Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Anthropometric Characteristics
2.2. Beverage Composition
2.3. Experimental Procedure
2.4. Fatty Acid Determination
2.5. Polyphenols Determination
2.6. Cytokine, Eicosanoids and Adhesion Molecules Determination
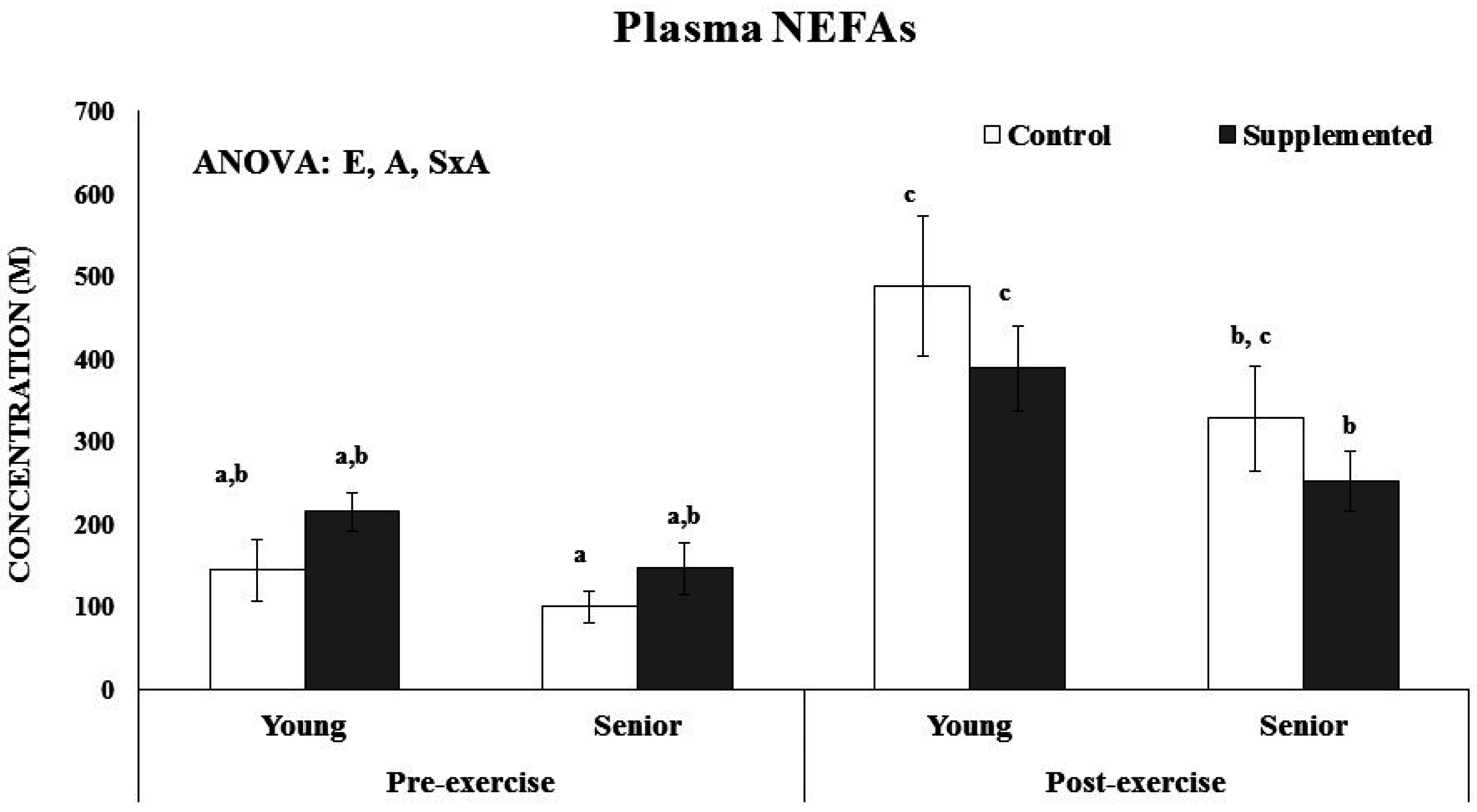
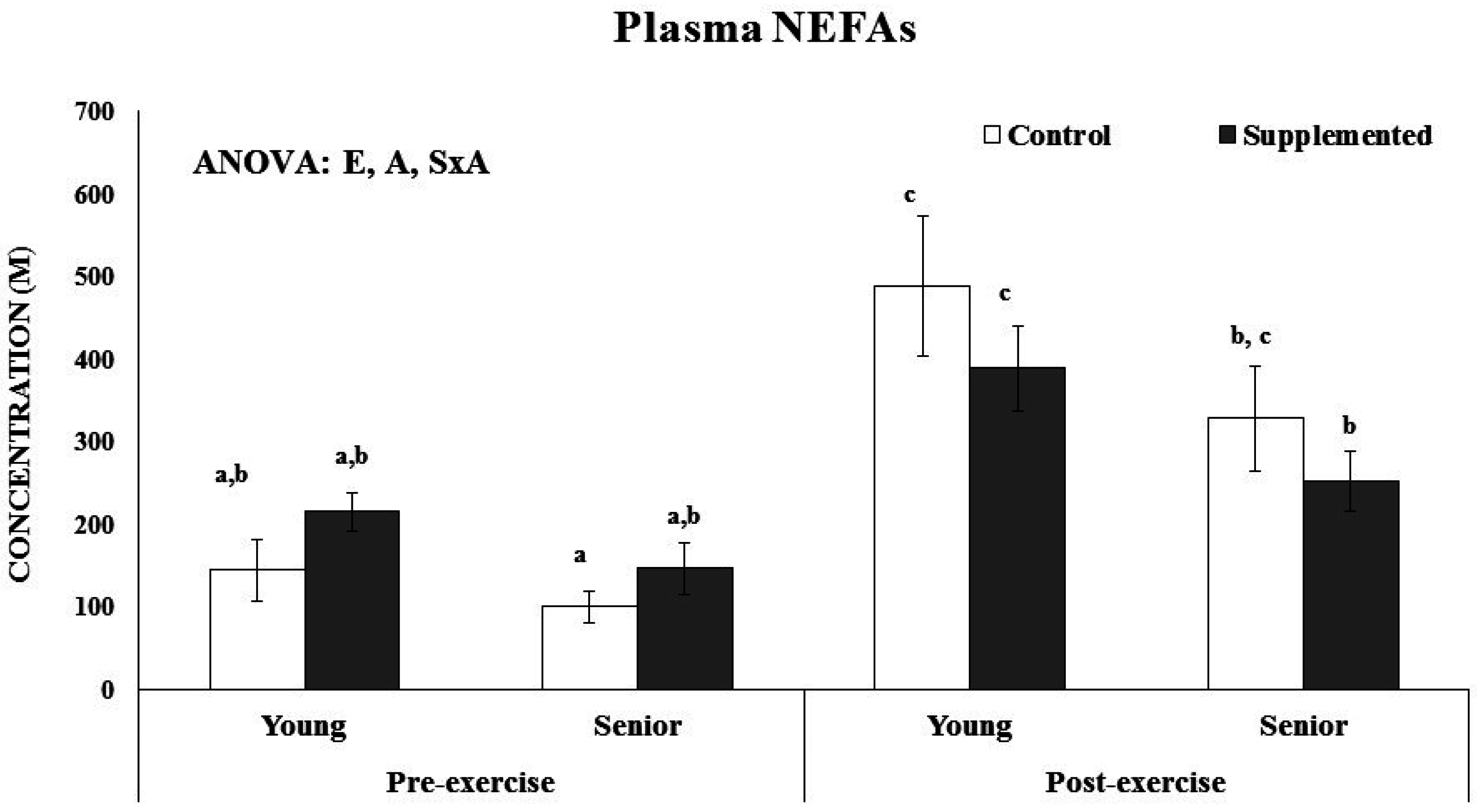
2.7. Non Esterified Fatty Acids Determination
2.8. Nuclear Factor κβ (NFκβ) Activation Quantification
2.9. Gene Expression
2.10. Statistical Analysis
3. Results
3.1. Effects on Exercise Performance Parameters
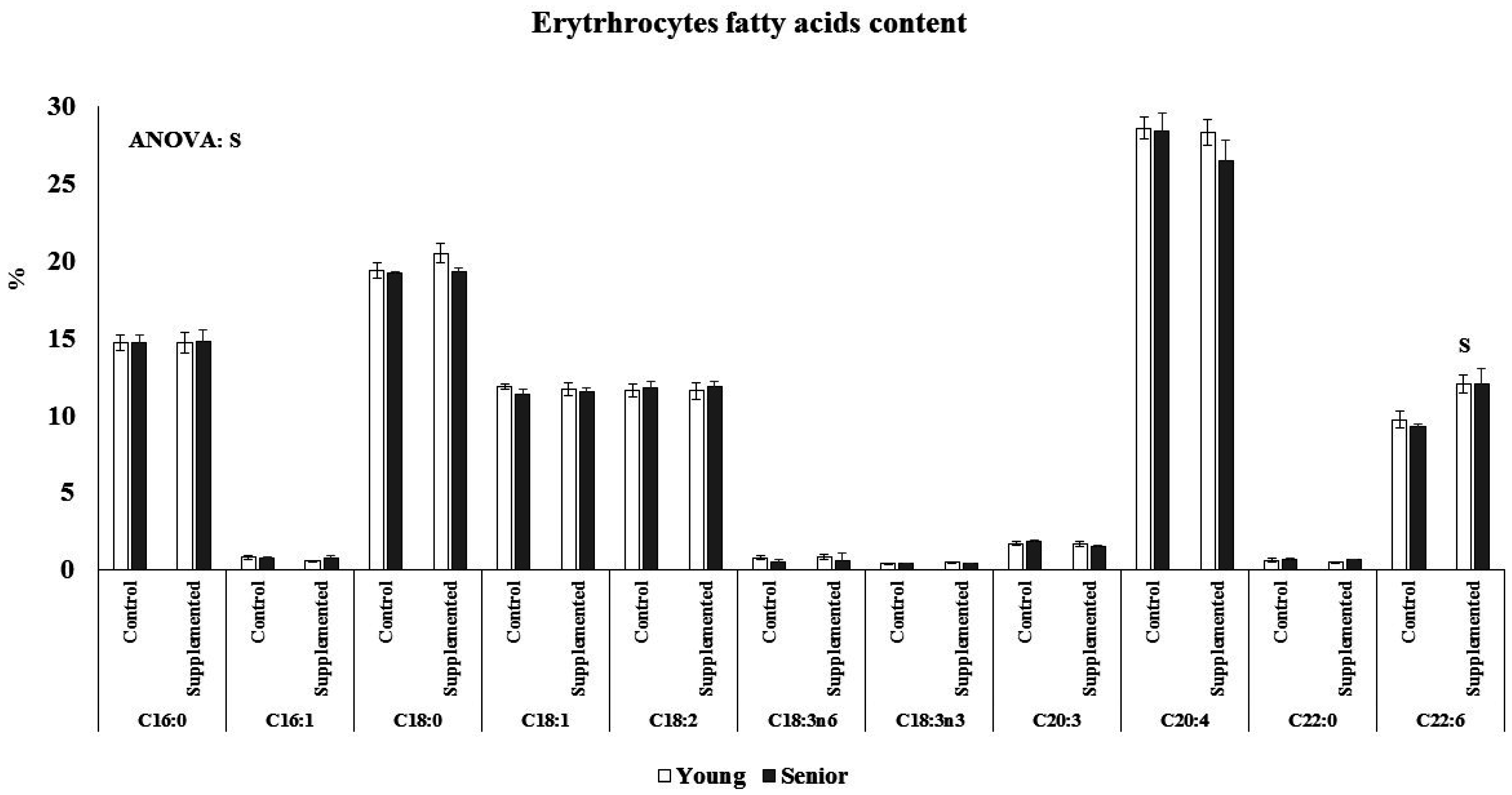
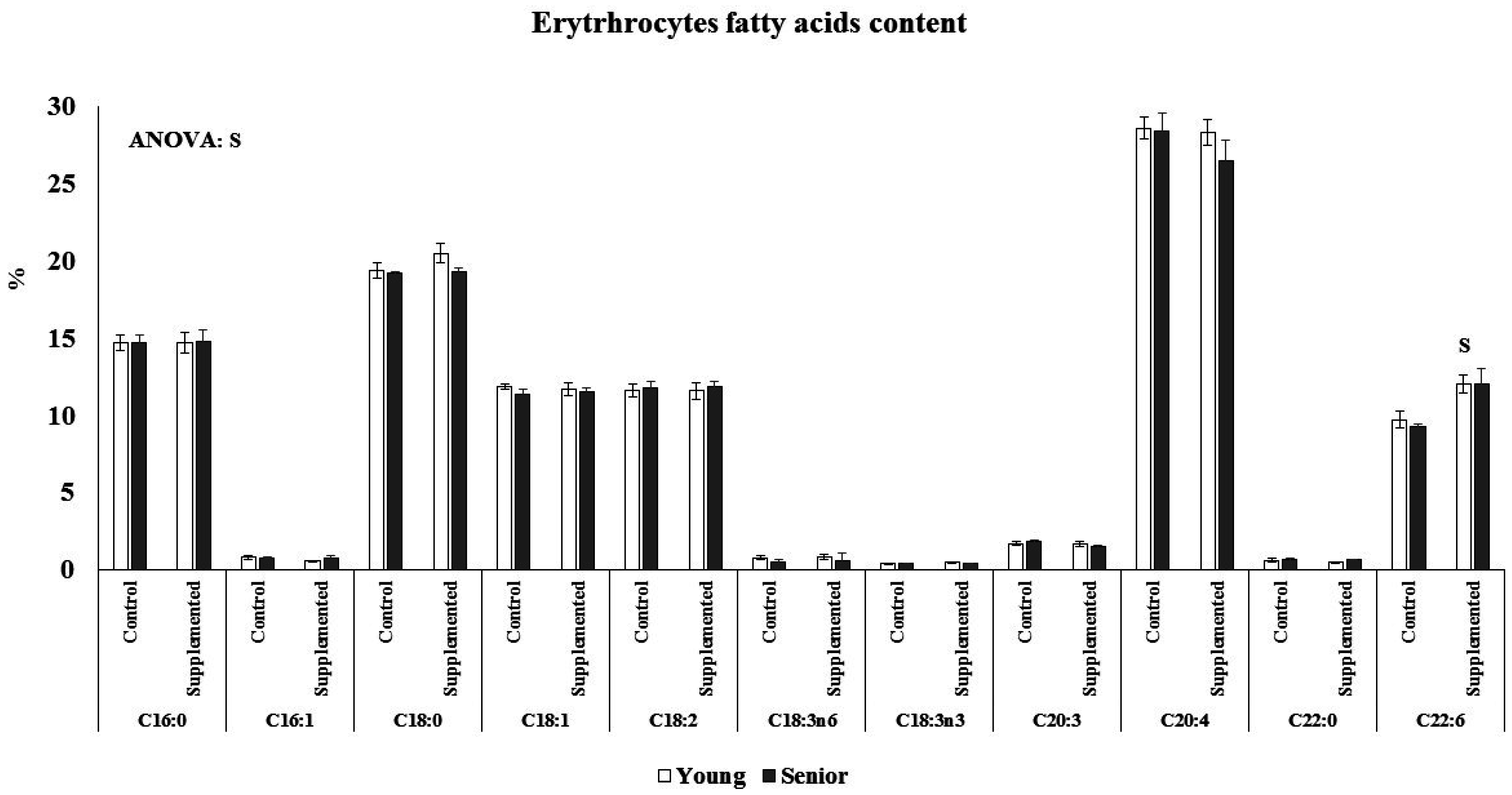
3.2. Effects on Fatty Acids Composition
3.3. Effects on Inflammatory, Heat Stress and Immune Priming Response Markers
3.4. Effects on Inflammatory Genes Expression in PBMCs
4. Discussion
5. Effects on Inflammatory Markers
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Milbury, P.E.; Chen, C.Y.; Dolnikowski, G.G.; Blumberg, J.B. Determination of flavonoids and phenolics and their distribution in almonds. J. Agric. Food Chem. 2006, 54, 5027–5033. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jia, X.; Chen, C.Y.; Blumberg, J.B.; Song, Y.; Zhang, W.; Zhang, X.; Ma, G.; Chen, J. Almond consumption reduces oxidative DNA damage and lipid peroxidation in male smokers. J. Nutr. 2007, 137, 2717–2722. [Google Scholar] [PubMed]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134 (Suppl. 12), 3479s–3485s. [Google Scholar] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Marchie, A.; Josse, A.R.; Nguyen, T.H.; Faulkner, D.A.; Lapsley, K.G.; Blumberg, J. Almonds reduce biomarkers of lipid peroxidation in older hyperlipidemic subjects. J. Nutr. 2008, 138, 908–913. [Google Scholar] [PubMed]
- Tresserra-Rimbau, A.; Medina-Remon, A.; Perez-Jimenez, J.; Martinez-Gonzalez, M.A.; Covas, M.I.; Corella, D.; Salas-Salvado, J.; Gomez-Gracia, E.; Lapetra, J.; Aros, F.; et al. Dietary intake and major food sources of polyphenols in a Spanish population at high cardiovascular risk: The predimed study. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferre, M.; Hu, F.B.; Martinez-Gonzalez, M.A.; Fito, M.; Bullo, M.; Estruch, R.; Ros, E.; Corella, D.; Recondo, J.; Gomez-Gracia, E.; et al. Olive oil intake and risk of cardiovascular disease and mortality in the predimed study. BMC Med. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Casas, R.; Chiva-Blanch, G.; Romero-Mamani, E.S.; Valderas-Martinez, P.; Arranz, S.; Andres-Lacueva, C.; Llorach, R.; Medina-Remon, A.; Lamuela-Raventos, R.M.; et al. Virgin olive oil and nuts as key foods of the mediterranean diet effects on inflammatory biomakers related to atherosclerosis. Pharmacol. Res. 2012, 65, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, M.A.; Toledo, E.; Aros, F.; Fiol, M.; Corella, D.; Salas-Salvado, J.; Ros, E.; Covas, M.I.; Fernandez-Crehuet, J.; Lapetra, J.; et al. Extravirgin olive oil consumption reduces risk of atrial fibrillation: The PREDIMED (prevencion con dieta mediterranea) trial. Circulation 2014, 130, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Medina-Remon, A.; Tresserra-Rimbau, A.; Pons, A.; Tur, J.A.; Martorell, M.; Ros, E.; Buil-Cosiales, P.; Sacanella, E.; Covas, M.I.; Corella, D.; et al. Effects of total dietary polyphenols on plasma nitric oxide and blood pressure in a high cardiovascular risk cohort. The predimed randomized trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Petroczi, A.; Naughton, D.P.; Mazanov, J.; Holloway, A.; Bingham, J. Performance enhancement with supplements: Incongruence between rationale and practice. J. Int. Soc. Sports Nutr. 2007, 4. [Google Scholar] [CrossRef] [PubMed]
