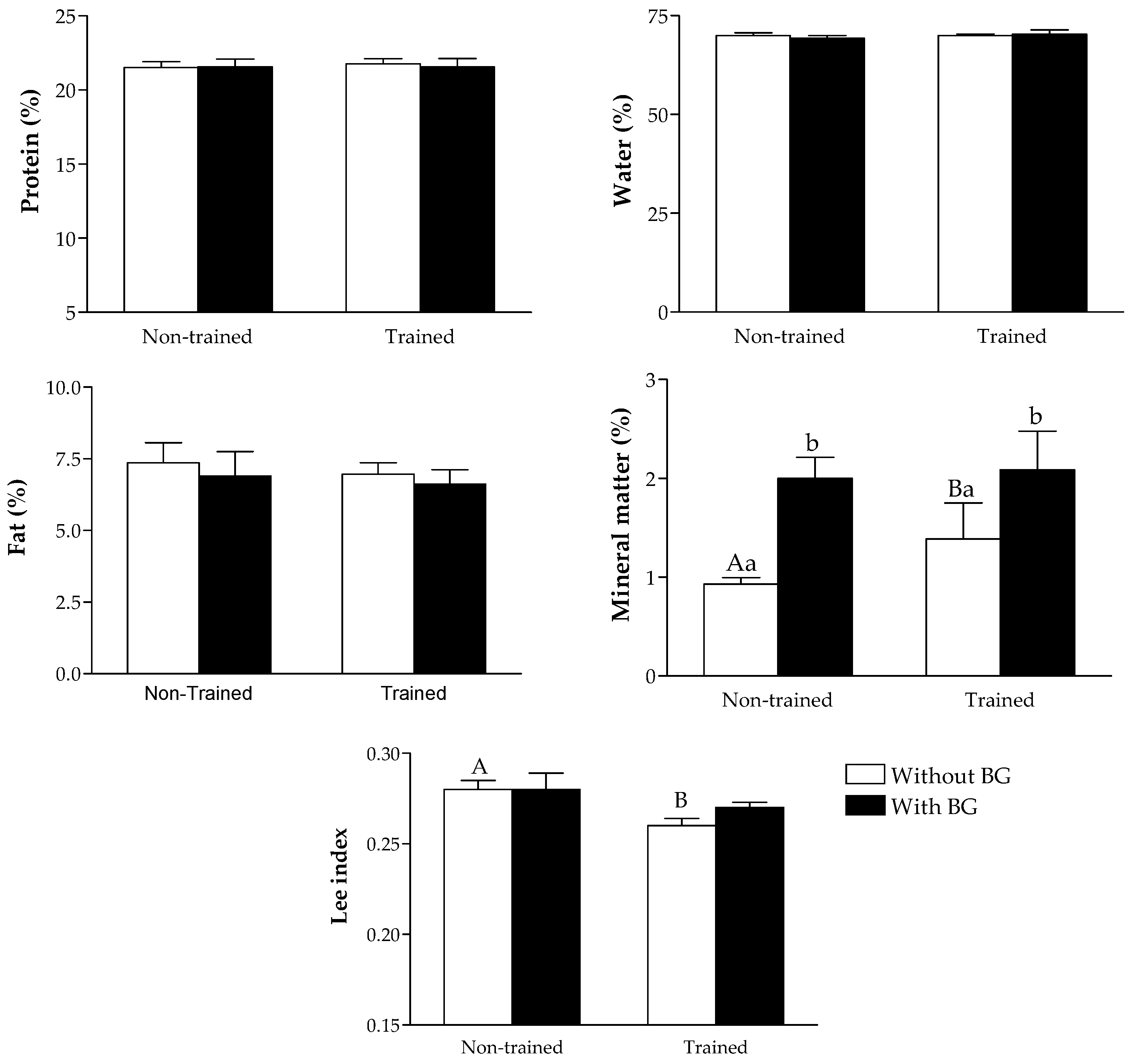
2.6. Lee Index Assessment and Chemical Composition of the Body
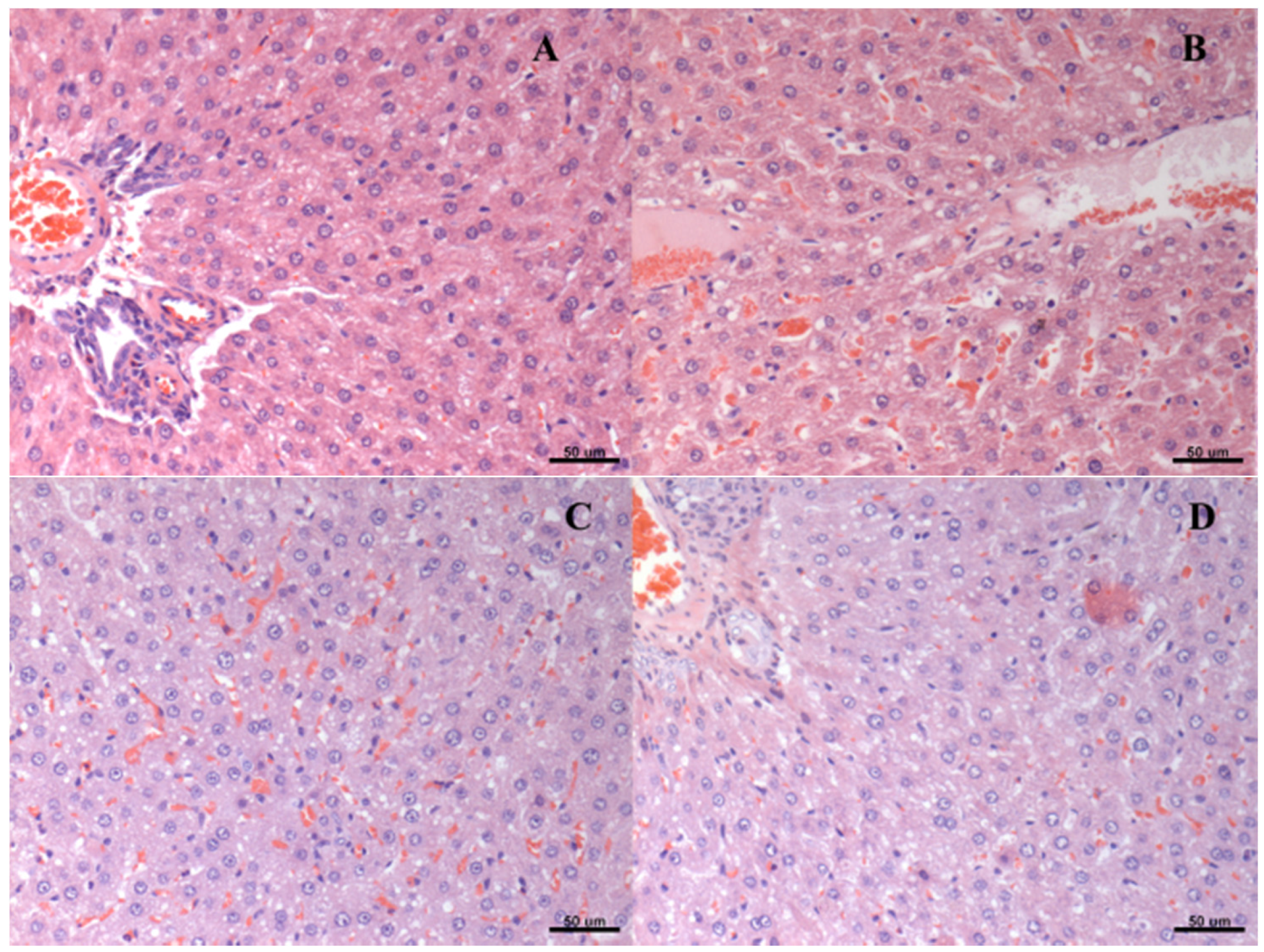
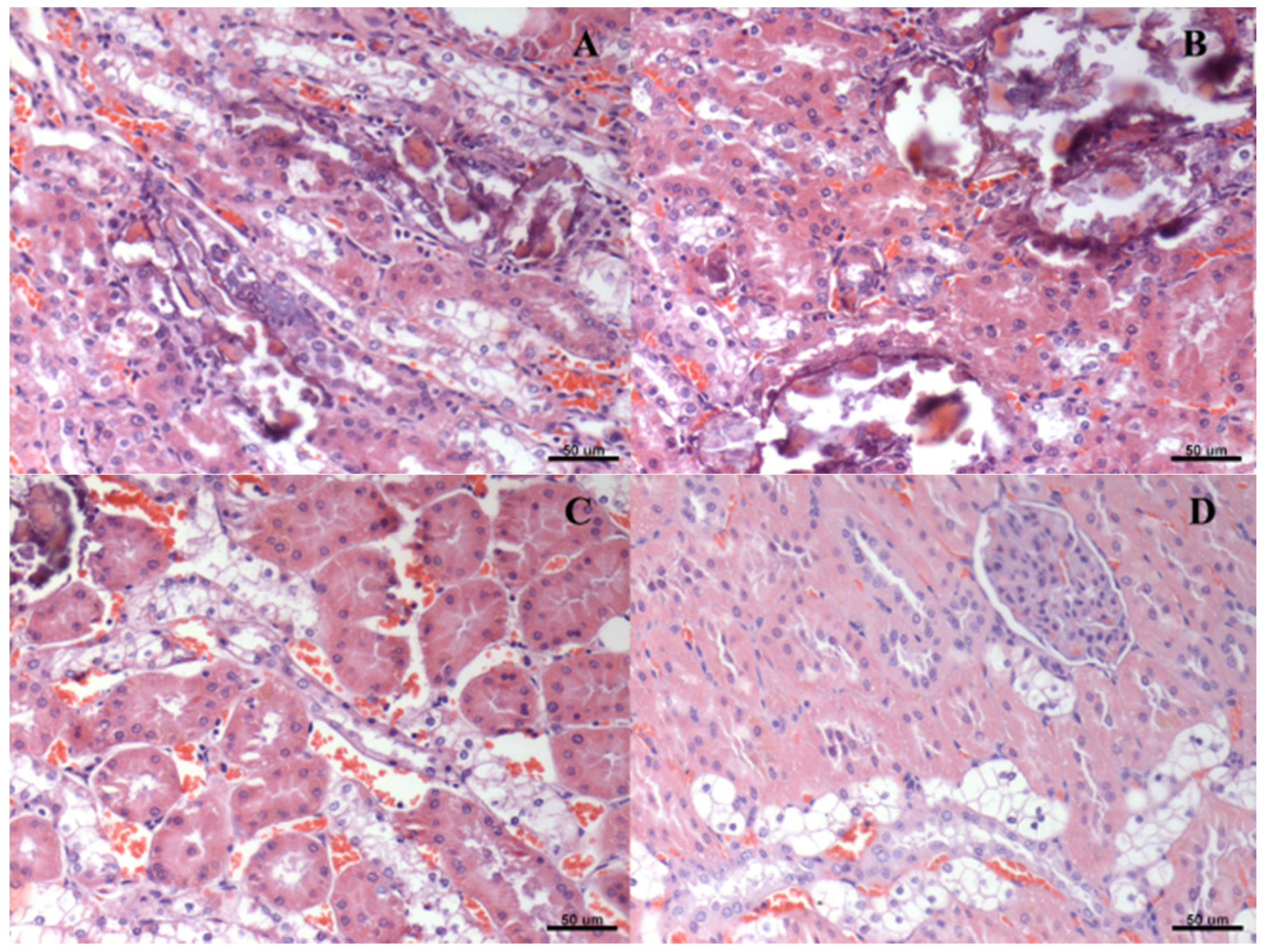
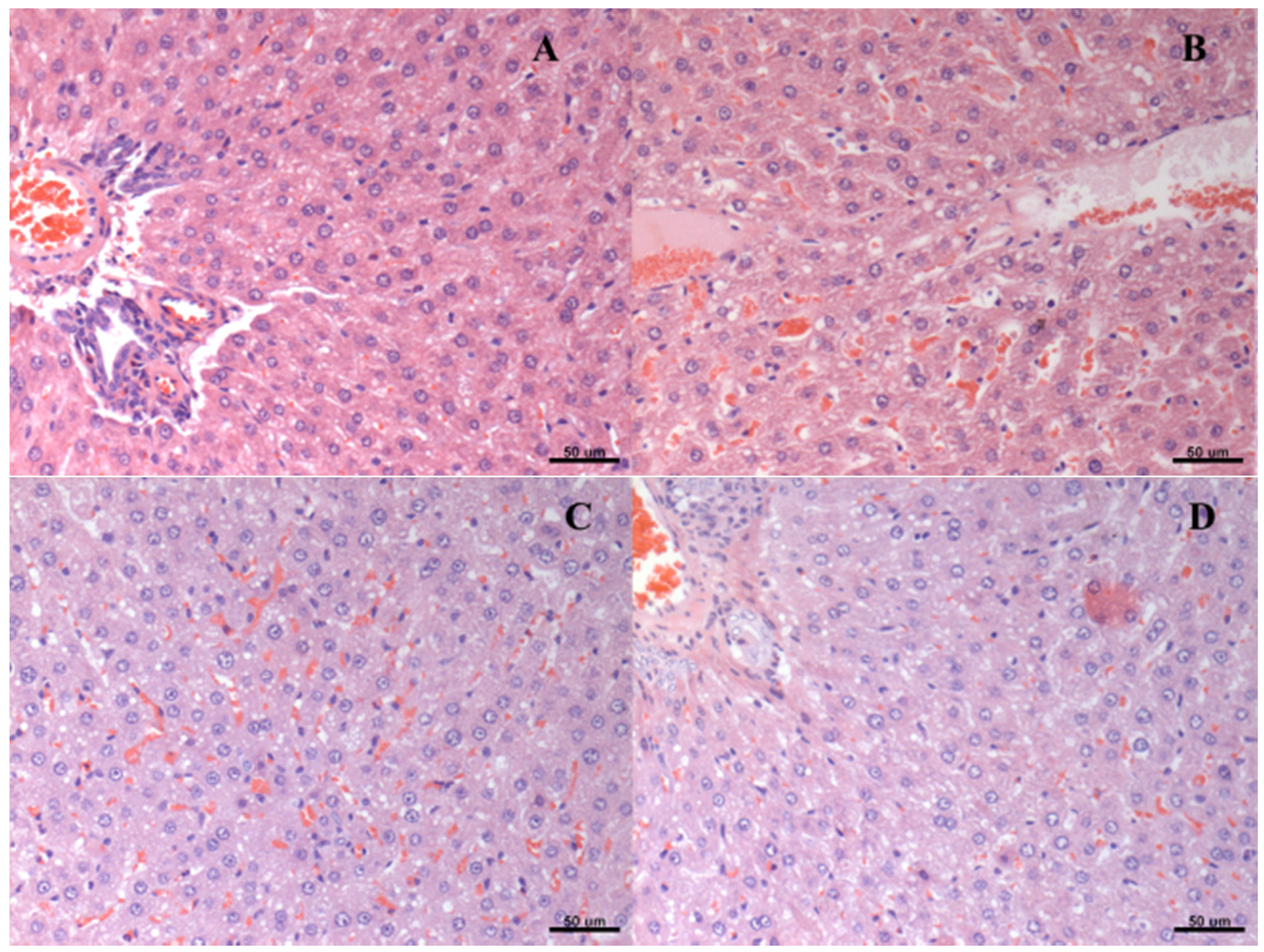
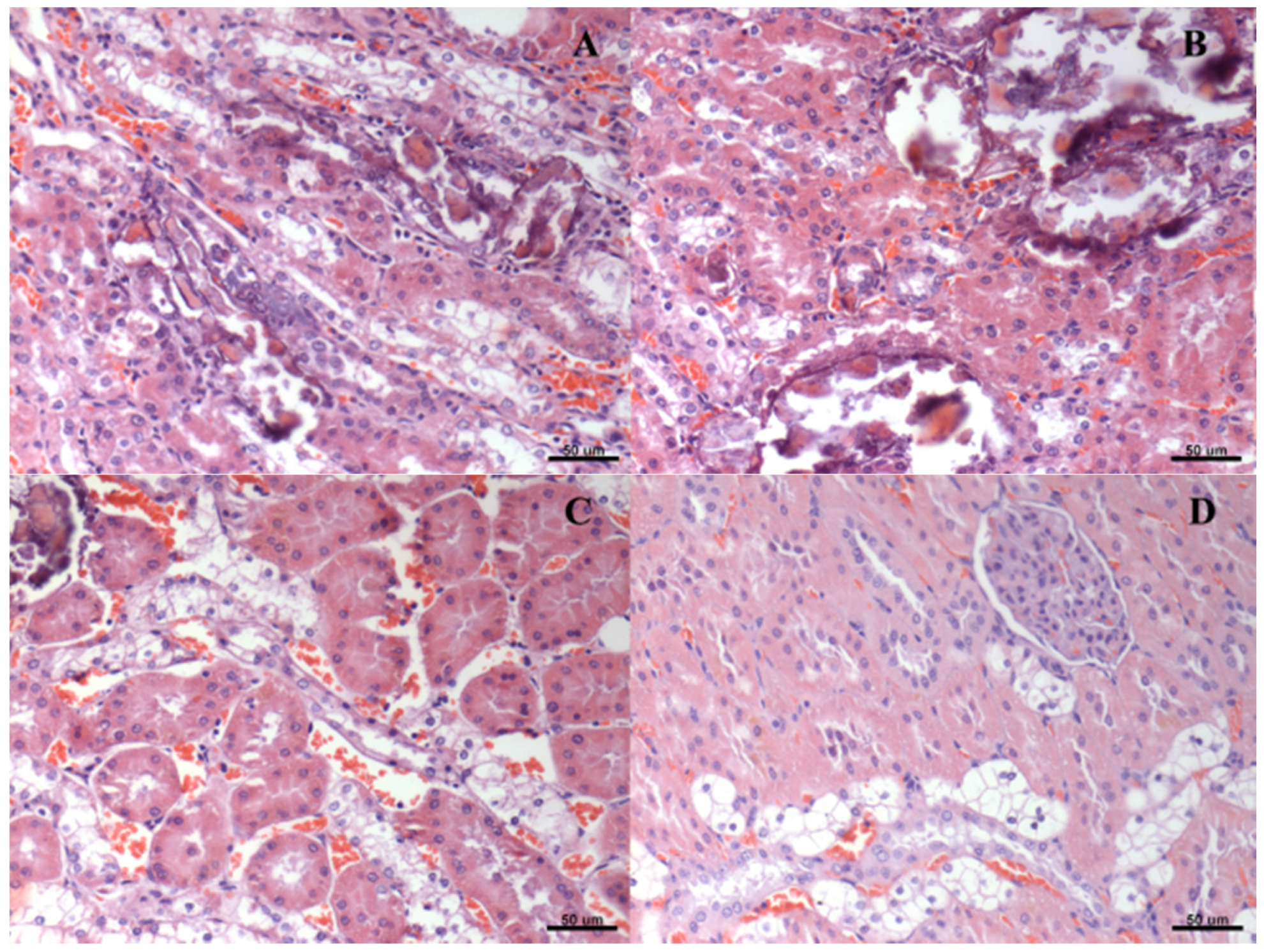
2.7. Histological Analysis
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus—Present and future perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet (Lond. Engl.) 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Atkinson, M.A.; Eisenbarth, G.S.; Michels, A.W. Type 1 diabetes. Lancet 2014, 383, 69–82. [Google Scholar] [CrossRef]
- American Diabetes Association. Standards of medical care in diabetes—2015 abridged for primary care providers. Clin. Diabetes 2015, 33, 97–111. [Google Scholar]
- Teixeira-Lemos, E.; Nunes, S.; Teixeira, F.; Reis, F. Regular physical exercise training assists in preventing type 2 diabetes development: Focus on its antioxidant and anti-inflammatory properties. Cardiovasc. Diabetol. 2011, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.W.; Khunti, K.; Harvey, R.; Johnson, M.; Preston, L.; Woods, H.B.; Davies, M.; Goyder, E. Preventing the progression to type 2 diabetes mellitus in adults at high risk: A systematic review and network meta-analysis of lifestyle, pharmacological and surgical interventions. Diabetes Res. Clin. Pract. 2015, 107, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Karpe, F.; Lafontan, M.; Frayn, K. Physical activity and exercise in the regulation of human adipose tissue physiology. Physiol. Rev. 2012, 92, 157–191. [Google Scholar] [CrossRef] [PubMed]
- Samuelsen, A.B.C.; Schrezenmeir, J.; Knutsen, S.H. Effects of orally administered yeast-derived beta-glucans: A review. Mol. Nutr. Food Res. 2014, 58, 183–193. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Silva, V.; Lobato, R.V.; Andrade, E.F.; de Macedo, C.G.; Napimoga, J.T.C.; Napimoga, M.H.; Messora, M.R.; Murata, R.M.; Pereira, L.J. β-Glucans (Saccharomyces cereviseae) Reduce Glucose Levels and Attenuate Alveolar Bone Loss in Diabetic Rats with Periodontal Disease. PLoS ONE 2015, 10, e0134742. [Google Scholar]
