The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers




{kind=link}
Abstract
:1. Introduction
2. Molecular Aspects of Human Lactase-Phlorizin Hydrolase Protein
2.1. Structure and Function
2.2. Biosynthesis and Trafficking
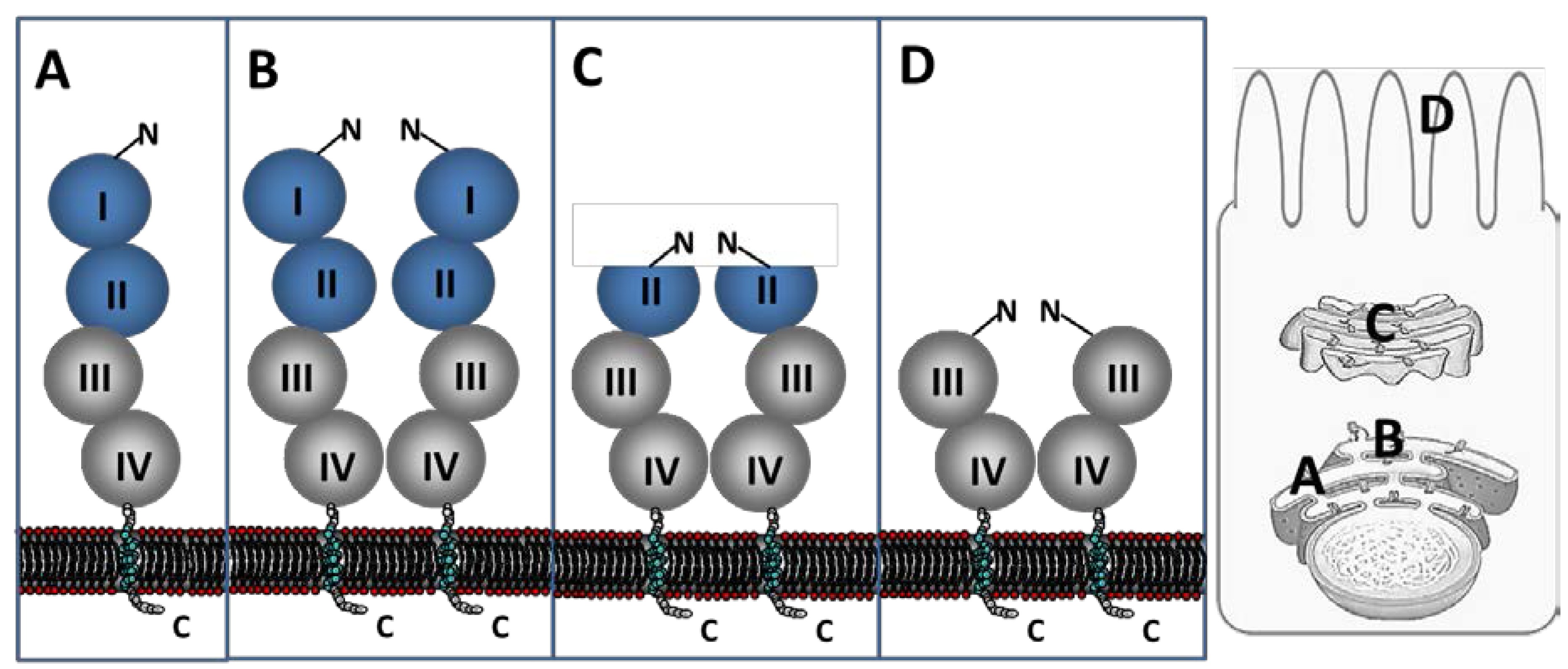
3. Lactose Intolerance
4. Adult Type of Hypolactasia
5. Congenital Lactase Deficiency
6. Secondary Lactose Intolerance Caused by Infections
7. Association of Lactase Persistence and Dairy Consumption with the Incidence of Some Common Cancers
7.1. Colorectal Cancer
7.2. Ovarian and Prostate Cancers
8. Concluding Remarks and Outlook
Conflicts of Interest
References
- Naim, H.Y.; Sterchi, E.E.; Lentze, M.J. Biosynthesis and maturation of lactase-phlorizin hydrolase in the human small intestinal epithelial cells. Biochem. J. 1987, 241, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.Y.; Sterchi, E.E.; Lentze, M.J. Structure, biosynthesis, and glycosylation of human small intestinal maltase-glucoamylase. J. Biol. Chem. 1988, 263, 19709–19717. [Google Scholar] [PubMed]
- Naim, H.Y.; Sterchi, E.E.; Lentze, M.J. Biosynthesis of the human sucrase-isomaltase complex. Differential O-glycosylation of the sucrase subunit correlates with its position within the enzyme complex. J. Biol. Chem. 1988, 263, 7242–7253. [Google Scholar] [PubMed]
- Semenza, G. Anchoring and biosynthesis of stalked brush border membrane proteins: Glycosidases and peptidases of enterocytes and renal tubuli. Annu. Rev. Cell. Biol. 1986, 2, 255–313. [Google Scholar] [CrossRef] [PubMed]
- Triadou, N.; Bataille, J.; Schmitz, J. Longitudinal study of the human intestinal brush border membrane proteins. Distribution of the main disaccharidases and peptidases. Gastroenterology 1983, 85, 1326–1332. [Google Scholar] [PubMed]
- Ferraris, R.P. Dietary and developmental regulation of intestinal sugar transport. Biochem. J. 2001, 360, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Hauri, H.P.; Sterchi, E.E.; Bienz, D.; Fransen, J.A.; Marxer, A. Expression and intracellular transport of microvillus membrane hydrolases in human intestinal epithelial cells. J. Cell Biol. 1985, 101, 838–851. [Google Scholar] [CrossRef] [PubMed]
- Sim, L.; Willemsma, C.; Mohan, S.; Naim, H.Y.; Pinto, B.M.; Rose, D.R. Structural basis for substrate selectivity in human maltase-glucoamylase and sucrase-isomaltase N-terminal domains. J. Biol. Chem. 2010, 285, 17763–17770. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Naim, H.Y. Miglustat-induced intestinal carbohydrate malabsorption is due to the inhibition of α-glucosidases, but not β-galactosidases. J. Inherit. Metab. Dis. 2012, 35, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Zecca, L.; Mesonero, J.E.; Stutz, A.; Poiree, J.C.; Giudicelli, J.; Cursio, R.; Gloor, S.M.; Semenza, G. Intestinal lactase-phlorizin hydrolase (LPH): The two catalytic sites; the role of the pancreas in pro-LPH maturation. FEBS. Lett. 1998, 435, 225–228. [Google Scholar] [CrossRef]
- Colombo, V.; Lorenz-Meyer, H.; Semenza, G. Small intestinal phlorizin hydrolase: The “β-glycosidase complex”. Biochim. Biophys. Acta. 1973, 327, 412–424. [Google Scholar] [CrossRef]
- Sterchi, E.E.; Mills, P.R.; Fransen, J.A.; Hauri, H.P.; Lentze, M.J.; Naim, H.Y.; Ginsel, L.; Bond, J. Biogenesis of intestinal lactase-phlorizin hydrolase in adults with lactose intolerance. Evidence for reduced biosynthesis and slowed-down maturation in enterocytes. J. Clin. Investig. 1990, 86, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Kruse, T.A.; Bolund, L.; Grzeschik, K.H.; Ropers, H.H.; Sjostrom, H.; Noren, O.; Mantei, N.; Semenza, G. The human lactase-phlorizin hydrolase gene is located on chromosome 2. FEBS. Lett. 1988, 240, 123–126. [Google Scholar] [CrossRef]
- Boll, W.; Wagner, P.; Mantei, N. Structure of the chromosomal gene and cdnas coding for lactase-phlorizin hydrolase in humans with adult-type hypolactasia or persistence of lactase. Am. J. Hum. Genet. 1991, 48, 889–902. [Google Scholar] [PubMed]
- Mantei, N.; Villa, M.; Enzler, T.; Wacker, H.; Boll, W.; James, P.; Hunziker, W.; Semenza, G. Complete primary structure of human and rabbit lactase-phlorizin hydrolase: Implications for biosynthesis, membrane anchoring and evolution of the enzyme. EMBO. J. 1988, 7, 2705–2713. [Google Scholar] [PubMed]
- Wacker, H.; Keller, P.; Falchetto, R.; Legler, G.; Semenza, G. Location of the two catalytic sites in intestinal lactase-phlorizin hydrolase. Comparison with sucrase-isomaltase and with other glycosidases, the membrane anchor of lactase-phlorizin hydrolase. J. Biol. Chem. 1992, 267, 18744–18752. [Google Scholar] [PubMed]