- Mickleborough, T.D. Omega-3 polyunsaturated fatty acids in physical performance optimization. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Martorell, M.; Capo, X.; Sureda, A.; Batle, J.M.; Llompart, I.; Argelich, E.; Tur, J.A.; Pons, A. Effect of DHA on plasma fatty acid availability and oxidative stress during training season and football exercise. Food Funct. 2014, 5, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Capo, X.; Martorell, M.; Llompart, I.; Sureda, A.; Tur, J.A.; Pons, A. Docosahexanoic acid diet supplementation attenuates the peripheral mononuclear cell inflammatory response to exercise following LPS activation. Cytokine 2014, 69, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Capo, X.; Martorell, M.; Sureda, A.; Batle, J.M.; Tur, J.A.; Pons, A. Docosahexaenoic diet supplementation, exercise and temperature affect cytokine production by lipopolysaccharide-stimulated mononuclear cells. J. Physiol. Biochem. 2016, 72, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.J.; Bote, E.; Hinchado, M.D.; Ortega, E. A single session of intense exercise improves the inflammatory response in healthy sedentary women. J. Physiol. Biochem. 2011, 67, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Rohde, T.; Zacho, M. Immunity in athletes. J. Sports Med. Phys. Fit. 1996, 36, 236–245. [Google Scholar]
- Nieman, D.C.; Pedersen, B.K. Exercise and immune function. Recent developments. Sports Med. 1999, 27, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Ozen, A.E.; Pons, A.; Tur, J.A. Worldwide consumption of functional foods: A systematic review. Nutr. Rev. 2012, 70, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Tauler, P.; Aguilo, A.; Cases, N.; Fuentespina, E.; Cordova, A.; Tur, J.A.; Pons, A. Relation between oxidative stress markers and antioxidant endogenous defences during exhaustive exercise. Free Radic. Res. 2005, 39, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Mestre-Alfaro, A.; Ferrer, M.D.; Sureda, A.; Tauler, P.; Martinez, E.; Bibiloni, M.M.; Micol, V.; Tur, J.A.; Pons, A. Phytoestrogens enhance antioxidant enzymes after swimming exercise and modulate sex hormone plasma levels in female swimmers. Eur. J. Appl. Physiol. 2011, 111, 2281–2294. [Google Scholar] [CrossRef] [PubMed]
- Watson, T.A.; MacDonald-Wicks, L.K.; Garg, M.L. Oxidative stress and antioxidants in athletes undertaking regular exercise training. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 131–146. [Google Scholar] [PubMed]
- Volpe, S.L. Micronutrient requirements for athletes. Clin. Sports Med. 2007, 26, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an endocrine organ: Focus on muscle-derived Interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.C.; Fleshner, M.; Green, C.; Pedersen, B.K.; Hoffman-Goetz, L.; et al. Position statement part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar] [PubMed]
