
Lacticaseibacillus paracasei 36 Mitigates Alcoholic-Associated Liver Disease Through Modulation of Microbiota and AMPK Signaling

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Preparation and Pretreatment of ALD Mice
2.3. Histopathological Analysis
2.4. Biochemical Analyses
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. The 16S rRNA Gene Sequencing and Data Processing
2.7. RNA-Seq Analysis
2.8. Statistical Analyses
3. Results
3.1. Effects of LP36 on Body Weight Change and Anti-Inflammatory Status in ALD Mice
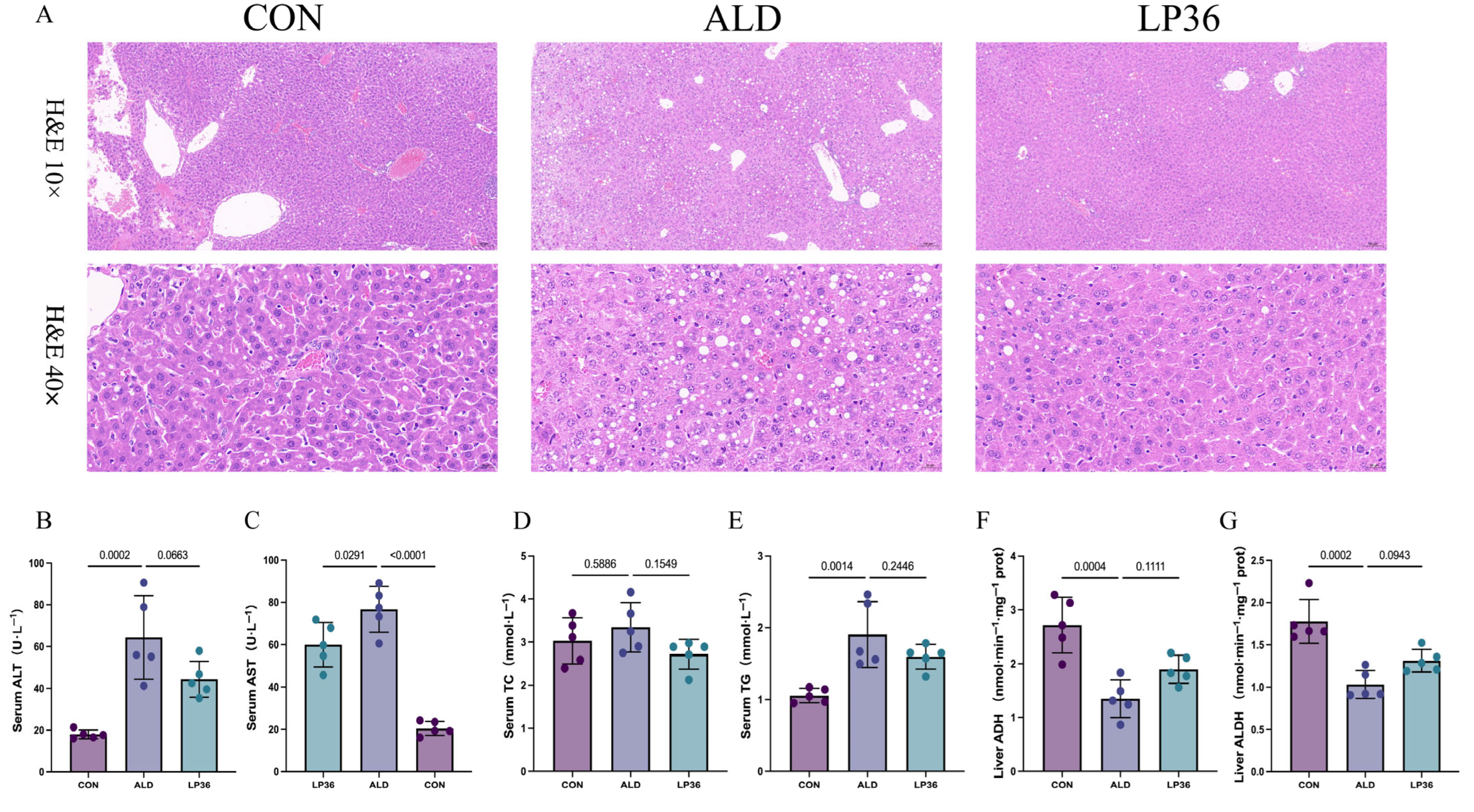
3.2. LP36 Pretreatment Alleviates Ethanol-Induced Liver Injury and Promotes Ethanol Metabolism in Mice
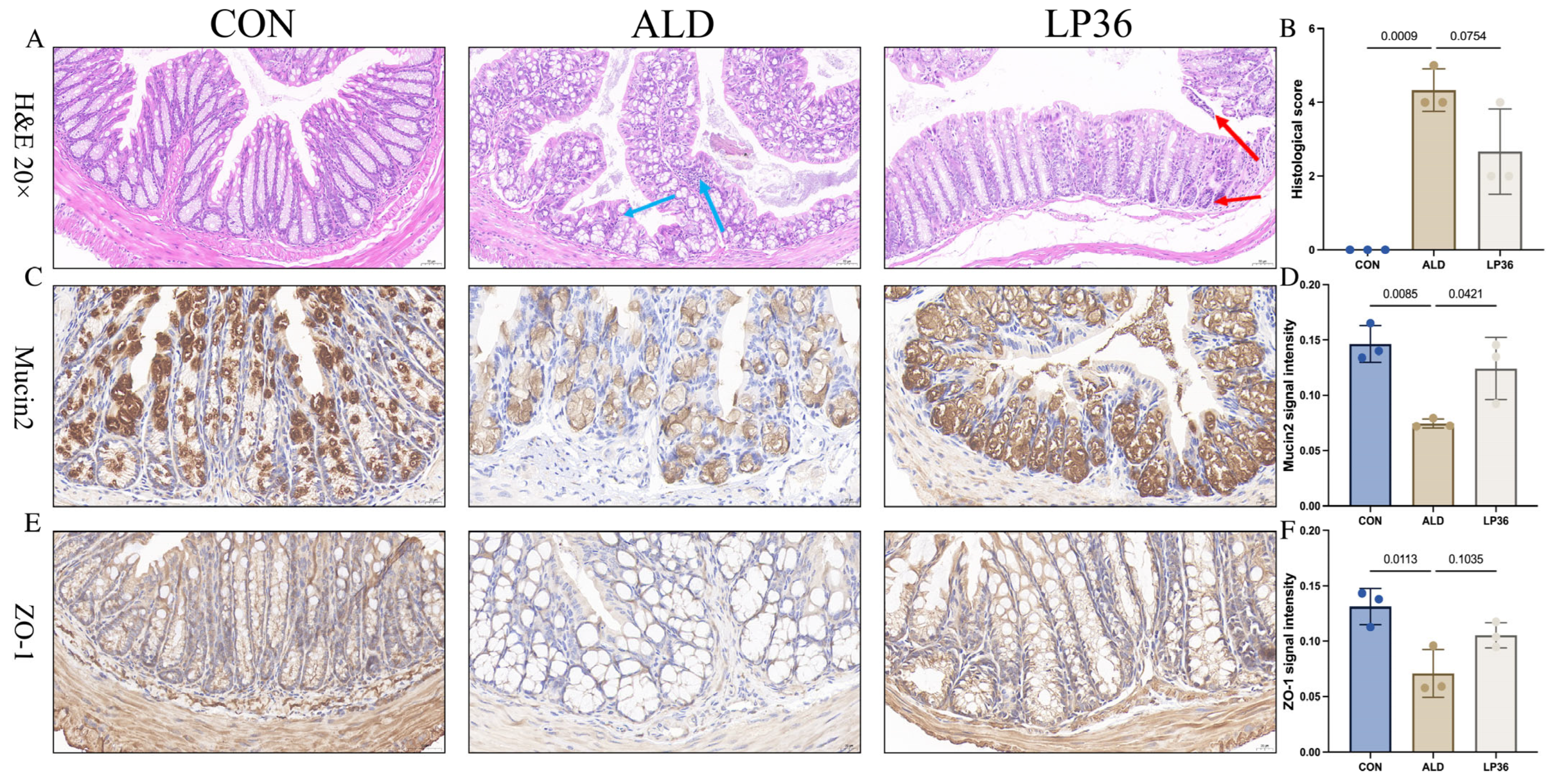
3.3. LP36 Pretreatment Improves Ethanol-Induced Intestinal Injury in Mice
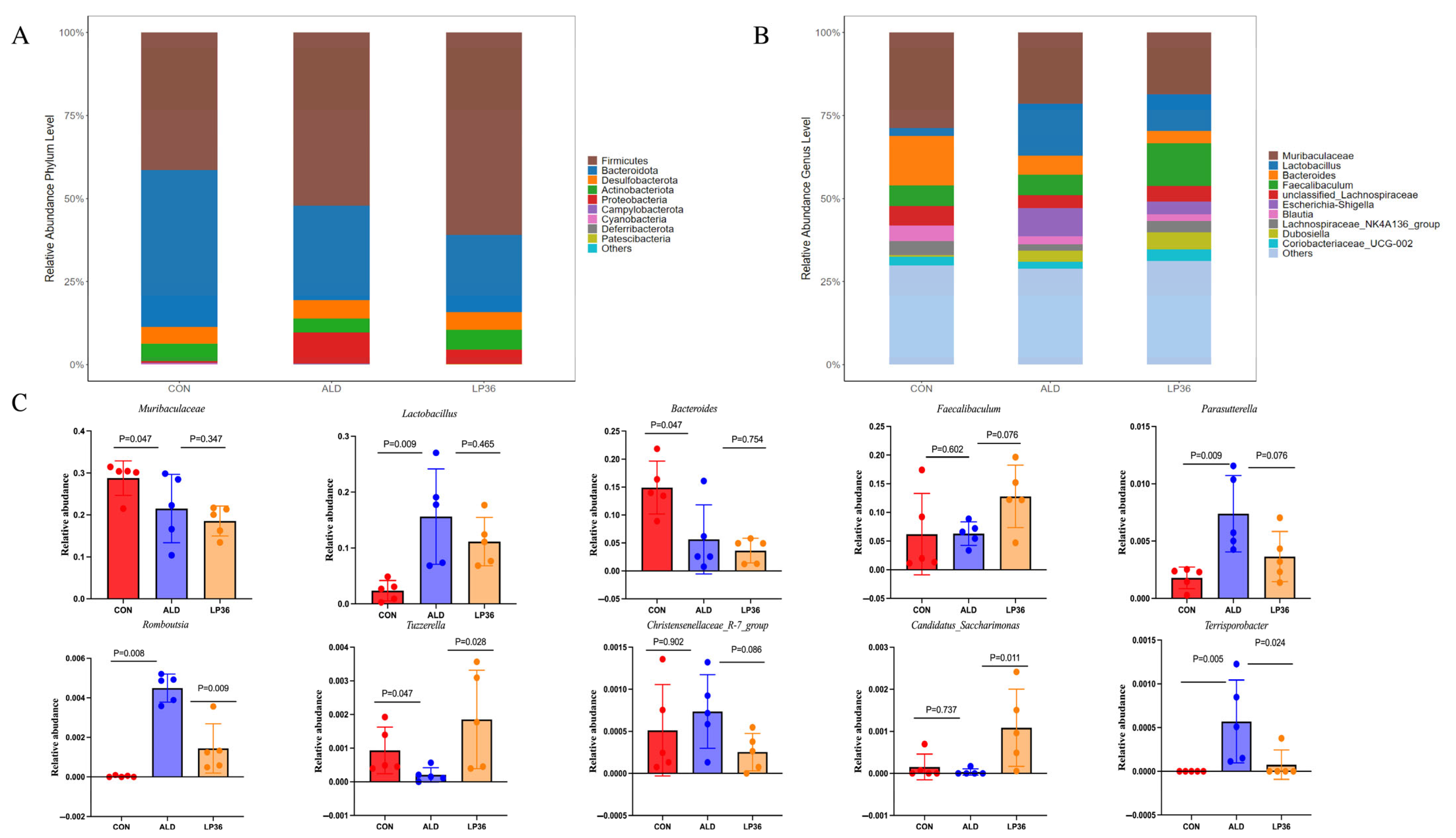
3.4. LP36 Improves the Gut Microbiota Composition in Mice
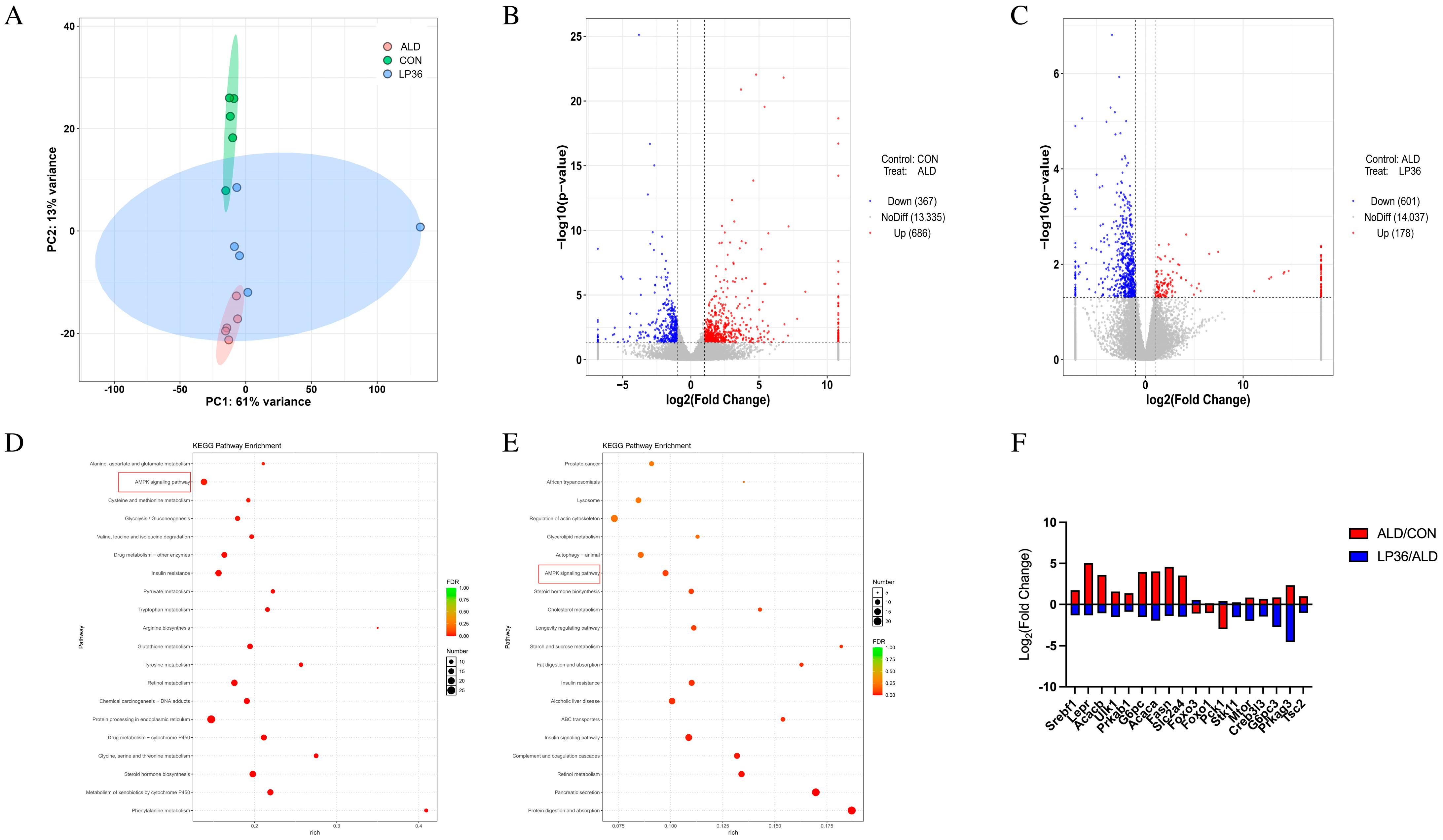
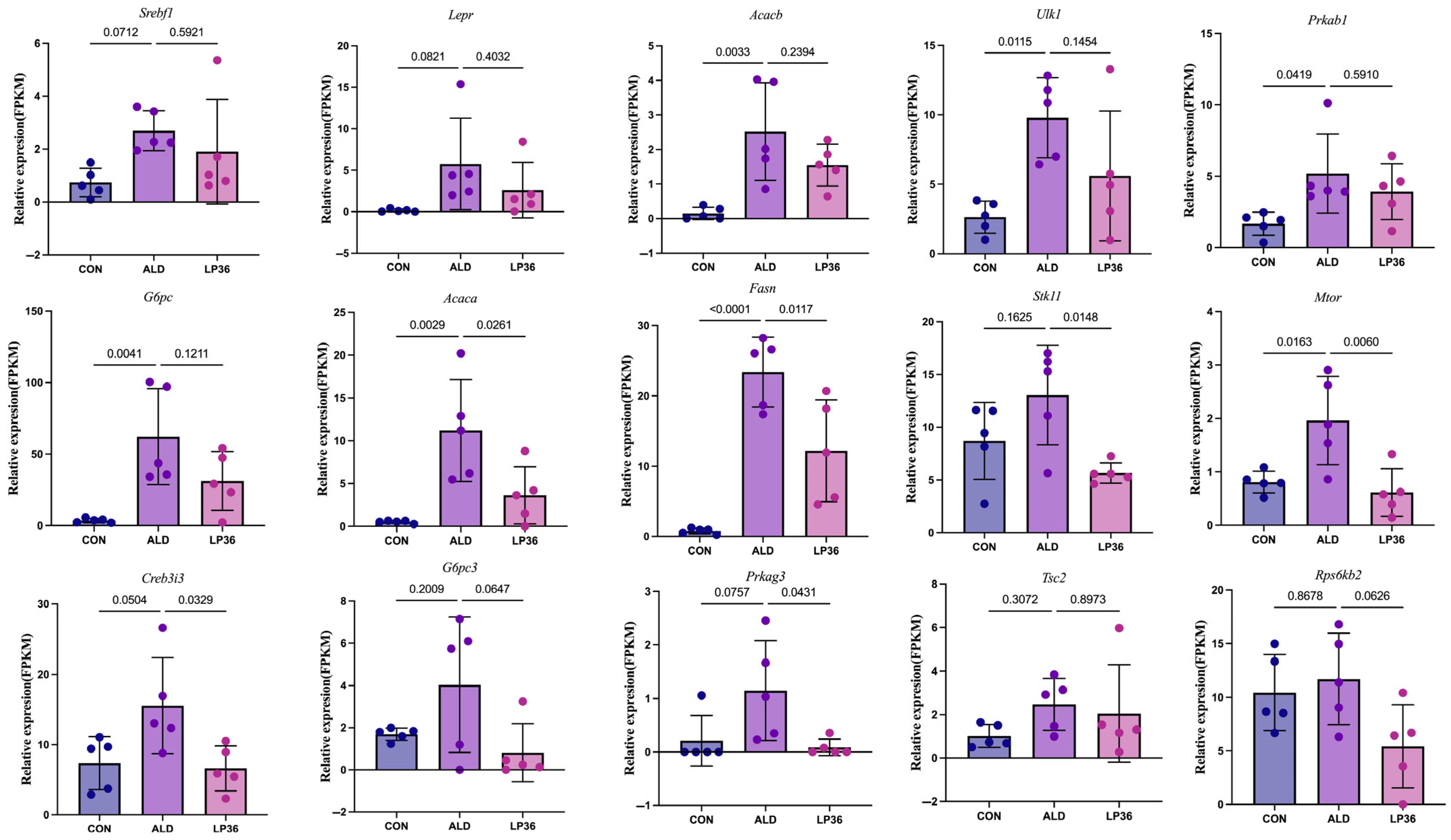
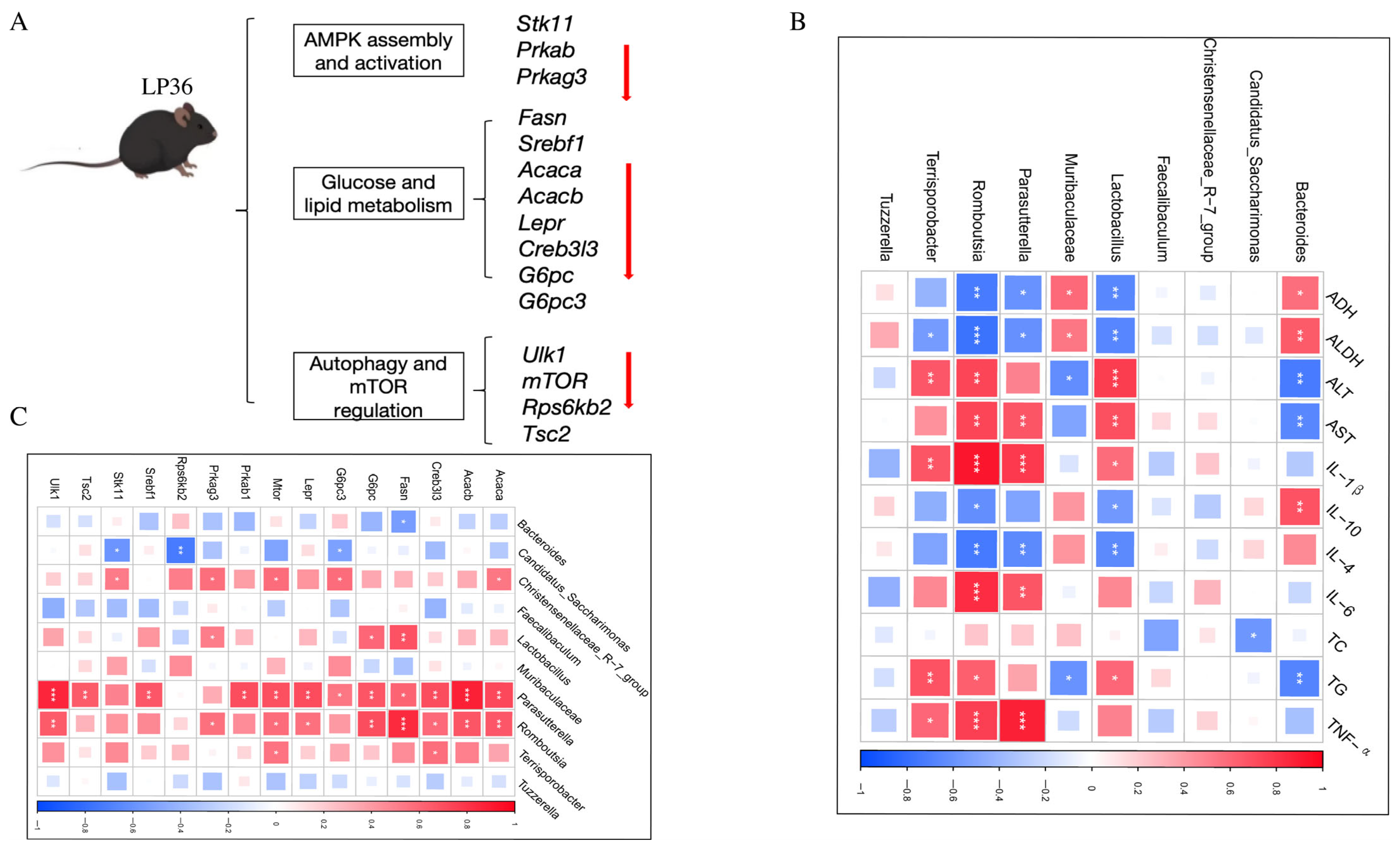
3.5. LP36 Regulates the Liver Transcriptomics in Mice
3.6. Correlation Among Liver Injury and Inflammatory Parameters, Gut Microbiota, and AMPK-Related Genes
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Åberg, F.; Jiang, Z.G.; Cortez-Pinto, H.; Männistö, V. Alcohol-associated liver disease-Global epidemiology. Hepatology 2024, 80, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Mehta, H.; Dunn, W. Determining Prognosis of ALD and Alcohol-associated Hepatitis. J. Clin. Exp. Hepatol. 2023, 13, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Solís-Urra, P.; Rodríguez-Rodríguez, F.; Olivares-Arancibia, J.; Navarro-Oliveros, M.; Abadía-Molina, F.; Álvarez-Mercado, A.I. The Gut Barrier, Intestinal Microbiota, and Liver Disease: Molecular Mechanisms and Strategies to Manage. Int. J. Mol. Sci. 2020, 21, 8351. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Zhou, X.; Xia, F.; Zhou, B. Intestinal Barrier Dysfunction and Gut Microbiota in Non-Alcoholic Fatty Liver Disease: Assessment, Mechanisms, and Therapeutic Considerations. Biology 2024, 13, 243. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Lee, I.O.; Tan, P.L.; Eor, J.Y.; Hwang, J.K.; Kim, S.H. Protective Effect of Lactobacillus fermentum LA12 in an Alcohol-Induced Rat Model of Alcoholic Steatohepatitis. Korean J. Food Sci. Anim. Resour. 2017, 37, 931–939. [Google Scholar] [PubMed]
