
Brain-Derived Neurotrophic Factor as a Potential Mediator of the Beneficial Effects of Myo-Inositol Supplementation during Suckling in the Offspring of Gestational-Calorie-Restricted Rats

 ,
,  ,
,  and
and 
Highlights
- Myo-inositol supplementation during suckling enhances BDNF signaling in the offspring of gestational calorie-restricted dams, in a sex-dependent manner;
- Myo-inositol supplementation promotes anorexigenic pathways in the hypothalamus, increases browning markers, and enhances adipose tissues’ glucose uptake capacity;
- BDNF is suggested as a key mediator of the metabolic benefits of myo-inositol supplementation during suckling.
Abstract
1. Introduction
2. Materials and Methods
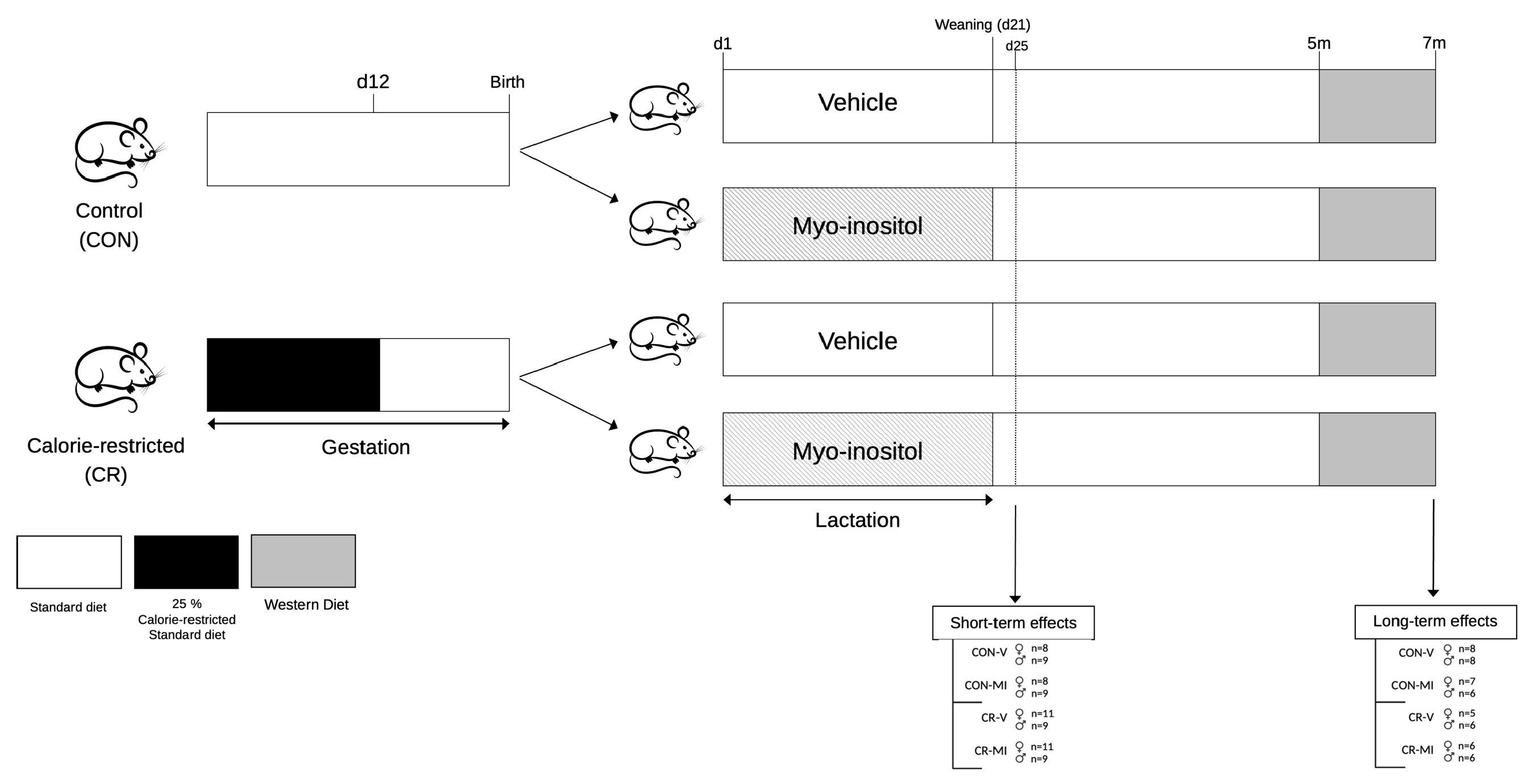
2.1. Animals
- Study 1. Short-term effects of myo-inositol supplementation during suckling in the offspring of gestational-calorie-restricted rats
- Study 2. Long-term effects of myo-inositol supplementation during lactation in the offspring of gestational-calorie-restricted rats
2.2. BDNF Quantification by Enzyme-Linked Immunosorbent Assay (ELISA)
2.3. Gene Expression Analyses
2.4. Protein Expression Analyses
2.5. Statistical Analysis
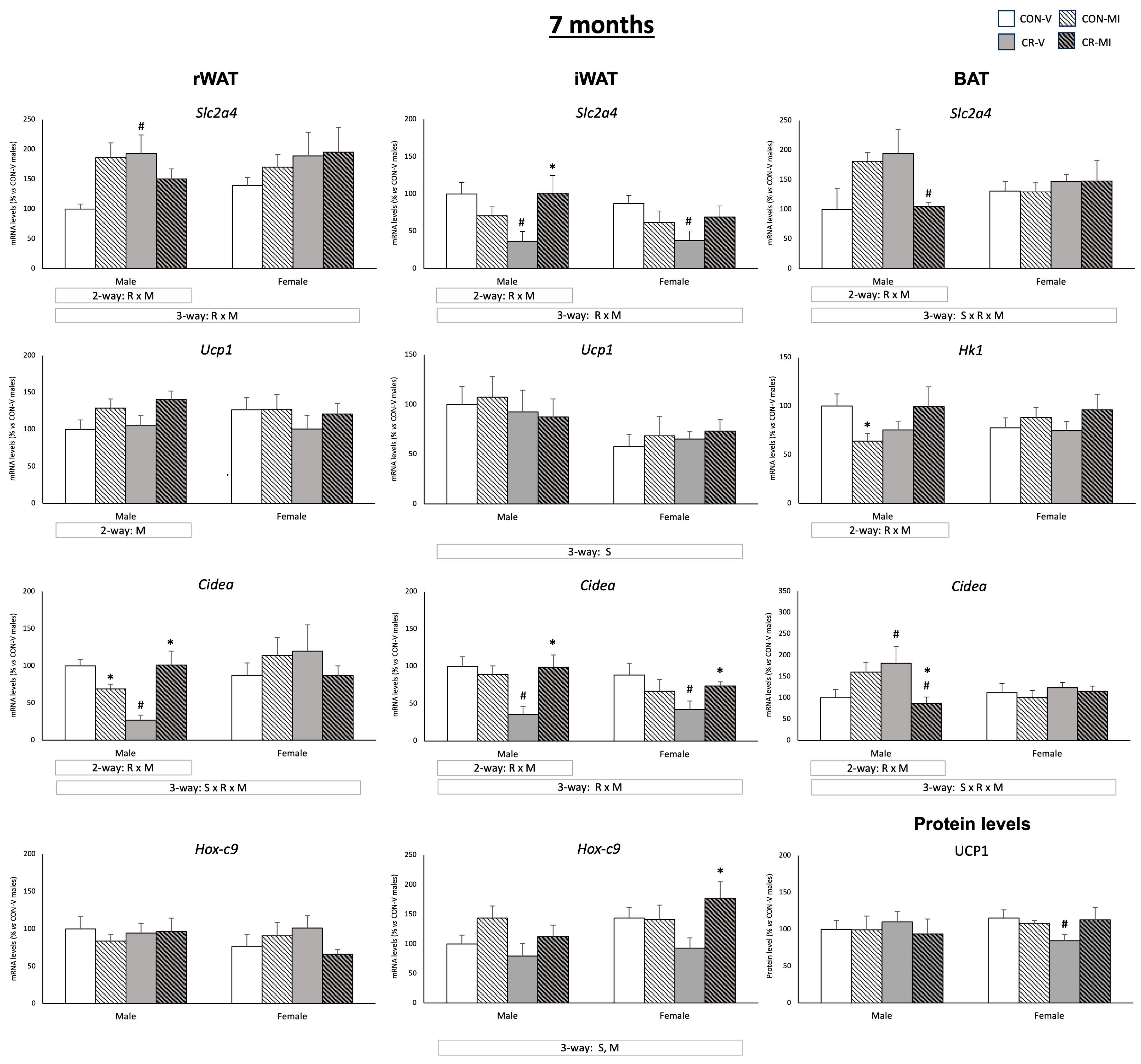
3. Results
3.1. Weight-Related Parameters
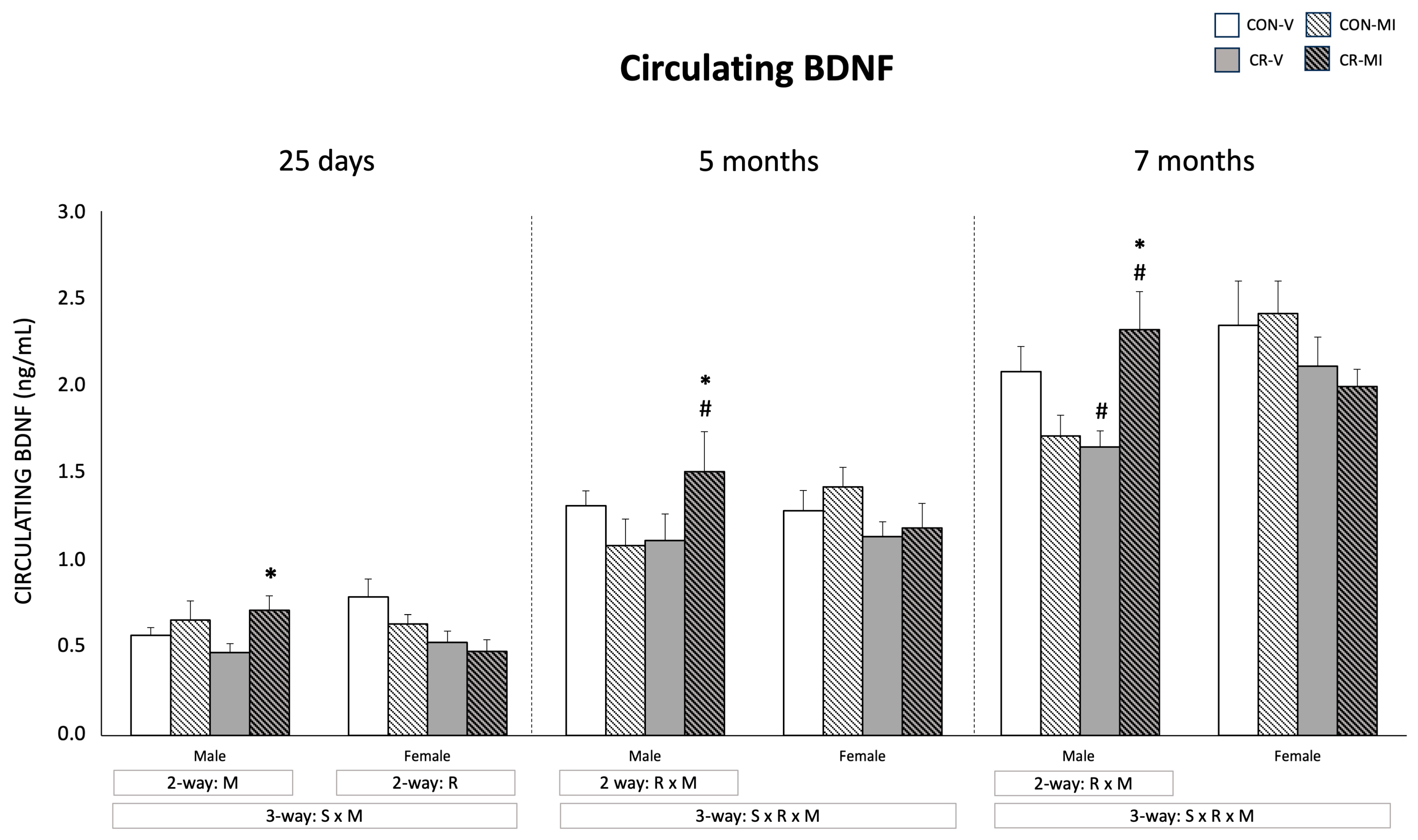
3.2. Circulating BDNF
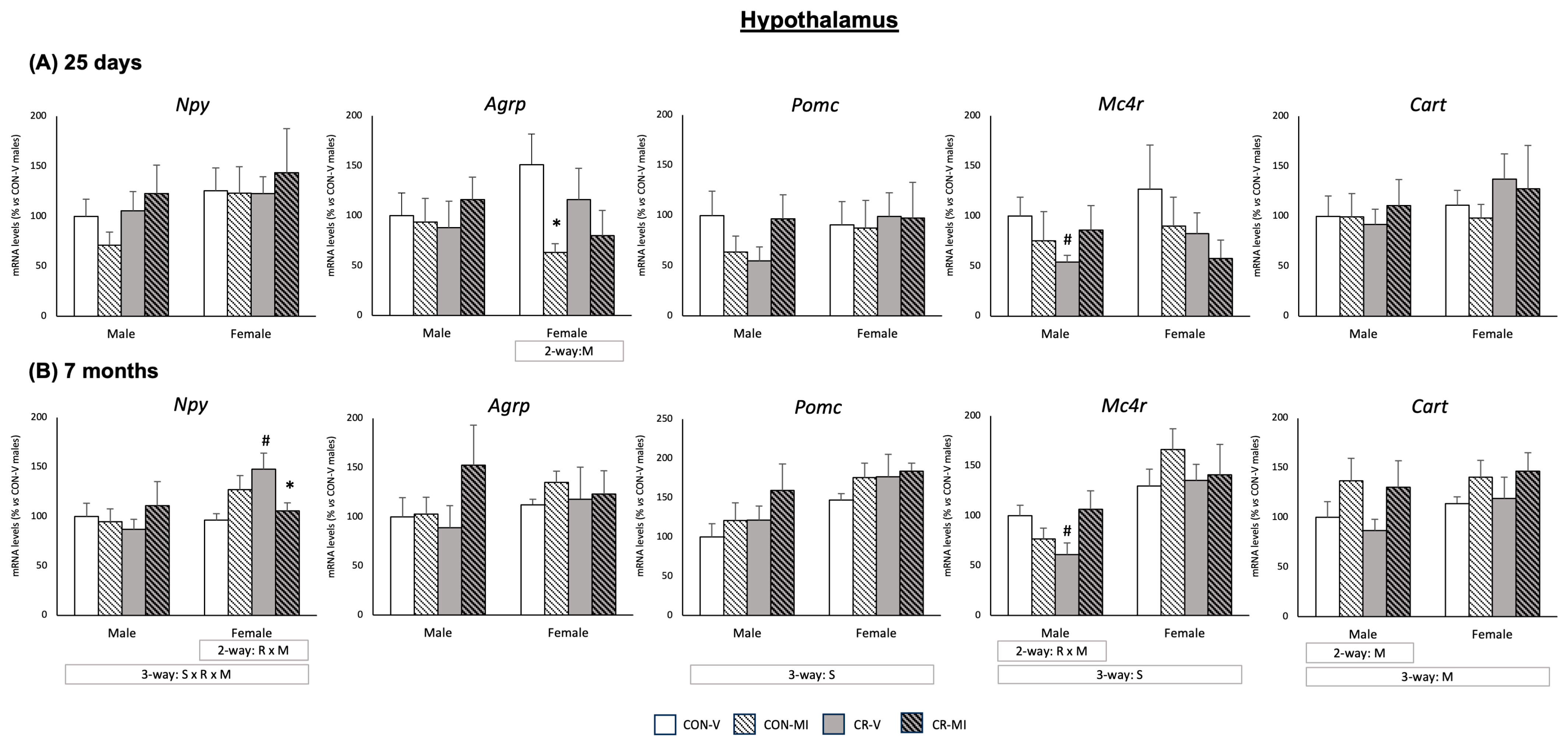
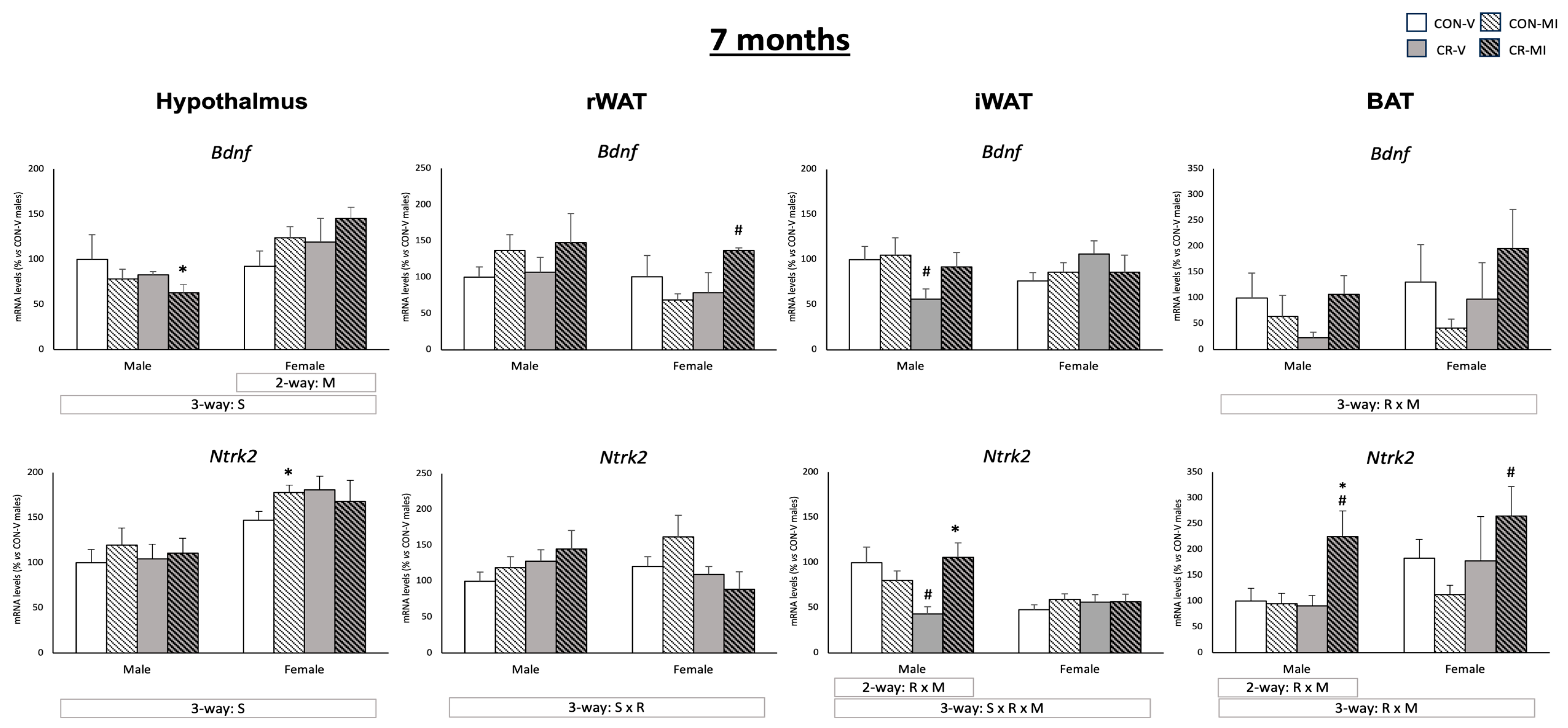
3.3. Gene and Protein Expression Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Plagemann, A.; Harder, T.; Schellong, K.; Schulz, S.; Stupin, J.H. Early Postnatal Life as a Critical Time Window for Determination of Long-Term Metabolic Health. Best Pr. Res. Clin. Endocrinol. Metab. 2012, 26, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Picó, C.; Palou, M.; Priego, T.; Sánchez, J.; Palou, A. Metabolic Programming of Obesity by Energy Restriction during the Perinatal Period: Different Outcomes Depending on Gender and Period, Type and Severity of Restriction. Front. Physiol. 2012, 3, 436. [Google Scholar] [CrossRef] [PubMed]
- Langley-Evans, S.C. Developmental Programming of Health and Disease. Proc. Nutr. Soc. 2006, 65, 97. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. The Developing World of DOHaD. J. Dev. Orig. Health Dis. 2018, 9, 266–269. [Google Scholar] [CrossRef]
