Milk Intolerance, Beta-Casein and Lactose


{kind=link}
Abstract
:1. Introduction
2. Literature Search and Selection of Studies for Review
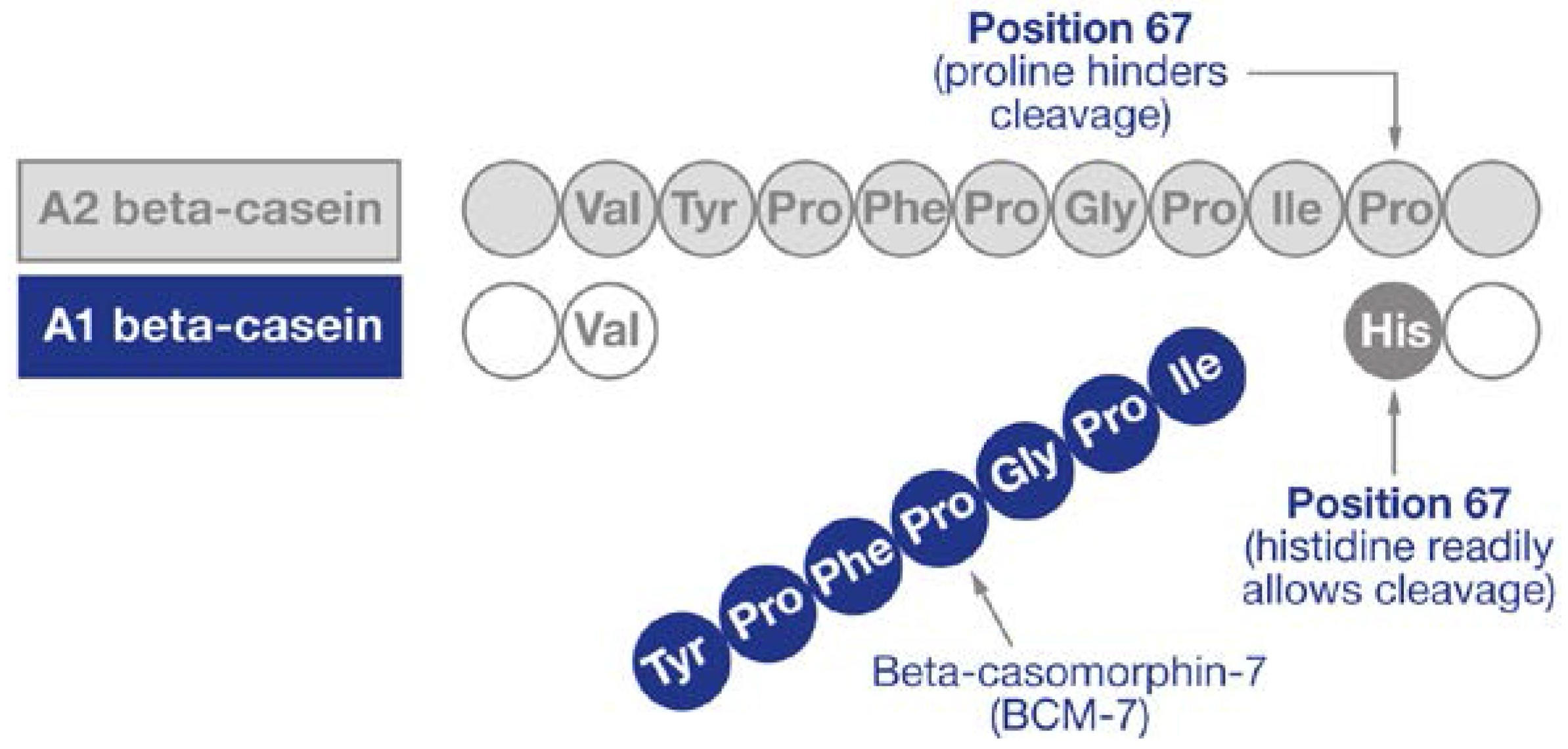
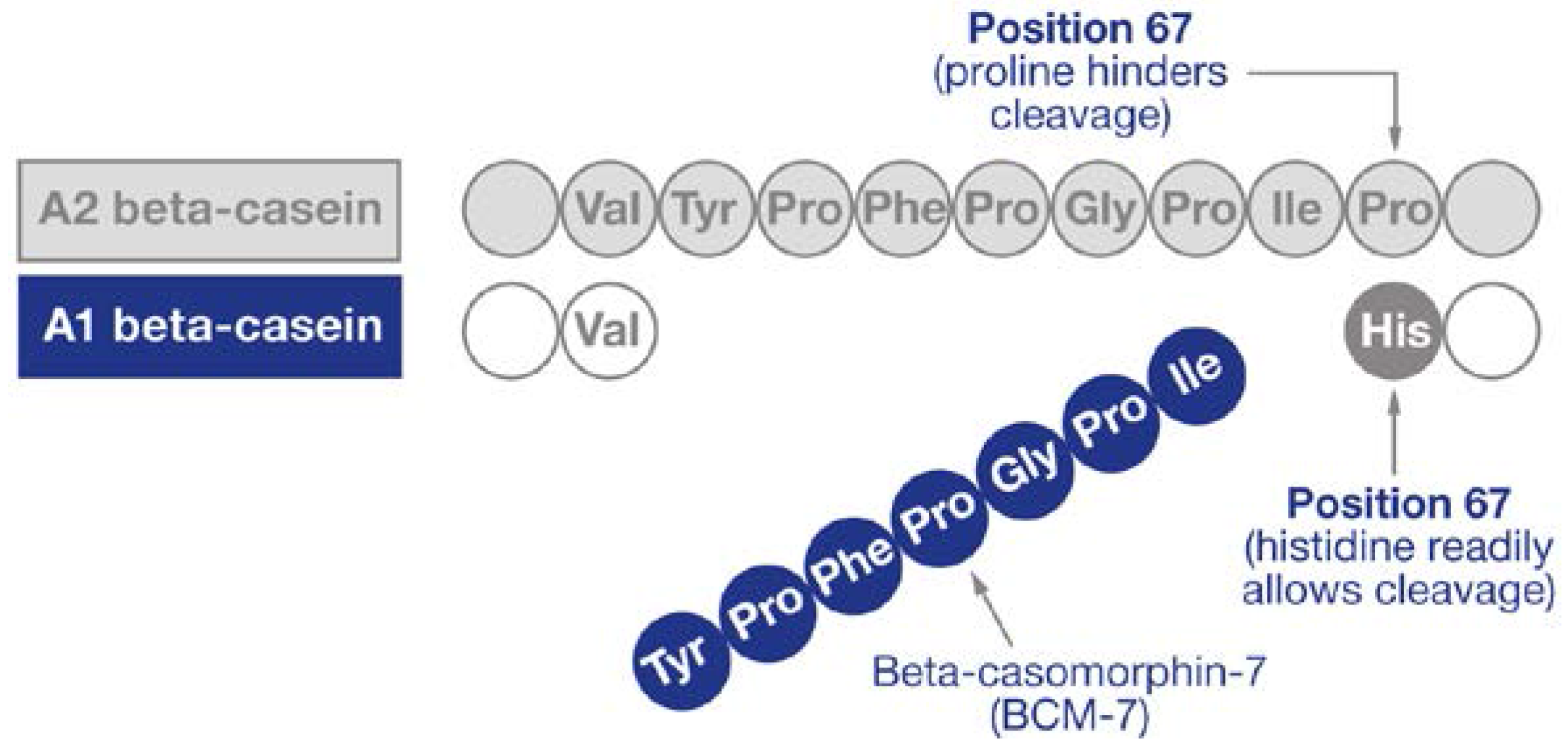
3. Beta-Caseins and BCM-7
4. Opioid Characteristics of Casomorphins
5. Beta-Caseins, Beta-Casomorphins and Delayed Intestinal Transit
6. Inflammatory and Immune Responses to Casomorphins in the Gastrointestinal System
7. Clinical Studies of Beta-Casein Effects in the Gastrointestinal System
8. The Potential for BCM-7 and Lactose Interactions
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Suchy, F.J.; Brannon, P.M.; Carpenter, T.O.; Fernandez, J.R.; Gilsanz, V.; Gould, J.B.; Hall, K.; Hui, S.L.; Lupton, J.; Mennella, J.; et al. NIH consensus development conference statement: Lactose intolerance and health. NIH Consens. State Sci. Statements 2010, 27, 1–27. [Google Scholar] [PubMed]
- Phelan, M.; Aherne, A.; FitzGerald, R.J.; O’Brien, N.M. Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatory status. Int. Dairy J. 2009, 19, 643–654. [Google Scholar] [CrossRef]
- Formaggioni, P.; Summer, A.; Malacarne, M.; Mariani, P. Milk protein polymorphism: Detection and diffusion of the genetic variants in Bos genus. Ann. Fac. Med. Vet. Univ. Parma 1999, 19, 127–165. [Google Scholar]
- Ng-Kwai-Hang, K.F.; Grosclaude, F. Genetic polymorphism of milk proteins. In Advanced Dairy Chemistry: Volume 1: Proteins, Parts A & B; Fox, P.F., McSweeney, P.L.H., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 739–816. [Google Scholar]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tomé, D.; Leonil, J. Sequential release of milk protein-derived bioactive peptides in the jejunum in healthy humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- De Noni, I.; Cattaneo, S. Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- De Noni, I. Release of β-casomorphins 5 and 7 during simulated gastro-intestinal digestion of bovine β-casein variants and milk-based infant formulas. Food Chem. 2008, 110, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and β-casomorphin from bovine beta-casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef]
- Ul Haq, M.R.; Kapila, R.; Kapila, S. Release of β-casomorphin-7/5 during simulated gastrointestinal digestion of milk β-casein variants from Indian crossbred cattle (Karan Fries). Food Chem. 2015, 168, 70–79. [Google Scholar] [CrossRef] [PubMed]
