Protection against Oxygen-Glucose Deprivation/Reperfusion Injury in Cortical Neurons by Combining Omega-3 Polyunsaturated Acid with Lyciumbarbarum Polysaccharide

Abstract
:1. Introduction
2. Materials and Method
2.1. Animals
2.2. Primary Cortical Neuron Cultures and Oxygen-Glucose Deprivation/Reperfusion (OGD/R)
2.3. Drugs
2.4. Antibodies
2.5. Immunocytochemistry
2.6. Genomic DNA Extractions and PCR Amplification
2.7. Fatty Acid Analysis
2.8. Cell Viability Assay
2.9. TUNEL Staining
2.10. Intracellular Calcium (Ca2+) Measurements
2.11. Western Blotting Analysis
2.12. Statistical Analysis
3. Results
3.1. Identification, Genotyping and Fatty Acid Profiles of Primary Cortical Neurons


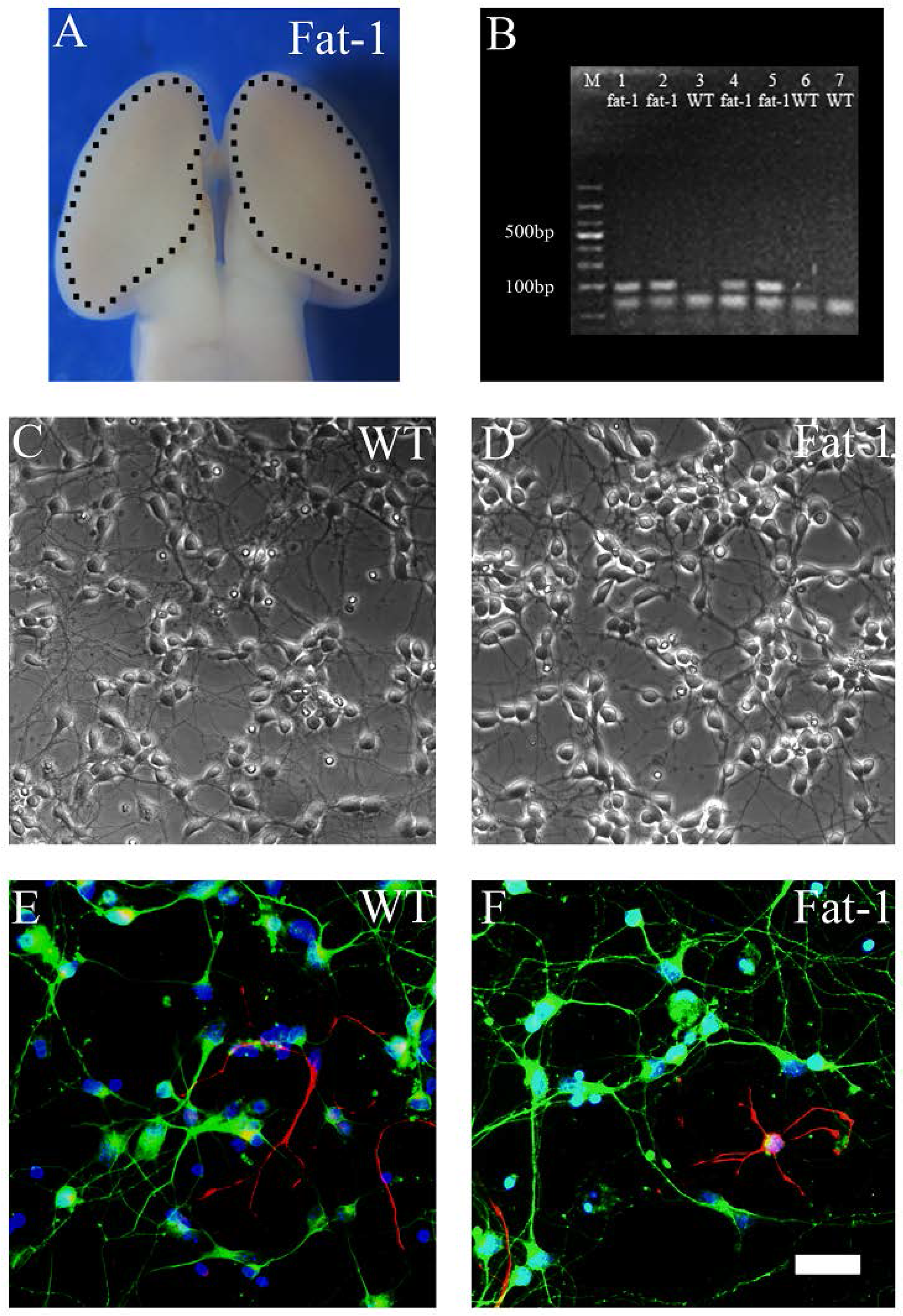{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | WT | fat-1 |
|---|---|---|
| C14:0 | 3.01 ± 0.25 | 1.77 ± 0.33 * |
| C16:0 | 25.22 ± 0.27 | 24.20 ± 0.46 |
| C16:1,9 | 8.65 ± 0.23 | 5.32 ± 0.57 ** |
| C18:0 | 13.12 ± 0.85 | 16.11 ± 0.21 * |
| C18:1,9 | 33.12 ± 0.34 | 28.97 ± 0.12 ** |
| C18:2,6 | 0.78 ± 0.23 | 0.77 ± 0.02 |
| C18:3,3 (ALA) | 0.11 ± 0.06 | 0.47 ± 0.11 ** |
| C20:0 | 0.30 ± 0.05 | 0.27 ± 0.02 |
| C20:1,9 | 1.22 ± 0.03 | 1.14 ± 0.02 |
| C20:2,6 | 4.661 ± 0.24 | 2.88 ± 0.41 * |
| C20:4,6 (AA) | 5.11 ± 0.18 | 0.88 ± 0.09 ** |
| C20:5,3 (EPA) | 0.33 ± 0.00 | 3.79 ± 0.73 ** |
| C22:0 | 0.20 ± 0.03 | 0.54 ± 0.09 * |
| C22:1,9 | 3.56 ± 0.70 | 3.95 ± 0.63 |
| C22:5,3 (DPA) | 0.99 ± 0.03 | 4.37 ± 0.31 ** |
| C22:6,3 (DHA) | 1.02 ± 0.14 | 2.90 ± 0.03 ** |
| C24:1 | 0.92 ± 0.12 | 1.32 ± 0.17 |
| SFA | 41.85 ± 0.34 | 42.89 ± 1.09 |
| MUFA | 47.47 ± 1.25 | 40.70 ± 1.09 ** |
| PUFA | 13.00 ± 1.05 | 16.06 ± 1.72 * |
| ω-6/ω-3 | 4.31 ± 4.03 | 0.39 ± 0.26 ** |
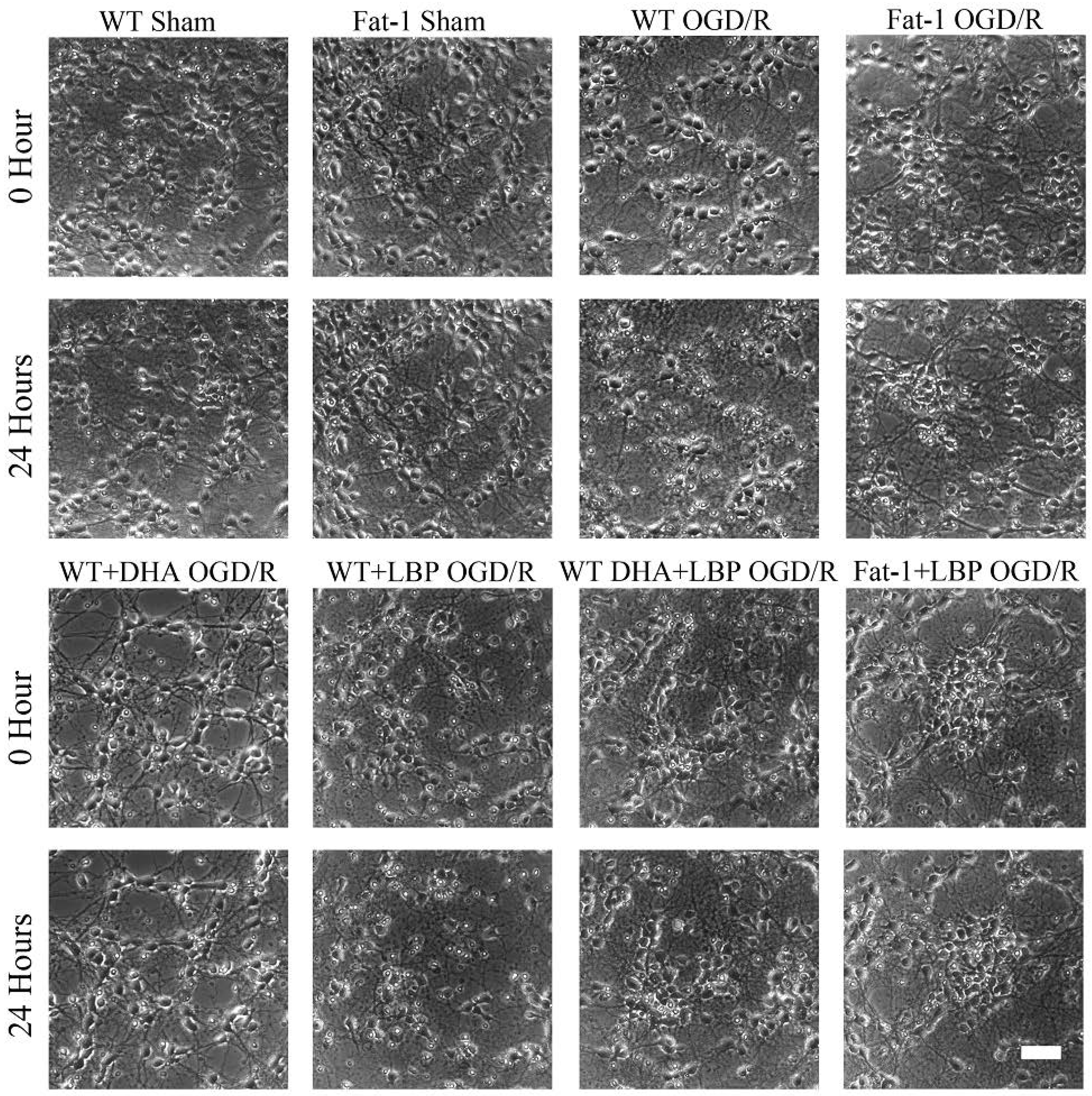