- Barker, D.J.P. Fetal Origins of Coronary Heart Disease. Br. Heart J. 1993, 69, 195. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, J.C.; Lisboa, P.C.; de Moura, E.G.; Barella, L.F.; Miranda, R.A.; Malta, A.; da Silva Franco, C.C.; da Silva Ribeiro, T.A.; Torrezan, R.; Gravena, C.; et al. Poor Pubertal Protein Nutrition Disturbs Glucose-Induced Insulin Secretion Process in Pancreatic Islets and Programs Rats in Adulthood to Increase Fat Accumulation. J. Endocrinol. 2013, 216, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.L.; Loret De Mola, C.; Victora, C.G. Long-Term Consequences of Breastfeeding on Cholesterol, Obesity, Systolic Blood Pressure and Type 2 Diabetes: A Systematic Review and Meta-Analysis. Acta Paediatr. 2015, 104, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Dai, L.J.; Zhang, Q.; Ouyang, Y.Q. A Meta-Analysis of the Association Between Breastfeeding and Early Childhood Obesity. J. Pediatr. Nurs. 2020, 53, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.L. Developmental Origins of Obesity: Early Feeding Environments, Infant Growth, and the Intestinal Microbiome. Am. J. Hum. Biol. 2012, 24, 350–360. [Google Scholar] [CrossRef]
- Palou, M.; Picó, C.; Palou, A. Leptin as a Breast Milk Component for the Prevention of Obesity. Nutr Rev. 2018, 76, 875–892. [Google Scholar] [CrossRef]
- Picó, C.; Oliver, P.; Sánchez, J.; Miralles, O.; Caimari, A.; Priego, T.; Palou, A. The Intake of Physiological Doses of Leptin during Lactation in Rats Prevents Obesity in Later Life. Int. J. Obes. 2007, 31, 1199–1209. [Google Scholar] [CrossRef]
- Konieczna, J.; García, A.P.; Sánchez, J.; Palou, M.; Palou, A.; Picó, C. Oral Leptin Treatment in Suckling Rats Ameliorates Detrimental Effects in Hypothalamic Structure and Function Caused by Maternal Caloric Restriction during Gestation. PLoS ONE 2013, 8, 81906. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.; Palou, M.; Otero, D.; Núñez, P.; Palou, A.; Picó, C.; Castillo, P.; Palou, M.; Otero, D.; Núñez, P.; et al. Sex-Specific Effects of Myo-Inositol Ingested During Lactation in the Improvement of Metabolic Health in Adult Rats. Mol. Nutr. Food Res. 2021, 65, 2000965. [Google Scholar] [CrossRef] [PubMed]