- De Noni, R.J.; FitzGerald, H.J.T.; Korhonen, Y.; Le Roux, C.T.; Livesey, I.; Thorsdottir, D.; Tomé, R.W. Scientific Report of EFSA prepared by a DATEX Working Group on the potential health impact of β-casomorphins and related peptides. EFSA Sci. Rep. 2009, 231, 1–107. [Google Scholar]
- Cielinkska, A.; Kostyra, E.B.; Kostyra, H.; Olenski, K.; Fiedorowicz, E.; Kaminski, S.A. Milk from cows of different β-casein genotypes as a source of β-casomorphin-7. Int. J. Food Sci. Nutr. 2012, 63, 426–430. [Google Scholar]
- Woodford, K. Devil in the Milk: Illness, Health and Politics: A1 and A2 Milk; Craig Potton Publishing: Wellington, New Zealand, 2007. [Google Scholar]
- Kaminski, S.; Cieslinska, A.; Kostyra, E. Polymorphism of bovine β-casein and its potential effect on human health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Mencarini, I.R.; Woodford, K.B.; Old, K.M. Comparing herd selection strategies for A2 beta-casein. Proc. N. Z. Soc. Anim. Prod. 2013, 73, 149–154. [Google Scholar]
- Nguyen, D.D.; Solah, V.A.; Johnson, S.K.; Charrois, J.W.; Busetti, F. Isotope dilution liquid chromatography-tandem mass spectrometry for simultaneous identification and quantification of beta-casomorphin 5 and beta-casomorphin 7 in yoghurt. Food Chem. 2014, 146, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Janer, C.; Arigoni, F.; Lee, B.H.; Peláez, C.; Requena, T. Enzymatic ability of Bifidobacterium animalis subsp. lactis to hydrolyze milk proteins: Identification and characterization of endopeptidase O. Appl. Environ. Microbiol. 2005, 71, 8460–8465. [Google Scholar] [PubMed]
- Hamosh, M.; Hong, H.; Hamosh, P. Beta-Casomorphins: Milk-β-casein derived opioid peptides. In Textbook of Gastroenterology and Nutrition in Infancy, 2nd ed.; Lebenthal, E., Ed.; Raven Press: New York, NY, USA, 1989; pp. 143–150. [Google Scholar]
- Wada, Y.; Lonnerdal, B. Bioactive peptides released from in vitro digestion of human milk with or without pasteurization. Pediatr. Res. 2015, 77, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Wiedemann, K.; Teschemacher, H. Opioid activities of human beta-casomorphins. Naunyn Schmiedebergs Arch. Pharmacol. 1985, 33, 351–354. [Google Scholar] [CrossRef]
- Brantl, V. Novel opioid peptides derived from human beta-casein: Human beta-casomorphins. Eur. J. Pharmacol. 1984, 106, 213–214. [Google Scholar] [CrossRef]