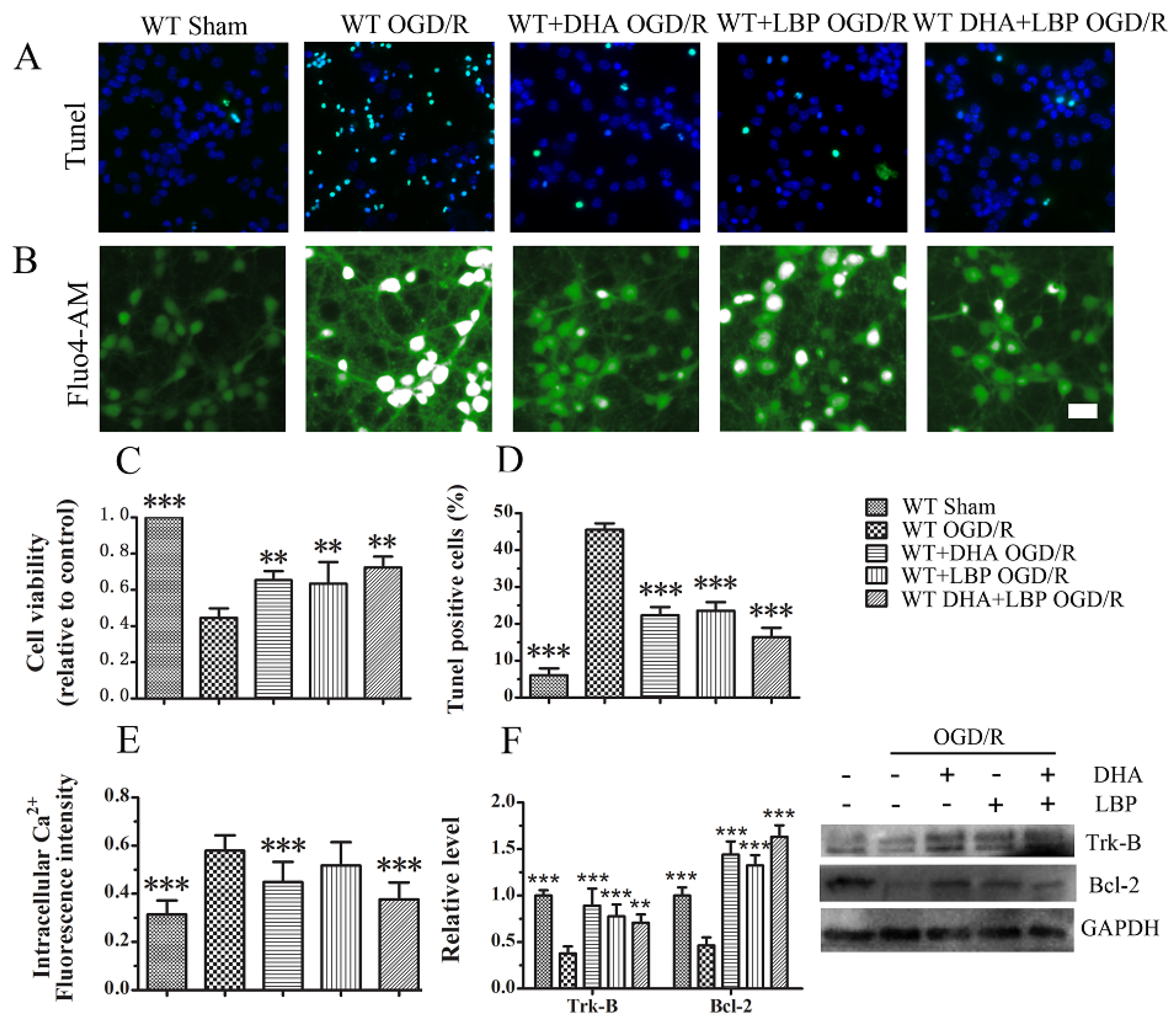
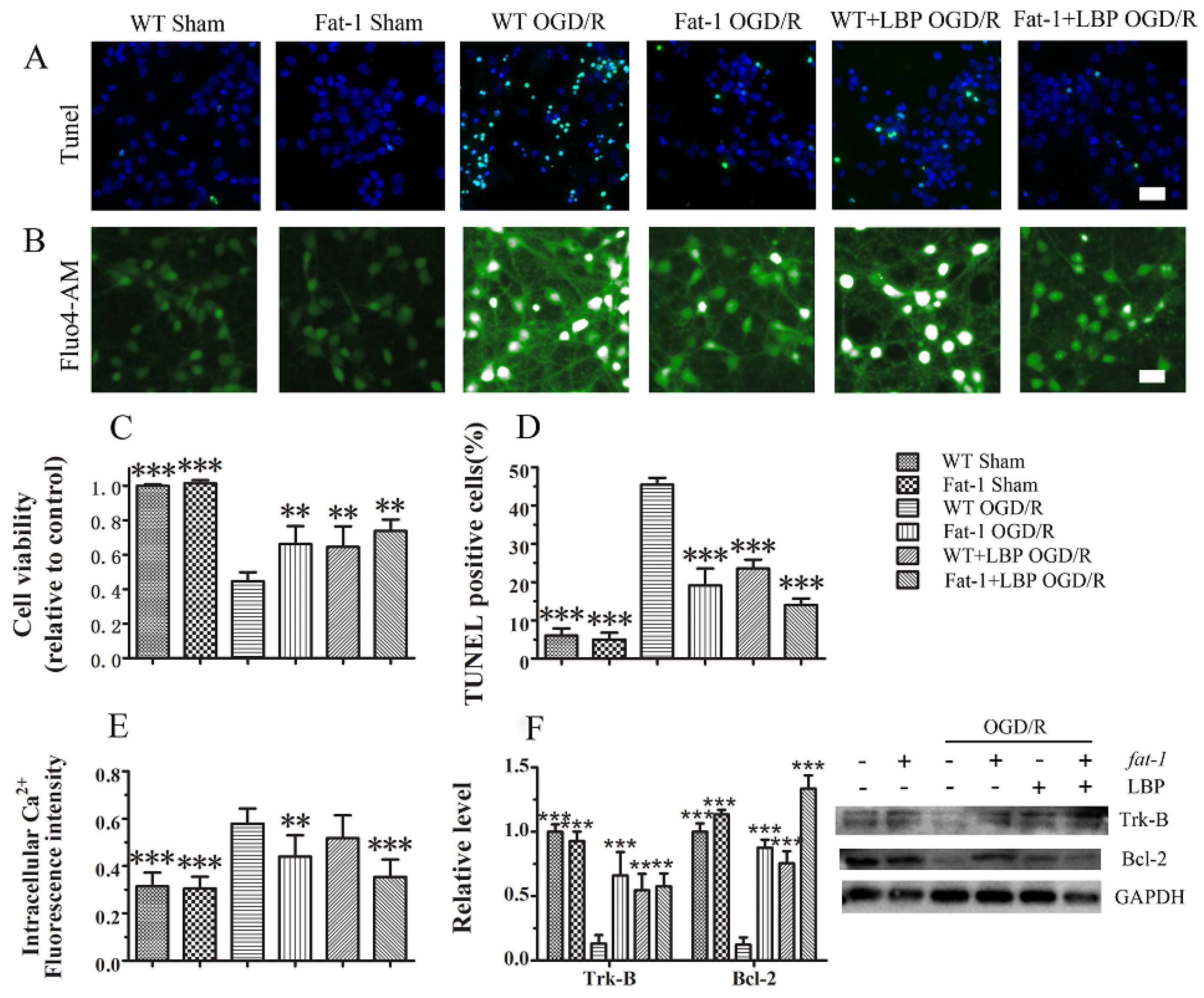
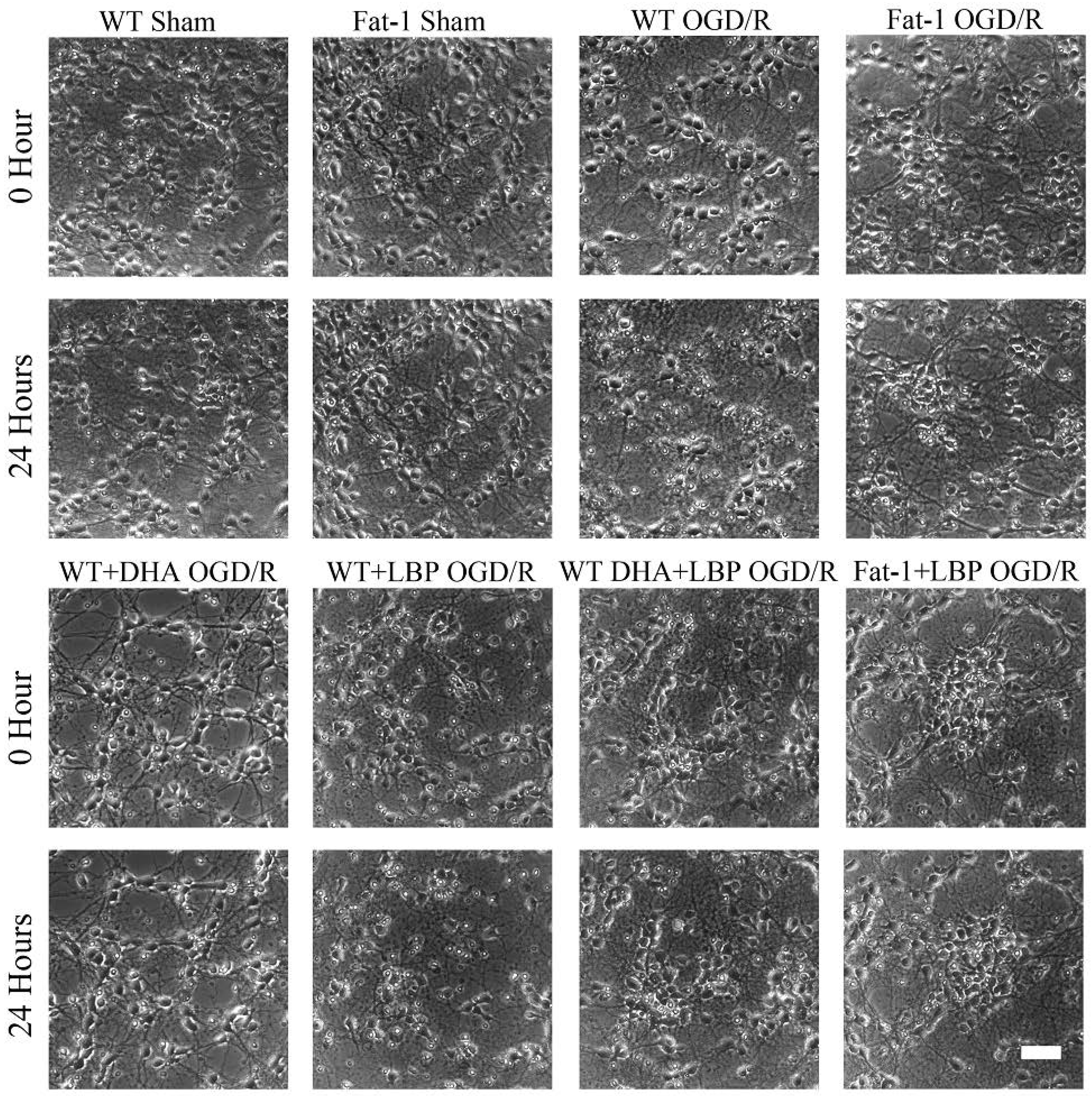
3.2. LBP Either Together with DHA or Endogenous ω-3 PUFAs Rescues Cortical Neurons from OGD/R Insults
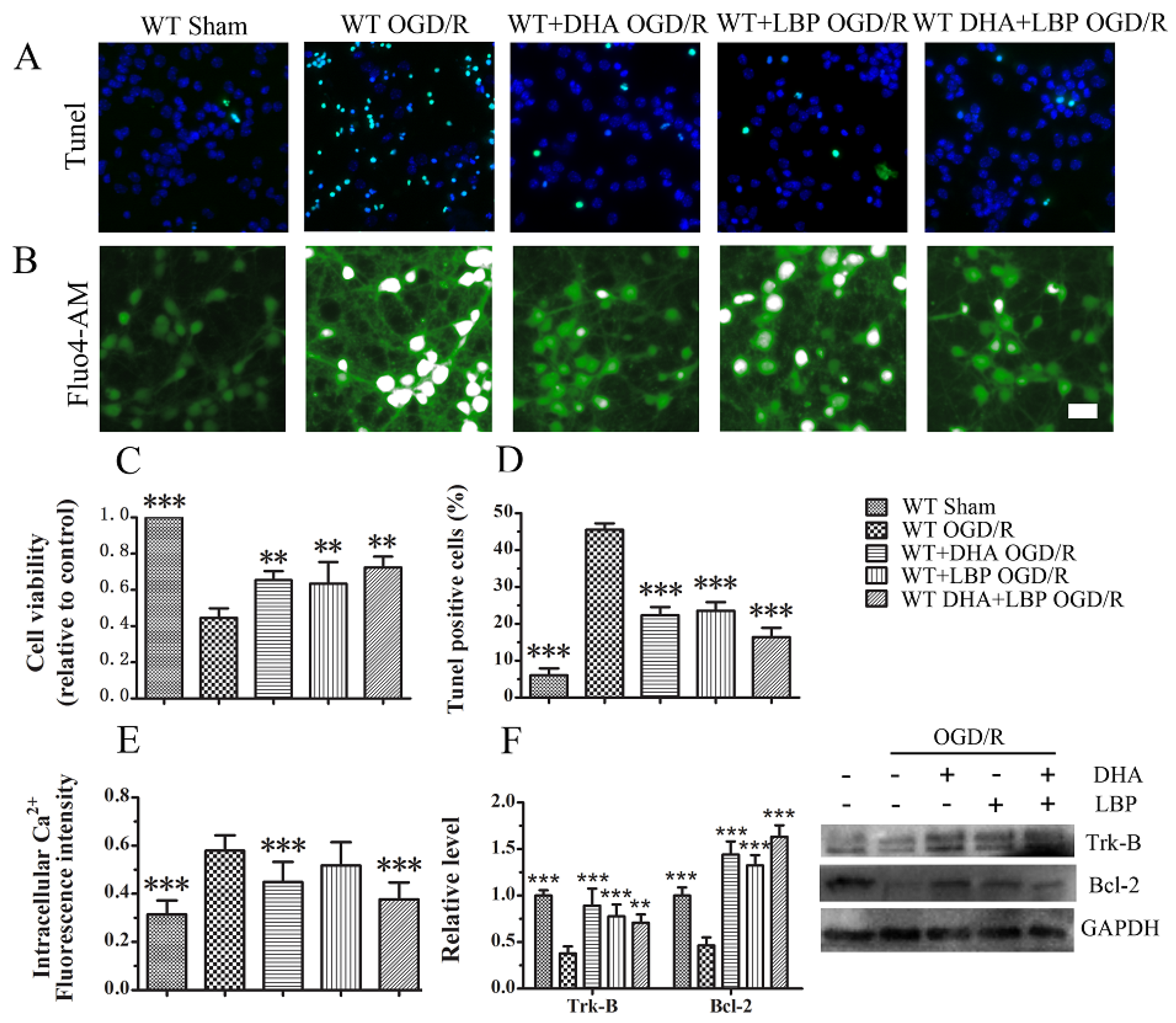
3.3. LBP Either Together with DHA or Endogenous ω-3 PUFAs Significantly Prevents OGD/R-Induced Neuronal Apoptosis via Intracellular Ca2+ Handling or Neurotrophic Pathway Activation
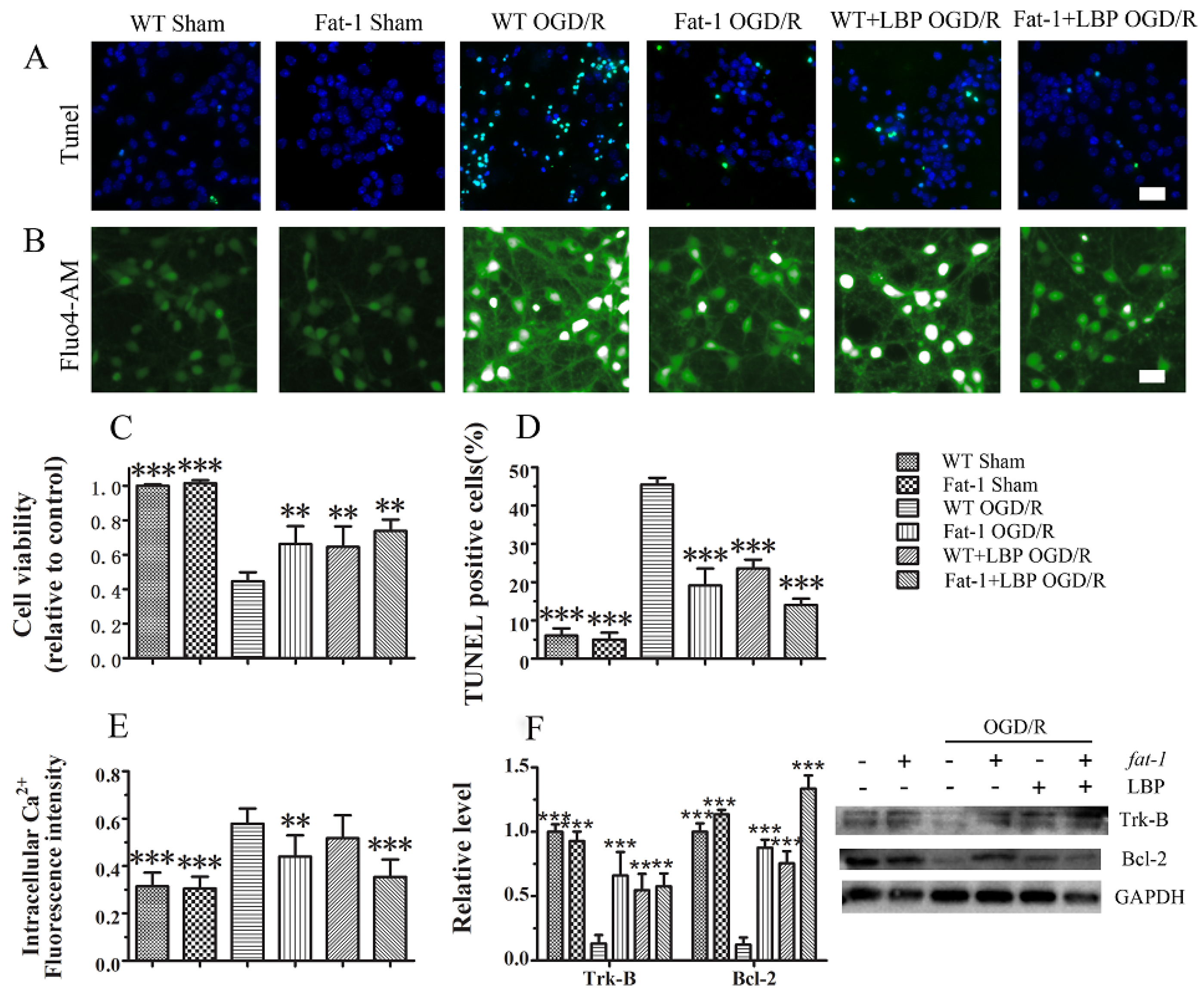
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rothwell, P.M.; Algra, A.; Amarenco, P. Medical treatment in acute and long-term secondary prevention after transient ischaemic attack and ischaemic stroke. Lancet 2011, 377, 1681–1692. [Google Scholar] [CrossRef]
- O’Collins, V.E.; Macleod, M.R.; Donnan, G.A.; Horky, L.L.; van der Worp, B.H.; Howells, D.W. 1026 experimental treatments in acute stroke. Ann. Neurol. 2006, 59, 467–477. [Google Scholar] [PubMed]
- Stankowski, J.N.; Gupta, R. Therapeutic targets for neuroprotection in acute ischemic stroke: Lost in translation? Antioxid. Redox Signal. 2011, 14, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- George, P.M.; Steinberg, G.K. Novel stroke therapeutics: Unraveling stroke pathophysiology and its impact on clinical treatments. Neuron 2015, 87, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Khoutorova, L.; Atkins, K.D.; Bazan, N.G. Robust docosahexaenoic acid-mediated neuroprotection in a rat model of transient, focal cerebral ischemia. Stroke 2009, 40, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.T.; Hu, X.M.; Yang, W.; Gao, Y.Q.; Chen, J. Omega-3 polyunsaturated fatty acid supplementation confers long-term neuroprotection against neonatal hypoxic-ischemic brain injury through anti-inflammatory actions. Stroke 2010, 41, 2341–2347. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Khoutorova, L.; Atkins, K.D.; Eady, T.N.; Hong, S.; Lu, Y.; Obenaus, A.; Bazan, N.G. Docosahexaenoic acid therapy of experimental ischemic stroke. Transl. Stroke Res. 2011, 2, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Oudin, A.; Wennberg, M. Fish consumption and ischemic stroke in southern sweden. Nutr. J. 2011, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhang, F.; Leak, R.K.; Zhang, W.; Iwai, M.; Stetler, R.A.; Dai, Y.; Zhao, A.; Gao, Y.; Chen, J. Transgenic overproduction of omega-3 polyunsaturated fatty acids provides neuroprotection and enhances endogenous neurogenesis after stroke. Curr. Mol. Med. 2013, 13, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, S.; Mao, L.; Leak, R.K.; Shi, Y.; Zhang, W.; Hu, X.; Sun, B.; Cao, G.; Gao, Y.; et al. Omega-3 fatty acids protect the brain against ischemic injury by activating Nrf2 and upregulating heme oxygenase 1. J. Neurosci. 2014, 34, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Kichev, A.; Rousset, C.I.; Baburamani, A.A.; Levison, S.W.; Wood, T.L.; Gressens, P.; Thornton, C.; Hagberg, H. Tnf-related apoptosis-inducing ligand (TRAIL) signaling and cell death in the immature central nervous system after hypoxia-ischemia and inflammation. J. Biol. Chem. 2014, 289, 9430–9439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Spite, M. Resolvins: Anti-inflammatory and proresolving mediators derived from omega-3 polyunsaturated fatty acids. Annu. Rev. Nutr. 2012, 32, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Ren, H.; Wan, J.B.; Yao, X.; Zhang, X.; He, C.; So, K.F.; Kang, J.X.; Pei, Z.; Su, H. Enriched endogenous omega-3 fatty acids in mice protect against