The Protective Effect of Rhizoma Dioscoreae Extract against Alveolar Bone Loss in Ovariectomized Rats via Regulating Wnt and p38 MAPK Signaling

 ,
,
Abstract
:
1. Introduction
2. Experimental Section
2.1. Preparation of Aqueous Extract
2.2. Animal Grouping and Treatments
2.3. Preparation of Specimens
2.4. Biochemical Markers of Bone Turnover
2.5. Micro-CT Analysis
2.6. Histological Observation
2.7. Microarray Data Analysis
2.8. Ingenuity Pathway Analysis
2.9. Quantitative Real-Time RT-PCR (qRT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript | Sequence (5′–3′) |
|---|---|
| Gapdh | F: 5′-GGGAAACTGTGGCGTGAT-3′ R: 5′-GAGTGGGTGTCGCTGTTGA-3′ |
| Fzd2 | F: 5′-CAGGGCACTAAGAAAGAAGGCT-3′ R: 5′-AGGAACCAGGTGAGGGACAGA-3′ |
| Il1rn | F: 5′-CTTACCTTCATCCGCTCCGA-3′ R: 5′-GATCAGGCAGTTGGTGGTCAT-3′ |
| Mapk14 | F: 5′-ATAGACGAATGGAAGAGCCTGAC-3′ R: 5′-CAAAGATACATGGACAAACGGAC-3′ |
| Sfrp2 | F: 5′-CCGAAAGGGACCTGAAGAAAT-3′ R: 5′-ACCAGATACGGAGCGTTGATG-3′ |
| Sfrp4 | F: 5′-AAGTCTTTGTCACCTATCCCTCG-3′ R: 5′-CGGCTGGCTATCTGCTTCTT-3′ |
| Spp1 | F: 5′-TTTCACTCCAATCGTCCCTACA-3′ R: 5′-AGTCCATAAGCCAAGCTATCACC-3′ |
| Tcf3 | F: 5′-ACAGTCTCAGCAGCAAATCCAA-3′ R: 5′-GAAGACGCAGGGCTATCACAA-3′ |
| Ptk2b | F: 5′-TCTGTGACCCGTCTACCCATC-3′ R: 5′-CTTTCTCCAGCACTCCGATGA-3′ |
| Wnt7a | F: 5′-CTCTGCCGACATCCGCTAC-3′ R: 5′-CGACCCGCCTCGTTATTG-3′ |
| Bmp1 | F: 5′-CAAAGGACCCGACTCAGCA-3′ R: 5′-CCACATAGTCATACCAGCACAGG-3′ |
| Csf1r | F: 5′-AAGCCGAAATATCAGGTGCG-3′ R: 5′-GGGTCGATGAAGGTGTAGTTGTT-3′ |
| Frzb | F: 5′-TCCAAGGGATACCGTCAACC-3′ R: 5′-ATCCTTCCACTTCTCAGCGATAG-3′ |
2.10. Western Analysis
2.11. Statistical Analysis
3. Results
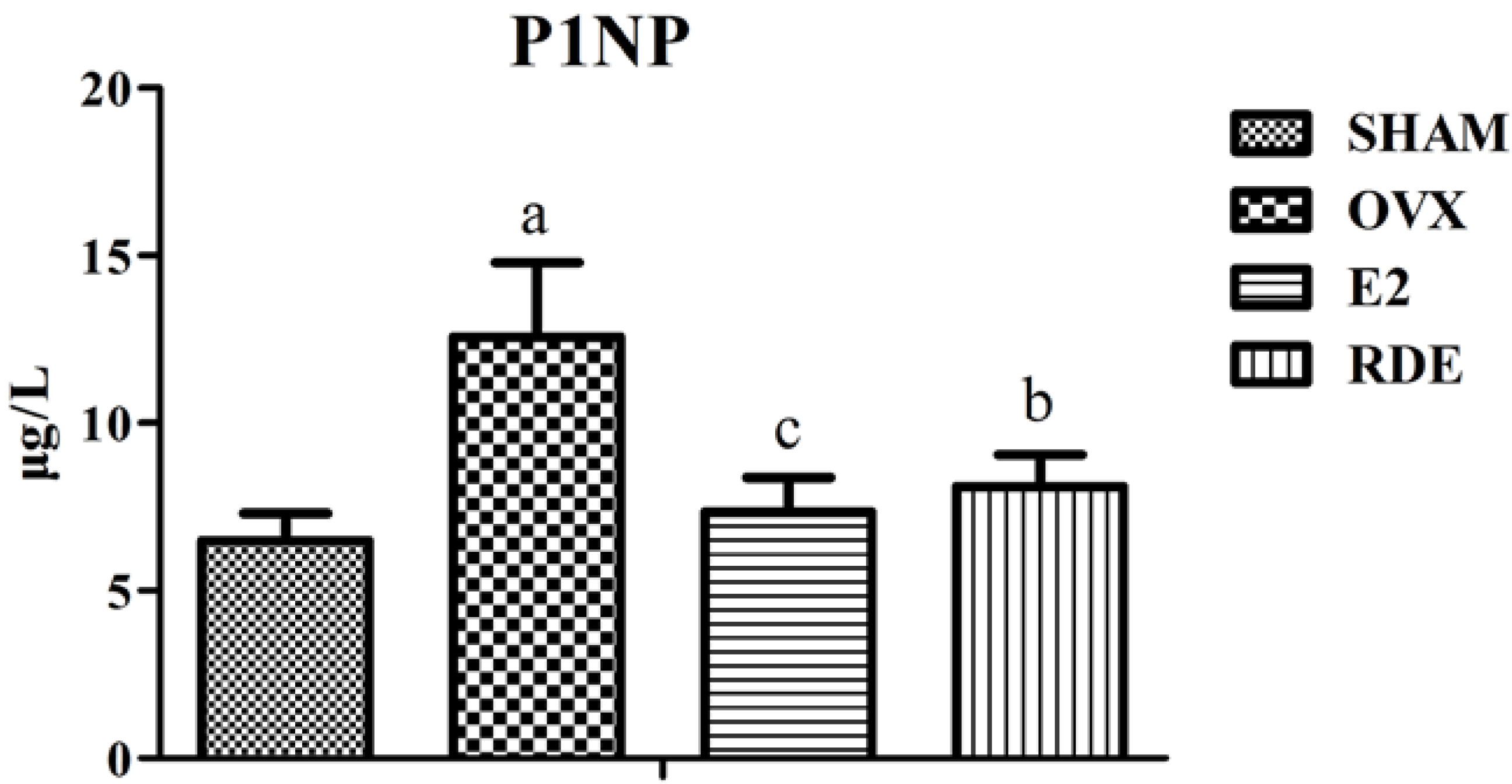
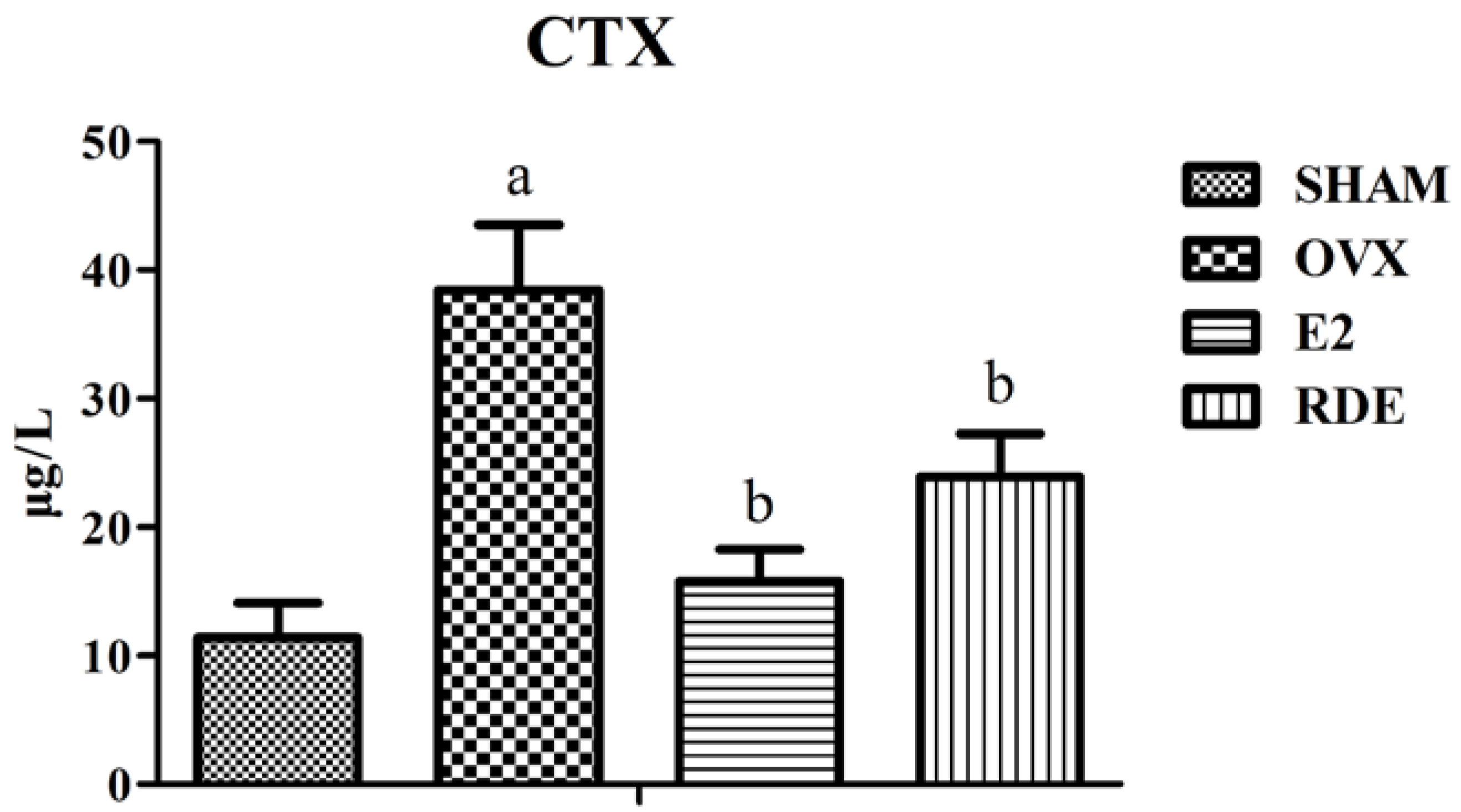