- Jarmolowska, B.; Sidor, K.; Iwan, M.; Bielikowicz, K.; Kaczmarski, M.; Kostyra, E.; Kostrya, H. Changes of β-casomorphin content in human milk during lactation. Peptides 2007, 28, 1982–1986. [Google Scholar] [CrossRef] [PubMed]
- Brantl, V.; Teschemacher, H.; Henschen, A.; Lottspeich, F. Novel opioid peptides derived from casein (β-casomorphins). I. Isolation from bovine casein peptone. Hoppe Seylers Z. Physiol. Chem. 1979, 360, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Henschen, A.; Lottspeich, F.; Brantl, V.; Teschemacher, H. Novel opioid peptides derived from casein (β-casomorphins). II. Structure of active components from bovine casein peptone. Hoppe Seylers Z. Physiol. Chem. 1979, 360, 1217–1224. [Google Scholar] [PubMed]
- Lottspeich, F.; Henschen, A.; Brantl, V.; Teschemacher, H. Novel opioid peptides derived from casein (β-casomorphins). III. Synthetic peptides corresponding to components from bovine casein peptone. Hoppe Seylers Z. Physiol. Chem. 1980, 361, 1835–1839. [Google Scholar] [CrossRef] [PubMed]
- Brantl, V.; Teschemacher, H.; Blasig, J.; Henschen, A.; Lottspeich, F. Opioid activities of beta-casomorphins. Life Sci. 1981, 28, 1903–1909. [Google Scholar] [CrossRef]
- Wasilewska, J.; Kaczmarski, M.; Kostyra, E.; Iwan, M. Cow’s-milk-induced infant apnoea with increased serum content of bovine β-casomorphin-5. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, J.; Sienkiewicz-Szlapka, E.; Kuzbida, E.; Jarmolowska, B.; Kaczmarski, M.; Kostyra, E. The exogenous opioid peptides and DPPIV serum activity in infants with apnoea expressed as apparent life threatening events (ALTE). Neuropeptides 2011, 45, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kost, N.V.; Sokolov, O.Y.; Kurasova, O.B.; Dmitriev, A.D.; Tarakanova, J.N.; Gabaeva, M.V.; Zolotarev, Y.A.; Dadayan, A.K.; Grachev, S.A.; Korneeva, E.V.; et al. β-casomorphins-7 in infants on different type of feeding and different levels of psychomotor development. Peptides 2009, 30, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, O.; Kost, N.; Andreeva, O.; Korneeva, E.; Meshavkin, V.; Tarakanova, Y.; Dadayan, A.; Zolotarev, Y.; Grachev, S.; Mikheeva, I.; et al. Autistic children display elevated urine levels of bovine casomorphin-7 immunoreactivity. Peptides 2014, 56, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.P.