- Wang, W.; Zhao, X.; Ma, Y.; Zhang, J.; Xu, C.; Ma, J.; Hussain, M.A.; Hou, J.; Qian, S. Alleviating Effect of Lacticaseibacillus rhamnosus 1.0320 Combined with Dihydromyricetin on Acute Alcohol Exposure-Induced Hepatic Impairment: Based on Short-Chain Fatty Acids and Adenosine 5′-Monophosphate-Activated Protein Kinase-Mediated Lipid Metabolism Signaling Pathway. J. Agric. Food Chem. 2023, 71, 4837–4850. [Google Scholar] [PubMed]
- Hsieh, P.S.; Chen, C.W.; Kuo, Y.W.; Ho, H.H. Lactobacillus spp. reduces ethanol-induced liver oxidative stress and inflammation in a mouse model of alcoholic steatohepatitis. Exp. Ther. Med. 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Nighot, P.; Nighot, M.; Haque, M.; Rawat, M.; Ma, T.Y. Lactobacillus acidophilus Induces a Strain-specific and Toll-Like Receptor 2-Dependent Enhancement of Intestinal Epithelial Tight Junction Barrier and Protection Against Intestinal Inflammation. Am. J. Pathol. 2021, 191, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Feng, Y.; Cheng, X.; Xiao, Y.; Zhao, J.; Lu, W.; Tian, F.; Chen, W. The alleviative effects of viable and inactive Lactobacillus paracasei CCFM1120 against alcoholic liver disease via modulation of gut microbiota and the Nrf2/HO-1 and TLR4/MyD88/NF-κB pathways. Food Funct. 2024, 15, 8797–8809. [Google Scholar] [CrossRef] [PubMed]
- Huan, R.; Zhai, Z.; An, J.; Ma, X.; Hao, Y. L-Malic Acid Protects Lacticaseibacillus paracasei L9 from Glycodeoxycholic Acid Stress via the Malolactic Enzyme Pathway. J. Agric. Food Chem. 2022, 70, 9007–9016. [Google Scholar] [CrossRef] [PubMed]
- Wanchai, K.; Yasom, S.; Tunapong, W.; Chunchai, T.; Eaimworawuthikul, S.; Thiennimitr, P.; Chaiyasut, C.; Pongchaidecha, A.; Chatsudthipong, V.; Chattipakorn, S.; et al. Probiotic Lactobacillus paracasei HII01 protects rats against obese-insulin resistance-induced kidney injury and impaired renal organic anion transporter 3 function. Clin. Sci. 2018, 132, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeon, H.J.; Kim, D.G.; Kim, J.Y.; Shim, J.J.; Lee, J.H. Lacticaseibacillus paracsei HY7207 Alleviates Hepatic Steatosis, Inflammation, and Liver Fibrosis in Mice with Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2024, 25, 9870. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Cai, Y.; Jia, C.; Zhang, X.; Huang, Q.; Wei, J.; Chen, Q.; Chen, T. BefA protein alleviates progression of non-alcoholic fatty liver disease by modulating the AMPK signaling pathway through the gut-liver axis. Int. J. Biol. Macromol. 2025, 294, 139446. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Guo, F.; Li, Y.; Liang, Y.; Li, G.; Fu, P.; Ma, L. Activation of AMPK by triptolide alleviates nonalcoholic fatty liver disease by improving hepatic lipid metabolism, inflammation and fibrosis. Phytomedicine 2021, 92, 153739. [Google Scholar] [CrossRef] [PubMed]
- Chyau, C.C.; Wang, H.F.; Zhang, W.J.; Chen, C.C.; Huang, S.H.; Chang, C.C.; Peng, R.Y. Antrodan Alleviates High-Fat and High-Fructose Diet-Induced Fatty Liver Disease in C57BL/6 Mice Model via AMPK/Sirt1/SREBP-1c/PPARγ Pathway. Int. J. Mol. Sci. 2020, 21, 360. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Duan, K.; Wang, C.; McClain, C.; Feng, W. Probiotics and Alcoholic Liver Disease: Treatment and Potential Mechanisms. Gastroenterol. Res. Pract. 2016, 2016, 5491465. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Na, G.H.; Yim, D.J.; Liu, C.F.; Lin, T.H.; Shih, T.W.; Pan, T.M.; Lee, C.L.; Koo, Y.K. Lactobacillus paracasei subsp. paracasei NTU 101 prevents obesity by regulating AMPK pathways and gut microbiota in obese rat. Biochem. Biophys. Res. Commun. 2024, 731, 150279. [Google Scholar]
- Zhao, Y.; Xie, W.; Duan, J.; Li, F. Probiotic Limosilactobacillus reuteri DSM 17938 Alleviates Acute Liver Injury by Activating the AMPK Signaling via Gut Microbiota-Derived Propionate. Probiotics Antimicrob. Proteins 2025. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, X.; Wang, Q.; Jin, Q.; Xu, S.; Tang, L.; Zeng, Z.; Fu, A.; Zhu, J.; Zhang, Q.; et al. Screening of Lactic Acid Bacteria Strains from Chinese Fermented Food (Suanshui) and its Protective Effect on Acute Liver Injury in Mice. Probiotics Antimicrob. Proteins 2024. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, X.; Jin, Q.; Fu, A.; Zhang, Q.; Yang, R.; Deng, B.; Li, W. Based on whole genome sequencing and metabolomics analysis the safety and probiotic characteristics of Lacticaseibacillus paracasei 36 isolated from Chinese fermented vegetables. Food Biosci. 2024, 62, 105405. [Google Scholar] [CrossRef]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Tsai, I.T.; Hsu, Y.C. Alcohol-Related Liver Disease: Basic Mechanisms and Clinical Perspectives. Int. J. Mol. Sci. 2021, 22, 5170. [Google Scholar] [CrossRef] [PubMed]
- Burra, P.; Zanetto, A.; Schnabl, B.; Reiberger, T.; Montano-Loza, A.J.; Asselta, R.; Karlsen, T.H.; Tacke, F. Hepatic immune regulation and sex disparities. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Cao, F.; Lai, S.; Zhuge, H.; Chang, K.; Valencak, T.G.; Liu, J.; Li, S.; Ren, D. Lactobacillus plantarum ZY08 relieves chronic alcohol-induced hepatic steatosis and liver injury in mice via restoring intestinal flora homeostasis. Food Res. Int. 2022, 157, 111259. [Google Scholar] [CrossRef] [PubMed]
- Chayanupatkul, M.; Somanawat, K.; Chuaypen, N.; Klaikeaw, N.; Wanpiyarat, N.; Siriviriyakul, P.; Tumwasorn, S.; Werawatganon, D. Probiotics and their beneficial effects on alcohol-induced liver injury in a rat model: The role of fecal microbiota. BMC Complement. Med. Ther. 2022, 22, 168. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, F.; Liu, Y.; Gu, Z.; Zhang, L.; Lee, J.; He, L.; Vatsalya, V.; Zhang, H.G.; Deng, Z.; et al. Probiotic-derived nanoparticles inhibit ALD through intestinal miR194 suppression and subsequent FXR activation. Hepatology 2023, 77, 1164–1180. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Li, X.; Sun, J.; Pang, X.; Sun, Q.; Lu, Y. Regulatory mechanisms of the probiotic-targeted gut-liver axis for the alleviation of alcohol-related liver disease: A review. Crit. Rev. Food Sci. Nutr. 2025, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Wang, M.; Wen, S.; Hu, L.; Lan, Y.; Xu, H. Lactiplantibacillus plantarum P101 Alleviated Alcohol-Induced Hepatic Lipid Accumulation in Mice via AMPK Signaling Pathway: Gut Microbiota and Metabolomics Analysis. Probiotics Antimicrob. Proteins 2024. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, S.; Wu, L.; Yang, K.; Yang, F.; Yang, J.; Hu, S.; Yao, Y.; Xia, X.; Liu, Y.; et al. Puerarin inhibits inflammation and lipid accumulation in alcoholic liver disease through regulating MMP8. Chin. J. Nat. Med. 2023, 21, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Raya Tonetti, F.; Eguileor, A.; Mrdjen, M.; Pathak, V.; Travers, J.; Nagy, L.E.; Llorente, C. Gut-liver axis: Recent concepts in pathophysiology in alcohol-associated liver disease. Hepatology 2024, 80, 1342–1371. [Google Scholar] [CrossRef] [PubMed]
- Rangra, S.; Rana, D.; Prajapati, A.; Benival, D.; Dwivedi, P.; Mandoli, A. Nutritional and microbiota-based therapeutic interventions for alcohol-associated liver disease: From pathogenesis to therapeutic insights. Life Sci. 2024, 352, 122852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, Q.; Zhang, Y.; Ma, W.; Ning, K.; Xiang, J.Y.; Cui, J.; Xiang, H. Combination of probiotics with different functions alleviate DSS-induced colitis by regulating intestinal microbiota, IL-10, and barrier function. Appl. Microbiol. Biotechnol. 2020, 104, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Shataer, D.; Yan, H.; Dong, X.; Zhang, M.; Qin, Y.; Cui, J.; Wang, L. Probiotics and Non-Alcoholic Fatty Liver Disease: Unveiling the Mechanisms of Lactobacillus plantarum and Bifidobacterium bifidum in Modulating Lipid Metabolism, Inflammation, and Intestinal Barrier Integrity. Foods 2024, 13, 2992. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Chen, Z.; Xie, L.; Yang, S.; Li, Y.; Wu, J.; Wu, Y.; Li, S.; Zhang, X.; Ma, Y.; et al. Lactobacillus plantarum ST-III culture supernatant protects against acute alcohol-induced liver and intestinal injury. Aging 2023, 16, 2077–2089. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.C.; Ding, X.C.; Liu, H.J.; Ma, W.L.; Feng, X.Y.; Ma, L.N. Effects of Lactobacillus paracasei N1115 on gut microbial imbalance and liver function in patients with hepatitis B-related cirrhosis. World J. Gastroenterol. 2024, 30, 1556–1571. [Google Scholar] [CrossRef] [PubMed]