- Naim, H.Y.; Lacey, S.W.; Sambrook, J.F.; Gething, M.J. Expression of a full-length cDNA coding for human intestinal lactase-phlorizin hydrolase reveals an uncleaved, enzymatically active, and transport-competent protein. J. Biol. Chem. 1991, 266, 12313–12320. [Google Scholar] [PubMed]
- Hauri, H.P.; Sander, B.; Naim, H. Induction of lactase biosynthesis in the human intestinal epithelial cell line Caco-2. Eur. J. Biochem. 1994, 219, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.; Brewer, C.; Fransen, J.A.; Naim, H.Y. Transport, function, and sorting of lactase-phlorizin hydrolase in madin-darby canine kidney cells. J. Biol. Chem. 1994, 269, 2712–2721. [Google Scholar] [PubMed]
- Buller, H.A.; Montgomery, R.K.; Sasak, W.V.; Grand, R.J. Biosynthesis, glycosylation, and intracellular transport of intestinal lactase-phlorizin hydrolase in rat. J. Biol. Chem. 1987, 262, 17206–17211. [Google Scholar] [PubMed]
- Danielsen, E.M.; Skovbjerg, H.; Noren, O.; Sjostrom, H. Biosynthesis of intestinal microvillar proteins intracellular processing of lactase-phlorizin hydrolase. Biochem. Biophys. Res. Commun. 1984, 122, 82–90. [Google Scholar] [CrossRef]
- Naim, H.Y. Processing and transport of human small intestinal lactase-phlorizin hydrolase (LPH). Role of N-linked oligosaccharide modification. FEBS. Lett. 1994, 342, 302–307. [Google Scholar] [CrossRef]
- Danielsen, E.M. Biosynthesis of intestinal microvillar proteins. Dimerization of aminopeptidase N and lactase-phlorizin hydrolase. Biochemistry 1990, 29, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.Y.; Naim, H. Dimerization of lactase-phlorizin hydrolase occurs in the endoplasmic reticulum, involves the putative membrane spanning domain and is required for an efficient transport of the enzyme to the cell surface. Eur. J. Cell Biol. 1996, 70, 198–208. [Google Scholar] [PubMed]
- Jacob, R.; Weiner, J.R.; Stadge, S.; Naim, H.Y. Additional N-glycosylation and its impact on the folding of intestinal lactase-phlorizin hydrolase. J. Biol. Chem. 2000, 275, 10630–10637. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.Y.; Lentze, M.J. Impact of o-glycosylation on the function of human intestinal lactase-phlorizin hydrolase. Characterization of glycoforms varying in enzyme activity and localization of O-glycoside addition. J. Biol. Chem. 1992, 267, 25494–25504. [Google Scholar] [PubMed]
- Jacob, R.; Peters, K.; Naim, H.Y. The prosequence of human lactase-phlorizin hydrolase modulates the folding of the mature enzyme. J. Biol. Chem. 2002, 277, 8217–8225. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.Y.; Jacob, R.; Naim, H.; Sambrook, J.F.; Gething, M.J. The pro region of human intestinal lactase-phlorizin hydrolase. J. Biol. Chem. 1994, 269, 26933–26943. [Google Scholar] [PubMed]
- Naim, H.Y. The pro-region of human intestinal lactase-phlorizin hydrolase is enzymatically inactive towards lactose. Biol. Chem. Hoppe. Seyler. 1995, 376, 255–258. [Google Scholar] [PubMed]
- Behrendt, M.; Polaina, J.; Naim, H.Y. Structural hierarchy of regulatory elements in the folding and transport of an intestinal multidomain protein. J. Biol. Chem. 2010, 285, 4143–4152. [Google Scholar] [CrossRef] [PubMed]
- Stoops, E.H.; Caplan, M.J. Trafficking to the apical and basolateral membranes in polarized epithelial cells. J. Am. Soc. Nephrol. 2014, 25, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, E.M. Involvement of detergent-insoluble complexes in the intracellular transport of intestinal brush border enzymes. Biochemistry 1995, 34, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, E.; Simons, K. Protein and lipid sorting from the trans-golgi network to the plasma membrane in polarized cells. Semin. Cell. Dev. Biol. 1998, 9, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Alfalah, M.; Jacob, R.; Preuss, U.; Zimmer, K.P.; Naim, H.; Naim, H.Y. O-linked glycans mediate apical sorting of human intestinal sucrase-isomaltase through association with lipid rafts. Curr. Biol. 1999, 9, 593–596. [Google Scholar] [CrossRef]
- Jacob, R.; Alfalah, M.; Grunberg, J.; Obendorf, M.; Naim, H.Y. Structural determinants required for apical sorting of an intestinal brush-border membrane protein. J. Biol. Chem. 2000, 275, 6566–6572. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.; Parczyk, K.; Leutz, A.; Kayne, M.; Kondor-Koch, C. Constitutive apical secretion of an 80-kD sulfated glycoprotein complex in the polarized epithelial Madin-Darby canine kidney cell line. J. Cell. Biol. 1987, 105, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.; Preuss, U.; Panzer, P.; Alfalah, M.; Quack, S.; Roth, M.G.; Naim, H.; Naim, H.Y. Hierarchy of sorting signals in chimeras of intestinal lactase-phlorizin hydrolase and the influenza virus hemagglutinin. J. Biol. Chem. 1999, 274, 8061–8067. [Google Scholar] [CrossRef] [PubMed]