- Rahar, S.; Swami, G.; Nagpal, N.; Nagpal, M.A.; Singh, G.S. Preparation, characterization, and biological properties of β-glucans. J. Adv. Pharm. Technol. Res. 2011, 2, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Sonck, E.; Stuyven, E.; Goddeeris, B.; Cox, E. The effect of beta-glucans on porcine leukocytes. Vet. Immunol. Immunopathol. 2010, 135, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, M.S.; Bellini, M.F.; Angeli, J.P.F.; Oliveira, R.J.; Silva, A.F.; Ribeiro, L.R. Beta-Glucans in promoting health: Prevention against mutation and cancer. Mutat. Res. 2008, 658, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.F.; Lobato, R.V.; Araújo, T.V.; Zangerônimo, M.G.; Sousa, R.V.; Pereira, L.J. Effect of beta-glucans in the control of blood glucose levels of diabetic patients: A systematic review. Nutr. Hosp. 2014, 31, 170–177. [Google Scholar]
- Akramiene, D.; Kondrotas, A.; Didziapetriene, J.; Kevelaitis, E. Effects of beta-glucans on the immune system. Medicina (Kaunas) 2007, 43, 597–606. [Google Scholar] [PubMed]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary Yeast Beta-1,3/1,6-d-glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Lobato, R.V.; de Silva, V.O.; Andrade, E.F.; Orlando, D.R.; Zangeronimo, M.G.; de Sousa, R.V.; Pereira, L.J. Metabolic effects of β-glucans (Saccharomyces cerevisae) per os administration in rats with streptozotocin-induced diabetes. Nutr. Hosp. 2015, 32, 256–264. [Google Scholar]
- De Araújo, T.V.; Andrade, E.F.; Lobato, R.V.; Orlando, D.R.; Gomes, N.F.; de Sousa, R.V.; Zangeronimo, M.G.; Pereira, L.J. Effects of beta-glucans ingestion (Saccharomyces cerevisiae) on metabolism of rats receiving high-fat diet. J. Anim. Physiol. Anim. Nutr. (Berl) 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Duan, G.; Lu, Y.; Pang, S.; Huang, X.; Jiang, Q.; Dang, N. The effect of simvastatin on glucose homeostasis in streptozotocin induced type 2 diabetic rats. J. Diabetes Res. 2013, 2013, 274986. [Google Scholar] [CrossRef] [PubMed]