3.1. Effect of RDE on Biomarkers of Bone Turnover
3.2. Effect of RDE on BMD and Trabecular Bone Microarchitecture
| Parameters | SHAM | OVX | E2 | RDE |
|---|---|---|---|---|
| BMD (g/cm3) | 0.348 ± 0.005 | 0.295 ± 0.006 a | 0.328 ± 0.008 c | 0.313 ± 0.004 b |
| BV/TV (%) | 55.100 ± 4.718 | 6.777 ± 4.1569 a | 42.756 ± 0.843 c | 22.299 ± 2.535 c |
| Tb.Th (μm) | 81.592 ± 8.654 | 41.237 ± 2.592 a | 74.226 ± 6.361 c | 49.382 ± 3.976 c |
| Tb.Sp (μm) | 181.62 ± 26.48 | 678.27 ± 33.21 a | 319.63 ± 95.91 c | 519.29 ± 13.55 c |
| Tb.N (1/mm) | 6.84 ± 1.28 | 1.61 ± 0.91 a | 5.81 ± 0.61 c | 4.52 ± 0.40 c |
| SMI | 0.914 ± 0.275 | 2.327 ± 0.419 a | 1.637 ± 0.217 c | 1.712 ± 0.379 c |
| DA | 2.526 ± 0.837 | 7.770 ± 0.679 a | 2.185 ± 0.271 | 1.861 ± 0.188 |

3.3. Effect of RDE on Histological Morphology of Alveolar Bone
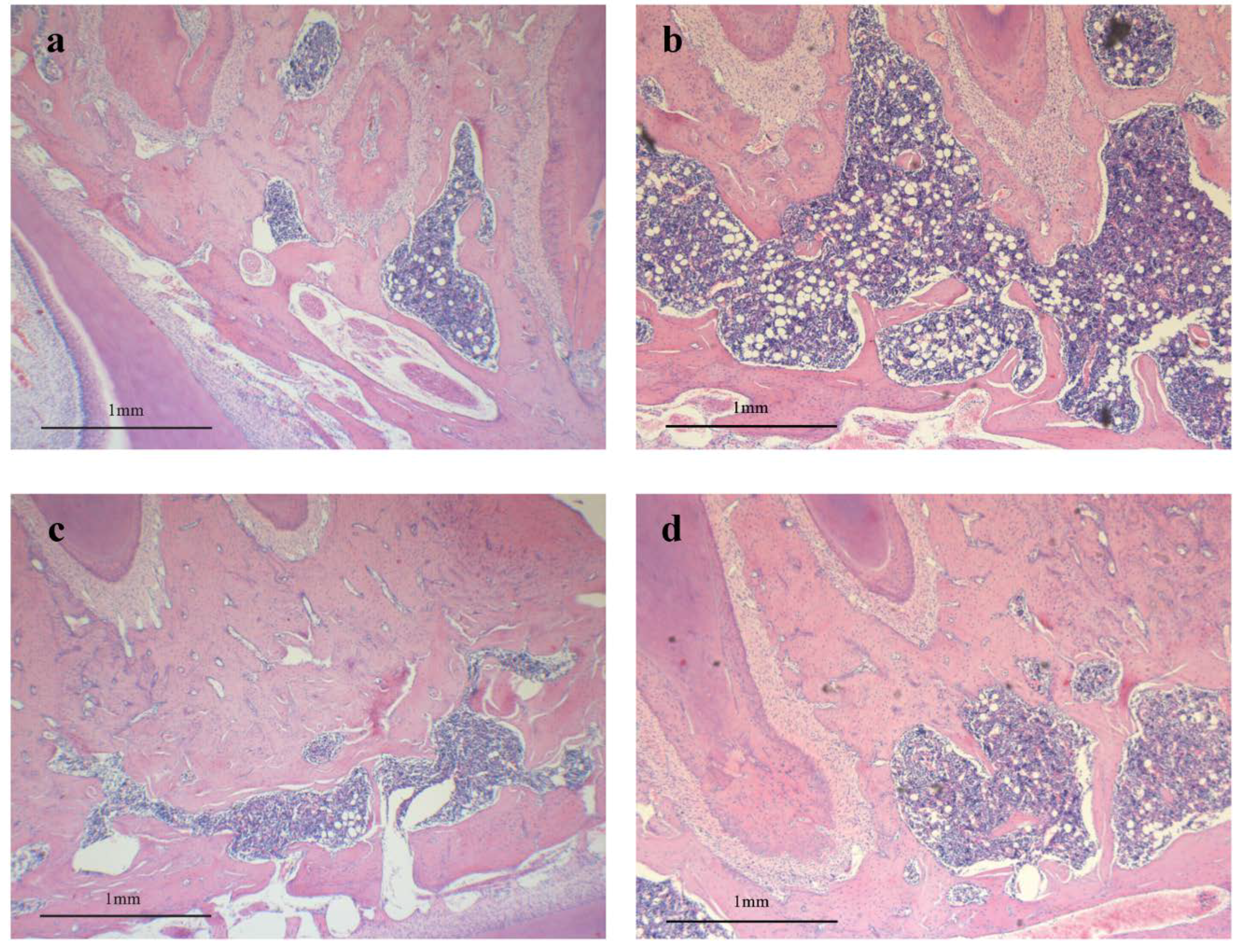
3.4. Effect of RDE on Gene Expression Profile
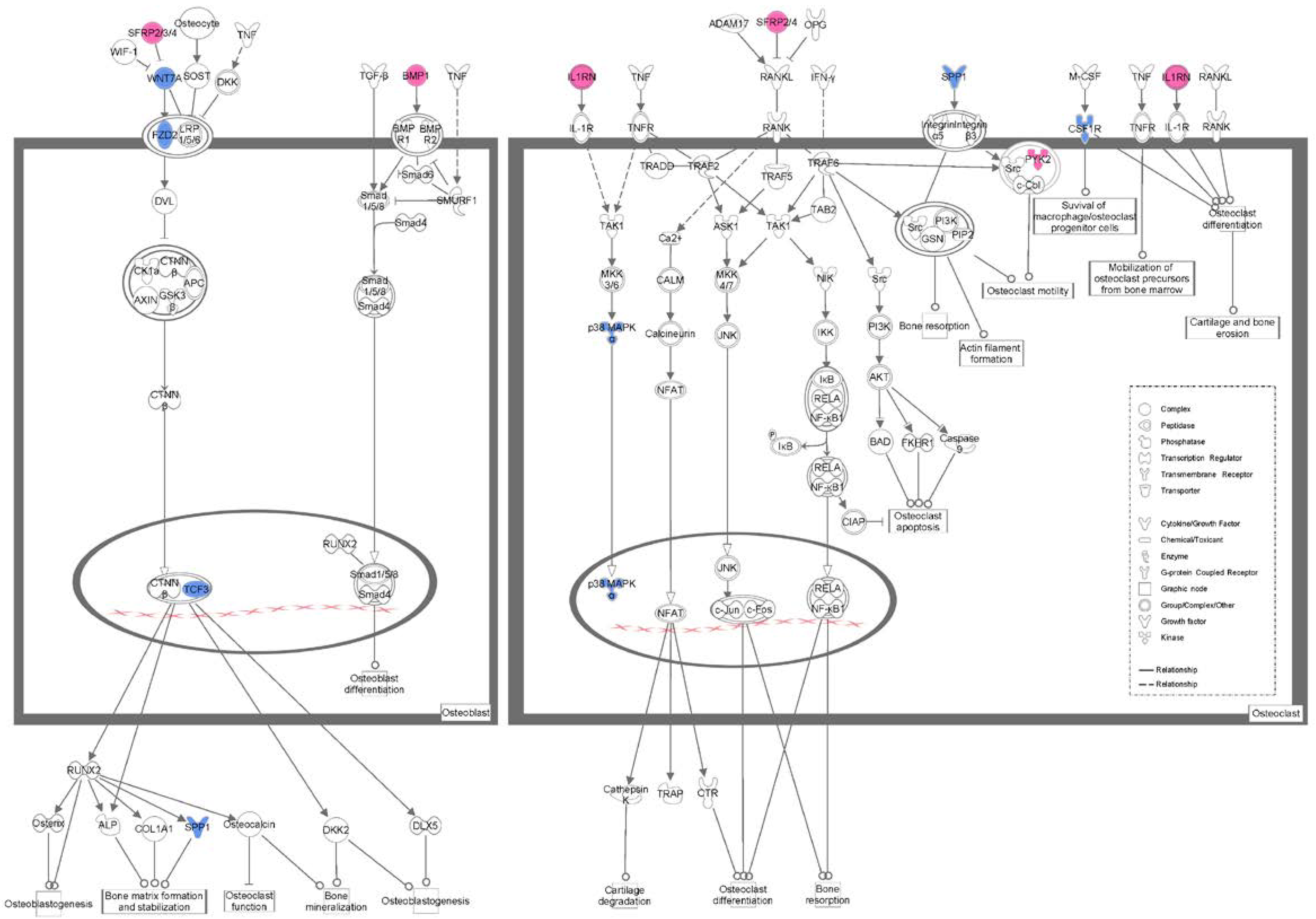
3.5. Pathway Analysis of Differentially Expressed Genes
| Ingenuity Canonical Pathways | p-Value | Number of Molecules |
|---|---|---|
| Role of osteoblasts, osteoclasts and chondrocytes in rheumatoid arthritis | 1.23 × 10−3 | 12 |
| Role of Wnt/GSK-3β signaling in the pathogenesis of influenza | 1.34 × 10−2 | 4 |
| Basal cell carcinoma signaling | 2.02 × 10−2 | 4 |
| Human embryonic stem cell pluripotency | 2.33 × 10−2 | 6 |
| Wnt/β-catenin signaling | 4.07 × 10−2 | 6 |
3.6. Confirmation of Differential Levels of Gene Expression by qRT-PCR


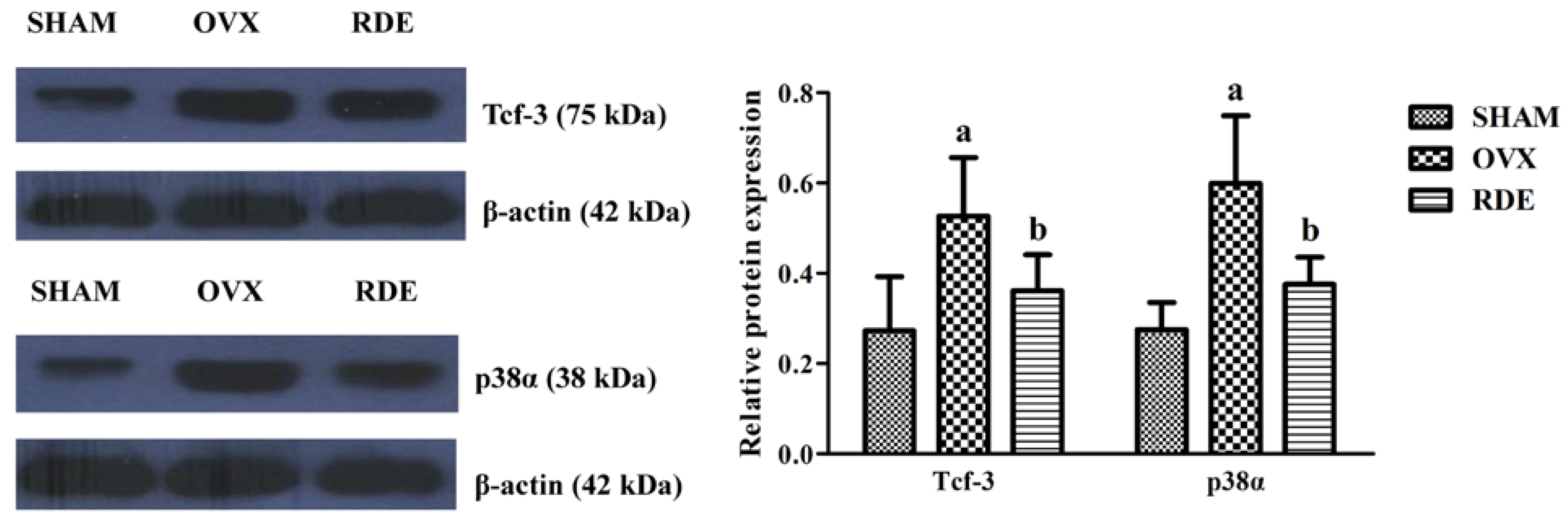
3.7. Confirmation of Proteins by Western Blotting
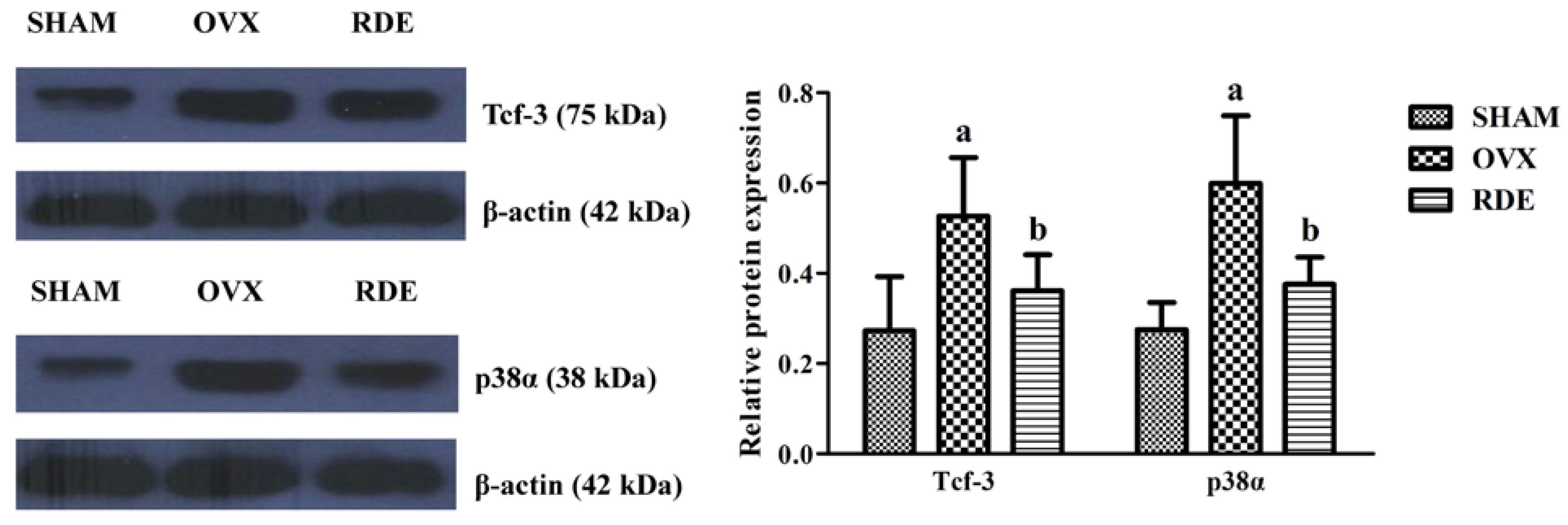