- Kundu, A.; Avalos, R.T.; Sanderson, C.M.; Nayak, D.P. Transmembrane domain of influenza virus neuraminidase, a type ii protein, possesses an apical sorting signal in polarized MDCK cells. J. Virol. 1996, 70, 6508–6515. [Google Scholar] [PubMed]
- Panzer, P.; Preuss, U.; Joberty, G.; Naim, H.Y. Protein domains implicated in intracellular transport and sorting of lactase-phlorizin hydrolase. J. Biol. Chem. 1998, 273, 13861–13869. [Google Scholar] [CrossRef] [PubMed]
- Grunberg, J.; Luginbuhl, U.; Sterchi, E.E. Proteolytic processing of human intestinal lactase-phlorizin hydrolase precursor is not a prerequisite for correct sorting in madin darby canine kidney (MDCK) cells. FEBS. Lett. 1992, 314, 224–228. [Google Scholar] [CrossRef]
- Jacob, R.; Zimmer, K.P.; Naim, H.; Naim, H.Y. The apical sorting of lactase-phlorizin hydrolase implicates sorting sequences found in the mature domain. Eur. J. Cell Biol. 1997, 72, 54–60. [Google Scholar] [PubMed]
- Jacob, R.; Naim, H.Y. Apical membrane proteins are transported in distinct vesicular carriers. Curr. Biol. 2001, 11, 1444–1450. [Google Scholar] [CrossRef]
- Jacob, R.; Heine, M.; Alfalah, M.; Naim, H.Y. Distinct cytoskeletal tracks direct individual vesicle populations to the apical membrane of epithelial cells. Curr. Biol. 2003, 13, 607–612. [Google Scholar] [CrossRef]
- Delacour, D.; Koch, A.; Jacob, R. The role of galectins in protein trafficking. Traffic 2009, 10, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Delacour, D.; Cramm-Behrens, C.I.; Drobecq, H.; le Bivic, A.; Naim, H.Y.; Jacob, R. Requirement for galectin-3 in apical protein sorting. Curr. Biol. 2006, 16, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Kuokkanen, M.; Enattah, N.S.; Oksanen, A.; Savilahti, E.; Orpana, A.; Jarvela, I. Transcriptional regulation of the lactase-phlorizin hydrolase gene by polymorphisms associated with adult-type hypolactasia. Gut 2003, 52, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Kuokkanen, M.; Kokkonen, J.; Enattah, N.S.; Ylisaukko-Oja, T.; Komu, H.; Varilo, T.; Peltonen, L.; Savilahti, E.; Jarvela, I. Mutations in the translated region of the lactase gene (LCT) underlie congenital lactase deficiency. Am. J. Hum. Genet. 2006, 78, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Robayo-Torres, C.C.; Nichols, B.L. Molecular differentiation of congenital lactase deficiency from adult-type hypolactasia. Nutr. Rev. 2007, 65, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Heyman, M.B. Lactose intolerance in infants, children, and adolescents. Pediatrics 2006, 118, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Lasekan, J.B.; Jacobs, J.; Reisinger, K.S.; Montalto, M.B.; Frantz, M.P.; Blatter, M.M. Lactose-free milk protein-based infant formula: Impact on growth and gastrointestinal tolerance in infants. Clin. Pediatr. (Phila) 2011, 50, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Antonowicz, I.; Lebenthal, E. Developmental pattern of small intestinal enterokinase and disaccharidase activities in the human fetus. Gastroenterology 1977, 72, 1299–1303. [Google Scholar] [PubMed]
- Erasmus, H.D.; Ludwig-Auser, H.M.; Paterson, P.G.; Sun, D.; Sankaran, K. Enhanced weight gain in preterm infants receiving lactase-treated feeds: A randomized, double-blind, controlled trial. J. Pediatr. 2002, 141, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Scrimshaw, N.S. Global and regional consequences of occult hunger. Arch. Latinoam. Nutr. 1994, 44, 75S–84S. [Google Scholar] [PubMed]
- Sahi, T. Genetics and epidemiology of adult-type hypolactasia. Scand. J. Gastroenterol. Suppl. 1994, 202, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Simoons, F.J. Primary adult lactose intolerance and the milking habit: A problem in biologic and cultural interrelations. Am. J. Dig. Dis. 1970, 15, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Sahi, T. Hypolactasia and lactase persistence. Historical review and the terminology. Scand. J. Gastroenterol. Suppl. 1994, 202, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.B.; Wang, Y.; Hughes, L.A.; Swallow, D.M.; Thurrell, W.P.; Sams, V.R.; Barton, R.; Lanzon-Miller, S.; Sarner, M. Studies on the expression of intestinal lactase in different individuals. Gut 1995, 36, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Enattah, N.S.; Sahi, T.; Savilahti, E.; Terwilliger, J.D.; Peltonen, L.; Jarvela, I. Identification of a variant associated with adult-type hypolactasia. Nat. Genet. 2002, 30, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Olds, L.C.; Sibley, E. Lactase persistence DNA variant enhances lactase promoter activity in vitro: Functional role as a cis regulatory element. Hum. Mol. Genet. 