- Skovsø, S. Modeling type 2 diabetes in rats using high-fat diet and streptozotocin. J. Diabetes Investig. 2014, 5, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Rambo, L.M.; Ribeiro, L.R.; Oliveira, M.S.; Furian, A.F.; Lima, F.D.; Souza, M.A.; Silva, L.F.A.; Retamoso, L.T.; Corte, C.L.D.; Puntel, G.O.; et al. Additive anticonvulsant effects of creatine supplementation and physical exercise against pentylenetetrazol-induced seizures. Neurochem. Int. 2009, 55, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Gobatto, C.A.; de Mello, M.A.; Sibuya, C.Y.; de Azevedo, J.R.; dos Santos, L.A.; Kokubun, E. Maximal lactate steady state in rats submitted to swimming exercise. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 21–27. [Google Scholar] [CrossRef]
- Andrade, E.F.; Lobato, R.V.; de Araújo, T.V.; Orlando, D.R.; da Costa, D.V.; de Oliveira Silva, V.; Rogatto, G.P.; Zangeronimo, M.G.; Rosa, P.V.; Pereira, L.J. Adaptation to physical training in rats orally supplemented with glycerol. Can. J. Physiol. Pharmacol. 2015, 93, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Amr, A.R.; Abeer, E.E.-K. Hypolipideimic and Hypocholestermic Effect of Pine Nuts in Rats Fed High Fat, Cholesterol-Diet. World Appl. Sci. J. 2011, 15, 1667–1677. [Google Scholar]
- Martinez-flores, H.E.; Kil, Y. Effect of high fiber products on blood lipids and lipoproteins in hamsters. Nutr. Res. 2004, 24, 85–93. [Google Scholar] [CrossRef]
- Hemmati, M.; Zohoori, E.; Mehrpour, O.; Karamian, M.; Asghari, S.; Zarban, A.; Nasouti, R. Anti-atherogenic potential of jujube, saffron and barberry: Anti-diabetic and antioxidant actions. EXCLI J. 2015, 14, 908–915. [Google Scholar] [PubMed]