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1 β-casein affects gastrointestinal transit time, dipeptidyl peptidase-4 activity, and inflammatory status relative to A2 β-casein in Wistar rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Nakamura, T.; Kitazawa, H.; Kawai, Y.; Itoh, T. Isolation and structural analysis of antihypertensive peptides that exist naturally in Gouda cheese. J. Dairy Sci. 2000, 83, 1434–1440. [Google Scholar] [CrossRef]
- Pleuvry, B.J. Opioid receptors and their ligands: Natural and unnatural. Br. J. Anaesth. 1991, 66, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.J.; Takemori, A.E. Relative involvement of receptor subtypes in opioid-induced inhibition of gastrointestinal transit in mice. J. Pharmacol. Exp. Ther. 1983, 224, 359–363. [Google Scholar] [PubMed]
- Zoghbi, S.; Trompette, A.; Claustre, J.; El Homsi, M.; Garzon, J.; Jourdan, G.; Scoazec, J.Y.; Plaisancié, P. β-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a μ-opioid pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.E.; Davison, J.S.; Bates, S.L.; Meddings, J.B. Opioid receptors on guinea-pig intestinal crypt epithelial cells. J. Physiol. 1996, 497, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Greenwood-Van Meerveld, B.; Gardner, C.J.; Little, P.J.; Hicks, G.A.; Dehaven-Hudkins, D.L. Preclinical studies of opioids and opioid antagonists on gastrointestinal function. Neurogastroenterol. Motil. 2004, 16, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Gonenne, J.; Camilleri, M.; Ferber, I.; Burton, D.; Baxter, K.; Keyashian, K.; Foss, J.; Wallin, B.; Du, W.; Zinsmeister, A. Effect of alvimopan and codeine on gastrointestinal transit: A randomized controlled study. Clin. Gastroenterol. Hepatol. 2005, 3, 784–791. [Google Scholar] [CrossRef]
- Stephen, A.M.; Wiggins, H.S.; Cummings, J.H. Effect of changing transit time on colonic microbial metabolism in man. Gut 1987, 28, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Hempel, G.; Grecksch, G.; Matthies, H. Effects of beta-casomorphin derivatives on gastrointestinal transit in mice. Biomed. Biochim. Acta 1990, 49, 1203–1207. [Google Scholar] [PubMed]
- Defilippi, C.; Gomez, E.; Charlin, V.; Silva, C. Inhibition of small intestinal motility by casein: A role of beta casomorphins? Nutrition 1995, 11, 751–754. [Google Scholar] [PubMed]