- Pedersen, B.K.; Saltin, B. Evidence for prescribing exercise as therapy in chronic disease. Scand. J. Med. Sci. Sports 2006, 16 (Suppl. 1), 3–63. [Google Scholar] [CrossRef] [PubMed]
- Calbet, J.A. Ageing, exercise and cardiovascular health: Good and bad news. J. Physiol. 2012, 590, 5265–5266. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Goldeck, D.; Derhovanessian, E. Inflammation, ageing and chronic disease. Curr. Opin. Immunol. 2014, 29, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.M.; McEneny, J.; Mathieu-Costello, O.; Henry, R.R.; James, P.E.; McCord, J.M.; Pietri, S.; Young, I.S.; Richardson, R.S. Sedentary aging increases resting and exercise-induced intramuscular free radical formation. J. Appl. Physiol. 2010, 109, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M. Vitamin E requirement in relation to dietary fish oil and oxidative stress in elderly. Exs 1992, 62, 411–418. [Google Scholar] [PubMed]
- Molfino, A.; Gioia, G.; Rossi Fanelli, F.; Muscaritoli, M. The role for dietary omega-3 fatty acids supplementation in older adults. Nutrients 2014, 6, 4058–4073. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Mitchell, J.B. Exercise in hot and cold environments: Differential effects on leukocyte number and NK cell activity. Aviat. Space Environ. Med. 2003, 74, 1231–1236. [Google Scholar] [PubMed]
- Borg, G. Perceived exertion as an indicator of somatic stress. Scand. J. Rehabil. Med. 1970, 2, 92–98. [Google Scholar] [PubMed]
- Capo, X.; Martorell, M.; Sureda, A.; Llompart, I.; Tur, J.A.; Pons, A. Diet supplementation with DHA-enriched food in football players during training season enhances the mitochondrial antioxidant capabilities in blood mononuclear cells. Eur. J. Nutr. 2015, 54, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Martorell, M.; Capo, X.; Bibiloni, M.M.; Sureda, A.; Mestre-Alfaro, A.; Batle, J.M.; Llompart, I.; Tur, J.A.; Pons, A. Docosahexaenoic acid supplementation promotes erythrocyte antioxidant defense and reduces protein nitrosative damage in male athletes. Lipids 2015, 50, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Kubiliene, L.; Laugaliene, V.; Pavilonis, A.; Maruska, A.; Majiene, D.; Barcauskaite, K.; Kubilius, R.; Kasparaviciene, G.; Savickas, A. Alternative preparation of propolis extracts: Comparison of their composition and biological activities. BMC Complement. Altern. Med. 2015, 15, 156. [Google Scholar] [CrossRef] [PubMed]
- Tepsic, J.; Vucic, V.; Arsic, A.; Blazencic-Mladenovic, V.; Mazic, S.; Glibetic, M. Plasma and erythrocyte phospholipid fatty acid profile in professional basketball and football players. Eur. J. Appl. Physiol. 2009, 107, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, N.R.; DiMarco, N.M.; Langley, S. Position of the American dietetic association, dietitians of Canada, and the American college of sports medicine: Nutrition and athletic performance. J. Am. Diet. Assoc. 2009, 109, 509–527. [Google Scholar] [PubMed]