- Gleissner, C.A.; Galkina, E.; Nadler, J.L.; Ley, K. Mechanisms by which diabetes increases cardiovascular disease. Drug Discov. Today Dis. Mech. 2007, 4, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.O. Determination of the surface area of the white rat with its application to the expression of metabolic results. Am. J. Physiol. 1929, 89, 24–33. [Google Scholar]
- Vickers, S.P.; Cheetham, S.; Headland, K.; Dickinson, K.; Grempler, R.; Mayoux, E.; Mark, M.; Klein, T. Combination of the sodium-glucose cotransporter-2 inhibitor empagliflozin with orlistat or sibutramine further improves the body-weight reduction and glucose homeostasis of obese rats fed a cafeteria diet. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.F.; Lobato, R.V.; Araújo, T.V.; Orlando, D.R.; Gomes, N.F.; Alvarenga, R.R.; Rogatto, G.P.; Zangeronimo, M.G.; Pereira, L.J. Metabolic effects of glycerol supplementation and aerobic physical training on Wistar rats. Can. J. Physiol. Pharmacol. 2014, 92, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.F. Sisvar: A computer statistical analysis system. Ciênc. Agrotec. 2011, 35, 1039–1042. [Google Scholar]
- Hall, K.E.; McDonald, M.W.; Grisé, K.N.; Campos, O.A.; Noble, E.G.; Melling, C.W.J. The role of resistance and aerobic exercise training on insulin sensitivity measures in STZ-induced Type 1 diabetic rodents. Metabolism 2013, 62, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Ghiasi, R.; Ghadiri Soufi, F.; Somi, M.H.; Mohaddes, G.; Mirzaie Bavil, F.; Naderi, R.; Alipour, M.R. Swim Training Improves HOMA-IR in Type 2 Diabetes Induced by High Fat Diet and Low Dose of Streptozotocin in Male Rats. Adv. Pharm. Bull. 2015, 5, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Silveira, A.P.S.; Bentes, C.M.; Costa, P.B.; Simão, R.; Silva, F.C.; Silva, R.P.; Novaes, J.S. Acute effects of different intensities of resistance training on glycemic fluctuations in patients with