- Palou, M.; Torrens, J.M.; Castillo, P.; Sánchez, J.; Palou, A.; Picó, C. Metabolomic Approach in Milk from Calorie-Restricted Rats during Lactation: A Potential Link to the Programming of a Healthy Phenotype in Offspring. Eur. J. Nutr. 2020, 59, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.R.; Baker, L.; Egler, J.; Corcoran, L.; Chiavacci, R. Serum Myoinositol Concentrations in Premature Infants Fed Human Milk, Formula for Infants, and Parenteral Nutrition. Am. J. Clin. Nutr. 1990, 51, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Bromberger, P.; Hallman, M. Myoinositol in Small Preterm Infants: Relationship between Intake and Serum Concentration. J. Pediatr. Gastroenterol. Nutr. 1986, 5, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.; Palou, M.; Yau-Qiu, Z.X.; Rodríguez, A.M.; Palou, A.; Picó, C. Myo-Inositol Supplementation in Suckling Rats Protects against Adverse Programming Outcomes on Hypothalamic Structure Caused by Mild Gestational Calorie Restriction, Partially Comparable to Leptin Effects. Nutrients 2021, 13, 3257. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol. Neurobiol. 2018, 38, 579. [Google Scholar] [CrossRef] [PubMed]
- Iu, E.C.Y.; Chan, C.B. Is Brain-Derived Neurotrophic Factor a Metabolic Hormone in Peripheral Tissues? Biology 2022, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.C.; Zimbone, S.; Saab, M.W.; Tomasello, M.F. The Pleiotropic Potential of BDNF beyond Neurons: Implication for a Healthy Mind in a Healthy Body. Life 2021, 11, 1256. [Google Scholar] [CrossRef]
- Colitti, M.; Montanari, T. Brain-Derived Neurotrophic Factor Modulates Mitochondrial Dynamics and Thermogenic Phenotype on 3T3-L1 Adipocytes. Tissue Cell 2020, 66, 101388. [Google Scholar] [CrossRef] [PubMed]
- McMurphy, T.; Huang, W.; Liu, X.; Siu, J.J.; Queen, N.J.; Xiao, R.; Cao, L. Hypothalamic Gene Transfer of BDNF Promotes Healthy Aging in Mice. Aging Cell 2019, 18, e12846. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, M.; Tsuchida, A.; Nakagawa, T.; Nonomura, T.; Ono-Kishino, M.; Sugaru, E.; Noguchi, H.; Taiji, M. Brain-Derived Neurotrophic Factor Enhances Glucose Utilization in Peripheral Tissues of Diabetic Mice. Diabetes Obes. Metab. 2007, 9, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ravelli, G.