Consumption of Polyphenol-Rich Zingiber Zerumbet Rhizome Extracts Protects against the Breakdown of the Blood-Retinal Barrier and Retinal Inflammation Induced by Diabetes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extraction
2.2. Measurement of Total Phenolic Content
2.3. Measurement of Total Flavonoids
2.4. Generation of Diabetic Rat Model
2.5. Treatment Protocols
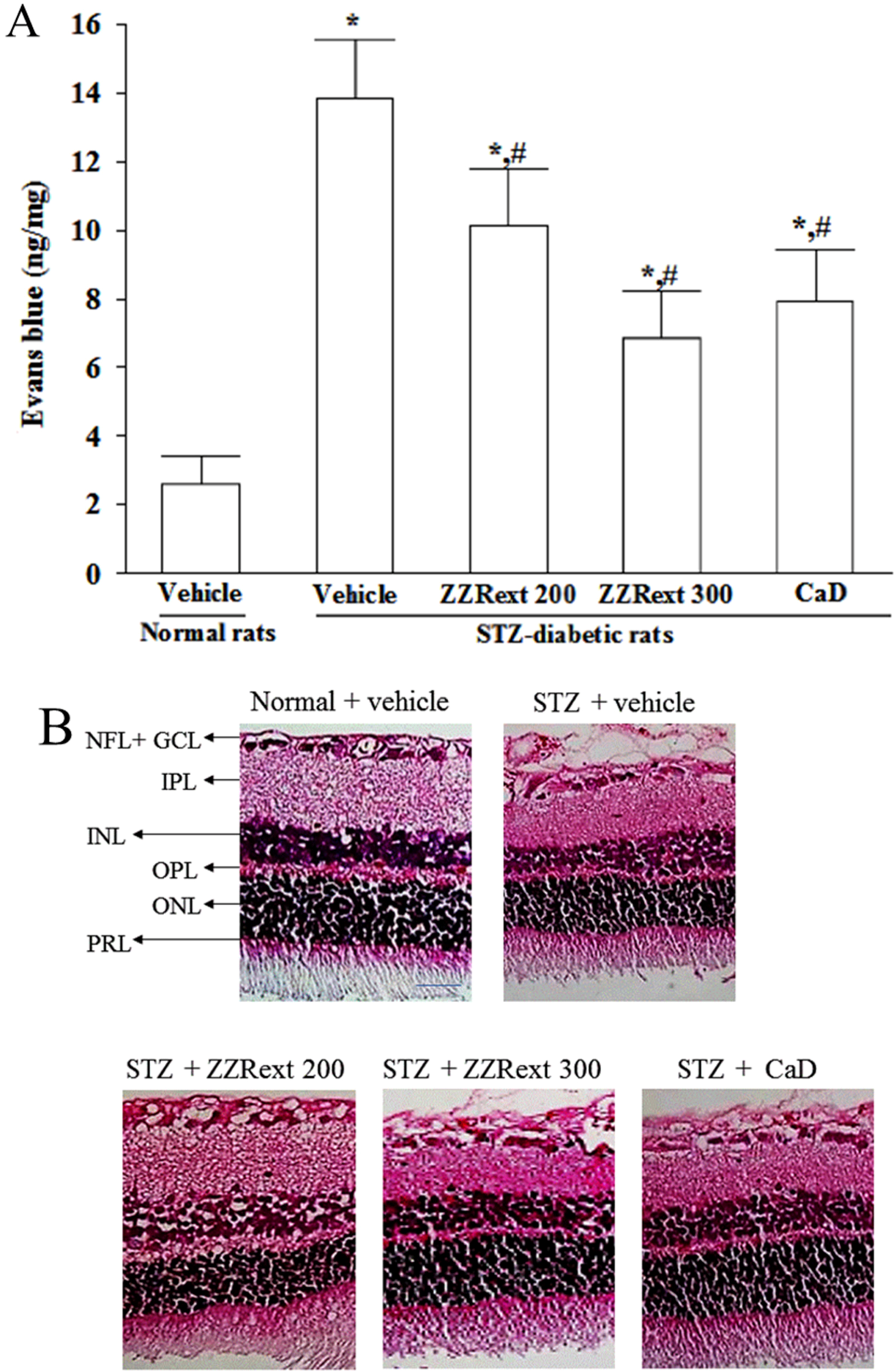
2.6. Measurement of BRB Permeability
2.7. Retinal Cytokines and Adhesion Molecules Determination
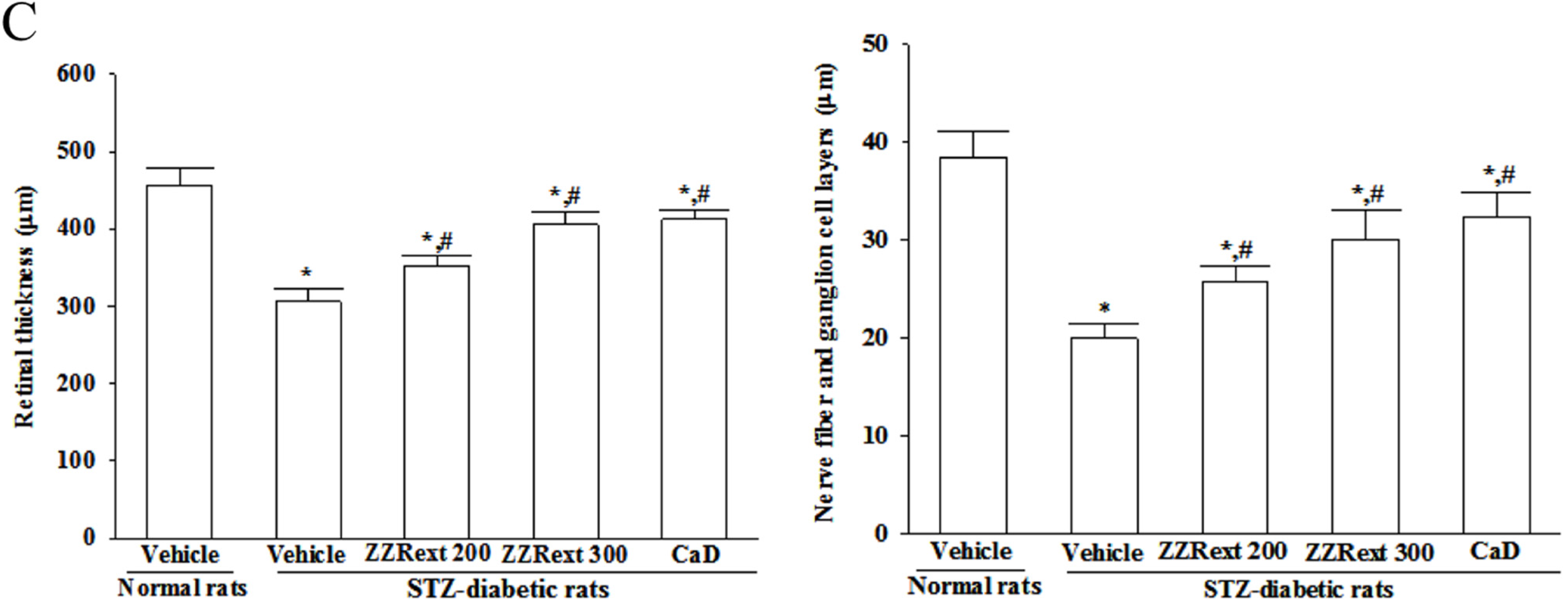
2.8. Light Microscopy and Morphometric Analysis
2.9. Cytoplasmic and Nuclear Extraction
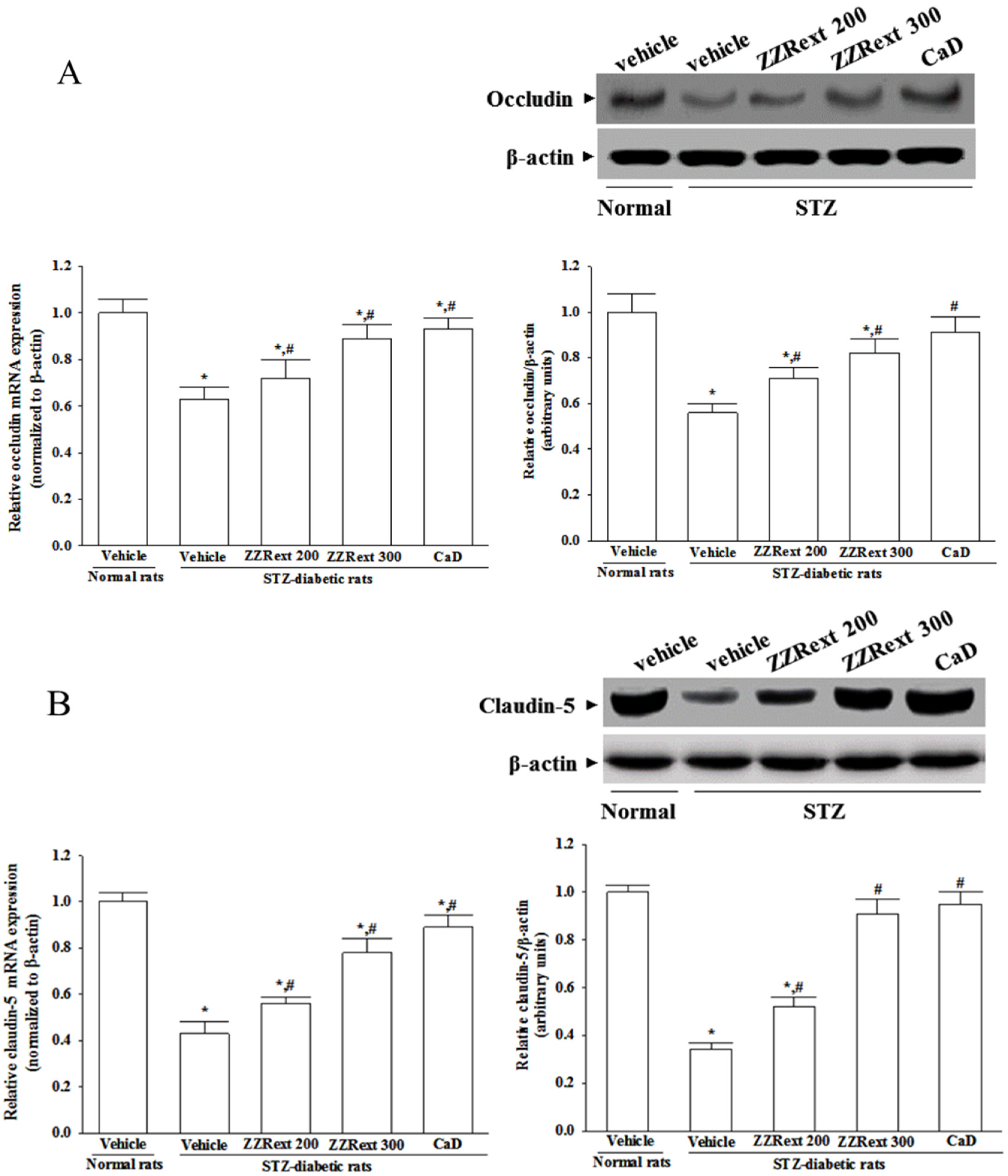
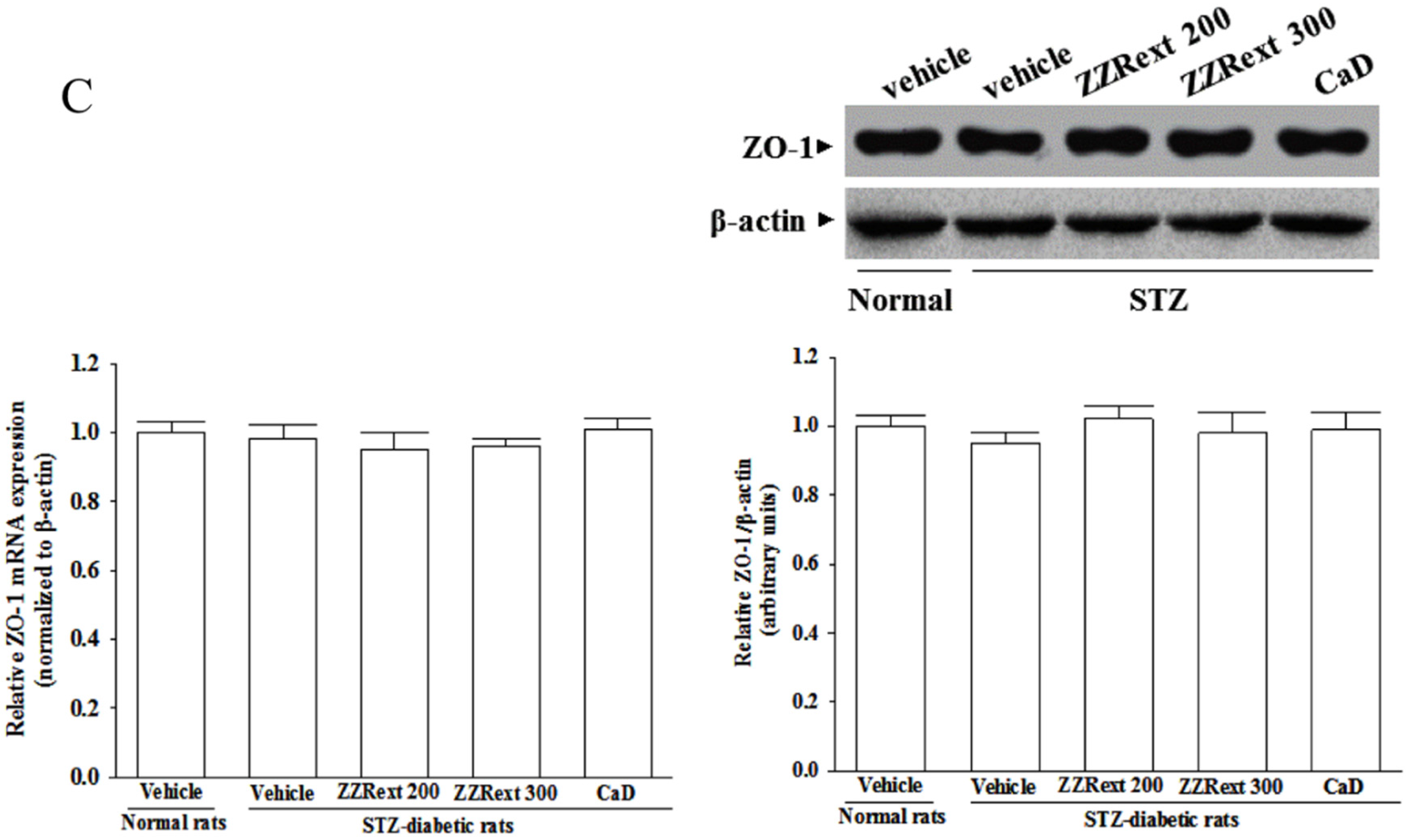
2.10. Western Blot Analysis
2.11. Real-Time Polymerase Chain Reaction (PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target gene | Primers | Sequence |
|---|---|---|
| TNFα | FP | ACACCATGAGCACGGAAAGC |
| RP | CCGCCACGAGCAGGAA | |
| IL-1β | FP | AATGGACAGAACATAAGCCAACA |
| RP | CCCAAGGCCACAGGGAT | |
| IL-6 | FP | GTTGCCTTCTTGGGACTGATG |
| RP | ATACTGGTCTGTTGTGGGTGGT | |
| VEGF | FP | ACAGGGAAGACAATGGGATGA |
| RP | GGGCCAGGGATGGGTTT | |
| ICAM-1 | FP | CGGGTTTGGGCTTCTCC |
| RP | GCCACTGCTCGTCCACATAG | |
| VCAM-1 | FP | ATCTTCGGAGCCTCAACGG |
| RP | CCAATCTGAGCGAGCGTTT | |
| β-actin | FP | TGTGATGGTGGGAATGGGTCAG |
| RP | TTTGATGTCACGCACGATTTCC |
2.12. Quantification of NF-κB Activation
2.13. Statistical Analysis
3. Results
3.1. Total Phenol and Flavonoid Contents of ZZRext
3.2. Status of Body Weight, Blood Glucose, and Glycosylated Hemoglobin