type 1 diabetes mellitus. Res. Sports Med. 2014, 22, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Borghouts, L.B.; Keizer, H.A. Exercise and insulin sensitivity: A review. Int. J. Sports Med. 2000, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhao, J.; Huang, Y.; Li, Y. The difference between oats and beta-glucan extract intake in the management of HbA1c, fasting glucose and insulin sensitivity: A meta-analysis of randomized controlled trials. Food Funct. 2016, 7, 1413–1428. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Gügolz, E.; Würsch, P. Effects of breakfast cereals containing various amounts of beta-glucan fibers on plasma glucose and insulin responses in NIDDM subjects. Diabetes Care 1996, 19, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Kim, H.; Jung, M.H.; Hong, S.; Song, J. Consumption of barley beta-glucan ameliorates fatty liver and insulin resistance in mice fed a high-fat diet. Mol. Nutr. Food Res. 2010, 54, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Rideout, T.C.; Harding, S.V.; Jones, P.J.; Fan, M.Z. Guar gum and similar soluble fibers in the regulation of cholesterol metabolism: Current understandings and future research priorities. Vasc. Health Risk Manag. 2008, 4, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Raa, J. Immune modulation by non-digestible and non-absorbable beta-1,3/1,6-glucan. Microb. Ecol. Health Dis. 2015, 26, 27824. [Google Scholar] [CrossRef] [PubMed]
- Rebello, C.J.; Burton, J.; Heiman, M.; Greenway, F.L. Gastrointestinal microbiome modulator improves glucose tolerance in overweight and obese subjects: A randomized controlled pilot trial. J. Diabetes Complicat. 2015, 29, 1272–1276. [Google Scholar] [CrossRef] [PubMed]