-P.; Stein, Z.A.; Susser, M.W. Obesity in Young Men after Famine Exposure in Utero and Early Infancy. N. Engl. J. Med. 1976, 295, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Nishimura, M.; Yajima, T.; Kuwata, T.; Suzuki, Y.I.; Goda, T.; Takase, S.; Harada, E. Effect of Intermittent Feeding on the Development of Disaccharidase Activities in Artificially Reared Rat Pups. Comp. Biochem. Physiol.—A Mol. Integr. Physiol. 1998, 121, 289–297. [Google Scholar] [CrossRef]
- Timper, K.; Brüning, J.C. Hypothalamic Circuits Regulating Appetite and Energy Homeostasis: Pathways to Obesity. Dis. Model Mech. 2017, 10, 679. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The Human Milk Metabolome Reveals Diverse Oligosaccharide Profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, Y.L.; Wang, L.P.; Liu, X.C. Impact of Different Stereoisomers of Inositol on Insulin Sensitivity of Gestational Diabetes Mellitus Patients. World J. Clin. Cases 2021, 9, 565. [Google Scholar] [CrossRef]
- Brown, J.; Crawford, T.J.; Alsweiler, J.; Crowther, C.A. Dietary Supplementation with Myo-inositol in Women during Pregnancy for Treating Gestational Diabetes. Cochrane Database Syst. Rev. 2016, 2016, CD012048. [Google Scholar] [CrossRef]
- Fraticelli, F.; Celentano, C.; Zecca, I.A.; Di Vieste, G.; Pintaudi, B.; Liberati, M.; Franzago, M.; Di Nicola, M.; Vitacolonna, E. Effect of Inositol Stereoisomers at Different Dosages in Gestational Diabetes: An Open-Label, Parallel, Randomized Controlled Trial. Acta Diabetol. 2018, 55, 805–812. [Google Scholar] [CrossRef]
- Akashi, M.; Akazawa, S.; Akazawa, M.; Trocino, R.; Hashimoto, M.; Maeda, Y.; Yamamoto, H.; Kawasaki, E.; Takino, H.; Yokota, A.; et al. Effects of Insulin and Myo-Inositol on Embryo Growth and Development during Early Organogenesis in Streptozocin-Induced Diabetic Rats. Diabetes 1991, 40, 1574–1579. [Google Scholar] [CrossRef] [PubMed]
- D’Anna, R.; Corrado, F.; Loddo, S.; Gullo, G.; Giunta, L.; Di Benedetto, A. Myoinositol plus α-Lactalbumin Supplementation, Insulin Resistance and Birth Outcomes in Women with Gestational Diabetes Mellitus: A Randomized, Controlled Study. Sci. Rep. 2021, 11, 8866. [Google Scholar] [CrossRef] [PubMed]