| Groups | Body weight (g/rat) | Plasma glucose (mg/dL) | HbAlc (%) |
|---|---|---|---|
| Normal rats | |||
| vehicle-treated | 337.1 ± 11.9 | 95.6 ± 5.7 | 4.9 ± 0.9 |
| STZ-diabetic rats | |||
| vehicle-treated | 217.1 ± 13.9 * | 423.3 ± 10.3 * | 14.2 ± 1.2 * |
| ZZRext 200-treated | 265.4 ± 10.6 *,# | 341.4 ± 12.4 *,# | 11.9 ± 1.0 *,# |
| ZZRext 300-treated | 285.9 ± 12.7 *,# | 285.3 ± 11.6 *,# | 10.3 ± 0.8 *,# |
| CaD-treated | 270.6 ± 14.5 *,# | 401.2 ± 12.9 * | 13.6 ± 1.3 * |
3.3. Retinal Vascular Permeability
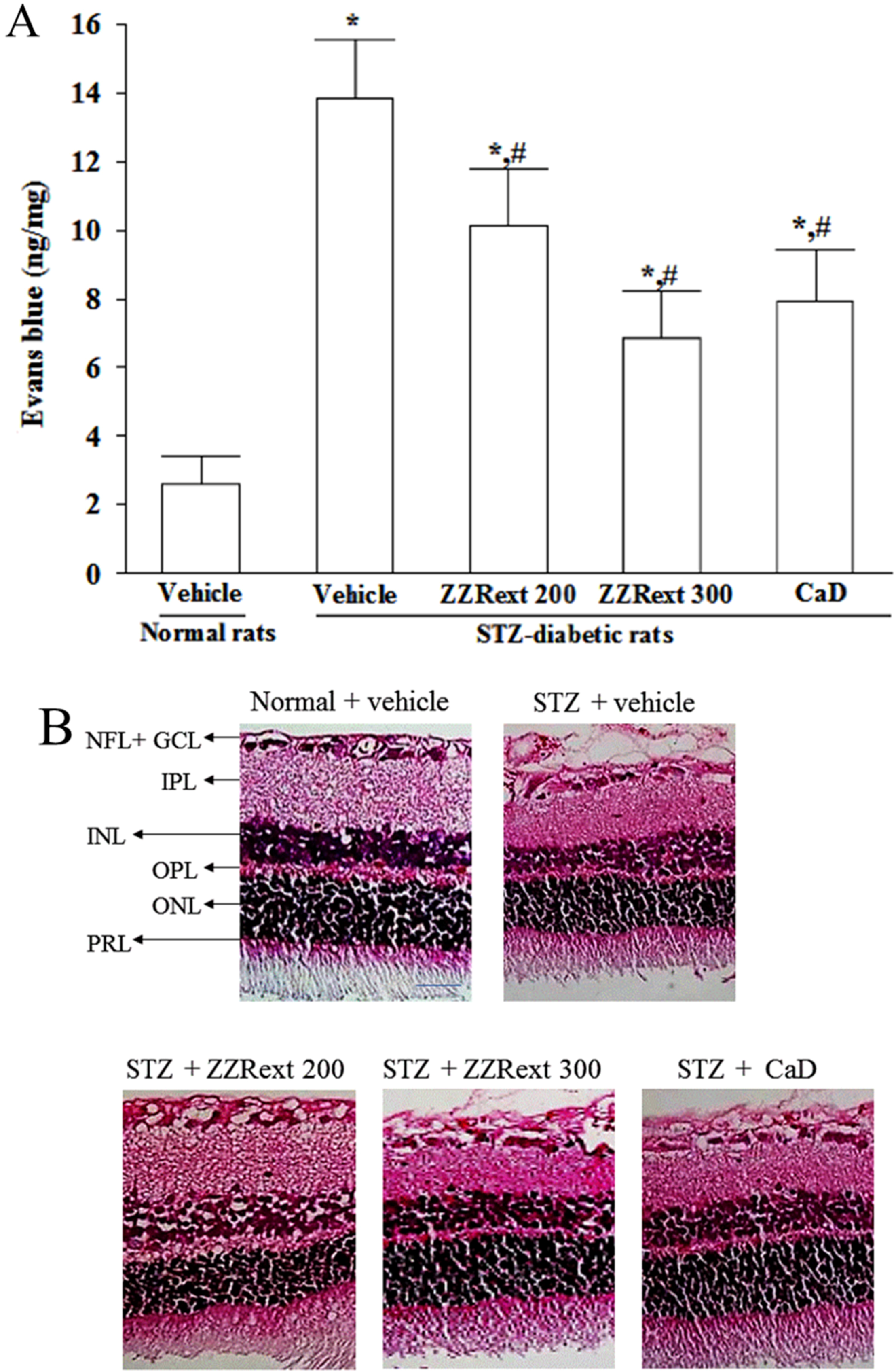
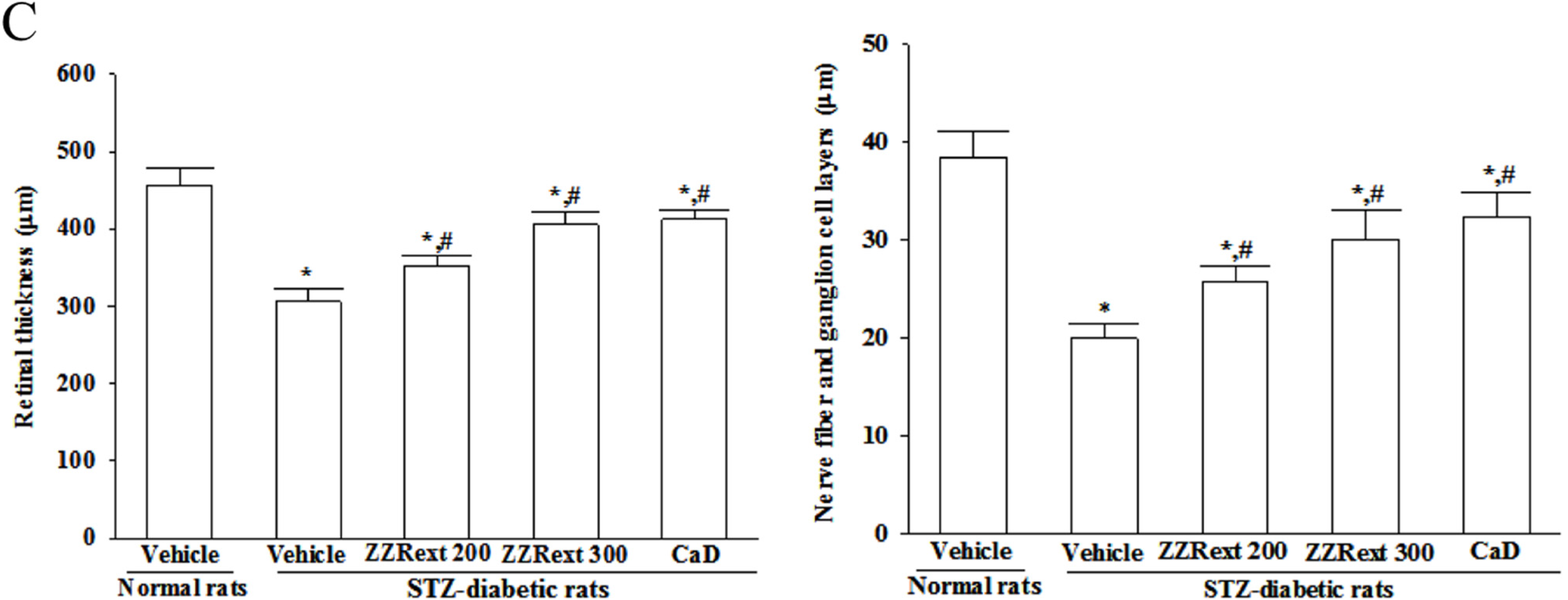
3.4. Light Microscopy and Morphometric Analysis
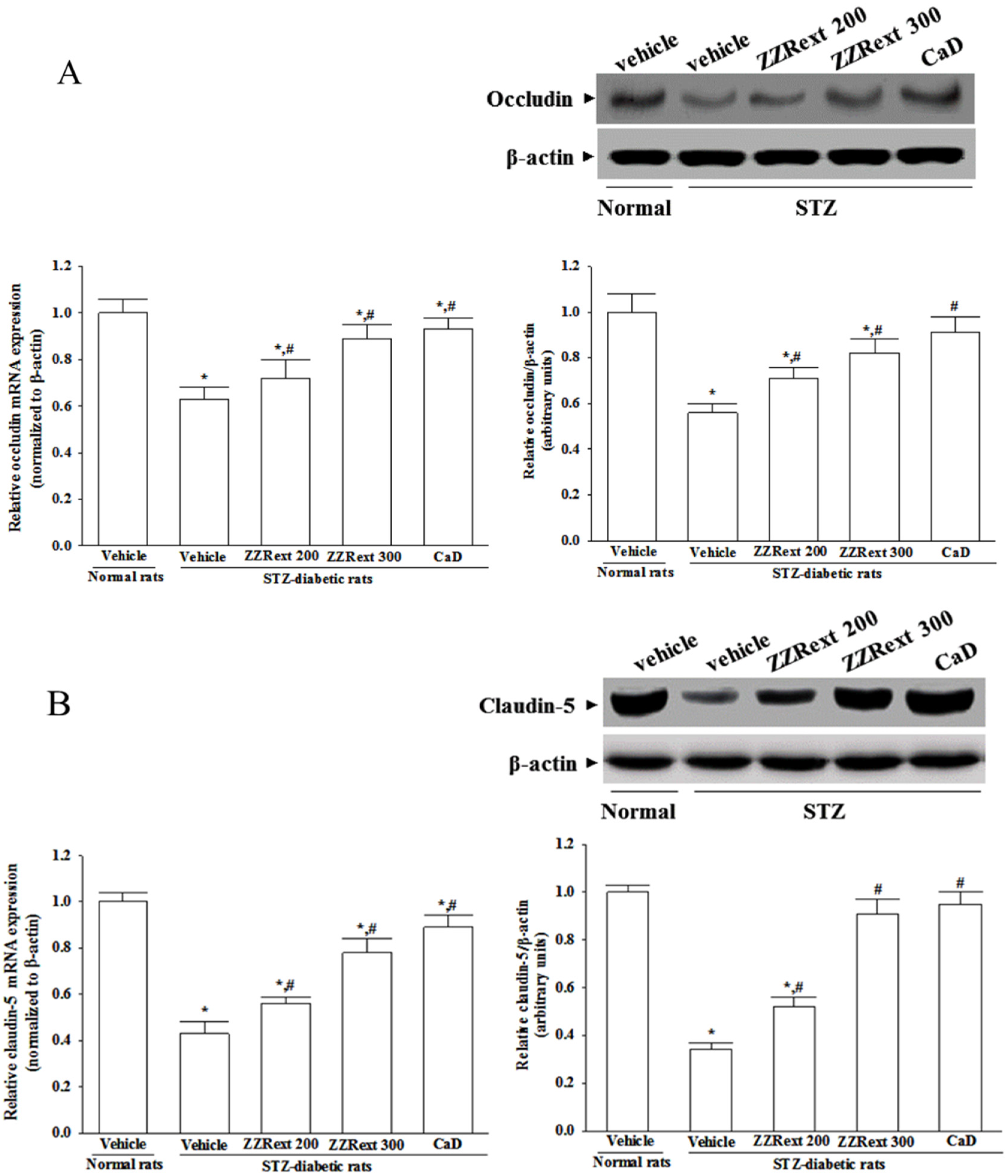
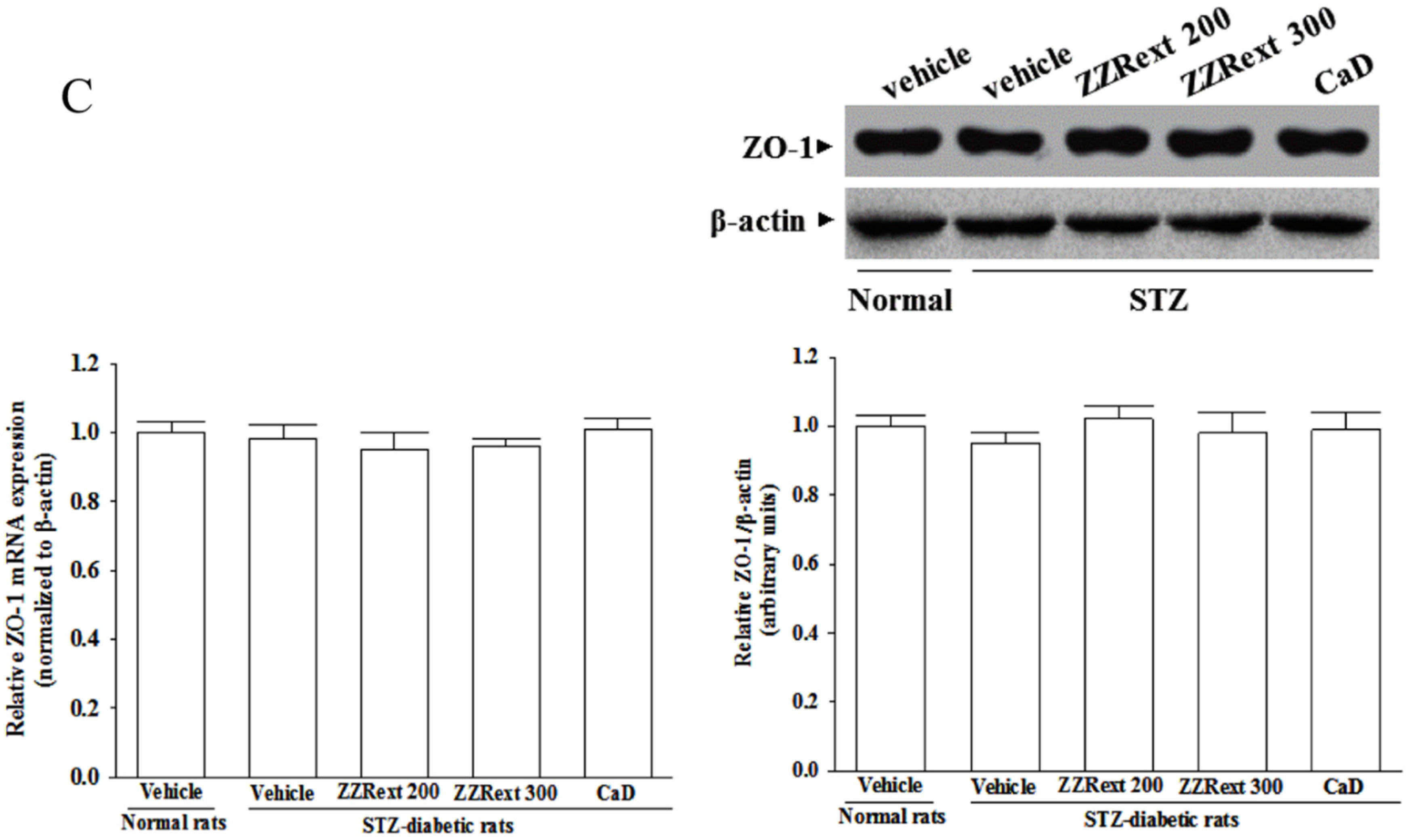
3.5. Retinal mRNA and Protein Expressions of Tight Junction Proteins
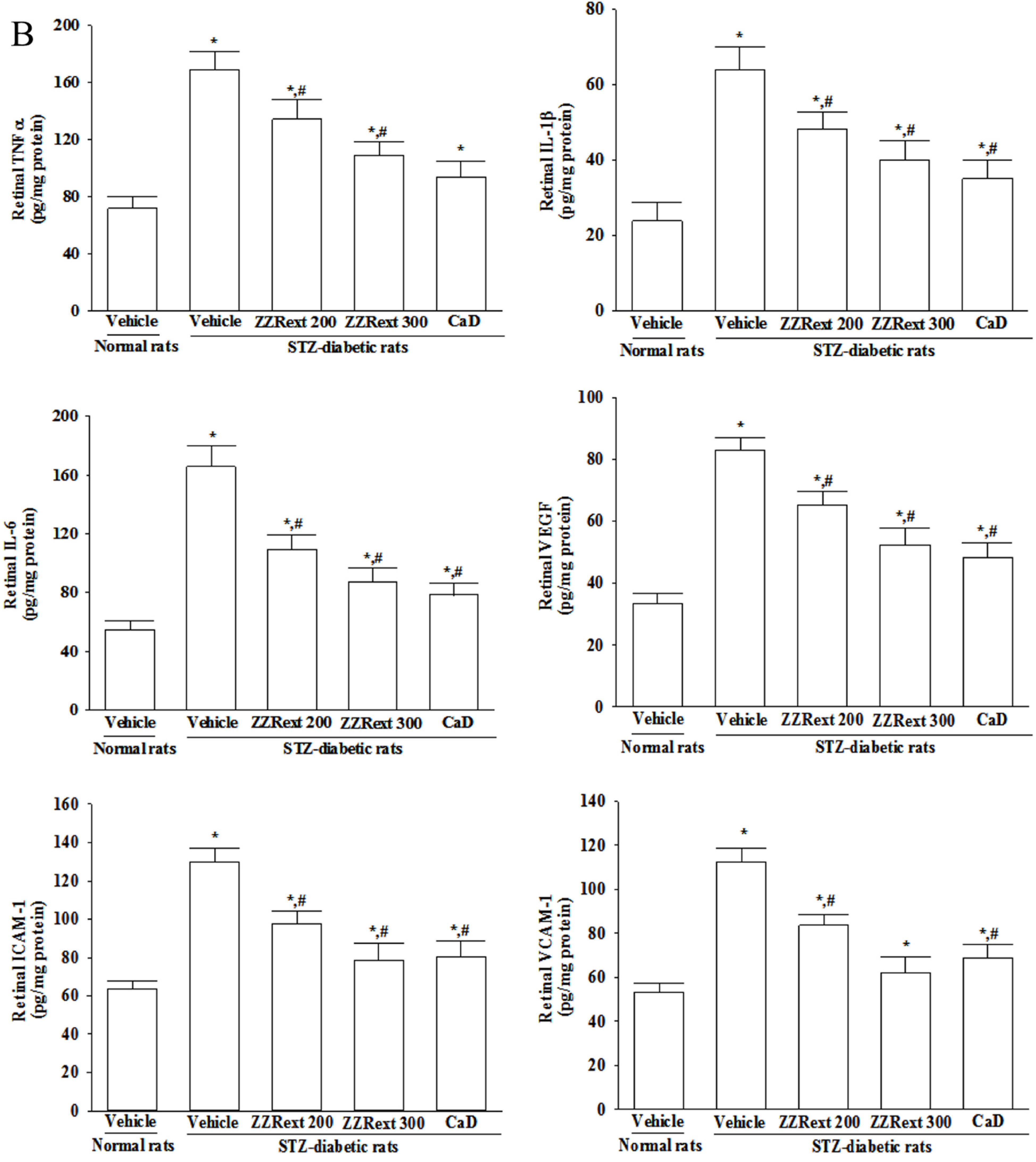
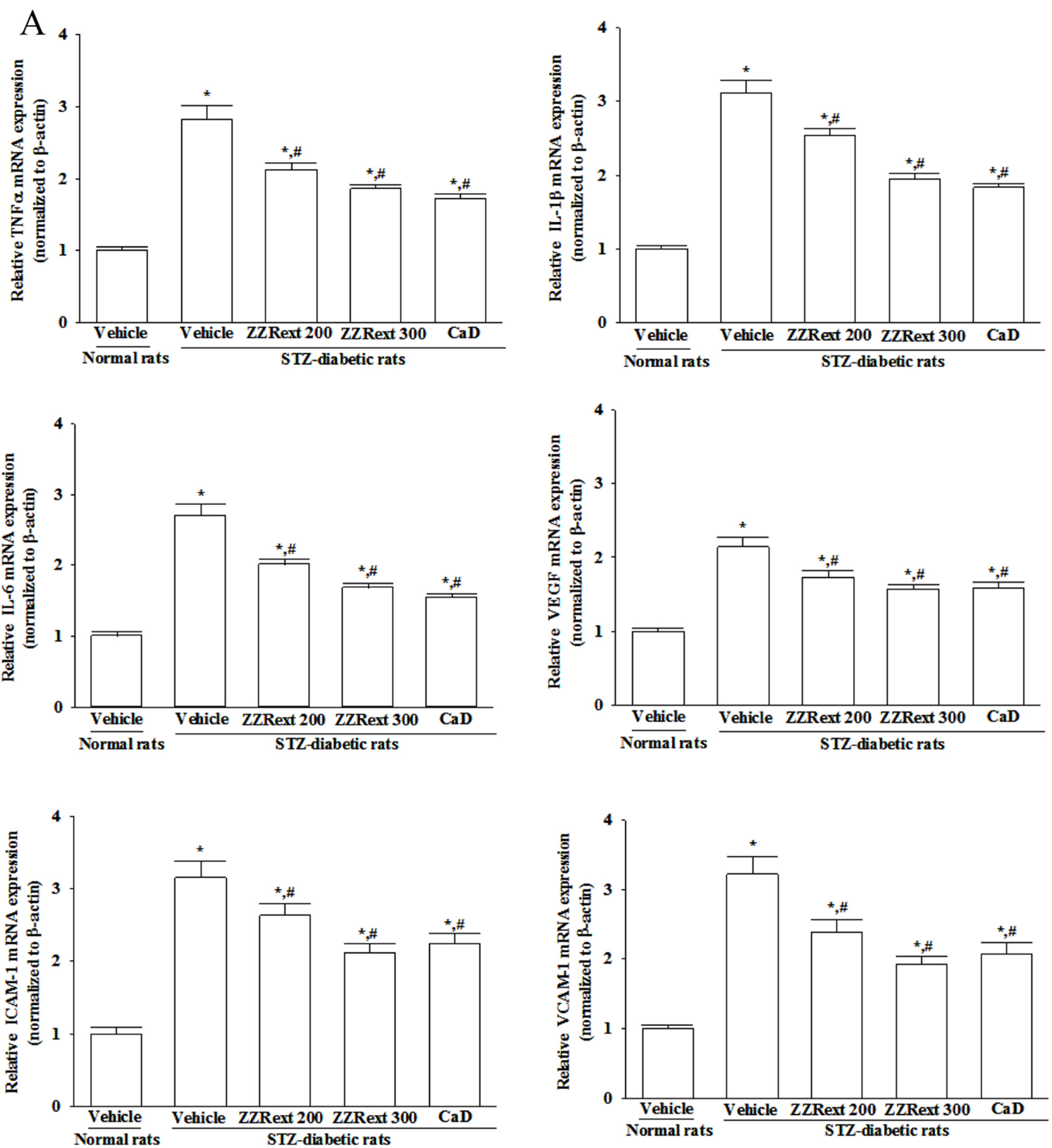
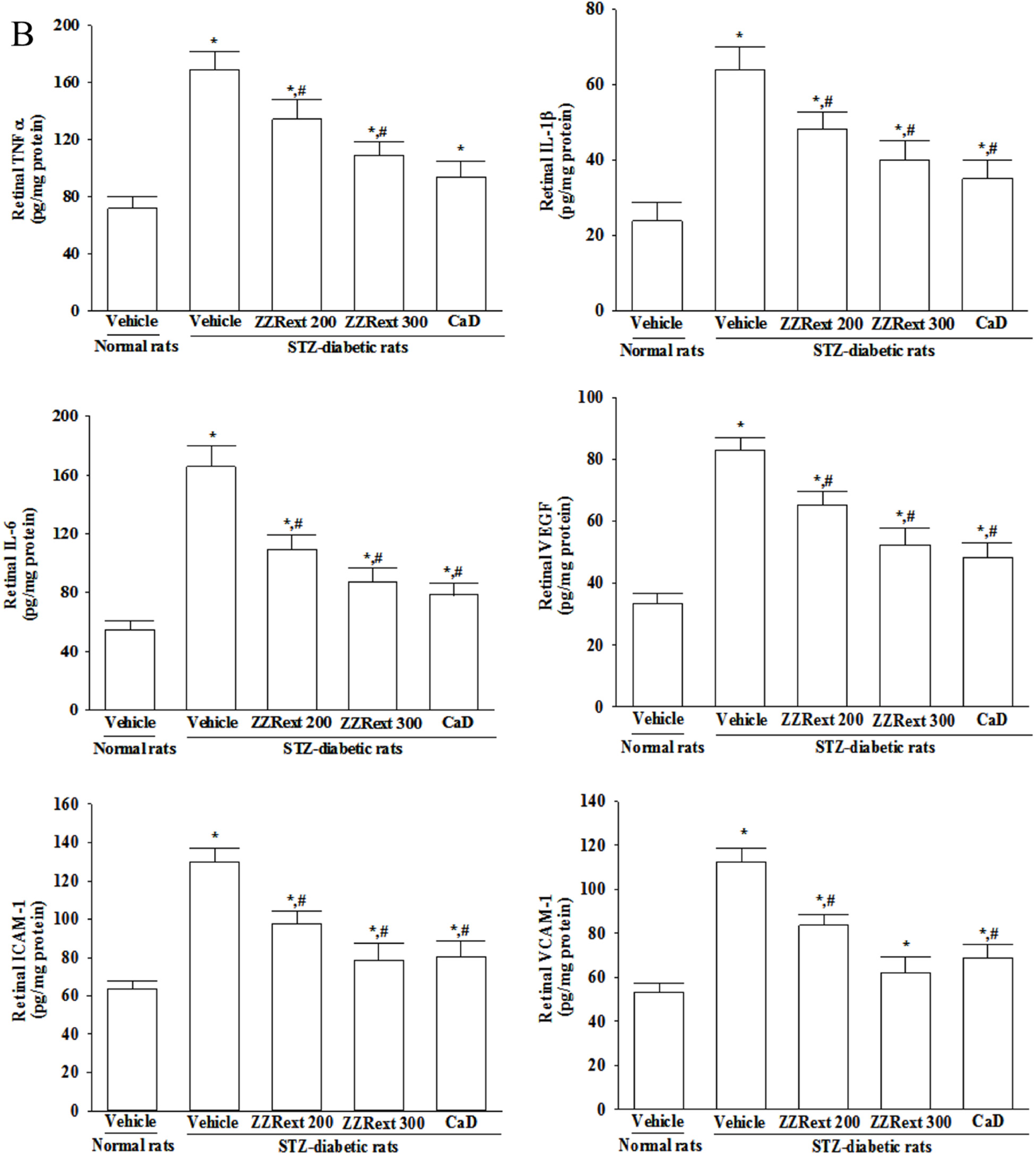
3.6. Retinal mRNA and Protein Expressions of Inflammatory Cytokines and Adhesion Molecules
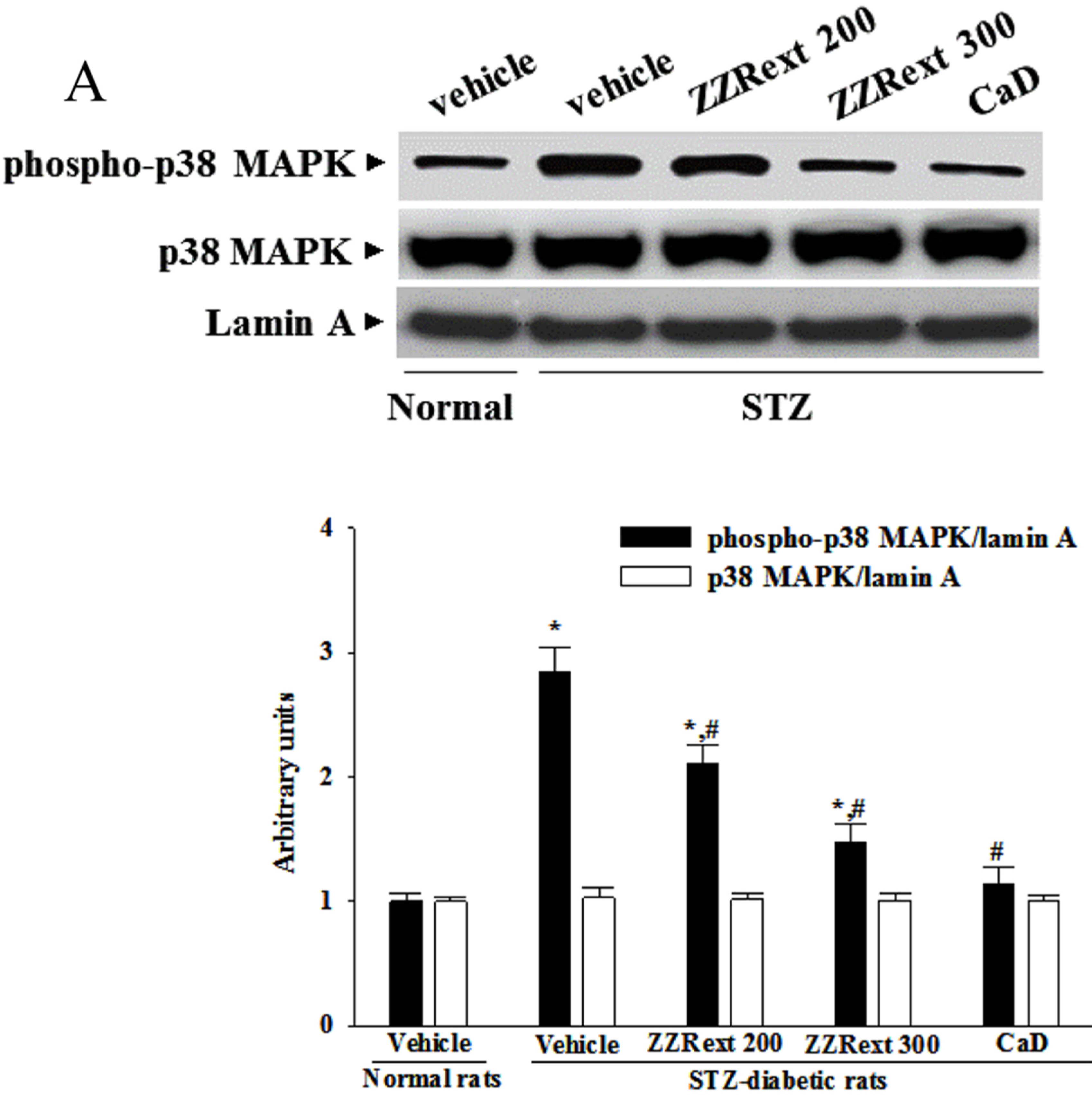
3.7. The Activation of p38 MAPK in Retina

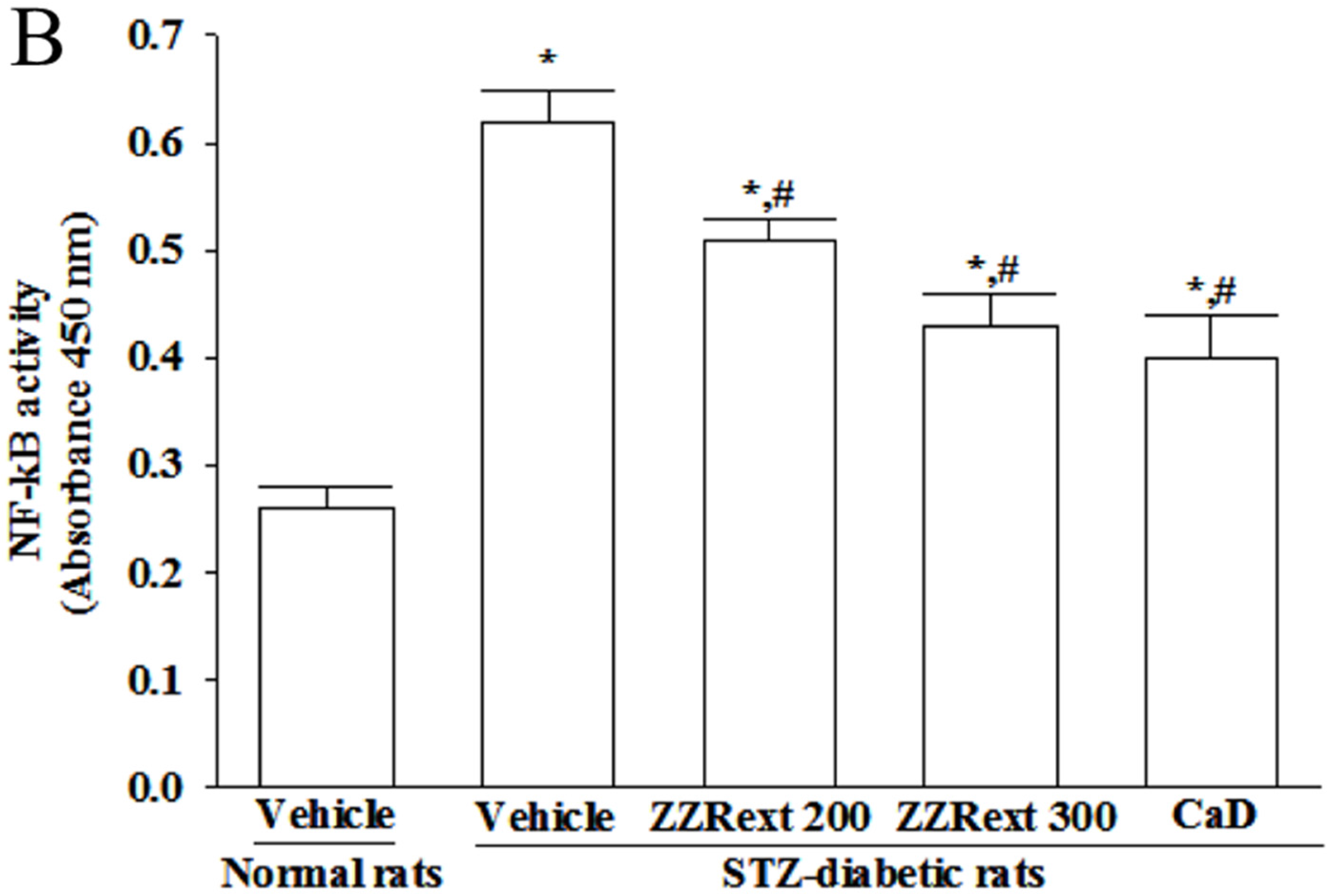
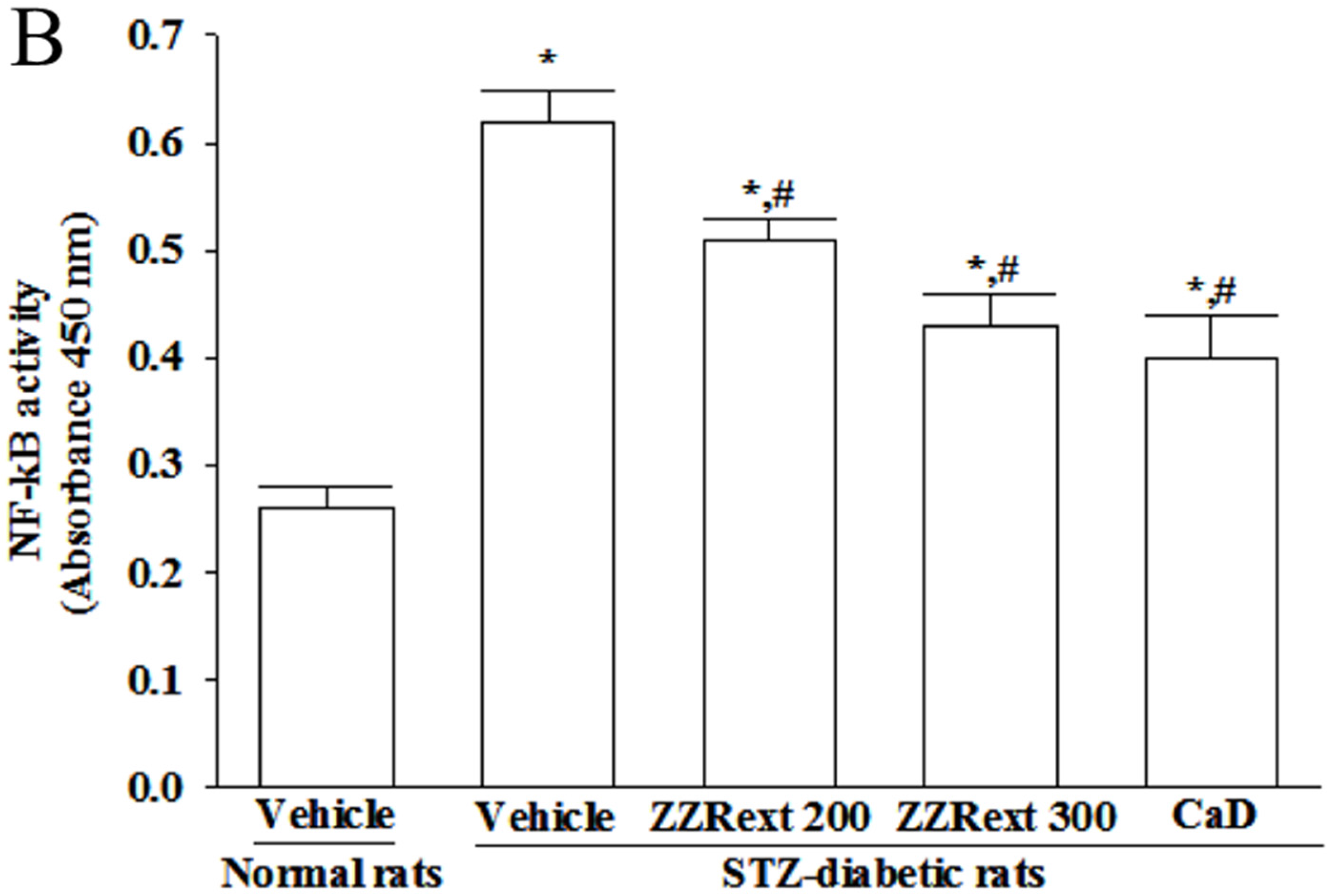
3.8. The NF-κB Activity in Retina
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Campbell, M.; Humphries, P. The blood-retina barrier: Tight junctions and barrier modulation. Adv. Exp. Med. Biol. 2012, 763, 70–84. [Google Scholar] [PubMed]