- Mihatsch, W.A.; Franz, A.R.; Kuhnt, B.; Hogel, J.; Pohlandt, F. Hydrolysis of casein accelerates gastrointestinal transit via reduction of opioid receptor agonists released from casein in rats. Biol. Neonate 2005, 87, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Frohlinde, E.; Schmid, R.; Brantl, V.; Schusdziarra, V. Effect of bovine β-casomorphin-4-amide on gastrointestinal transit and pancreatic endocrine function in man. In Beta-Casomorphins and Related Peptides: Recent Developments; Brantl, V., Teschemacher, H., Eds.; VCH Weinheim: New York, NY, USA, 1994; pp. 155–160. [Google Scholar]
- Daniel, H.; Vohwinkel, M.; Rehner, G. Effect of casein and beta-casomorphins on gastrointestinal motility in rats. J. Nutr. 1990, 120, 252–257. [Google Scholar] [PubMed]
- Ul Haq, M.R.; Kapila, R.; Sharma, R.; Saliganti, V.; Kapila, S. Comparative evaluation of cow β-casein variants (A1/A2) consumption on Th2-mediated inflammatory response in mouse gut. Eur. J. Nutr. 2014, 53, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Pol, O.; Sasaki, M.; Jimenez, N.; Dawson, V.L.; Dawson, T.M.; Puig, M.M. The involvement of nitric oxide in the enhanced expression of μ-opioid receptors during intestinal inflammation in mice. Br. J. Pharmacol. 2005, 145, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J.; Gromada, J. Role of incretin hormones in the regulation of insulin secretion in diabetic and nondiabetic humans. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E199–E206. [Google Scholar] [CrossRef] [PubMed]
- Schirra, J.; Nicolaus, M.; Roggel, R.; Katschinski, M.; Storr, M.; Woerle, H.J.; Goke, B. Endogenous glucagon-like peptide 1 controls endocrine pancreatic secretion andantro-pyloro-duodenal motility in humans. Gut 2006, 55, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claustre, J.; Toumi, F.; Trompette, A.; Jourdan, G.; Guignard, H.; Chayvialle, J.A.; Plaisancié, P. Effects of peptides derived from dietary proteins on mucus secretion in rat jejunum. Am. J. Physiol. Gastrointest Liver Physiol. 2002, 283, G521–G528. [Google Scholar] [CrossRef] [PubMed]
- Elitsur, Y.; Luk, G.D. Beta-casomorphin (BCM) and human colonic lamina propria lymphocyte proliferation. Clin. Exp. Immunol. 1991, 85, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Kayser, H.; Meisel, H. Stimulation of human peripheral blood lymphocytes by bioactive peptides derived from bovine milk proteins. FEBS Lett. 1996, 383, 18–20. [Google Scholar] [CrossRef]