- Neubauer, O.; Yfanti, C. Antioxidants in athlete’s basic nutrition: Considerations towards a guideline for the intake of vitamin C and vitamine. In Antioxidants in Sport Nutrition; Lamprecht, M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2015. [Google Scholar]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Martin, M.V.; Soto, M.J.; Rico, M.C.; Vallejo, F.; Tomas-Barberan, F.; Perez-de-la-Cruz, A.J.; Gil, A.; Mesa, M.D. Normal or high polyphenol concentration in orange juice affects antioxidant activity, blood pressure, and body weight in obese or overweight adults. J. Nutr. 2015, 145, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Bentley, D.J.; Ackerman, J.; Clifford, T.; Slattery, K.S. Acute and chronic effects of antioxidant supplementation on exercise performance. In Antioxidants in Sport Nutrition; Lamprecht, M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2015. [Google Scholar]
- Fleming, J.; Sharman, M.J.; Avery, N.G.; Love, D.M.; Gomez, A.L.; Scheett, T.P.; Kraemer, W.J.; Volek, J.S. Endurance capacity and high-intensity exercise performance responses to a high fat diet. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Rodacki, C.L.; Rodacki, A.L.; Pereira, G.; Naliwaiko, K.; Coelho, I.; Pequito, D.; Fernandes, L.C. Fish-oil supplementation enhances the effects of strength training in elderly women. Am. J. Clin. Nutr. 2012, 95, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Lembke, P.; Capodice, J.; Hebert, K.; Swenson, T. Influence of omega-3 (n-3) index on performance and wellbeing in young adults after heavy eccentric exercise. J. Sports Sci. Med. 2014, 13, 151–156. [Google Scholar] [PubMed]
- Sureda, A.; Mestre-Alfaro, A.; Banquells, M.; Riera, J.; Drobnic, F.; Camps, J.; Joven, J.; Tur, J.A.; Pons, A. Exercise in a hot environment influences plasma anti-inflammatory and antioxidant status in well-trained athletes. J. Therm. Biol. 2015, 47, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Devaraj, S.; Jialal, I. Dietary factors that promote or retard inflammation. Arterioscler. Thromb. Vasc. Boil. 2006, 26, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; Connell, K.M.; Sabate, J. Effect of almond-enriched high-monounsaturated fat diet on selected markers of inflammation: A randomised, controlled, crossover study. Br. J. Nutr. 2010, 103, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [PubMed]
- Peake, J.M.; Suzuki, K.; Hordern, M.; Wilson, G.; Nosaka, K.; Coombes, J.S. Plasma cytokine changes in relation to exercise intensity and muscle damage. Eur. J. Appl. Physiol. 2005, 95, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Pararasa, C.; Ikwuobe, J.; Shigdar, S.; Boukouvalas, A.; Nabney, I.T.; Brown, J.E.; Devitt, A.; Bailey, C.J.; Bennett, S.J.; Griffiths, H.R. Age-associated changes in long-chain fatty acid profile during healthy aging promote pro-inflammatory monocyte polarization via ppargamma. Aging Cell 2016, 15, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Pahl, H.L. Activators and target genes of Rel/NF-kappab transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [PubMed]
- Capo, X.; Martorell, M.; Sureda, A.; Tur, J.A.; Pons, A. Effects of docosahexaenoic supplementation and in vitro vitamin C on the oxidative and inflammatory neutrophil response to activation. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Markworth, J.F.; Vella, L.; Lingard, B.S.; Tull, D.L.; Rupasinghe, T.W.; Sinclair, A.J.; Maddipati, K.R.; Cameron-Smith, D. Human inflammatory and resolving lipid mediator responses to resistance exercise and ibuprofen treatment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1281–R1296. [Google Scholar] [CrossRef] [PubMed]