global ischemia injury. J. Lipid Res. 2014, 55, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.H.; Xu, J.F.; Xue, R.H.; Zheng, W.F.; Liu, Z.L. Protective effect of chronic ethyl docosahexaenoate administration on brain injury in ischemic gerbils. Pharmacol. Biochem. Behav. 2004, 79, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Siwek, M.; Sowa-Kucma, M.; Dudek, D.; Styczen, K.; Szewczyk, B.; Kotarska, K.; Misztakk, P.; Pilc, A.; Wolak, M.; Nowak, G. Oxidative stress markers in affective disorders. Pharmacol. Rep. 2013, 65, 1558–1571. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J.; Wu, L.; Kang, Z.B. Transgenic mice: Fat-1 mice convert n-6 to ω-3 fatty acids. Nature 2004, 427, 504. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Wang, Y.; Zhou, R.; Ma, L.; Hao, Y.; Jin, S.; Du, J.; Zhao, C.; Sun, T.; et al. Lycium barbarum polysaccharide prevents focal cerebral ischemic injury by inhibiting neuronal apoptosis in mice. PLoS ONE 2014, 9, e90780. [Google Scholar] [CrossRef] [PubMed]
- Rui, C.; Yuxiang, L.; Yinju, H.; Qingluan, Z.; Yang, W.; Qipeng, Z.; Hao, W.; Lin, M.; Juan, L.; Chengjun, Z.; et al. Protective effects of lycium barbarum polysaccharide on neonatal rat primary cultured hippocampal neurons injured by oxygen-glucose deprivation and reperfusion. J. Mol. Histol. 2012, 43, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Liu, M.; Cao, J.; Yao, M.; Lu, Y.; Li, J.; Zhu, X.; Yang, Z.; Wen, A. Protective effects of lycium barbarum polysaccharide on 6-OHDA-induced apoptosis in PC12 cells through the ROS-NO pathway. Molecules 2015, 20, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Li, Y.X.; Hao, Y.J.; Wang, T.F.; Lei, Z.; Wu, Y.; Zhao, Q.P.; Ang, H.; Ma, L.; Liu, J.; et al. Neuroprotective effects of LBP on brain ischemic reperfusion neurodegeneration. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2760–2765. [Google Scholar] [PubMed]