- Croze, M.L.; Vella, R.E.; Pillon, N.J.; Soula, H.A.; Hadji, L.; Guichardant, M.; Soulage, C.O. Chronic Treatment with Myo-Inositol Reduces White Adipose Tissue Accretion and Improves Insulin Sensitivity in Female Mice. J. Nutr. Biochem. 2013, 24, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Dang, N.T.; Mukai, R.; Yoshida, K.I.; Ashida, H. D-Pinitol and Myo-Inositol Stimulate Translocation of Glucose Transporter 4 in Skeletal Muscle of C57BL/6 Mice. Biosci. Biotechnol. Biochem. 2010, 74, 1062–1067. [Google Scholar] [CrossRef]
- Ortmeyer, H.K. Dietary Myoinositol Results in Lower Urine Glucose and in Lower Postprandial Plasma Glucose in Obese Insulin Resistant Rhesus Monkeys. Obes. Res. 1996, 4, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Eyileten, C.; Kaplon-Cieslicka, A.; Mirowska-Guzel, D.; Malek, L.; Postula, M. Antidiabetic Effect of Brain-Derived Neurotrophic Factor and Its Association with Inflammation in Type 2 Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 2823671. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.K.; Kim, J.G.; Lee, B.J. Participation of the Central Melanocortin System in Metabolic Regulation and Energy Homeostasis. Cell Mol. Life Sci. 2014, 71, 3799–3809. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.D. Anatomy and Regulation of the Central Melanocortin System. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef]
- Obici, S.; Feng, Z.; Tan, J.; Liu, L.; Karkanias, G.; Rossetti, L. Central Melanocortin Receptors Regulate Insulin Action. J. Clin. Investig. 2001, 108, 1079. [Google Scholar] [CrossRef]
- Heijboer, A.C.; Van Den Hoek, A.M.; Pijl, H.; Voshol, P.J.; Havekes, L.M.; Romijn, J.A.; Corssmit, E.P.M. Intracerebroventricular Administration of Melanotan II Increases Insulin Sensitivity of Glucose Disposal in Mice. Diabetologia 2005, 48, 1621–1626. [Google Scholar] [CrossRef][Green Version]
- Fan, W.; Dinulescu, D.M.; Butler, A.A.; Zhou, J.; Marks, D.L.; Cone, R.D. The Central Melanocortin System Can Directly Regulate Serum Insulin Levels. Endocrinology 2000, 141, 3072–3079. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.E.; Turnbull, A.V.; White, A. Pro-Opiomelanocortin Processing in the Hypothalamus: Impact on Melanocortin Signalling and Obesity. J. Endocrinol. 2002, 172, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Vanevski, F.; Xu, B. Molecular and Neural Bases Underlying Roles of BDNF in the Control of Body Weight. Front. Neurosci. 