2003, 12, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.B.; Waud, J.P.; Roberts, A.G.; Campbell, A.K. Systemic lactose intolerance: A new perspective on an old problem. Postgrad. Med. J. 2005, 81, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Lewinsky, R.H.; Jensen, T.G.; Moller, J.; Stensballe, A.; Olsen, J.; Troelsen, J.T. T-13910 DNA variant associated with lactase persistence interacts with Oct-1 and stimulates lactase promoter activity in vitro. Hum. Mol. Genet. 2005, 14, 3945–3953. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.G.; Liebert, A.; Lewinsky, R.; Swallow, D.M.; Olsen, J.; Troelsen, J.T. The -14010*C variant associated with lactase persistence is located between an Oct-1 and HNF 1α binding site and increases lactase promoter activity. Hum. Genet. 2011, 130, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Tishkoff, S.A.; Reed, F.A.; Ranciaro, A.; Voight, B.F.; Babbitt, C.C.; Silverman, J.S.; Powell, K.; Mortensen, H.M.; Hirbo, J.B.; Osman, M.; et al. Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet. 2007, 39, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ingram, C.J.; Elamin, M.F.; Mulcare, C.A.; Weale, M.E.; Tarekegn, A.; Raga, T.O.; Bekele, E.; Elamin, F.M.; Thomas, M.G.; Bradman, N.; et al. A novel polymorphism associated with lactose tolerance in Africa: Multiple causes for lactase persistence? Hum. Genet. 2007, 120, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.; Sequeira, F.; Luiselli, D.; Beleza, S.; Rocha, J. On the edge of bantu expansions: MtDNA, Y chromosome and lactase persistence genetic variation in southwestern Angola. BMC Evol. Biol. 2009, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Ingram, C.J.; Raga, T.O.; Tarekegn, A.; Browning, S.L.; Elamin, M.F.; Bekele, E.; Thomas, M.G.; Weale, M.E.; Bradman, N.; Swallow, D.M. Multiple rare variants as a cause of a common phenotype: Several different lactase persistence associated alleles in a single ethnic group. J. Mol. Evol. 2009, 69, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Gallego Romero, I.; Basu Mallick, C.; Liebert, A.; Crivellaro, F.; Chaubey, G.; Itan, Y.; Metspalu, M.; Eaaswarkhanth, M.; Pitchappan, R.; Villems, R.; et al. Herders of Indian and European cattle share their predominant allele for lactase persistence. Mol. Biol. Evol. 2012, 29, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Torniainen, S.; Parker, M.I.; Holmberg, V.; Lahtela, E.; Dandara, C.; Jarvela, I. Screening of variants for lactase persistence/non-persistence in populations from south Africa and Ghana. BMC Genet. 2009, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, D.C.; Santos, S.E.; Ribeiro-dos-Santos, A.K.; Hutz, M.H. Several different lactase persistence associated alleles and high diversity of the lactase gene in the admixed Brazilian population. PLoS ONE 2012, 7, e46520. [Google Scholar] [CrossRef] [PubMed]
- Vesa, T.H.; Marteau, P.; Korpela, R. Lactose intolerance. J. Am. Coll. Nutr. 2000, 19, 165S–175S. [Google Scholar] [CrossRef] [PubMed]
- Malagelada, J.R. Lactose intolerance. N. Engl. J. Med. 1995, 333, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Montes, R.; Monge, E. Lactose malabsorption and IBS. Scand. J. Gastroenterol 2004, 39, 1033. [Google Scholar] [CrossRef] [PubMed]
- Bolin, T. IBS or intolerance? Aust. Fam. Physician 2009, 38, 962–965. [Google Scholar] [PubMed]
- Wackerbauer, R.; Schmidt, T. Symbolic dynamics of jejunal motility in the irritable bowel. Chaos 1999, 9, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Arola, H. Diagnosis of hypolactasia and lactose malabsorption. Scand. J. Gastroenterol. Suppl. 1994, 202, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Forget, P.; Lombet, J.; Grandfils, C.; Dandrifosse, G.; Geubelle, F. Lactase insufficiency revisited. J. Pediatr. Gastroenterol. Nutr. 1985, 4, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, A.D.; McGill, D.B. Lactose tolerance tests in adults with normal lactase activity. Gastroenterology 1966, 50, 340–346. [Google Scholar] [PubMed]
- Rasinpera, H.; Savilahti, E.; Enattah, N.S.; Kuokkanen, M.; Totterman, N.; Lindahl, H.; Jarvela, I.; Kolho, K.L. A genetic test which can be used to diagnose adult-type hypolactasia in children. Gut 2004, 53, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Wilt, T.J.; Shaukat, A.; Shamliyan, T.; Taylor, B.C.; MacDonald, R.; Tacklind, J.; Rutks, I.; Schwarzenberg, S.J.; Kane, R.L.; Levitt, M. Lactose intolerance and health. Evid. Rep. Technol. Assess (Full Rep.) 2010, 192, 1–410. [Google Scholar] [PubMed]