- Capo, X.; Martorell, M.; Sureda, A.; Tur, J.A.; Pons, A. Effects of dietary docosahexaenoic, training and acute exercise on lipid mediators. J. Int. Soc. Sports Nutr. 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Hirose, L.; Nosaka, K.; Newton, M.; Laveder, A.; Kano, M.; Peake, J.; Suzuki, K. Changes in inflammatory mediators following eccentric exercise of the elbow flexors. Exerc. Immunol. Rev. 2004, 10, 75–90. [Google Scholar] [PubMed]
- Scheller, J.; Rose-John, S. Interleukin-6 and its receptor: From bench to bedside. Med. Microbial. Immunol. 2006, 195, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liu, J.; Pang, X.; Wang, S.; Zhao, J.; Zhang, X.; Feng, L. Palmitic acid exerts pro-inflammatory effects on vascular smooth muscle cells by inducing the expression of c-reactive protein, inducible nitric oxide synthase and tumor necrosis factor-alpha. Int. J. Mol. Med. 2014, 34, 1706–1712. [Google Scholar] [PubMed]
- Calder, P.C. N-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83 (Suppl. 6), 1505s–1519s. [Google Scholar] [PubMed]
- Petridou, A.; Chatzinikolaou, A.; Fatouros, I.; Mastorakos, G.; Mitrakou, A.; Chandrinou, H.; Papassotiriou, I.; Mougios, V. Resistance exercise does not affect the serum concentrations of cell adhesion molecules. Br. J. Sports Med. 2007, 41, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.; Liesen, H.; Enneper, J. Leucocytes, lymphocytes, activation parameters and cell adhesion molecules in middle-distance runners under different training conditions. Int. J. Sports Med. 1994, 15 (Suppl. 3), S122–S126. [Google Scholar] [CrossRef] [PubMed]
- Carlos, T.M.; Harlan, J.M. Leukocyte-endothelial adhesion molecules. Blood 1994, 84, 2068–2101. [Google Scholar] [PubMed]
- Akimoto, T.; Furudate, M.; Saitoh, M.; Sugiura, K.; Waku, T.; Akama, T.; Kono, I. Increased plasma concentrations of intercellular adhesion molecule-1 after strenuous exercise associated with muscle damage. Eur. J. Appl. Physiol. 2002, 86, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, M.A.; Pulido, R.; Munoz, C.; Alvarez, V.; Humbria, A.; Campanero, M.R.; Sanchez-Madrid, F. Regulation of ICAM-3 (CD50) membrane expression on human neutrophils through a proteolytic shedding mechanism. Eur. J. Immunol. 1994, 24, 2586–2594. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.L.; Anwar, A.; Fragen, M.; Rananto, C.; Johnson, R.; Holbert, D. Cytokines and cell adhesion molecules associated with high-intensity eccentric exercise. Eur. J. Appl. Physiol. 2000, 82, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.G.; Lyberg, T. Long-distance running modulates the expression of leucocyte and endothelial adhesion molecules. Scand. J. Immunol. 2004, 60, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.J.; Hong, S.; Redwine, L.; Carter, S.M.; Chiu, A.; Ziegler, M.G.; Dimsdale, J.E.; Maisel, A.S. Physical fitness attenuates leukocyte-endothelial adhesion in response to acute exercise. J. Appl. Physiol. 2006, 101, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Munoz, A.; Costa, M. Nutritionally mediated oxidative stress and inflammation. Oxid. Med. Cell. Longev. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Young | Senior | |