2013, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.Y.; An, J.J.; Gharami, K.; Waterhouse, E.G.; Vanevski, F.; Jones, K.R.; Xu, B. Dendritically Targeted Bdnf MRNA Is Essential for Energy Balance and Response to Leptin. Nat. Med. 2012, 18, 564. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Loh, K.H.; Wu, M.; Morgan, D.A.; Schneeberger, M.; Yu, X.; Chi, J.; Kosse, C.; Kim, D.; Rahmouni, K.; et al. A Leptin–BDNF Pathway Regulating Sympathetic Innervation of Adipose Tissue. Nature 2020, 583, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Ruud, J.; Steculorum, S.M.; Bruning, J.C. Neuronal Control of Peripheral Insulin Sensitivity and Glucose Metabolism. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, Z.; Kalyani, M.; Janik, J.M.; Shi, H. Effects of Energy Status and Diet on Bdnf Expression in the Ventromedial Hypothalamus of Male and Female Rats. Physiol. Behav. 2014, 130, 99. [Google Scholar] [CrossRef] [PubMed]
- Glud, M.; Christiansen, T.; Larsen, L.H.; Richelsen, B.; Bruun, J.M. Changes in Circulating BDNF in Relation to Sex, Diet, and Exercise: A 12-Week Randomized Controlled Study in Overweight and Obese Participants. J. Obes. 2019, 2019, 4537274. [Google Scholar] [CrossRef]
- Keifer, J.; Zheng, Z.; Ambigapathy, G. A MicroRNA-BDNF Negative Feedback Signaling Loop in Brain: Implications for Alzheimer’s Disease. MicroRNA 2015, 4, 101–108. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose Tissue in Control of Metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 178543. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Choi, E.Y.; Liu, X.; Martin, A.; Wang, C.; Xu, X.; During, M.J. White to Brown Fat Phenotypic Switch Induced by Genetic and Environmental Activation of a Hypothalamic-Adipocyte Axis. Cell Metab. 2011, 14, 324–338. [Google Scholar] [CrossRef]
- Wajchenberg, B.L.; Giannella-Neto, D.; Da Silva, M.E.R.; Santos, R.F. Depot-Specific Hormonal Characteristics of Subcutaneous and Visceral Adipose Tissue and Their Relation to the Metabolic Syndrome. Horm. Metab. Res. 2002, 34, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Mössenböck, K.; Vegiopoulos, A.; Rose, A.J.; Sijmonsma, T.P.; Herzig, S.; Schafmeier, T. Browning of White Adipose Tissue Uncouples Glucose Uptake from Insulin Signaling. PLoS ONE 2014, 9, 110428. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, J.; Dai, H.; Duan, Y.; An, Y.; Shi, L.; Lv, Y.; Li, H.; Wang, C.; Ma, Q.; et al. Brown and Beige Adipose Tissue: A Novel Therapeutic Strategy for Obesity and Type 2 Diabetes Mellitus. Adipocyte 2021, 10, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Brune, J.E.; Kern, M.; Kunath, A.; Flehmig, G.; Schön, M.R.; Lohmann, T.; Dressler, M.; Dietrich, A.; Fasshauer, M.; Kovacs, P.; et al. Fat Depot-Specific Expression of HOXC9 and HOXC10 May Contribute to Adverse Fat Distribution and Related Metabolic Traits. Obesity 2016, 24, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Palou, M.; Priego, T.; Sánchez, J.; Rodríguez, A.M.; Palou, A.; Picó, C. Gene Expression Patterns in Visceral and Subcutaneous Adipose Depots in Rats Are Linked to Their Morphologic Features. Cell. Physiol. Biochem. 2009, 24, 547–556. [Google Scholar] [CrossRef]
- Palou, M.; Sánchez, J.; Priego, T.; Rodríguez, A.M.; Picó, C.; Palou, A. Regional Differences in the Expression of Genes Involved in Lipid Metabolism in Adipose Tissue in Response to Short- and Medium-Term Fasting and Refeeding. J. Nutr. Biochem. 2010, 21, 23–33. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MALE | FEMALE | 3-Way | |||||||
|---|---|---|---|---|---|---|---|---|---|
| V | MI | 2-Way | V | MI | 2-Way | ||||
| Body weight | g | CON | 63.0 ± 1.4 | 64.7 ± 1.3 | R | 62.5 ± 1.2 | 59.8 ± 1.3 | R | R |
| CR | 57.2 ± 2.3 # | 57.1 ± 2.6 # | 58.9 ± 1.9 | 54.9 ± 2.5 | |||||
| Body fat | % | CON | 9.77 ± 0.34 | 9.9 ± 0.36 | - | 9.85 ± 0.17 | 9.58 ± 0.21 | - | R |
| CR | 9.20 ± 0.59 | 8.68 ± 0.62 | 9.40 ± 0.46 | 8.53 ± 0.51 | |||||
| White adipose tissue | Inguinal (g) | CON | 0.701 ± 0.034 | 0.787 ± 0.068 | R | 0.845 ± 0.068 | 0.775 ± 0.037 | R | R |
| CR | 0.648 ± 0.060 | 0.620 ± 0.053 | 0.679 ± 0.064 | 0.587 ± 0.064 # | |||||
| Retroperitoneal (g) | CON | 0.140 ± 0.013 | 0.135 ± 0.016 | - | 0.110 ± 0.012 | 0.114 ± 0.014 | R | S, R | |
| CR | 0.110 ± 0.010 | 0.109 ± 0.008 | 0.093 ± 0.009 | 0.079 ± 0.012 | |||||
| Brown adipose tissue | g | CON | 0.270 ± 0.024 | 0.288 ± 0.037 | - | 0.275 ± 0.032 | 0.258 ± 0.025 | - | - |
| CR | 0.258 ± 0.023 | 0.250 ± 0.023 | 0.254 ± 0.022 | 0.252 ± 0.018 | |||||
| MALE | FEMALE | 3-Way | |||||||
|---|---|---|---|---|---|---|---|---|---|
