
5-Heptadecylresorcinol Improves Aging-Associated Hepatic Fatty Acid Oxidation Dysfunction via Regulating Adipose Sirtuin 3



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animal and Experimental Design
2.3. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
2.4. Biochemical Analysis of Serum
2.5. Histological Analysis
2.6. RNA Isolation and qRT-PCR Analysis
2.7. Western Blotting
2.8. Acylcarnitine Analysis
2.9. Statistical Analysis
3. Results
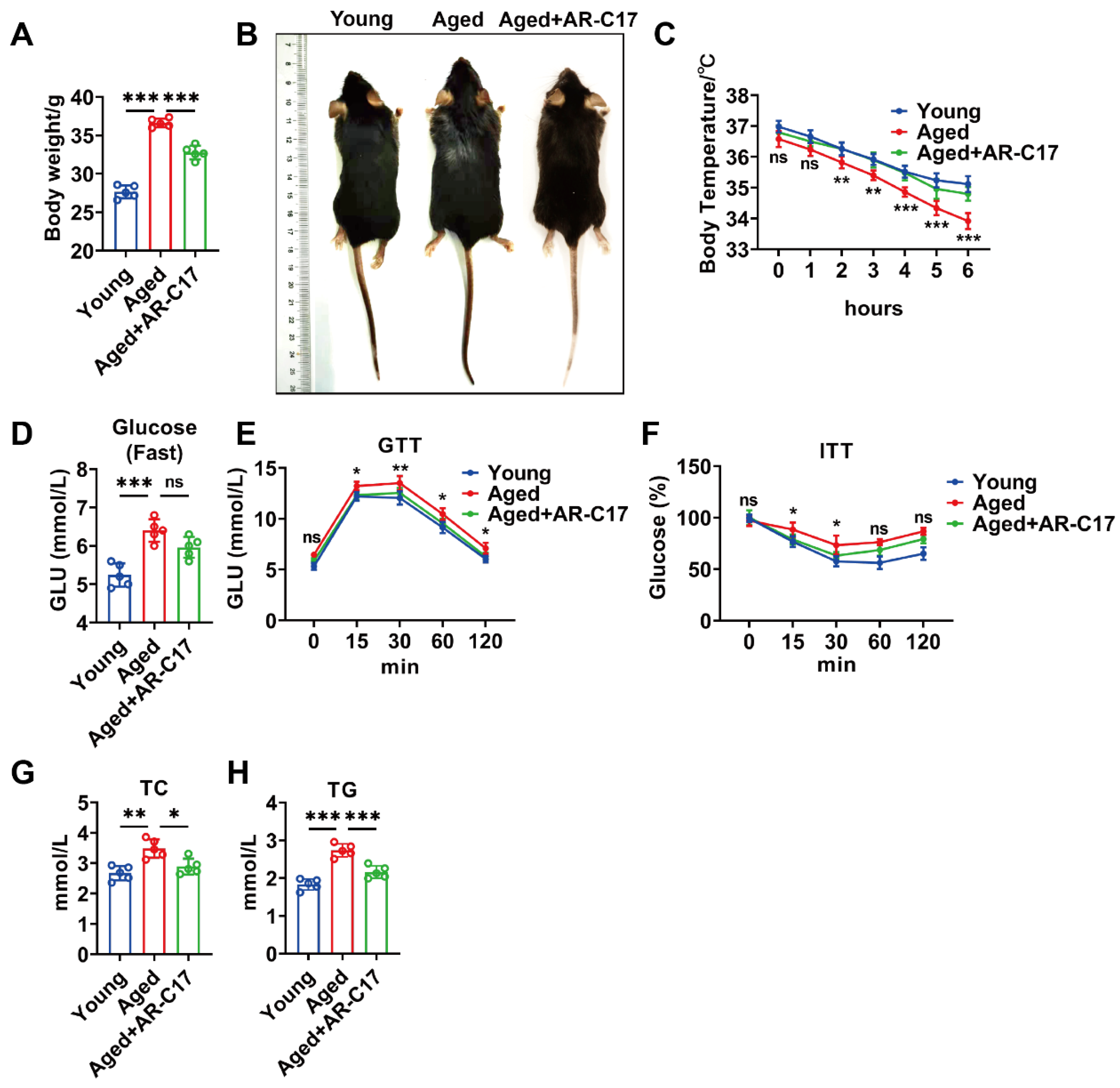
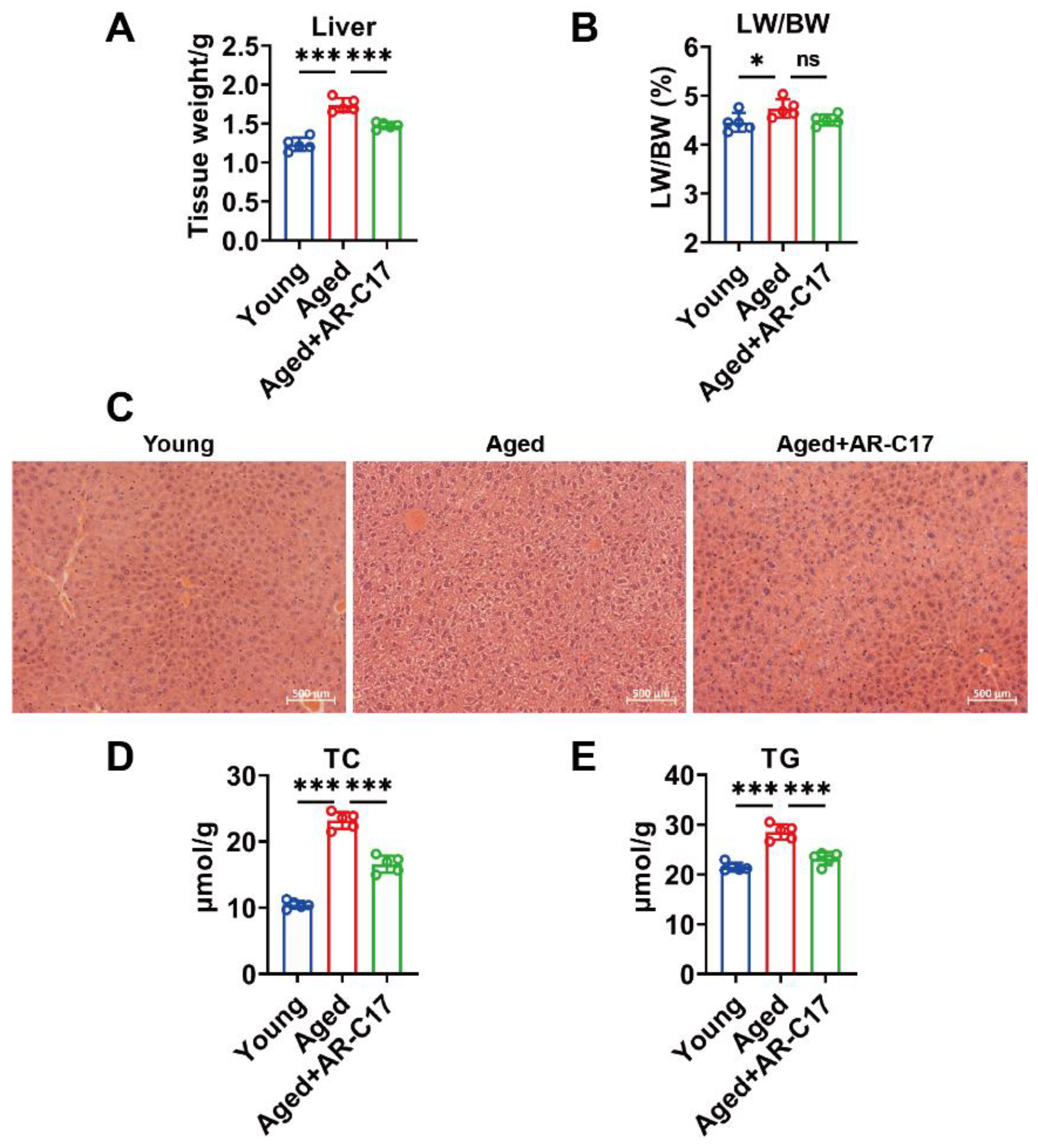
3.1. AR-C17 Improved Glucose and Lipid Metabolism during Aging
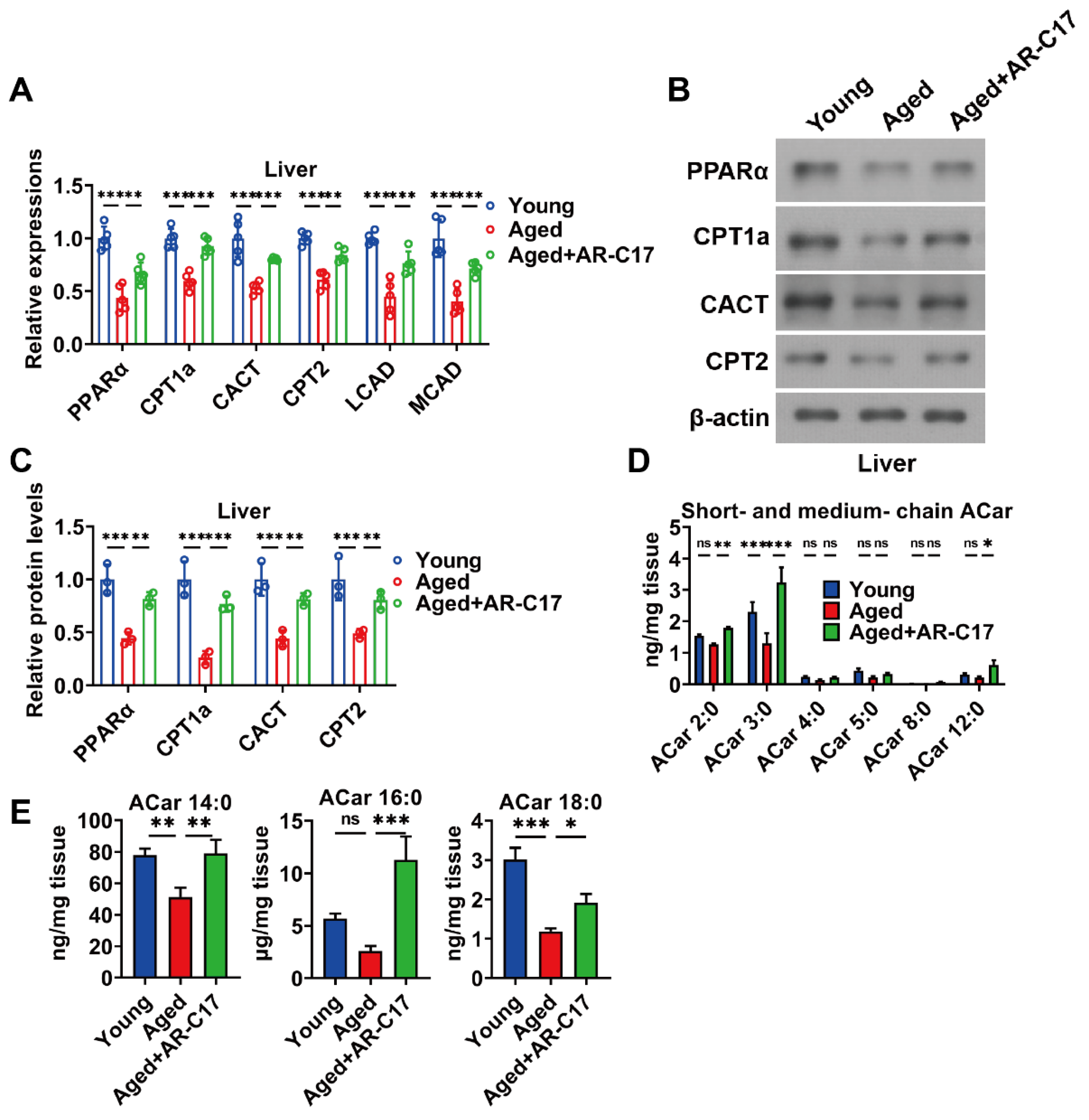
3.2. AR-C17 Improved Hepatic Fatty Acid Oxidation during Aging
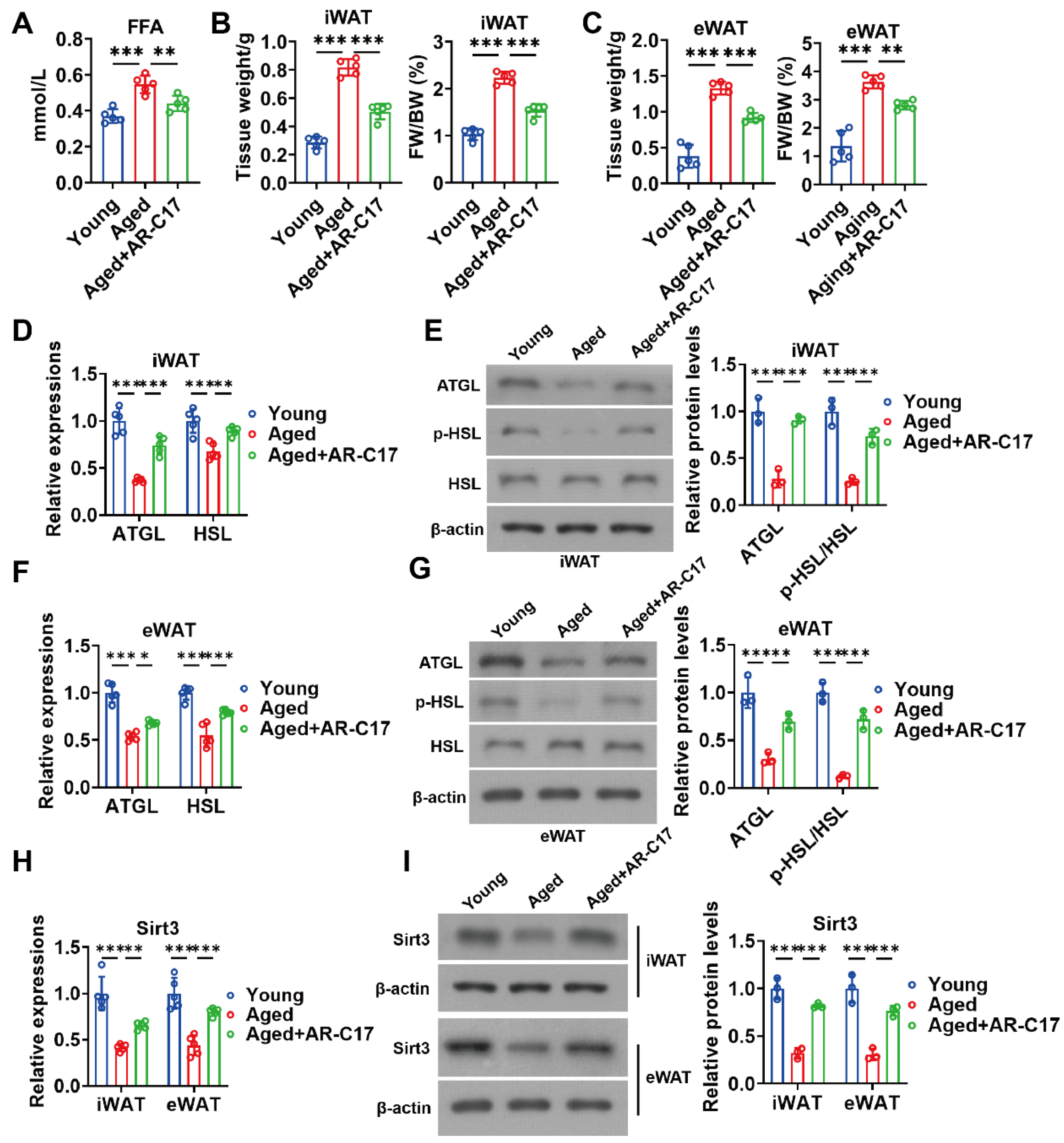
3.3. AR-C17 Improved WAT Lipolysis and Increased Adipose Sirt3 Expressions during Aging
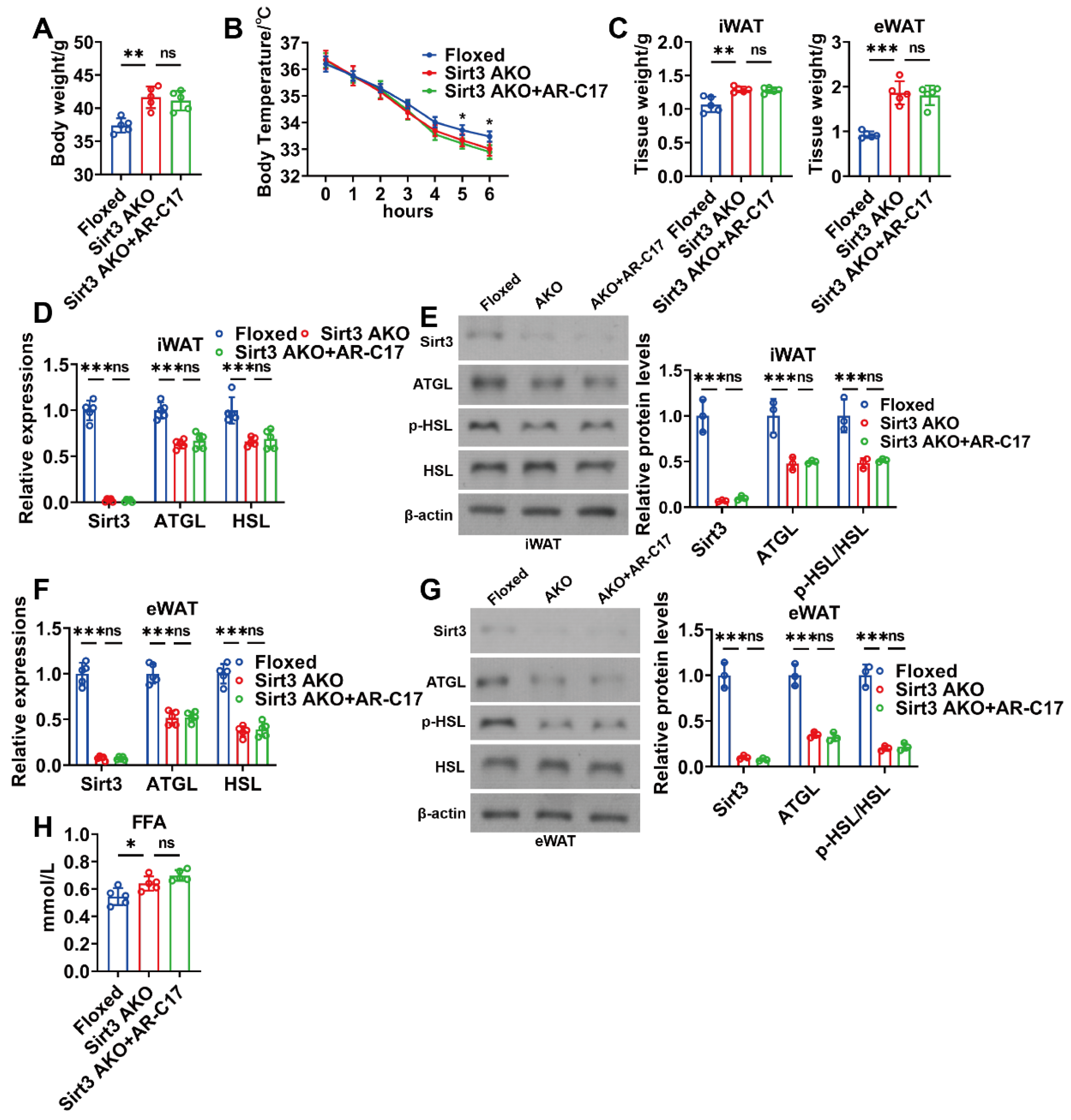
3.4. AR-C17 Failed to Reinforce WAT Lipolysis during Aging in the Absence of Adipose Sirt3
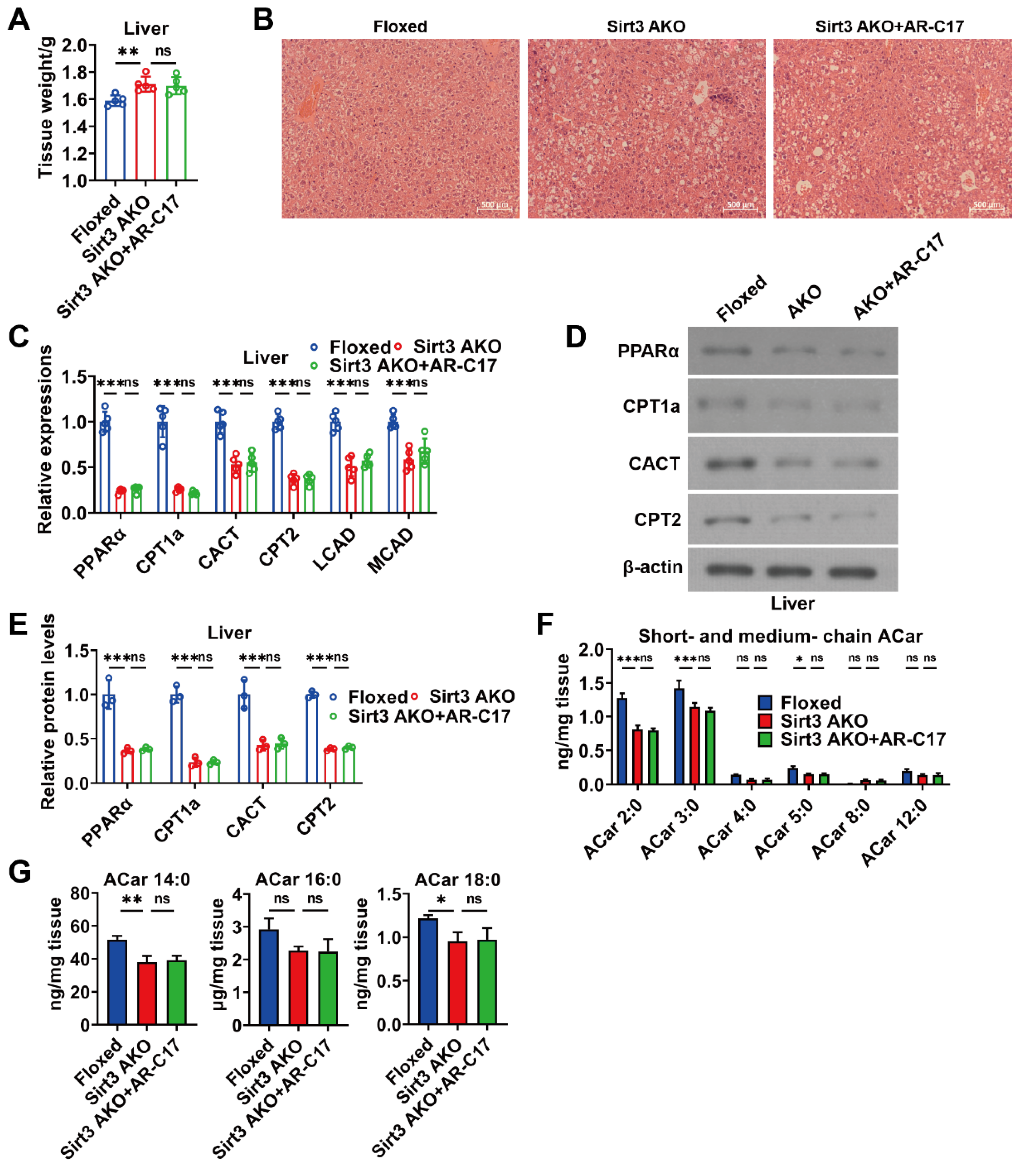
3.5. AR-C17 Failed to Improve Hepatic Fatty Acid Oxidation during Aging in the Absence of Adipose Sirt3
4. Discussion
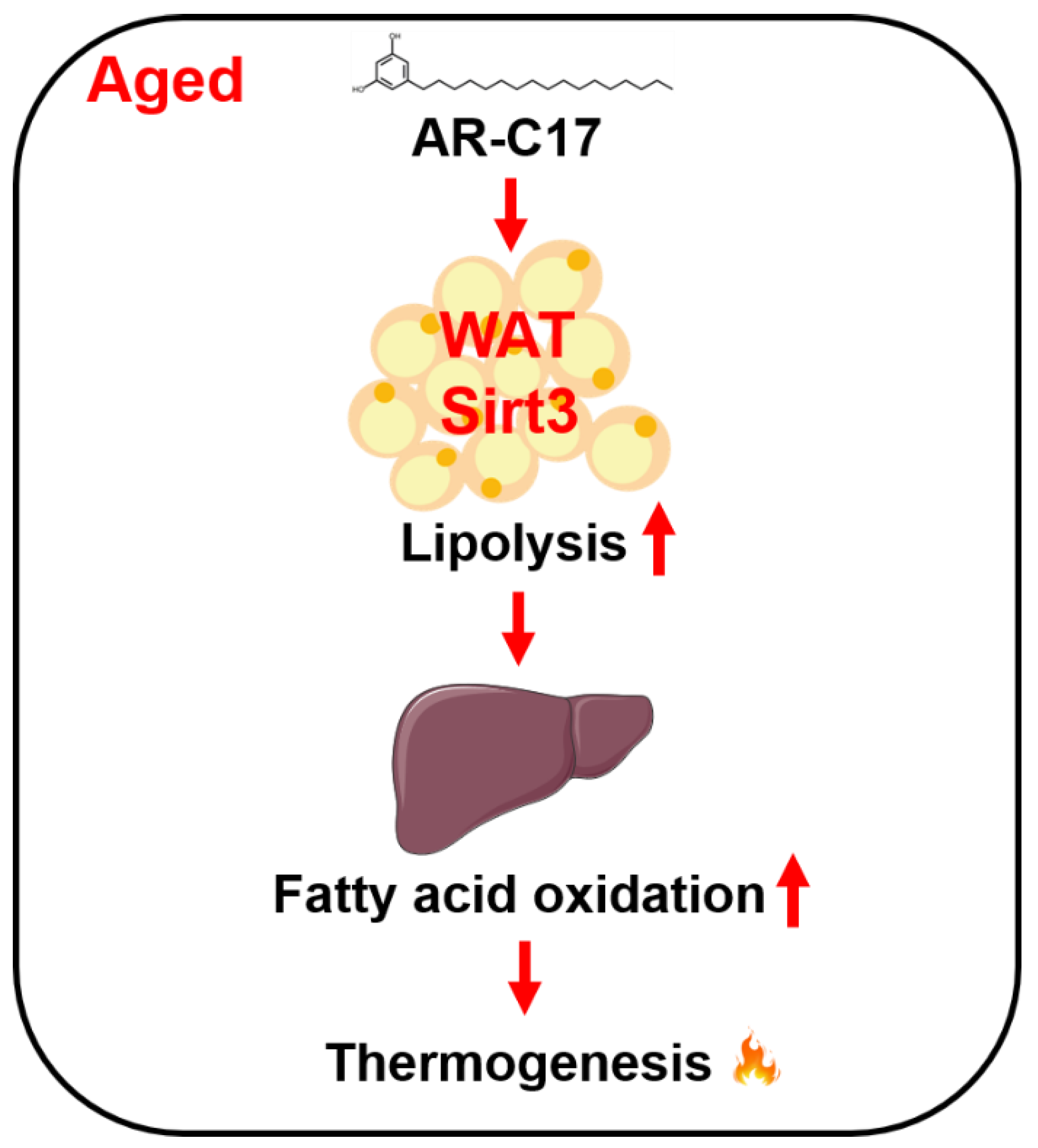
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nikopoulou, C.; Kleinenkuhnen, N.; Parekh, S.; Sandoval, T.; Ziegenhain, C.; Schneider, F.; Giavalisco, P.; Donahue, K.F.; Vesting, A.J.; Kirchner, M.; et al. Spatial and single-cell profiling of the metabolome, transcriptome and epigenome of the aging mouse liver. Nat. Aging 2023, 3, 1430–1445. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Rhyu, J.; Yu, R. Newly discovered endocrine functions of the liver. World J. Hepatol. 2021, 13, 1611–1628. [Google Scholar] [CrossRef]