|---|---|---|
| Age (years) | 22.8 ± 3.8 $ | 45.6 ± 1.6 |
| Weight (kg) | 71.0 ± 4.8 | 76.1 ± 2.9 |
| Height (cm) | 176 ± 3.8 | 177 ± 3.8 |
| Fat-free mass (%) | 9.50 ± 1.1 | 11.6 ± 1.0 |
| Body surface (m2) | 1.86 ± 0.05 | 1.93 ± 0.07 |
| VO2 max (L/min*kg) | 58.8 ± 2.5 | 50.4 ± 3.4 |
| Fatty Acid | Composition |
|---|---|
| C16:0 (%) | 7.62 ± 0.97 |
| C16:1 (%) | 1.20 ± 0.14 |
| C18:0 (%) | 4.24 ± 0.7 |
| C18:1 (%) | 42.5 ± 4.8 |
| C18:2 (%) | 22.9 ± 2.5 |
| C18:3n6 (%) | 0.949 ± 0.55 |
| C18:3n3 (%) | 1.21 ± 0.35 |
| C20:0 (%) | 0.368 ± 0.105 |
| C20:1(%) | 0.113 ± 0.032 |
| C20:2 (%) | 0.884 ± 0.252 |
| C20:3 (%) | 0.065 ± 0.019 |
| C20:4n6 (%) | 0.434 ± 0.124 |
| C22:0 (%) | 0.262 ± 0.075 |
| C22:5 (%) | 5.68 ± 1.62 |
| C22:6n3 (%) | 11.6 ± 3.3 |
| Total Fatty Acids (µM) | 47400 ± 9586 |
| SFA (%) | 9.90 ± 1.15 |
| MUFA (%) | 51.7 ± 5.0 |
| PUFA (%) | 38.3 ± 4.4 |
| VITAMIN E (mg/L) | 45.7 ± 27.7 |
| POLYPHENOLS (mM) | 2.85 ± 0.29 |
| Gene | Primer | Conditions | ||
|---|---|---|---|---|
| 18S | Fw: | 5′-ATG TGA AGT CAC TGT GCC AG-3′ | 95 °C | 10 s |
| Rv: | 5′-GTG TAA TCC GTC TCC ACA GA-3′ | 60 °C | 10 s | |
| 72 °C | 12 s | |||
| TLR2 | Fw: | 5′-GGGTTGAAGCACTGGACAAT-3′ | 95 °C | 10 s |
| Rv: | 5′-TTCTTCCTTGGAGAGGCTGA-3′ | 60 °C | 10 s | |
| 72 °C | 15 s | |||
| TLR4 | Fw: | 5′-GGTCACCTTTTCTTGATTCCA-3′ | 95 °C | 10 s |
| Rv: | 5′-TCAGAGGTCCATCAAACATCAC-3′ | 60 °C | 10 s | |
| 72 °C | 15 s | |||
| TNFα | Fw: | 5′-CCCAGGCAGTCAGATCATCTTCTCGGAA-3′ | 94 °C | 10 s |
| Rv: | 5′-CTGGTTATCTCTCAGCTCCACGCCATT-3′ | 63 °C | 10 s | |
| 72 °C | 15 s | |||
| NFκβ | Fw: | 5′-AAA CAC TGT GAG GAT GGG ATC TG-3′ | 95 °C | 10 s |
| Rv: | 5′-CGA AGC CGA CCA CCA TGT-3′ | 60 °C | 10 s | |
| 72 °C | 15 s | |||
| COX2 | Fw: | 5-TTG CTG GCA GGG TTG CTG GTG GTA-3′ | 95 °C | 10 s |
| Rv: | 5′-CAT CTG CCT GCT CTG GTC AAT GGA A-3′ | 67 °C | 10 s | |
| 72 °C | 15 s | |||
| 15 LOX 2 | Fw: | 5′-GCA TCC ACT GAT TGG ACC TT-3′ | 95 °C | 10 s |
| Rv: | 5′-GCT GGC CTT GAA CTT CTG AC-3′ | 61 °C | 10 s | |
| 72 °C | 15 s | |||
| IL1β | Fw: | 5′-GGA CAG GAT ATG GAG CAA CA-3′ | 95 °C | 10 s |
| Rv: | 5′-GGC AGA CTC AAA TTC CAG CT-3′ | 58 °C | 10 s | |
| 72 °C | 15 s | |||
| 5 LOX | Fw: | 5′-GGG CAT GGA GAG CAA AGA AG-3 | 95 °C | 10 s |
| Rv: | 5′-ACC TCG GCC GTG AAC GT-3′ | 59 °C | 10 s | |
| 72 °C | 15 s | |||
| IL-8 | Fw: | 5′-GCTCTGTGTGAAGGTGCAGTTTTGCCAA-3′ | 95 °C | 10 s |
| Rv: | 5′-GGCGCAGTGTGGTCCACTCTCAAT-3′ | 63 °C | 10 s | |
| 72 °C | 15 s | |||
| IL-10 | Fw: | 5′-AGAACCTGAAGACCCTCAGGC-3′ | 95 °C | 10 s |
| Rv: | 5′-CCACGGCCTTGCTCTTGTT-3′ | 60 °C | 10 s | |
| 72 °C | 15 s | |||
| IL-15 | Fw: | 5′-CCGTGGCTTTGAGTAATGAGAATTTCGAA-3′ | 95 °C | 10 s |
| Rv: | 5′-CCTGCACTGAAACAGCCCAAAATGAA-3′ | 60 °C | 10 s | |
| 72 °C | 15 s | |||
| HSP72 | Fw: | 5′-CCGGCAAGGCCAACAAGATC-3′ | 95 °C | 10 s |
| Rv: | 5′-CCTCCACGGCGCTCTTCATG-3′ | 62 °C | 10 s | |
| 72 °C | 15 s | |||
| ANOVA | ||||||
|---|---|---|---|---|---|---|
| Young | Senior | S | A | S × A | ||
| Maximum Borg Index (Borg scale) | Control | 17.8 ± 0.49 | 16.2 ± 0.49 | |||
| Supplemented | 17.8 ± 0.48 | 16.6 ± 0.68 | ||||