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Z.; Song, C.; Fu, X.; Liu, M.; Li, Y.; Pan, J.; Liu, H.; Wang, S.; Xiang, L.; Xiao, G.G.; et al. High-dose diosgenin reduces bone loss in ovariectomized rats via attenuation of the RANKL/OPG ratio. Int. J. Mol. Sci. 2014, 15, 17130–17147. [Google Scholar] [CrossRef] [PubMed]
- Pinkerton, J.V.; Thomas, S.; Dalkin, A.C. Osteoporosis treatment and prevention for postmenopausal women: Current and future therapeutic options. Clin. Obstet. Gynecol. 2013, 56, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.D.; White, S.C. Age and trabecular features of alveolar bone associated with osteoporosis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2005, 100, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Streckfus, C.F.; Johnson, R.B.; Nick, T.; Tsao, A.; Tucci, M. Comparison of alveolar bone loss, alveolar bone density and second metacarpal bone density, salivary and gingival crevicular fluid interleukin-6 concentrations in healthy premenopausal and postmenopausal women on estrogen therapy. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 1997, 52, M343–M351. [Google Scholar] [CrossRef]
- Sultan, N.; Rao, J. Association between periodontal disease and bone mineral density in postmenopausal women: A cross sectional study. Med. Oral Patol. Oral Cir. Bucal 2011, 16, e440–e447. [Google Scholar] [CrossRef] [PubMed]
- Tezal, M.; Wactawski-Wende, J.; Grossi, S.G.; Ho, A.W.; Dunford, R.; Genco, R.J. The relationship between bone mineral density and periodontitis in postmenopausal women. J. Periodontol. 2000, 71, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Filho, I.S.; Passos Jde, S.; Cruz, S.S.; Vianna, M.I.; Cerqueira Ede, M.; Oliveira, D.C.; dos Santos, C.A.; Coelho, J.M.; Sampaio, F.P.; Freitas, C.O.; et al. The association between postmenopausal osteoporosis and periodontal disease. J. Periodontol. 