- Reyna, N.Y.; Cano, C.; Bermúdez, V.J.; Medina, M.T.; Souki, A.J.; Ambard, M.; Nuñez, M.; Ferrer, M.A.; Inglett, G.E. Sweeteners and beta-glucans improve metabolic and anthropometrics variables in well controlled type 2 diabetic patients. Am. J. Ther. 2003, 10, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Battilana, P.; Ornstein, K.; Minehira, K.; Schwarz, J.M.; Acheson, K.; Schneiter, P.; Burri, J.; Jéquier, E.; Tappy, L. Mechanisms of action of beta-glucan in postprandial glucose metabolism in healthy men. Eur. J. Clin. Nutr. 2001, 55, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Lumaga, R.B.; Stanzione, A.; Scalfi, L.; Fogliano, V. β-Glucan-enriched bread reduces energy intake and modifies plasma ghrelin and peptide YY concentrations in the short term. Appetite 2009, 53, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Moura, L.P.; Puga, G.M.; Beck, W.R.; Teixeira, I.P.; Ghezzi, A.C.; Silva, G.A.; Mello, M.A.R. Exercise and spirulina control non-alcoholic hepatic steatosis and lipid profile in diabetic Wistar rats. Lipids Health Dis. 2011, 10, 2–7. [Google Scholar] [CrossRef] [PubMed]
- De Lemos, E.T.; Pinto, R.; Oliveira, J.; Garrido, P.; Sereno, J.; Mascarenhas-Melo, F.; Páscoa-Pinheiro, J.; Teixeira, F.; Reis, F. Differential effects of acute (extenuating) and chronic (training) exercise on inflammation and oxidative stress status in an animal model of type 2 diabetes mellitus. Mediat. Inflamm. 2011, 2011, 253061. [Google Scholar]
- Bays, H.E.; Evans, J.L.; Maki, K.C.; Evans, M.; Maquet, V.; Cooper, R.; Anderson, J.W. Chitin-glucan fiber effects on oxidized low-density lipoprotein: A randomized controlled trial. Eur. J. Clin. Nutr. 2013, 67, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Chen, A.K.; Barnard, R.J. Effect of a short-term diet and exercise intervention in youth on atherosclerotic risk factors. Atherosclerosis 2007, 191, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Alsaad, K.O.; Herzenberg, A.M. Distinguishing diabetic nephropathy from other causes of glomerulosclerosis: An update. J. Clin. Pathol. 