- Kull, M.; Kallikorm, R.; Lember, M. Impact of molecularly defined hypolactasia, self-perceived milk intolerance and milk consumption on bone mineral density in a population sample in Northern Europe. Scand. J. Gastroenterol. 2009, 44, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Suchy, F.J.; Brannon, P.M.; Carpenter, T.O.; Fernandez, J.R.; Gilsanz, V.; Gould, J.B.; Hall, K.; Hui, S.L.; Lupton, J.; Mennella, J.; et al. National institutes of health consensus development conference: Lactose intolerance and health. Ann. Intern. Med. 2010, 152, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Lifshitz, F. Congenital lactase deficiency. J. Pediatr. 1966, 69, 229–237. [Google Scholar] [CrossRef]
- Savilahti, E.; Launiala, K.; Kuitunen, P. Congenital lactase deficiency. A clinical study on 16 patients. Arch. Dis. Child. 1983, 58, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, M.; Keiser, M.; Hoch, M.; Naim, H.Y. Impaired trafficking and subcellular localization of a mutant lactase associated with congenital lactase deficiency. Gastroenterology 2009, 136, 2295–2303. [Google Scholar] [CrossRef] [PubMed]
- Torniainen, S.; Freddara, R.; Routi, T.; Gijsbers, C.; Catassi, C.; Hoglund, P.; Savilahti, E.; Jarvela, I. Four novel mutations in the lactase gene (LCT) underlying congenital lactase deficiency (CLD). BMC Gastroenterol. 2009, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Uchida, N.; Sakamoto, O.; Irie, M.; Abukawa, D.; Takeyama, J.; Kure, S.; Tsuchiya, S. Two novel mutations in the lactase gene in a Japanese infant with congenital lactase deficiency. Tohoku. J. Exp. Med. 2012, 227, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, L.; Pfeiffer, K.; Naim, H.Y. Congenital lactose intolerance is triggered by severe mutations on both alleles of the lactase gene. BMC Gastroenterol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, W.; Kaczmarek, S.; Kirschstein, M.; Santer, R. A novel mutation within the lactase gene (LCT): The first report of congenital lactase deficiency diagnosed in central Europe. BMC Gastroenterol. 2015, 15, 90. [Google Scholar] [CrossRef] [PubMed]
- Gendrel, D.; Richard-Lenoble, D.; Kombila, M.; Dupont, C.; Moreno, J.L.; Gendrel, C.; Nardou, M.; Chaussain, M. Influence of intestinal parasitism on lactose absorption in well-nourished African children. Am. J. Trop. Med. Hyg. 1992, 46, 137–140. [Google Scholar] [PubMed]
- Beau, I.; Cotte-Laffitte, J.; Geniteau-Legendre, M.; Estes, M.K.; Servin, A.L. An NSP4-dependant mechanism by which rotavirus impairs lactase enzymatic activity in brush border of human enterocyte-like Caco-2 cells. Cell. Microbiol. 2007, 9, 2254–2266. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.M.; Mitchell, D.M.; Gibbons, T.F.; Parr, R.D. Rotavirus NSP4: A multifunctional viral enterotoxin. Viral. Immun. 2005, 18, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Berkova, Z.; Crawford, S.E.; Blutt, S.E.; Morris, A.P.; Estes, M.K. Expression of rotavirus NSP4 alters the actin network organization through the actin remodeling protein cofilin. J. Virol. 2007, 81, 3545–3553. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Parsons, D.A.; Staempfli, H.R. Association of clostridium difficile with enterocolitis and lactose intolerance in a foal. J. Am. Vet. Med. Assoc. 1999, 214, 229–232. [Google Scholar] [PubMed]
- Kwak, Y.K.; Vikstrom, E.; Magnusson, K.E.; Vecsey-Semjen, B.; Colque-Navarro, P.; Mollby, R. The staphylococcus aureus alpha-toxin perturbs the barrier function in Caco-2 epithelial cell monolayers by altering junctional integrity. Infect. Immun. 2012, 80, 1670–1680. [Google Scholar] [CrossRef] [PubMed]
- Miko, B.A.; Uhlemann, A.C.; Gelman, A.; Lee, C.J.; Hafer, C.A.; Sullivan, S.B.; Shi, Q.; Miller, M.; Zenilman, J.; Lowy, F.D. High prevalence of colonization with Staphylococcus aureus clone USA 300 at multiple body sites among sexually transmitted disease clinic patients: An unrecognized reservoir. Microbes. Infect. 2012, 14, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Hess, D.J.; Henry-Stanley, M.J.; Erickson, E.A.; Wells, C.L. Intracellular survival of Staphylococcus aureus within cultured enterocytes. J. Surg. Res. 2003, 114, 42–49. [Google Scholar] [CrossRef]
- Michael, L.; Timothy, W.; Aasma, S. Clinical implications of lactose malabsorption versus lactose intolerance. J. Clin. Gastroenterol. 2013, 47, 471. [Google Scholar]
- Dallas, M.S. Genetics of lactase persistence and lactose intolerance. Genetics 2003, 37, 197–219. [Google Scholar]
- Virginie, A.; Patrick, C.E.; Christiane, L.-A.; Jean-Marc, B.; François, B.; Robert, B.; Daniel, T.; Anne-Marie, D. Lactose malabsorption and colonic fermentations alter host metabolism in rats. Br. J. Nutr. 2013, 110, 625–631. [Google Scholar]