2007, 78, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Tezal, M.; Wactawski-Wende, J.; Grossi, S.G.; Dmochowski, J.; Genco, R.J. Periodontal disease and the incidence of tooth loss in postmenopausal women. J. Periodontol. 2005, 76, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Civitelli, R.; Pilgram, T.K.; Dotson, M.; Muckerman, J.; Lewandowski, N.; Armamento-Villareal, R.; Yokoyama-Crothers, N.; Kardaris, E.E.; Hauser, J.; Cohen, S.; et al. Alveolar and postcranial bone density in postmenopausal women receiving hormone/estrogen replacement therapy: A randomized, double-blind, placebo-controlled trial. Arch. Intern. Med. 2002, 162, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Palomo, L.; Bissada, N.F.; Liu, J. Periodontal assessment of postmenopausal women receiving risedronate. Menopause 2005, 12, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, Z.; Li, C. Intermittent PTH administration: A novel therapy method for periodontitis-associated alveolar bone loss. Med. Hypotheses 2009, 72, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Strom, B.L.; Schinnar, R.; Weber, A.L.; Bunin, G.; Berlin, J.A.; Baumgarten, M.; DeMichele, A.; Rubin, S.C.; Berlin, M.; Troxel, A.B.; et al. Case-control study of postmenopausal hormone replacement therapy and endometrial cancer. Am. J. Epidemiol. 2006, 164, 775–786. [Google Scholar] [PubMed]