- Hunt, N.J.; Kang, S.W.S.; Lockwood, G.P.; Le Couteur, D.G.; Cogger, V.C. Hallmarks of Aging in the Liver. Comput. Struct. Biotechnol. J. 2019, 17, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, C.; Jiang, M.; Liu, X.; Geng, L.; Zhang, Y.; Sun, S.; Wang, K.; Yin, J.; Ma, S.; et al. A single-nucleus transcriptomic atlas of primate liver aging uncovers the pro-senescence role of SREBP2 in hepatocytes. Protein Cell 2024, 15, 98–120. [Google Scholar] [CrossRef]
- Gregg, S.Q.; Gutierrez, V.; Robinson, A.R.; Woodell, T.; Nakao, A.; Ross, M.A.; Michalopoulos, G.K.; Rigatti, L.; Rothermel, C.E.; Kamileri, I.; et al. A mouse model of accelerated liver aging caused by a defect in DNA repair. Hepatology 2012, 55, 609–621. [Google Scholar] [CrossRef]
- Abumrad, N.A. The Liver as a Hub in Thermogenesis. Cell Metab. 2017, 26, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Simcox, J.; Geoghegan, G.; Maschek, J.A.; Bensard, C.L.; Pasquali, M.; Miao, R.; Lee, S.; Jiang, L.; Huck, I.; Kershaw, E.E.; et al. Global Analysis of Plasma Lipids Identifies Liver-Derived Acylcarnitines as a Fuel Source for Brown Fat Thermogenesis. Cell Metab. 2017, 26, 509–522. [Google Scholar] [CrossRef]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef]
- Rona, R.R.; Richard, D.G.; Feike, R.v.d.L. Molecular enzymology of carnitine transfer and transport. Biochim. Biophys. Acta 2001, 1546, 21–43. [Google Scholar] [CrossRef]