- Suarez, F.L.; Savaiano, D.A.; Levitt, M.D. Review article the treatment of lactose intolerance. Aliment. Pharmacol. Ther. 1995, 9, 589–597. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Priebe, M.G.; Harmsen, H.J.; Stellaard, F.; Sun, X.; Welling, G.W.; Vonk, R.J. Colonic fermentation may play a role in lactose intolerance in humans. J. Nutr. 2006, 136, 58–63. [Google Scholar] [PubMed]
- Benjamin, M.; Daniel, P.; Heiko, F.; Michael, F.; Stephan, R.V.; Mark, F. Lactose malabsorption and intolerance: Pathogenesis, diagnosis and treatment. United Eur. Gastroent. J. 2013, 1, 151–159. [Google Scholar]
- Sanchez, B.; GavilÁN, D.; Margolles, A.; Gueimonde, M. Probiotic fermented milks: Present and future. Int. J. Dairy Technol. 2009, 62, 472–483. [Google Scholar] [CrossRef]
- Robert, T.G.; Taylor, M.; Sherry, B.; Phyllis, A.W. Cancer statistics, 2000. CA. Cancer. J. Clin. 2000, 50, 7–33. [Google Scholar]
- Kuokkanen, M.; Butzow, R.; Rasinpera, H.; Medrek, K.; Nilbert, M.; Malander, S.; Lubinski, J.; Jarvela, I. Lactase persistence and ovarian carcinoma risk in Finland, Poland and Sweden. Int. J. Cancer 2005, 117, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Peter, R.H. New insights into calcium, dairy and colon cancer. World J. Gastroenterol. 2008, 14, 4429–4433. [Google Scholar]
- Ma, R.W.L.; Chapman, K. A systematic review of the effect of diet in prostate cancer prevention and treatment. J. Hum. Nutr. Diet. 2009, 22, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.J.; Thompson, P.A. Diet and nutrient factors in colorectal cancer risk. Nutr. Clin. Pract. 2012, 27, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Raúl Baena, R.; Pedro Salinas, H. Diet and cancer: Risk factors and epidemiological evidence. Maturitas 2014, 77, 202208. [Google Scholar]
- Rock, C.L. Milk and the risk and progression of cancer. Nestle. Nutr. Workshop Ser. Pediatr. Program 2011, 67, 173–185. [Google Scholar] [PubMed]
- Glade, M.J. World Cancer Research Fund/American Institute for Cancer Research. Food, Nutrition, Physical Activity and the Prevention of Cancer: A Global Perspective. Nutrition 2008, 24, 393–398. [Google Scholar] [CrossRef]
- Renate, C.H.B.; Renate, M.W.; Jacoba, M.S.R.; Lea, K.; Anne-Claire, B.O.L.; Ettje, F.T.; Doris, S.M.C.; Teresa, N.; Ellen, K. Dietary supplement use and colorectal cancer risk: A systematic review and meta-analyses of prospective cohort studies. Int. J. Cancer 2015, 136, 2388–2401. [Google Scholar]
- Van der Meer, R.; Lapré, J.A.; Govers, M.J.; Kleibeuker, J.H. Mechanisms of the intestinal effects of dietary fats and milk products on colon carcinogenesis. Cancer Lett. 1997, 114, 75–83. [Google Scholar] [CrossRef]
- Govers, M.J.; Termont, D.S.; Lapré, J.A.; Kleibeuker, J.H.; Vonk, R.J.; van der Meer, R. Calcium in milk products precipitates intestinal fatty acids and secondary bile acids and thus inhibits colonic cytotoxicity in humans. Cancer Res. 1996, 56, 3270–3275. [Google Scholar] [PubMed]
- Rasinperä, H.; Forsblom, C.; Enattah, N.S.; Halonen, P.; Salo, K.; Victorzon, M.; Mecklin, J.P.; Järvinen, H.; Enholm, S.; Sellick, G.; et al. The c/c−13910 genotype of adult-type hypolactasia is associated with an increased risk of colorectal cancer in the Finnish population. Gut 2005, 54, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Krisztián, B.; Erika, H.; János, P.K.; Henrik, H.; Áron, L.; Péter, L.L.; Bernadett, B.; Barna, B.; Péter, L.; Gábor, S. Effects of the lactase 13910 c/t and calcium-sensor receptor a986s g/t gene polymorphisms on the incidence and recurrence of colorectal cancer in Hungarian population. BMC Cancer 2008, 8, 317. [Google Scholar]
- Ada, P.; Enrico, S.; Angela, M.; Annamaria, G.; Rosa, C.; Michele, Q.; Antonio, M.; Mauro, C.; Paolo Usai, S.; Francesco, P. Genotyping of the lactase-phlorizin hydrolase c/t-13910 polymorphism by means of a new rapid denaturing high-performance liquid chromatography-based assay in healthy subjects and colorectal cancer patients. J. Biomol. Screen. 2007, 12, 733–739. [Google Scholar]
- Elena, T.; Paola, P.; Elisabetta, B.; Giovanni, C.A.; Gianfranco, T.; Luciana, F.; Mohammad, A.; Francesca, C.; Nicola, L.; Giovanni, S.; et al. Lct-13910c > t polymorphism-associated lactose malabsorption and risk for colorectal cancer in Italy. Dig. Liver Dis. 2010, 42, 741743. [Google Scholar]