- Rossing, M.A.; Cushing-Haugen, K.L.; Wicklund, K.G.; Doherty, J.A.; Weiss, N.S. Menopausal hormone therapy and risk of epithelial ovarian cancer. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.B.; Hellstein, J.W.; Kalmar, J.R. Narrative corrected review: Bisphosphonates and osteonecrosis of the jaws. Ann. Intern. Med. 2006, 144, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R.; Reginster, J.Y.; Boonen, S.; Breart, G.; Diez-Perez, A.; Felsenberg, D.; Kaufman, J.M.; Kanis, J.A.; Cooper, C. Adverse reactions and drug-drug interactions in the management of women with postmenopausal osteoporosis. Calcif. Tissue Int. 2011, 89, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Clemett, D.; Spencer, C.M. Raloxifene: A review of its use in postmenopausal osteoporosis. Drugs 2000, 60, 379–411. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, C.; Cao, H.; Chan, K.; Lu, A. The role of chinese medicine in the treatment of chronic diseases in China. Planta Med. 2011, 77, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiang, L.; Bai, D.; Fu, X.; Wang, W.; Li, Y.; Liu, H.; Pan, J.; Xiao, G.G.; Ju, D. Treatment with Rhizoma Dioscoreae extract has protective effect on osteopenia in ovariectomized rats. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Li, C.M.; Dong, X.L.; Fan, X.D.; Wu, J.H.; Wang, Q.H.; Tian, X.L.; Guo, D.J.; Wong, M.S.; Qiu, T.Q.; Chan, S.W. Aqueous extract of danshen (Salvia miltiorrhiza Bunge) protects ovariectomized rats fed with high-fat diet from endothelial dysfunction. Menopause 2013, 20, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Maimoun, L.; Brennan-Speranza, T.C.; Rizzoli, R.; Ammann, P. Effects of ovariectomy on the changes in microarchitecture and material level properties in response to hind leg disuse in female rats. Bone 2012, 51, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.E.; Yao, W.; Kinney, J.H.; Modin, G.; Balooch, M.; Wronski, T.J. Both hPTH(1–34) and bFGF increase trabecular bone mass in osteopenic rats but they have different effects on trabecular bone architecture. J. Bone Miner. Res. 2003, 18, 2105–2115. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Pham, S.M.; Crabbe, D.L. High-resolution Micro-CT evaluation of mid- to long-term effects of estrogen deficiency on rat trabecular bone. Acad. Radiol. 2003, 10, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]
- Ikeo, T.; Goda, S.; Domae, E. Metabolism of alveolar bone. Clin. Calcium 2006, 16, 117–121. [Google Scholar] [PubMed]
- Taguchi, A.; Tanimoto, K.; Suei, Y.; Wada, T. Tooth loss and mandibular osteopenia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1995, 79, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Krall, E.A.; Garcia, R.I.; Dawson-Hughes, B. Increased risk of tooth loss is related to bone loss at the whole body, hip, and spine. Calcif. Tissue Int. 1996, 59, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, P.; Liu, W.; Maye, P.; Zhang, J.; Zhang, Y.; Hurley, M.; Guo, C.; Boskey, A.; Sun, L.; et al. Dkk2 has a role in terminal osteoblast differentiation and mineralized matrix formation. Nat. Genet. 2005, 37, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Michigami, T.; Ozono, K. Wnt signaling in bone metabolism. J. Bone Miner. Metab. 2009, 27, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R. Wnt signaling. Cold Spring Harb. Perspect. Biol. 2012. [Google Scholar] [CrossRef]
- Ueno, K.; Hirata, H.; Hinoda, Y.; Dahiya, R. Frizzled homolog proteins, microRNAs and Wnt signaling in cancer. Int. J. Cancer 2013, 132, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Bovolenta, P.; Esteve, P.; Ruiz, J.M.; Cisneros, E.; Lopez-Rios, J. Beyond Wnt inhibition: New functions of secreted frizzled-related proteins in development and disease. J. Cell Sci. 2008, 121, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. The complex world of Wnt receptor signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Deepak, V.; Meng, L.; Wang, L.; Li, Y.; Jiang, Q.; Zeng, X.; Liu, W. Analysis of HDAC1-mediated regulation of RUNX2-induced osteopontin gene expression in C3h10t1/2 cells. Biotechnol. Lett. 2012, 34, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Lekic, P.; Sodek, J.; McCulloch, C.A. Relationship of cellular proliferation to expression of osteopontin and bone sialoprotein in regenerating rat periodontium. Cell Tissue Res. 1996, 285, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, E.H.; Shin, M.Y.; Sohn, H.Y.; Park, Y.M.; Kim, T.; Lim, J.H.; Jeong, H.J.; Kwon, S.T.; Kwun, I.S. Diosgenin stimulates osteogenic activity by increasing bone matrix protein synthesis and bone-specific transcription factor RUNX2 in osteoblastic MC3T3-E1 cells. J. Nutr. Biochem. 2011, 22, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Cobb, M.H.; Goldsmith, E.J. How map kinases are regulated. J. Biol. Chem. 1995, 270, 14843–14846. [Google Scholar] [CrossRef] [PubMed]
- Whitmarsh, A.J.; Davis, R.J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. Berl. 1996, 74, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Kitaura, H.; Zhou, P.; Ross, F.P.; Teitelbaum, S.L. IL-1 mediates TNF-induced osteoclastogenesis. J. Clin. Investig. 2005, 115, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.E.; Li, F.; Coatney, D.D.; Otremba, J.; Kriegl, J.M.; Protter, T.A.; Higgins, L.S.; Medicherla, S.; Kirkwood, K.L. A p38 mitogen-activated protein kinase inhibitor arrests active alveolar bone loss in a rat periodontitis model. J. Periodontol. 2007, 78, 1992–1998. [Google Scholar]