- Ureshino, R.P.; Rocha, K.K.; Lopes, G.S.; Bincoletto, C.; Smaili, S.S. Calcium signaling alterations, oxidative stress, and autophagy in aging. Antioxid. Redox Signal. 2014, 21, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; You, Y.; Gupta, V.B.; Klistorner, A.; Graham, S.L. Trkb receptor signalling: Implications in neurodegenerative, psychiatric and proliferative disorders. Int. J. Mol. Sci. 2013, 14, 10122–10142. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.S.; Leung, S.K.; Lai, S.W.; Che, C.M.; Zee, S.Y.; So, K.F.; Yuen, W.H.; Chang, R.C. Neuroprotective effects of anti-aging oriental medicine lycium barbarum against beta-amyloid peptide neurotoxicity. Exp. Gerontol. 2005, 40, 716–727. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Qu, X.; Cui, L.; Wang, J.; Kang, J.X. Improved spatial learning performance of fat-1 mice is associated with enhanced neurogenesis and neuritogenesis by docosahexaenoic acid. Proc. Natl. Acad. Sci. USA 2009, 106, 11370–11375. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, D.; Ni, N.; Ren, H.; Luo, C.; He, C.; Kang, J.X.; Wan, J.B.; Su, H. Omega-3 polyunsaturated fatty acids protect neural progenitor cells against oxidative injury. Mar. Drugs 2014, 12, 2341–2356. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.B.; Huang, L.L.; Rong, R.; Tan, R.; Wang, J.; Kang, J.X. Endogenously decreasing tissue n-6/ω-3 fatty acid ratio reduces atherosclerotic lesions in apolipoprotein e-deficient mice by inhibiting systemic and vascular inflammation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Doyle, K.P.; Simon, R.P.; Stenzel-Poore, M.P. Mechanisms