2007, 60, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Birkenfeld, A.L.; Shulman, G.I. Nonalcoholic fatty liver disease, hepatic insulin resistance, and type 2 Diabetes. Hepatology 2014, 59, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.-S.; Couturier, K.; Latour, J.-G.; Lavoie, J.-M. Concurrent exercise prevents high-fat-diet-induced macrovesicular hepatic steatosis. J. Appl. Physiol. 2003, 94, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Khazaei, M.; Moien-Afshari, F.; Ang, L.S.; Granville, D.J.; Verchere, C.B.; Dunn, S.R.; McCue, P.; Mizisin, A.; Sharma, K.; et al. Moderate exercise attenuates caspase-3 activity, oxidative stress, and inhibits progression of diabetic renal disease in db/db mice. Am. J. Physiol. Renal Physiol. 2009, 296, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kume, S.; Araki, S.; Isshiki, K.; Chin-Kanasaki, M.; Sakaguchi, M.; Sugimoto, T.; Koya, D.; Haneda, M.; Kashiwagi, A.; et al. Fenofibrate, a PPARα agonist, has renoprotective effects in mice by enhancing renal lipolysis. Kidney Int. 2011, 79, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Boor, P.; Celec, P.; Behuliak, M.; Grančič, P.; Kebis, A.; Kukan, M.; Pronayová, N.; Liptaj, T.; Ostendorf, T.; Šebeková, K. Regular moderate exercise reduces advanced glycation and ameliorates early diabetic nephropathy in obese Zucker rats. Metabolism 2009, 58, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.-I.; Matsui, T. Advanced glycation end products, oxidative stress and diabetic nephropathy. Oxid. Med. Cell. Longev. 2010, 3, 101–108. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Beta-Glucan | Physical Training | ||
|---|---|---|---|
| Without | With | ||
| Glucose (mg/dL) | Without | 371.0 (±21.4) A,a | 335.0 (±10.4) b |
| With | 311.0 (±25.0) b | 327.5 (±42.6) | |
| HbA1c (mg/dL) | Without | 9.4 (±0.4) A,a | 8.8 (±0.3) B |
| With | 8.33 (±0.1) b | 8.8 (±0.4) | |
| Triacylglycerols (mg/dL) | Without | 105.8 (±11.1) a | 99.7 (±2.2) a |
| With | 71.5 (±7.2) b | 57.5 (±12.8) b | |