- Henneke, L.; Schlicht, K.; Andreani, N.A.; Hollstein, T.; Demetrowitsch, T.; Knappe, C.; Hartmann, K.; Jensen-Kroll, J.; Rohmann, N.; Pohlschneider, D.; et al. A dietary carbohydrate-gut Parasutterella-human fatty acid biosynthesis metabolic axis in obesity and type 2 diabetes. Gut Microbes 2022, 14, 2057778. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Wu, H.; Wu, S.D.; Lu, N.; Wang, Y.T.; Liu, H.N.; Dong, L.; Liu, T.T.; Shen, X.Z. Parasutterella, in association with irritable bowel syndrome and intestinal chronic inflammation. J. Gastroenterol. Hepatol. 2018, 33, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Huang, J.; Guo, X.; Xia, J.; Hu, M. Romboutsia lituseburensis JCM1404 supplementation ameliorated endothelial function via gut microbiota modulation and lipid metabolisms alterations in obese rats. FEMS Microbiol. Lett. 2023, 370, fnad016. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Kong, J.Y.; Stothard, P.; Willing, B.P. Defining the role of Parasutterella, a previously uncharacterized member of the core gut microbiota. ISME J. 2019, 13, 1520–1534. [Google Scholar] [CrossRef] [PubMed]
- Schaffert, C.S.; Duryee, M.J.; Hunter, C.D.; Hamilton, B.C., 3rd; DeVeney, A.L.; Huerter, M.M.; Klassen, L.W.; Thiele, G.M. Alcohol metabolites and lipopolysaccharide: Roles in the development and/or progression of alcoholic liver disease. World J. Gastroenterol. 2009, 15, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yang, Y.; Ren, L.; Shao, T.; Li, F.; Zhao, C.; Liu, L.; Zhang, H.; McClain, C.J.; Feng, W. Activation of autophagy attenuates EtOH-LPS-induced hepatic steatosis and injury through MD2 associated TLR4 signaling. Sci. Rep. 2017, 7, 9292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.P.; Domingo, M.C.; Lévesque, S.; Yansouni, C.P. A case report of a deep surgical site infection with Terrisporobacter glycolicus/T. Mayombei and review of the literature. BMC Infect. Dis. 2016, 16, 529. [Google Scholar] [CrossRef] [PubMed]
- Yadegar, A.; Salahi-Niri, A.; Dai, C.; Shen, J. Editorial: Recent advances in understanding the role and mechanisms of gut microbiota in inflammatory bowel disease and irritable bowel syndrome. Front. Cell Infect. Microbiol. 2024, 14, 1521676. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; You, H.S.; Kang, H.G.; Kang, S.S.; Hyun, S.H. Association between Altered Blood Parameters and Gut Microbiota after Synbiotic Intake in Healthy, Elderly Korean Women. Nutrients 2020, 12, 3112. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jiang, Y.; Zeng, G.; Huda, N.; Thoudam, T.; Yang, Z.; Liangpunsakul, S.; Ma, J. Cell-to-cell and organ-to-organ crosstalk in the pathogenesis of alcohol-associated liver disease. eGastroenterology 2024, 2, e100104. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Ben-Zeev Brik, R.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e16. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xuan, Y.H.; Luo, M.X.; Ni, X.G.; Ling, L.Q.; Hu, S.J.; Chen, J.Q.; Xu, J.Y.; Jiang, L.Y.; Si, W.Z.; et al. Kaempferol alleviates acute alcoholic liver injury in mice by regulating intestinal tight junction proteins and butyrate receptors and transporters. Toxicology 2020, 429, 152338. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Guo, P.; Wang, Y.; Teng, X.; Zhang, H.; Sun, L.; Xue, M.; Liang, H. Propolis Ameliorates Alcohol-Induced Depressive Symptoms in C57BL/6J Mice by Regulating Intestinal Mucosal Barrier Function and Inflammatory Reaction. Nutrients 2022, 14, 1213. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.C.; Xu, L.M.; Du, S.J.; Huang, S.S.; Wu, H.; Dong, J.J.; Huang, J.R.; Wang, X.D.; Feng, W.K.; Chen, Y.P. Lactobacillus rhamnosus GG supernatant promotes intestinal barrier function, balances Treg and TH17 cells and ameliorates hepatic injury in a mouse model of chronic-binge alcohol feeding. Toxicol. Lett. 2016, 241, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, X.; Ren, H.; Chang, J.; Guo, J.; He, Z.; Shi, R.; Hu, X.; Jin, Y.; Lu, S.; et al. Lactobacillus paracasei Jlus66 relieves DSS-induced ulcerative colitis in a murine model by maintaining intestinal barrier integrity, inhibiting inflammation, and improving intestinal microbiota structure. Eur. J. Nutr. 2024, 63, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Ross, F.A.; MacKintosh, C.; Hardie, D.G. AMP-activated protein kinase: A cellular energy sensor that comes in 12 flavours. FEBS J. 2016, 283, 2987–3001. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Wang, C.; Zhao, H.; Zhao, C.; Chen, Y.; Wang, Y.; McClain, C.; Feng, W. Enhanced AMPK phosphorylation contributes to the beneficial effects of Lactobacillus rhamnosus GG supernatant on chronic-alcohol-induced fatty liver disease. J. Nutr. Biochem. 