of ischemic brain damage. Neuropharmacology 2008, 55, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G.; Marcheselli, V.L.; Cole-Edwards, K. Brain response to injury and neurodegeneration: Endogenous neuroprotective signaling. Ann. N. Y. Acad. Sci. 2005, 1053, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Akbar, M.; Kim, K.Y. Inhibition of neuronal apoptosis by polyunsaturated fatty acids. J. Mol. Neurosci. 2001, 16, 223–227. [Google Scholar] [CrossRef]
- D’Orsi, B.; Kilbride, S.M.; Chen, G.; Perez Alvarez, S.; Bonner, H.P.; Pfeiffer, S.; Plesnila, N.; Engel, T.; Henshall, D.C.; Dussmann, H.; et al. Bax regulates neuronal Ca2+ homeostasis. J. Neurosci. 2015, 35, 1706–1722. [Google Scholar] [CrossRef] [PubMed]
- Begum, G.; Kintner, D.; Liu, Y.; Cramer, S.W.; Sun, D. Dha inhibits er Ca2+ release and er stress in astrocytes following in vitro ischemia. J. Neurochem. 2012, 120, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Khairallah, R.J.; O’Shea, K.M.; Brown, B.H.; Khanna, N.; des Rosiers, C.; Stanley, W.C. Treatment with docosahexaenoic acid, but not eicosapentaenoic acid, delays Ca2+-induced mitochondria permeability transition in normal and hypertrophied myocardium. J. Pharmacol. Exp. Ther. 2010, 335, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Malagelada, C.; Xifro, X.; Minano, A.; Sabria, J.; Rodriguez-Alvarez, J. Contribution of caspase-mediated apoptosis to the cell death caused by oxygen-glucose deprivation in cortical cell cultures. Neurobiol. Dis. 2005, 20, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Broughton, B.R.; Reutens, D.C.; Sobey, C.G. Apoptotic mechanisms after cerebral ischemia. Stroke 2009, 40, e331–e339. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Cao, G.; Johnnides, M.J.; Signore, A.P.; Luo, Y.; Hickey, R.W.; Chen, J. TAT-mediated delivery of Bcl-xL protein is neuroprotective against neonatal hypoxic-ischemic brain injury via inhibition of caspases and AIF. Neurobiol. Dis. 2006, 21, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yenari, M.A.; Cheng, D.; Sapolsky, R.M.; Steinberg, G.K. Bcl-2 overexpression protects against neuron loss within the ischemic margin following experimental stroke and inhibits cytochrome C translocation and caspase-3 activity. J. Neurochem. 2003, 85, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Pei, W.; Ge, H.; Liang, Q.; Luo, Y.; Sharp, F.R.; Lu, A.; Ran, R.; Graham, S.H.; Chen, J. In vivo delivery of a Bcl-xL fusion protein containing the TAT protein transduction domain protects against ischemic brain injury and neuronal apoptosis. J. Neurosci. 2002, 22, 5423–5431. [Google Scholar] [PubMed]
- Rong, Y.P.; Bultynck, G.; Aromolaran, A.S.; Zhong, F.; Parys, J.B.; de Smedt, H.; Mignery, G.A.; Roderick, H.L.; Bootman, M.D.; Distelhorst, C.W. The BH4 domain of Bcl-2 inhibits er calcium release and apoptosis by binding the regulatory and coupling domain of the IP3 receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 14397–14402. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Rizzuto, R. Bcl-2 and Ca2+ homeostasis in the endoplasmic reticulum. Cell Death Differ. 2006, 13, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Trk receptors: Roles in neuronal signal transemuction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [PubMed]