- Szilagyi, A.; Nathwani, U.; Vinokuroff, C.; Correa, J.A.A.; Shrier, I. Evaluation of relationships among national colorectal cancer mortality rates, genetic lactase non-persistence status, and per capita yearly milk and milk product consumption. Nutr. Cancer 2006, 55, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Nathwani, U.; Vinokuroff, C.; Correa, J.A.A.; Shrier, I. The effect of lactose maldigestion on the relationship between dairy food intake and colorectal cancer: A systematic review. Nutr. Cancer 2006, 55, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Martha, L.S.; Carol, S.; Maureen, M.; Khe-Ni, M.; Bette, J.C.; John, D.P.; Roger, W. Associations between vitamin D, vitamin D receptor gene and the androgen receptor gene with colon and rectal cancer. Int. J. Cancer 2006, 118, 3140–3146. [Google Scholar]
- Christakos, S.; Dhawan, P.; Porta, A.; Mady, L.J.; Seth, T. Vitamin D and intestinal calcium absorption. Mol. Cell. Endocrinol. 2011, 347, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Andrew, S.; Henry, L.; Barry, B.; Xue, X.Q. Latitude, sunshine, and human lactase phenotype distributions may contribute to geographic patterns of modern disease: The inflammatory bowel disease model. Clin. Epidemiol. 2014, 6. [Google Scholar]
- George Davey, S.; Debbie, A.L.; Nic, J.T.; Jamil, B.; Matt, K.; Ian, N.M.D.; Shah, E. Lactase persistence-related genetic variant: Population substructure and health outcomes. Eur. J. Hum. Genet. 2008, 17, 357–367. [Google Scholar]
- Andrew, S. Determinants of geographic patterns of diseases: Interaction of lactose/lactase status and sunshine exposure. Med. Hypotheses 2010, 75, 466–470. [Google Scholar]
- Orel, R.; Trop, T.K. Intestinal microbiota, probiotics and prebiotics in inflammatory bowel disease. World J. Gastroenterol. 2014, 20. [Google Scholar] [CrossRef] [PubMed]
- Shiby, V.K.; Mishra, H.N. Fermented milks and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Peter, W.P. Dairy product consumption and the risk of prostate cancer. Int. Dairy J. 2009, 19, 551–565. [Google Scholar]
- Herrinton, L.J.; Weiss, N.S.; Beresford, S.A.; Stanford, J.L.; Wolfla, D.M.; Feng, Z.; Scott, C.R. Lactose and galactose intake and metabolism in relation to the risk of epithelial ovarian cancer. Am. J. Epidemiol. 1995, 141, 407–416. [Google Scholar] [PubMed]
- Gian Franco, M.; Carla, C.; Carlo La, V.; Giuseppina, R.; Maria Chiara, M.; Guido, A.; Pier Luigi, C. Lactose absorption in patients with ovarian cancer. Am. J. Epidemiol. 1999, 150, 183–186. [Google Scholar]
- Fathalla, M.F. Incessant ovulation—A factor in ovarian neoplasia? Lancet 1971, 2, 163. [Google Scholar] [CrossRef]
- Cramer, D.W.; Welch, W.R. Determinants of ovarian cancer risk. II. Inferences regarding pathogenesis. J. Natl. Cancer Inst. 1983, 71, 717–721. [Google Scholar] [PubMed]
- Kathleen, M.F.; David, J.H.; Graham, A.C.; Charles, S.F.; Daniel, W.C.; Frank, E.S.; Walter, C.W.; Susan, E.H. A prospective study of dietary lactose and ovarian cancer. Int. J. Cancer 2004, 110, 271–277. [Google Scholar]
- Cramer, D.W. Lactase persistence and milk consumption as determinants of ovarian cancer risk. Am. J. Epidemiol. 1989, 130, 904–910. [Google Scholar] [PubMed]
- Koralek, D.O.; Bertone-Johnson, E.R.; Leitzmann, M.F.; Sturgeon, S.R.; Lacey, J.V.; Schairer, C.; Schatzkin, A. Relationship between calcium, lactose, vitamin D, and dairy products and ovarian cancer. Nutr. Cancer 2006, 56, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Torniainen, S.; Hedelin, M.; Autio, V.; Rasinpera, H.; Balter, K.A.; Klint, A.; Bellocco, R.; Wiklund, F.; Stattin, P.; Ikonen, T.; et al. Lactase persistence, dietary intake of milk, and the risk for prostate cancer in Sweden and Finland. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 956–961. [Google Scholar] [CrossRef] [PubMed]
- Ruth, C.T.; Paul, N.A.; Afshan, S.; Naomi, E.A.; Rudolf, K.; Federico, C.; Silke, F.; Anne, T.; Nina Føns, J.; Kim, O.; et al. Genetic variation in the lactase gene, dairy product intake and risk for prostate cancer in the European prospective investigation into cancer and nutrition. Int. J. Cancer 2013, 132, 1901–1910. [Google Scholar]
- Mayank mohan, A.; Satyavati, V.R.; Arup kumar, M.; Sunita, M.; Niranjan, K.; Santosh, K.; Naveen chandra, A.; Shrawan kumar, S. Lactose intolerance in prostate cancer patients: Incidence and associated factors. Scand. J. Gastroenterol. 2008, 43, 270276. [Google Scholar]
- June, M.C.; Crystal, N.H.; Michael, F.L.; Eric, B.R.; Walter, C.W.; Meir, J.S.; Edward, L.G. Diet after diagnosis and the risk of prostate cancer progression, recurrence, and death (United States). Cancer Causes Control 2005, 17, 199–208. [Google Scholar]
- Parodi, A.; Dulbecco, P.; Savarino, E.; Giannini, E.G.; Bodini, G.; Corbo, M.; Isola, L.; De Conca, S.; Marabotto, E.; Savarino, V. Positive glucose breath testing is more prevalent in patients with IBS-like symptoms compared with controls of similar age and gender distribution. J. Clin. Gastroenterol 2009, 43, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Davaasambuu, G.; Akio, S. The possible role of female sex hormones in milk from pregnant cows in the development of breast, ovarian and corpus uteri cancers. Med. Hypotheses 2005, 65, 1028–1037. [Google Scholar]