| V | MI | 2-Way | V | MI | 2-Way | ||||
| Body weight | g | CON | 567 ± 15 | 595 ± 10 | - | 303 ± 6 | 301 ± 9 | - | S |
| CR | 559 ± 17 | 557 ± 15 | 293 ± 8 | 314 ± 11 | |||||
| Body fat | % | CON | 25.1 ± 0.8 | 22.5 ± 1.5 | - | 22.3 ± 1.2 | 21.9 ± 1.8 | - | - |
| CR | 24.1 ± 2.0 | 24.0 ± 1.4 | 23.2 ± 1.8 | 24.7 ± 1.9 | |||||
| White adipose tissue | Inguinal (g) | CON | 21.7 ± 1.6 | 21.7 ± 2.1 | - | 7.79 ± 0.46 | 7.27 ± 0.81 | - | S |
| CR | 21.6 ± 2.0 | 21.5 ± 2.2 | 7.59 ± 0.47 | 8.63 ± 0.77 | |||||
| Retroperitoneal (g) | CON | 22.5 ± 1.5 | 20.9 ± 2.7 | - | 8.28 ± 0.49 | 7.08 ± 0.93 | - | S × R | |
| CR | 21.2 ± 2.1 | 19.7 ± 1.0 | 8.87 ± 0.93 | 8.76 ± 0.92 | |||||
| Brown adipose tissue | g | CON | 1.06 ± 0.06 | 1.19 ± 0.15 | R | 0.689 ± 0.026 | 0.659 ± 0.072 | R | S × R |
| CR | 0.816 ± 0.050 # | 0.885 ± 0.052 | 0.709 ± 0.034 | 0.836 ± 0.008 * | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valle, A.; Castillo, P.; García-Rodríguez, A.; Palou, A.; Palou, M.; Picó, C. Brain-Derived Neurotrophic Factor as a Potential Mediator of the Beneficial Effects of Myo-Inositol Supplementation during Suckling in the Offspring of Gestational-Calorie-Restricted Rats. Nutrients 2024, 16, 980. https://doi.org/10.3390/nu16070980
Valle A, Castillo P, García-Rodríguez A, Palou A, Palou M, Picó C. Brain-Derived Neurotrophic Factor as a Potential Mediator of the Beneficial Effects of Myo-Inositol Supplementation during Suckling in the Offspring of Gestational-Calorie-Restricted Rats. Nutrients. 2024; 16(7):980. https://doi.org/10.3390/nu16070980
Chicago/Turabian StyleValle, Ana, Pedro Castillo, Adrián García-Rodríguez, Andreu Palou, Mariona Palou, and Catalina Picó. 2024. "Brain-Derived Neurotrophic Factor as a Potential Mediator of the Beneficial Effects of Myo-Inositol Supplementation during Suckling in the Offspring of Gestational-Calorie-Restricted Rats" Nutrients 16, no. 7: 980. https://doi.org/10.3390/nu16070980
APA StyleValle, A., Castillo, P., García-Rodríguez, A., Palou, A., Palou, M., & Picó, C. (2024). Brain-Derived Neurotrophic Factor as a Potential Mediator of the Beneficial Effects of Myo-Inositol Supplementation during Suckling in the Offspring of Gestational-Calorie-Restricted Rats. Nutrients, 16(7), 980. https://doi.org/10.3390/nu16070980