- Scanlon, P.H.; Aldington, S.J.; Stratton, I.M. Epidemiological issues in diabetic retinopathy. Middle East Afr. J. Ophthalmol. 2013, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Sayin, N.; Kara, N.; Pekel, G. Ocular complications of diabetes mellitus. World J. Diabetes 2015, 6, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Zhang, Z.; Mima, A.; King, G.L. Molecular mechanisms of diabetic vascular complications. J. Diabetes Investig. 2010, 1, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Blonde, L. Benefits and risks for intensive glycemic control in patients with diabetes mellitus. Am. J. Med. Sci. 2012, 343, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Ostri, C.; la Cour, M.; Lund-Andersen, H. Diabetic vitrectomy in a large type 1 diabetes patient population: Long-term incidence and risk factors. Acta. Ophthalmol. 2014, 92, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Romero-Aroca, P.; Reyes-Torres, J.; Baget-Bernaldiz, M.; Blasco-Suñe, C. Laser treatment for diabetic macular edema in the 21st century. Curr. Diabetes Rev. 2014, 10, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Nalawade, S.M.; Sagare, A.P.; Lee, C.Y.; Kao, C.L.; Tsay, H.S. Studies on tissue culture of Chinese medicinal plant resources in Taiwan and their sustainable utilization. Bot. Bull. Aca. Sinica. 2003, 44, 79–98. [Google Scholar]
- Dung, N.X.; Chinh, T.D.; Rang, D.D.; Leclercq, P.A. The constituents of the rhizome oil of Zingiber zerumbet (L.) Sm. From Vietnam. J. Essentl. Oil Res. 1993, 5, 553–555. [Google Scholar] [CrossRef]
- Yob, N.J.; Jofrry, S.M.; Affandi, M.M.; Teh, L.K.; Salleh, M.Z.; Zakaria, Z.A. Zingiber zerumbet (L.) Smith: A Review of Its Ethnomedicinal, Chemical, and Pharmacological Uses. Evid. Based Complement. Alternat. Med. 2011, 2011, 543216. [Google Scholar] [CrossRef] [PubMed]