| Time running at 90% of VO2max (min) | Control | 9.02 ± 2.99 | 8.84 ± 2.28 | |||
| Supplemented | 7.62 ± 2.59 | 10.0 ± 1.97 | ||||
| Initial body temperature (°C) | Control | 37.0 ± 0.18 | 37.1 ± 0.14 | |||
| Supplemented | 37.0 ± 0.07 | 37.0 ± 0.05 | ||||
| Maximum body temperature (°C) | Control | 39.8 ± 0.26 | 39.4 ± 0.15 | |||
| Supplemented | 39.4 ± 0.12 | 39.3 ± 0.20 | ||||
| Body temperature 5 min after the end (°C) | Control | 39.9 ± 0.23 | 39.5 ± 0.16 | |||
| Supplemented | 39.4 ± 0.15 | 39.3 ± 0.18 | ||||
| Maximum skin temperature (°C) | Control | 35.1 ± 0.21 | 34.7 ± 0.36 | |||
| Supplemented | 34.8 ± 0.19 | 34.8 ± 0.21 | ||||
| Skin temperature 5 min after the end (°C) | Control | 35.2 ± 0.24 | 34.5 ± 0.70 | |||
| Supplemented | 35.1 ± 0.15 | 34.7 ± 0.12 | ||||
| Maximum Lactate (mM) | Control | 4.08 ± 0.61 | 4.32 ± 0.68 | |||
| Supplemented | 3.86 ± 0.85 | 5.36 ± 0.41 | ||||
| Maximal heart rate (beats/min) | Control | 186 ± 4.60 | 175 ± 7.46 | |||
| Supplemented | 183 ± 6.39 | 175 ± 6.56 | ||||
| % of maximal heart rate during the exercise | Control | 97.4 ± 0.81 | 98.0 ±2.69 | |||
| Supplemented | 96.3 ± 0.68 | 98.0 ± 1.91 | ||||
| Water intake (L) | Control | 0.42 ± 0.06 | 0.41 ± 0.17 | |||
| Supplemented | 0.50 ± 0.09 | 0.42 ± 0.17 | ||||
| Absolute weight loss (%) | Control | 1.63 ± 0.08 | 1.41 ± 0.19 | |||
| Supplemented | 1.52 ± 0.17 | 1.66 ± 0.19 | ||||
| Wight loss without water intake (%) | Control | 2.21 ± 0.13 | 1.93 ± 0.09 | |||
| Supplemented | 2.22 ± 0.09 | 2.15 ± 0.13 | ||||
| Physiological Strain Index [36] | Control | 10.5 ± 0.50 | 9.73 ± 0.31 | |||
| Supplemented | 9.79 ± 0.24 | 9.49 ± 0.41 | ||||
| Heat Storage(W/m2) | Control | 348 ± 34.1 | 325 ± 19.2 | |||
| Supplemented | 318 ± 18.7 | 344 ± 47.8 | ||||
| Pre-Exercise | Post-Exercise | ANOVA | ||||
|---|---|---|---|---|---|---|
| Young | Senior | Young | Senior | |||
| sL-Selectin (ng × mL plasma) | Control | 106 ± 2.8 ab | 80.1 ± 9.7 ac | 146 ± 26 b | 89.1 ± 11.9 ac | S, S × E |
| Supplemented | 77.7± 8.1 ac | 92.4 ± 23.1 ac | 61.4 ± 6.2 ac | 47.8 ± 13.4 c | ||
| ICAM3 (ng of × mL plasma) | Control | 458 ± 122 ade | 507 ± 55 acd | 670 ± 104 c | 705 ± 43 c | A, S, S × A, S × E |
| Supplemented | 324 ± 65 ab | 604 ± 46 cd | 206 ±73 b | 618 ± 72 ce | ||
| HSP 70 (Pg × mL of plasma) | Control | 5.12 ± 0.31 | 5.01 ± 0.19 | 5.09 ± 0.33 | 4.97 ± 0.22 | |
| Supplemented | 4.98 ± 0.42 | 5.03 ± 0.18 | 4.87 ± 0.47 | 4.74 ± 0.09 | ||
| IL-6 (pg × mL of plasma) | Control | 4.51 ± 0.35 ac | 4.27 ± 0.46 ac | 4.56 ± 0.40 ac | 5.39 ± 0.49 ab | S × E × A |
| Supplemented | 3.54 ± 0.13 c | 4.87 ± 0.36 ac | 7.38 ± 1.21 b | 4.24 ± 0.36 ac | ||
| TNFα (pg × mL of plasma) | Control | 627 ± 43 afd | 679 ± 67 acfd | 697 ± 52 acfd | 524 ± 35 f | S, S × E × A |
| Supplemented | 831 ± 66 cg | 781 ± 32.4 abcd | 758 ± 43 gd | 936 ± 140 gb | ||
| Lipoxina (pg × mL of plasma) | Control | 54.8 ± 11.7 | 67.9 ± 19.9 | 65.2 ± 12.5 | 33.5 ±10.6 # | E |
| Supplemented | 104 ± 29 | 83.4 ± 18.5 | 67.2 ± 9.9 | 36.7 ± 8.8 # | ||
| PGE2 (pg × mL of plasma) | Control | 286 ± 63 | 398 ± 168 | 315 ± 71 | 1541± 579 # | E |
| Supplemented | 390 ± 112 | 781 ± 231 | 1557 ± 585 # | 993 ± 476 | ||
| PGE1 (pg × mL of plasma) | Control | 1976 ± 168 | 2598 ± 735 | 3087± 681 | 5645 ± 3038 | |