| Total cholesterol (mg/dL) | Without | 88.8 (±22.9) a | 85.4 (±9.4) a |
| With | 65.1 (±3.3) b | 63.8 (±7.2) b | |
| HDL-C (mg/dL) | Without | 34.33 (±3.8) a | 37.7 (±6.9) |
| With | 42.66 (±5.2) b | 44.26 (±4.0) | |
| LDL-C (mg/dL) | Without | 34.6 (±20.1) a | 33.4 (±5.8) a |
| With | 10.95 (±3.4) b | 19.6 (±4.9) b | |
| Atherogenic index of plasma | Without | 1.6 (±0.6) a | 1.3 (±0.2) a |
| With | 0.6 (±0.1) A,b | 0.4 (±0.1) B,b | |
| Group | Score of Steatosis | ||||
|---|---|---|---|---|---|
| * | ** | *** | **** | ***** | |
| A | 0 | 3 | 0 | 3 | 0 |
| B | 1 | 3 | 2 | 0 | 0 |
| C | 2 | 3 | 0 | 1 | 0 |
| D | 1 | 5 | 0 | 0 | 0 |
| Score of Renal Degeneration | |||||
|---|---|---|---|---|---|
| Group | * | ** | *** | **** | ***** |
| A | 0 | 0 | 0 | 2 | 4 |
| B | 0 | 0 | 0 | 6 | 0 |
| C | 0 | 0 | 1 | 5 | 0 |
| D # | 0 | 0 | 1 | 5 | 0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, E.F.; Lima, A.R.V.; Nunes, I.E.; Orlando, D.R.; Gondim, P.N.; Zangeronimo, M.G.; Alves, F.H.F.; Pereira, L.J. Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ). Nutrients 2016, 8, 792. https://doi.org/10.3390/nu8120792
Andrade EF, Lima ARV, Nunes IE, Orlando DR, Gondim PN, Zangeronimo MG, Alves FHF, Pereira LJ. Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ). Nutrients. 2016; 8(12):792. https://doi.org/10.3390/nu8120792
Chicago/Turabian StyleAndrade, Eric Francelino, Andressa Ribeiro Veiga Lima, Ingrid Edwiges Nunes, Débora Ribeiro Orlando, Paula Novato Gondim, Márcio Gilberto Zangeronimo, Fernando Henrique Ferrari Alves, and Luciano José Pereira. 2016. "Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ)" Nutrients 8, no. 12: 792. https://doi.org/10.3390/nu8120792
APA StyleAndrade, E. F., Lima, A. R. V., Nunes, I. E., Orlando, D. R., Gondim, P. N., Zangeronimo, M. G., Alves, F. H. F., & Pereira, L. J. (2016). Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ). Nutrients, 8(12), 792. https://doi.org/10.3390/nu8120792