- Prakash, R.O.; Rabinarayan, A.; Kumar, M.S. Zingiber zerumbet (L.) Sm., a reservoir plant for therapeutic uses: A review. Int. J. Pharma. World Res. 2011, 2, 1–23. [Google Scholar]
- Jantan, I.; Rafi, I.A.; Jalil, J. Platelet-activating factor (PAF) receptor-binding antagonist activity of Malaysian medicinal plants. Phytomedicine 2005, 12, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.K.; Pippen, R.; Rohana, Y.; Noorsaadah, A.R.; Halijah, I.; Norzulaani, K. Screening of selected Zingiberaceae extracts for dengue-2 virus protease inhibitory activities. Sunway Acad. J. 2006, 3, 1–7. [Google Scholar]
- Husen, R.; Pihie, A.H.; Nallappan, M. Screening for antihyperglycaemic activity in several local herbs of Malaysia. J. Ethnopharmacol. 2004, 95, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Tzeng, T.F.; Chang, Y.S.; Liu, I.M. Beneficial impact of Zingiber zerumbet on insulin sensitivity in fructose-fed rats. Planta Med. 2012, 78, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, T.F.; Liou, S.S.; Chang, C.J.; Liu, I.M. The ethanol extract of Zingiber zerumbet attenuates streptozotocin-induced diabetic nephropathy in rats. Evid. Based Complement. Alternat. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Lai, A.K.W.; Lo, A.C.Y. Animal models of diabetic retinopathy: Summary and comparison. J. Diabetes Res. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.R. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Padilla, E.; Ganado, P.; Sanz, M.; Zeini, M.; Ruiz, E.; Triviño, A.; Ramírez, A.I.; Salazar, J.J.; Ramírez, J.M.; Rojas, B.; Hoz, Rd.; Tejerina, T. Calcium dobesilate attenuates vascular injury and the progression of diabetic retinopathy in streptozotocin-induced diabetic rats. Diabetes Metab. Res. Rev. 2005, 21, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Qaum, T.; Adamis, A.P. Sensitive blood-retinal barrier breakdown quantitation using Evans blue. Invest. Ophthalmol. Vis. Sci. 2001, 42, 789–794. [Google Scholar] [PubMed]
- Russo, R.; Cavaliere, F.; Varano, G.P.; Milanese, M.; Adornetto, A.; Nucci, C.; Bonanno, G.; Morrone, L.A.; Corasaniti, M.T.; Bagetta, G. Impairment of neuronal glutamate uptake and modulation of the glutamate transporter GLT-1 induced by retinal ischemia. PLoS ONE 2013, 8, e69250. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Berthet, P.; Farine, J.C.; Barras, J.P. Calcium dobesilate: Pharmacological profile related to its use in diabetic retinopathy. Int. J. Clin. Pract. 1999, 53, 631–636. [Google Scholar] [PubMed]
- van Dijk, H.W.; Verbraak, F.D.; Stehouwer, M.; Kok, P.H.; Garvin, M.K.; Sonka, M.; DeVries, J.H.; Schlingemann, R.O.; Abràmoff, M.D. Association of visual function and ganglion cell layer thickness in patients with diabetes mellitus type 1 and no or minimal diabetic retinopathy. Vision Res. 2011, 51, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Matter, K.; Balda, M.S. Occludin and the functions of tight junctions. Int. Rev. Cytol. 1999, 186, 117–146. [Google Scholar] [PubMed]