| Supplemented | 3593 ± 995 | 3273 ± 1204 | 2465 ± 427 | 3947 ± 980 | ||
| NFκβ (U.A.) | Control | 2628 ± 140 | 2720 ± 249 | 2877 ±135 | 2889 ±194 | E |
| Supplemented | 2437 ± 210 | 2730 ± 210 | 3099 ± 343 # | 3400 ± 167 # | ||
| Pre-Exercise | Post-Exercise | ANOVA | ||||
|---|---|---|---|---|---|---|
| Young | Senior | Young | Senior | |||
| TLR2 | Control | 1.00 ± 0.28 | 1.59 ± 0.56 | 0.99 ± 0.29 | 0.77 ± 0.10 | E × A |
| Supplemented | 1.13 ± 0.37 | 1.01 ± 0.26 | 2.51 ± 1.11 | 0.84 ± 0.13 | ||
| TLR4 | Control | 1.00 ± 0.24 | 1.18 ± 0.41 | 1.25 ± 0.47 | 0.86 ± 0.12 | |
| Supplemented | 1.46 ± 0.55 | 1.15 ± 0.29 | 2.62 ± 1.19 | 0.84 ± 0.11 | ||
| NFκβ | Control | 1.00 ± 0.21 | 1.12 ± 0.34 | 1.11 ± 0.30 | 0.89 ± 0.12 | |
| Supplemented | 1.75 ± 0.79 | 1.74 ± 0.67 | 1.39 ± 0.46 | 0.87 ± 0.09 | ||
| COX2 | Control | 1.00 ± 0.09 | 1.07 ± 0.17 | 1.39 ± 0.38 | 1.50 ± 0.41 | A (0.094) |
| Supplemented | 2.31 ± 1.09 | 1.36 ± 0.36 | 3.99 ± 1.96 | 1.20 ± 0.24 | S × A (0.066) | |
| 5LOX | Control | 1.00 ± 0.19 | 1.21 ± 0.40 | 1.14 ± 0.28 | 1.10 ± 0.28 | |
| Supplemented | 1.15 ± 0.34 | 1.57 ± 0.53 | 3.59 ± 1.97 | 0.96 ± 0.13 | ||
| 15LOX2 | Control | 1.00 ± 0.33 | 1.63 ± 0.45 | 0.96 ± 0.22 | 2.14 ± 0.77 | A (0.035) |
| Supplemented | 2.14 ± 0.77 | 6.86 ± 4.21 *,$ | 1.13 ± 0.36 | 7.65 ± 2.52 *,$ | S (0.048) | |
| IL1β | Control | 1.00 ± 0.14 a | 0.99 ± 0.17 a | 1.17 ± 0.25 a | 1.68 ± 0.73 a | S (0.084) |
| Supplemented | 1.92 ± 0.59 a | 1.10 ± 0.22 a | 3.86 ± 1.54 b | 1.28 ± 0.34 a | S × A (0.043) | |
| IL-8 | Control | 1.00 ± 0.28 a | 0.92 ± 0.20 a | 0.74 ± 0.11 a | 1.74 ± 0.75 a | S × A (0.029) |
| Supplemented | 2.39 ± 1.19 ab | 0.94 ± 0.29 a | 4.10 ± 2.48 b | 0.73 ± 0.11 a | ||
| TNFα | Control | 1.00 ± 0.11 | 1.06 ± 0.19 | 1.07 ± 0.21 | 1.19 ± 0.39 | S (0.032) |
| Supplemented | 1.16 ± 0.21 | 1.21 ± 0.42 | 1.72 ± 0.58 | 2.37 ± 0.59 | ||
| IL-10 | Control | 1.00 ± 0.23 | 1.56 ± 0.60 | 1.45 ± 0.59 | 1.15 ± 0.37 | |
| Supplemented | 1.71 ± 0.75 | 1.51 ± 0.45 | 3.41 ± 1.66 | 1.54 ± 0.57 | ||
| IL-15 | Control | 1.00 ± 0.25 | 0.90 ± 0.23 | 0.91 ± 0.17 | 1.35 ± 0.52 | |
| Supplemented | 1.18 ± 0.35 | 1.16 ± 0.35 | 1.97 ± 0.89 | 0.84 ± 0.15 | ||
| HSP72 | Control | 1.00 ± 0.28 | 1.07 ± 0.31 | 1.01 ± 0.31 | 0.81 ± 0.13 | |
| Supplemented | 2.48 ± 1.39 | 1.23 ± 0.49 | 2.53 ± 1.17 | 0.79 ± 0.12 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capó, X.; Martorell, M.; Sureda, A.; Riera, J.; Drobnic, F.; Tur, J.A.; Pons, A. Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age. Nutrients 2016, 8, 619. https://doi.org/10.3390/nu8100619
Capó X, Martorell M, Sureda A, Riera J, Drobnic F, Tur JA, Pons A. Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age. Nutrients. 2016; 8(10):619. https://doi.org/10.3390/nu8100619
Chicago/Turabian StyleCapó, Xavier, Miquel Martorell, Antoni Sureda, Joan Riera, Franchek Drobnic, Josep Antoni Tur, and Antoni Pons. 2016. "Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age" Nutrients 8, no. 10: 619. https://doi.org/10.3390/nu8100619
APA StyleCapó, X., Martorell, M., Sureda, A., Riera, J., Drobnic, F., Tur, J. A., & Pons, A. (2016). Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age. Nutrients, 8(10), 619. https://doi.org/10.3390/nu8100619