- Qin, L.; Xu, J.; Wang, P.; Tong, J.; Hoshi, K. Milk consumption is a risk factor for prostate cancer in western countries: Evidence from cohort studies. Asia Pac. J. Clin. Nutr. 2007, 16, 467–476. [Google Scholar] [PubMed]
- Qin, L.Q.; Wang, P.Y.; Takashi, K.; Kazuhiko, H.; Akio, S. Estrogen: One of the risk factors in milk for prostate cancer. Med. Hypotheses 2004, 62, 133–142. [Google Scholar] [CrossRef]
- Shi, W.F.; Jeffrey, S.B. Estrogen plays a critical role in AAV2-mediated gene transfer in ovarian cancer. Acta Pharmacol. Sin. 2008, 29, 1440–1450. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Peter, S.; Aldert, A.B. Naturally occurring estrogens in processed milk and in raw milk (from gestated cows). J. Agri. Food Chem. 2006, 54, 9785–9791. [Google Scholar]
- John, V.; Terry, E.; Penny, K.-E.; Joan, B.; Steven, D.; Robin, S.; Daniel, G.; Roger, C.; Michael, M.; Matthew, L. Survey of retail milk composition as affected by label claims regarding farm-management practices. J. Am. Diet. Assoc. 2008, 108, 1198–1203. [Google Scholar]
- Daniel, W.F.; Xia, X.; Timothy, D.V. Quantitative measurement of endogenous estrogen metabolites, risk-factors for development of breast cancer, in commercial milk products by lc-ms/ms. J. Chromatogr. B. Analyt. Technol. Biomed. Life. Sci. 2009, 877, 1327–1334. [Google Scholar]
- Pape-Zambito, D.A.; Magliaro, A.L.; Kensinger, R.S. 17β-estradiol and estrone concentrations in plasma and milk during bovine pregnancy. J. Dairy Sci. 2008, 91, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Pape-Zambito, D.A.; Roberts, R.F.; Kensinger, R.S. Estrone and 17β-estradiol concentrations in pasteurized-homogenized milk and commercial dairy products. J. Dairy Sci. 2010, 93, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Furnari, C.; Maroun, D.; Gyawali, S.; Snyder, B.W.; Davis, A.M. Lack of biologically active estrogens in commercial cow milk. J. Dairy Sci. 2012, 95, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Peter, W.P. Impact of cows’ milk estrogen on cancer risk. Int. Dairy J. 2012, 22, 314. [Google Scholar]
- Arthur, C.O.; Seppo, S.; Erika, I. Probiotics: An overview of beneficial effects. Antonie Van Leeuwenhoek 2002, 82, 279–289. [Google Scholar]
- Johnson, I.T. New approaches to the role of diet in the prevention of cancers of the alimentary tract. Mutat. Res. 2004, 551, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Lourdes, S.L.; Adrián, H.M.; Hugo, S.G.; Verónica, M.H.; Belinda, V.C.; Aarón, F.G.C. The effects of consuming probiotic fermented milk on the immune system: A review of scientific evidence. Int. J. Dairy Technol. 2015, 68, 153–165. [Google Scholar]
- Jarvis, J.K.; Miller, G.D. Overcoming the barrier of lactose intolerance to reduce health disparities. J. Natl. Med. Assoc. 2002, 94, 55–66. [Google Scholar] [PubMed]
- Money, M.E.; Camilleri, M. Review: Management of postprandial diarrhea syndrome. Am. J. Med. 2012, 125, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Austin, G.L.; Dalton, C.B.; Hu, Y.; Morris, C.B.; Hankins, J.; Weinland, S.R.; Westman, E.C.; Yancy, W.S., Jr.; Drossman, D.A. A very low-carbohydrate diet improves symptoms and quality of life in diarrhea-predominant irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2009, 7, 706–708. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Priebe, M.G.; Zhong, Y.; Huang, C.; Harmsen, H.J.; Raangs, G.C.; Antoine, J.M.; Welling, G.W.; Vonk, R.J. Effects of yogurt and bifidobacteria supplementation on the colonic microbiota in lactose-intolerant subjects. J. Appl. Microbiol. 2008, 104, 595–604. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amiri, M.; Diekmann, L.; Von Köckritz-Blickwede, M.; Naim, H.Y. The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers. Nutrients 2015, 7, 7209-7230. https://doi.org/10.3390/nu7095332
Amiri M, Diekmann L, Von Köckritz-Blickwede M, Naim HY. The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers. Nutrients. 2015; 7(9):7209-7230. https://doi.org/10.3390/nu7095332
Chicago/Turabian StyleAmiri, Mahdi, Lena Diekmann, Maren Von Köckritz-Blickwede, and Hassan Y. Naim. 2015. "The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers" Nutrients 7, no. 9: 7209-7230. https://doi.org/10.3390/nu7095332
APA StyleAmiri, M., Diekmann, L., Von Köckritz-Blickwede, M., & Naim, H. Y. (2015). The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers. Nutrients, 7(9), 7209-7230. https://doi.org/10.3390/nu7095332