- Turksen, K.; Troy, T.C. Barriers built on claudins. J. Cell Sci. 2004, 117, 2435–2447. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Wittchen, E.S.; Haskins, J.; Stevenson, B.R. Protein interactions at the tight junction. Actin has multiple binding partners, and ZO-1 forms independent complexes with ZO-2 and ZO-3. J. Biol. Chem. 1999, 274, 35179–35185. [Google Scholar] [CrossRef] [PubMed]
- Leal, E.C.; Martins, J.; Voabil, P.; Liberal, J.; Chiavaroli, C.; Bauer, J.; Cunha-Vaz, J.; Ambrósio, A.F. Calcium dobesilate inhibits the alterations in tight junction proteins and leukocyte adhesion to retinal endothelial cells induced by diabetes. Diabetes 2010, 59, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- El-Remessy, A.B.; Al-Shabrawey, M.; Khalifa, Y.; Tsai, N.T.; Caldwell, R.B.; Liou, G.I. Neuroprotective and blood-retinal barrier-preserving effects of cannabidiol in experimental diabetes. Am. J. Pathol. 2006, 168, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Mattila, T.K.; de Boer, A. Influence of intensive versus conventional glucose control on microvascular and macrovascular complications in type 1 and 2 diabetes mellitus. Drugs 2010, 70, 2229–2245. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Koppolu, P.; Chakrabarti, S.; Chen, S. Diabetes-induced activation of nuclear transcriptional factor in the retina, and its inhibition by antioxidants. Free Radic. Res. 2003, 37, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, S.; Xia, X. Role of intravitreal inflammatory cytokines and angiogenic factors in proliferative diabetic retinopathy. Curr. Eye Res. 2012, 37, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Yan, H. Roles of elevated intravitreal IL-1β and IL-10 levels in proliferative diabetic retinopathy. Indian J. Ophthalmol. 2014, 62, 699–701. [Google Scholar] [PubMed]