- Capurso, C. Whole-Grain Intake in the Mediterranean Diet and a Low Protein to Carbohydrates Ratio Can Help to Reduce Mortality from Cardiovascular Disease, Slow Down the Progression of Aging, and to Improve Lifespan: A Review. Nutrients 2021, 13, 2540. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Beck, T.; Dhana, K.; Desai, P.; Krueger, K.R.; Tangney, C.C.; Holland, T.M.; Agarwal, P.; Evans, D.A.; Rajan, K.B. Association of Whole Grain Consumption and Cognitive Decline: An Investigation From a Community-Based Biracial Cohort of Older Adults. Neurology 2023, 101, 2277–2287. [Google Scholar] [CrossRef] [PubMed]
- Sang, S. Biomarkers of Whole Grain Intake. J. Agric. Food Chem. 2018, 66, 10347–10352. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Landberg, R. The Chemistry behind Health Effects of Whole Grains. Mol. Nutr. Food Res. 2017, 61, 1770074. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Yang, Z.; Liu, J.; Wang, Z.; Speakman, J.R.; Niu, C.; Sun, B.; Wang, J. Protective effects of 5-heptadecylresorcinol against adipocyte mitochondrial dysfunction through upregulation of Sirt3-mediated autophagy. J. Nutr. Biochem. 2022, 103, 108956. [Google Scholar] [CrossRef]
- Xie, M.; Liu, J.; Wang, Z.; Sun, B.; Wang, J. Inhibitory effects of 5-heptadecylresorcinol on the proliferation of human MCF-7 breast cancer cells through modulating PI3K/Akt/mTOR pathway. J. Funct. Food 2020, 69, 103946. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Wang, Z.; Hao, Y.; Bai, W.; Wang, Z.; Wang, J. 5-Heptadecylresorcinol, a Biomarker for Whole Grain Rye Consumption, Ameliorates Cognitive Impairments and Neuroinflammation in APP/PS1 Transgenic Mice. Mol. Nutr. Food Res. 2020, 64, 1901218. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Hao, Y.; Wang, Z.; Yang, Z.; Wang, Z.; Wang, J. 5-Heptadecylresorcinol attenuates oxidative damage and mitochondria-mediated apoptosis through activation of the SIRT3/FOXO3a signaling pathway in neurocytes. Food Funct. 2020, 11, 2535–2542. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Yang, H.; Zhang, D.; Zhang, Y.; Wang, J.; Liu, J. 5-Heptadecylresorcinol Ameliorates Obesity-Associated Skeletal Muscle Mitochondrial Dysfunction through SIRT3-Mediated Mitophagy. J. Agric. Food Chem. 2023, 71, 16032–16042. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, Z.; Li, Q.; Wang, Z.; Liu, J.; Wang, J. 5-Heptadecylresorcinol Protects against Atherosclerosis in Apolipoprotein E-Deficient Mice by Modulating SIRT3 Signaling: The Possible Beneficial Effects of Whole Grain Consumption. Mol. Nutr. Food Res. 2022, 66, 2101114. [Google Scholar] [CrossRef]
- Zhang, K.; Li, T.; Li, Q.; Nie, C.; Sun, Y.; Xue, L.; Wang, Y.; Fan, M.; Qian, H.; Li, Y.; et al. 5-Heptadecylresorcinol Regulates the Metabolism of Thermogenic Fat and Improves the Thermogenic Capacity of Aging mice via a Sirtuin 3-Adenosine Monophosphate-Activated Protein Kinase Pathway. J. Agric. Food Chem. 2023, 71, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Jia, X.; Cui, Y.; Song, Y.; Wang, S.; Geng, Y.; Li, R.; Gao, W.; Fu, D. Sirt3-mediated mitophagy regulates AGEs-induced BMSCs senescence and senile osteoporosis. Redox Biol. 2021, 41, 101915. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2021, 22, 119–141. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Pi, C.; Yu, X.; Gao, X.; Zhang, C.; Sun, H.; Zhang, H.; Shi, Y.; He, X. Nicotinamide Mononucleotide Supplementation Improves Mitochondrial Dysfunction and Rescues Cellular Senescence by NAD+/Sirt3 Pathway in Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 14739. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jiang, L.; Xue, L.; Wang, Y.; Sun, Y.; Fan, M.; Qian, H.; Wang, L.; Li, Y. The Enhancement of Acylcarnitine Metabolism by 5-Heptadecylresorcinol in Brown Adipose Tissue Contributes to Improving Glucose and Lipid Levels in Aging Male Mice. Nutrients 2023, 15, 4579. [Google Scholar] [CrossRef] [PubMed]
- Jawhara, M.; Sorensen, S.B.; Heitmann, B.L.; Andersen, V. Biomarkers of Whole-Grain and Cereal-Fiber Intake in Human Studies: A Systematic Review of the Available Evidence and Perspectives. Nutrients 2019, 11, 2994. [Google Scholar] [CrossRef] [PubMed]
- Nemkov, T.; Cendali, F.; Stefanoni, D.; Martinez, J.L.; Hansen, K.C.; San-Millan, I.; D’Alessandro, A. Metabolic Signatures of Performance in Elite World Tour Professional Male Cyclists. Sports Med. 2023, 53, 1651–1665. [Google Scholar] [CrossRef] [PubMed]
- Ruankham, W.; Suwanjang, W.; Wongchitrat, P.; Prachayasittikul, V.; Prachayasittikul, S.; Phopin, K. Sesamin and sesamol attenuate H2O2-induced oxidative stress on human neuronal cells via the SIRT1-SIRT3-FOXO3a signaling pathway. Nutr. Neurosci. 2021, 24, 90–101. [Google Scholar] [CrossRef]