- Zhang, R.; Wang, L.; Peng, B. Activation of p38 mitogen-activated protein kinase in rat periapical lesions. J. Endod. 2008, 34, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Xiang, L.; Bai, D.; Wang, W.; Li, Y.; Pan, J.; Liu, H.; Wang, S.; Xiao, G.G.; Ju, D. The Protective Effect of Rhizoma Dioscoreae Extract against Alveolar Bone Loss in Ovariectomized Rats via Regulating Wnt and p38 MAPK Signaling. Nutrients 2014, 6, 5853-5870. https://doi.org/10.3390/nu6125853
Zhang Z, Xiang L, Bai D, Wang W, Li Y, Pan J, Liu H, Wang S, Xiao GG, Ju D. The Protective Effect of Rhizoma Dioscoreae Extract against Alveolar Bone Loss in Ovariectomized Rats via Regulating Wnt and p38 MAPK Signaling. Nutrients. 2014; 6(12):5853-5870. https://doi.org/10.3390/nu6125853
Chicago/Turabian StyleZhang, Zhiguo, Lihua Xiang, Dong Bai, Wenlai Wang, Yan Li, Jinghua Pan, Hong Liu, Shaojun Wang, Gary Guishan Xiao, and Dahong Ju. 2014. "The Protective Effect of Rhizoma Dioscoreae Extract against Alveolar Bone Loss in Ovariectomized Rats via Regulating Wnt and p38 MAPK Signaling" Nutrients 6, no. 12: 5853-5870. https://doi.org/10.3390/nu6125853
APA StyleZhang, Z., Xiang, L., Bai, D., Wang, W., Li, Y., Pan, J., Liu, H., Wang, S., Xiao, G. G., & Ju, D. (2014). The Protective Effect of Rhizoma Dioscoreae Extract against Alveolar Bone Loss in Ovariectomized Rats via Regulating Wnt and p38 MAPK Signaling. Nutrients, 6(12), 5853-5870. https://doi.org/10.3390/nu6125853