- Simó, R.; Sundstrom, J.M.; Antonetti, D.A. Ocular Anti-VEGF therapy for diabetic retinopathy: The role of VEGF in the pathogenesis of diabetic retinopathy. Diabetes Care 2014, 37, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Uğurlu, N.; Gerceker, S.; Yülek, F.; Ugurlu, B.; Sarı, C.; Baran, P.; Çağil, N. The levels of the circulating cellular adhesion molecules ICAM-1, VCAM-1 and endothelin-1 and the flow-mediated vasodilatation values in patients with type 1 diabetes mellitus with early-stage diabetic retinopathy. Intern. Med. 2013, 52, 2173–2178. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Kern, T.S. Inflammation in diabetic retinopathy. Prog. Retin. Eye Res. 2011, 30, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-κB Signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Szabo, C.; Kern, T.S. Poly (ADP-ribose) polymerase is involved in the development of diabetic retinopathy via regulation of nuclear factor-kappa B. Diabetes 2004, 53, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Romeo, G.; Liu, W.H.; Asnaghi, V.; Kern, T.S.; Lorenzi, M. Activation of nuclear factor-kappaB induced by diabetes and high glucose regulates a Proapoptotic Program in retinal pericytes. Diabetes 2002, 51, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Kowall, B.; Rathmann, W. HbA1c for diagnosis of type 2 diabetes. Is there an optimal cut point to assess high risk of diabetes complications, and how well does the 6.5% cutoff perform? Diabetes Metab. Syndr. Obes. 2013, 6, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Cases, J.; Romain, C.; Dallas, C.; Gerbi, A.; Cloarec, M. Regular