- Bernal-Ramirez, J.; Silva-Platas, C.; Jerjes-Sanchez, C.; Ramos-Gonzalez, M.R.; Vazquez-Garza, E.; Chapoy-Villanueva, H.; Ramirez-Rivera, A.; Zarain-Herzberg, A.; Garcia, N.; Garcia-Rivas, G. Resveratrol Prevents Right Ventricle Dysfunction, Calcium Mishandling, and Energetic Failure via SIRT3 Stimulation in Pulmonary Arterial Hypertension. Oxid. Med. Cell Longev. 2021, 2021, 9912434. [Google Scholar] [CrossRef]
- Lu, H.; Fang, L.; Wang, J.; Zhao, F.; Liu, C.; Gao, Y.; Liu, J.; Min, W. Pine nut antioxidant peptides ameliorate the memory impairment in a scopolamine-induced mouse model via SIRT3-induced synaptic plasticity. Food Funct. 2021, 12, 8026–8036. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, X.; Tang, N.; Tao, H.; Xia, R.; Cheng, Y. Melanoidins, extracted from Chinese traditional vinegar powder, inhibit alcohol-induced inflammation and oxidative stress in macrophages via activation of SIRT1 and SIRT3. Food Funct. 2021, 12, 8120–8129. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zeng, X.; Li, B.; Wang, C.; Zhou, M.; Lang, H.; Yi, L.; Mi, M. Dihydromyricetin attenuates palmitic acid-induced oxidative stress by promoting autophagy via SIRT3-ATG4B signaling in hepatocytes. Nutr. Metab. 2021, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; Wang, B.; Fan, M.; Wang, Y.; Sun, Y.; Qian, H.; Li, Y.; Wang, L. Highland Barley Tea Polyphenols Extract Alleviates Skeletal Muscle Fibrosis in Mice by Reducing Oxidative Stress, Inflammation, and