- Ul Haq, M.R.; Kapila, R.; Saliganti, V. Consumption of β-casomorphins-7/5 induce inflammatory immune response in mice gut through Th2 pathway. J. Funct. Foods 2014, 8, 150–160. [Google Scholar] [CrossRef]
- Crowley, E.T.; Williams, L.T.; Roberts, T.K.; Dunstan, R.H.; Jones, P.D. Does milk cause constipation? A crossover dietary trial. Nutrients 2013, 5, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Andiran, F.; Dayi, S.; Mete, E. Cows milk consumption in constipation and anal fissure in infants and young children. J. Paediatr. Child Health 2003, 39, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Iacono, G.; Cavataio, F.; Montalto, G.; Florena, A.; Tumminello, M.; Soresi, M.; Notarbartolo, A.; Carroccio, A. Intolerance of cow’s milk and chronic constipation in children. N. Engl. J. Med. 1998, 339, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Woodford, K.; Kukuljan, S.; Pal, S. Comparative effects of A1 versus A2 beta-casein on gastrointestinal measures: A blinded randomised cross-over pilot study. Eur. J. Clin. Nutr. 2014, 68, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Mourad, F.H.; Barada, K.A.; Bou Rached, N.A.; Khoury, C.I.; Saade, N.E.; Nassar, C.F. Inhibitory effect of experimental colitis on fluid absorption in rat jejunum: Role of the enteric nervous system, VIP, and nitric oxide. Am. J. Physiol. Gastrointest Liver Physiol. 2006, 290, G262–G268. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, U.; Wisel, S.; Coon, S. Neutral Na-amino acid cotransport is differentially regulated by glucocorticoids in the normal and chronically inflamed rabbit small intestine. Am. J. Physiol. Gastrointest Liver Physiol. 2007, 292, G467–G474. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Provot, C.; Persuy, M.A.; Mercier, J.C. Complete nucleotide sequence of ovine β-casein cDNA: Inter-species comparison. Biochimie 1989, 71, 827–832. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, S.; Woodford, K.; Kukuljan, S.; Ho, S. Milk Intolerance, Beta-Casein and Lactose. Nutrients 2015, 7, 7285-7297. https://doi.org/10.3390/nu7095339
Pal S, Woodford K, Kukuljan S, Ho S. Milk Intolerance, Beta-Casein and Lactose. Nutrients. 2015; 7(9):7285-7297. https://doi.org/10.3390/nu7095339
Chicago/Turabian StylePal, Sebely, Keith Woodford, Sonja Kukuljan, and Suleen Ho. 2015. "Milk Intolerance, Beta-Casein and Lactose" Nutrients 7, no. 9: 7285-7297. https://doi.org/10.3390/nu7095339