consumption of Fiit-ns, a polyphenol extract from fruit and vegetables frequently consumed within the Mediterranean diet, improves metabolic ageing of obese volunteers: A randomized, double-blind, parallel trial. Int. J. Food Sci. Nutr. 2015, 66, 120–125. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzeng, T.-F.; Hong, T.-Y.; Tzeng, Y.-C.; Liou, S.-S.; Liu, I.-M. Consumption of Polyphenol-Rich Zingiber Zerumbet Rhizome Extracts Protects against the Breakdown of the Blood-Retinal Barrier and Retinal Inflammation Induced by Diabetes. Nutrients 2015, 7, 7821-7841. https://doi.org/10.3390/nu7095369
Tzeng T-F, Hong T-Y, Tzeng Y-C, Liou S-S, Liu I-M. Consumption of Polyphenol-Rich Zingiber Zerumbet Rhizome Extracts Protects against the Breakdown of the Blood-Retinal Barrier and Retinal Inflammation Induced by Diabetes. Nutrients. 2015; 7(9):7821-7841. https://doi.org/10.3390/nu7095369
Chicago/Turabian StyleTzeng, Thing-Fong, Tang-Yao Hong, Yu-Cheng Tzeng, Shorong-Shii Liou, and I-Min Liu. 2015. "Consumption of Polyphenol-Rich Zingiber Zerumbet Rhizome Extracts Protects against the Breakdown of the Blood-Retinal Barrier and Retinal Inflammation Induced by Diabetes" Nutrients 7, no. 9: 7821-7841. https://doi.org/10.3390/nu7095369
APA StyleTzeng, T.-F., Hong, T.-Y., Tzeng, Y.-C., Liou, S.-S., & Liu, I.-M. (2015). Consumption of Polyphenol-Rich Zingiber Zerumbet Rhizome Extracts Protects against the Breakdown of the Blood-Retinal Barrier and Retinal Inflammation Induced by Diabetes. Nutrients, 7(9), 7821-7841. https://doi.org/10.3390/nu7095369