Cell Senescence. J. Agric. Food Chem. 2023, 71, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; Li, Y.; Guan, Y.; Zhang, K.; Liu, J.; Fan, M.; Qian, H.; Wang, L. Highland barley tea represses palmitic acid-induced apoptosis and mitochondrial dysfunction via regulating AMPK/SIRT3/FoxO3a in myocytes. Food Biosci. 2021, 40, 100893. [Google Scholar] [CrossRef]
- Villarroya, F.; Cereijo, R.; Villarroya, J.; Giralt, M. Brown adipose tissue as a secretory organ. Nat. Rev. Endocrinol. 2017, 13, 26–35. [Google Scholar] [CrossRef]
- Wang, G.X.; Zhao, X.Y.; Meng, Z.X.; Kern, M.; Dietrich, A.; Chen, Z.; Cozacov, Z.; Zhou, D.; Okunade, A.L.; Su, X.; et al. The brown fat-enriched secreted factor Nrg4 preserves metabolic homeostasis through attenuation of hepatic lipogenesis. Nat. Med. 2014, 20, 1436–1443. [Google Scholar] [CrossRef]
- Sponton, C.H.; Hosono, T.; Taura, J.; Jedrychowski, M.P.; Yoneshiro, T.; Wang, Q.; Takahashi, M.; Matsui, Y.; Ikeda, K.; Oguri, Y.; et al. The regulation of glucose and lipid homeostasis via PLTP as a mediator of BAT-liver communication. EMBO Rep. 2020, 21, 49828. [Google Scholar] [CrossRef]
- Mills, E.L.; Harmon, C.; Jedrychowski, M.P.; Xiao, H.; Garrity, R.; Tran, N.V.; Bradshaw, G.A.; Fu, A.; Szpyt, J.; Reddy, A.; et al. UCP1 governs liver extracellular succinate and inflammatory pathogenesis. Nat. Metab. 2021, 3, 604–617. [Google Scholar] [CrossRef]
- Sugimoto, S.; Mena, H.A.; Sansbury, B.E.; Kobayashi, S.; Tsuji, T.; Wang, C.H.; Yin, X.; Huang, T.L.; Kusuyama, J.; Kodani, S.D.; et al. Brown adipose tissue-derived MaR2 contributes to cold-induced resolution of inflammation. Nat. Metab. 2022, 4, 775–790. [Google Scholar] [CrossRef]