- Balogun, K.A.; Cheema, S.K. The expression of neurotrophins is differentially regulated by omega-3 polyunsaturated fatty acids at weaning and postweaning in C57BL/6 mice cerebral cortex. Neurochem. Int. 2014, 66, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gladman, S.J.; Huang, W.; Lim, S.N.; Dyall, S.C.; Boddy, S.; Kang, J.X.; Knight, M.M.; Priestley, J.V.; Michael-Titus, A.T. Improved outcome after peripheral nerve injury in mice with increased levels of endogenous omega-3 polyunsaturated fatty acids. J. Neurosci. 2012, 32, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X. Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 263–267. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Wu, D.; Yao, J.-P.; Yao, X.; Huang, Z.; Li, P.; Wan, J.-B.; He, C.; Su, H. Protection against Oxygen-Glucose Deprivation/Reperfusion Injury in Cortical Neurons by Combining Omega-3 Polyunsaturated Acid with Lyciumbarbarum Polysaccharide. Nutrients 2016, 8, 41. https://doi.org/10.3390/nu8010041
Shi Z, Wu D, Yao J-P, Yao X, Huang Z, Li P, Wan J-B, He C, Su H. Protection against Oxygen-Glucose Deprivation/Reperfusion Injury in Cortical Neurons by Combining Omega-3 Polyunsaturated Acid with Lyciumbarbarum Polysaccharide. Nutrients. 2016; 8(1):41. https://doi.org/10.3390/nu8010041
Chicago/Turabian StyleShi, Zhe, Di Wu, Jian-Ping Yao, Xiaoli Yao, Zhijian Huang, Peng Li, Jian-Bo Wan, Chengwei He, and Huanxing Su. 2016. "Protection against Oxygen-Glucose Deprivation/Reperfusion Injury in Cortical Neurons by Combining Omega-3 Polyunsaturated Acid with Lyciumbarbarum Polysaccharide" Nutrients 8, no. 1: 41. https://doi.org/10.3390/nu8010041
APA StyleShi, Z., Wu, D., Yao, J.-P., Yao, X., Huang, Z., Li, P., Wan, J.-B., He, C., & Su, H. (2016). Protection against Oxygen-Glucose Deprivation/Reperfusion Injury in Cortical Neurons by Combining Omega-3 Polyunsaturated Acid with Lyciumbarbarum Polysaccharide. Nutrients, 8(1), 41. https://doi.org/10.3390/nu8010041