2015, 26, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.J.; Cui, R.; Hu, T.G.; Wu, H. Hypoglycemic effects of Lactiplantibacillus plantarum B19 via promoting AMPK/PI3K/AKT signaling pathway. Food Biosci. 2025, 68, 106414. [Google Scholar] [CrossRef]
- Xu, H.; Luo, J.; Ma, G.; Zhang, X.; Yao, D.; Li, M.; Loor, J.J. Acyl-CoA synthetase short-chain family member 2 (ACSS2) is regulated by SREBP-1 and plays a role in fatty acid synthesis in caprine mammary epithelial cells. J. Cell Physiol. 2018, 233, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Gomez-Llorente, C.; Fontana, L.; Gil, A. Modulation of immunity and inflammatory gene expression in the gut, in inflammatory diseases of the gut and in the liver by probiotics. World J. Gastroenterol. 2014, 20, 15632–15649. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Zhang, X.; Kuang, M.; Yu, J. The gut-liver axis in immune remodeling of hepatic cirrhosis. Front. Immunol. 2022, 13, 946628. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Chen, X.; Wang, F.; Chen, T.; Yin, M.; Liu, Z.; Li, W.; Zhu, J. Lacticaseibacillus paracasei 36 Mitigates Alcoholic-Associated Liver Disease Through Modulation of Microbiota and AMPK Signaling. Nutrients 2025, 17, 2340. https://doi.org/10.3390/nu17142340
Wang C, Chen X, Wang F, Chen T, Yin M, Liu Z, Li W, Zhu J. Lacticaseibacillus paracasei 36 Mitigates Alcoholic-Associated Liver Disease Through Modulation of Microbiota and AMPK Signaling. Nutrients. 2025; 17(14):2340. https://doi.org/10.3390/nu17142340
Chicago/Turabian StyleWang, Chongyu, Xi Chen, Fei Wang, Tianyu Chen, Mengqiu Yin, Ziyu Liu, Weifen Li, and Jinhui Zhu. 2025. "Lacticaseibacillus paracasei 36 Mitigates Alcoholic-Associated Liver Disease Through Modulation of Microbiota and AMPK Signaling" Nutrients 17, no. 14: 2340. https://doi.org/10.3390/nu17142340
APA StyleWang, C., Chen, X., Wang, F., Chen, T., Yin, M., Liu, Z., Li, W., & Zhu, J. (2025). Lacticaseibacillus paracasei 36 Mitigates Alcoholic-Associated Liver Disease Through Modulation of Microbiota and AMPK Signaling. Nutrients, 17(14), 2340. https://doi.org/10.3390/nu17142340