- Xu, J.; Cui, L.; Wang, J.; Zheng, S.; Zhang, H.; Ke, S.; Cao, X.; Shi, Y.; Li, J.; Zen, K.; et al. Cold-activated brown fat-derived extracellular vesicle-miR-378a-3p stimulates hepatic gluconeogenesis in male mice. Nat. Commun. 2023, 14, 5480. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Jiang, L.; Xue, L.; Wang, Y.; Sun, Y.; Fan, M.; Qian, H.; Wang, L.; Li, Y. 5-Heptadecylresorcinol Improves Aging-Associated Hepatic Fatty Acid Oxidation Dysfunction via Regulating Adipose Sirtuin 3. Nutrients 2024, 16, 978. https://doi.org/10.3390/nu16070978
Zhang K, Jiang L, Xue L, Wang Y, Sun Y, Fan M, Qian H, Wang L, Li Y. 5-Heptadecylresorcinol Improves Aging-Associated Hepatic Fatty Acid Oxidation Dysfunction via Regulating Adipose Sirtuin 3. Nutrients. 2024; 16(7):978. https://doi.org/10.3390/nu16070978
Chicago/Turabian StyleZhang, Kuiliang, Lei Jiang, Lamei Xue, Yu Wang, Yujie Sun, Mingcong Fan, Haifeng Qian, Li Wang, and Yan Li. 2024. "5-Heptadecylresorcinol Improves Aging-Associated Hepatic Fatty Acid Oxidation Dysfunction via Regulating Adipose Sirtuin 3" Nutrients 16, no. 7: 978. https://doi.org/10.3390/nu16070978
APA StyleZhang, K., Jiang, L., Xue, L., Wang, Y., Sun, Y., Fan, M., Qian, H., Wang, L., & Li, Y. (2024). 5-Heptadecylresorcinol Improves Aging-Associated Hepatic Fatty Acid Oxidation Dysfunction via Regulating Adipose Sirtuin 3. Nutrients, 16(7), 978. https://doi.org/10.3390/nu16070978