
Research Progress of Plant-Derived Natural Products against Drug-Resistant Cancer


Abstract
1. Introduction
2. Method
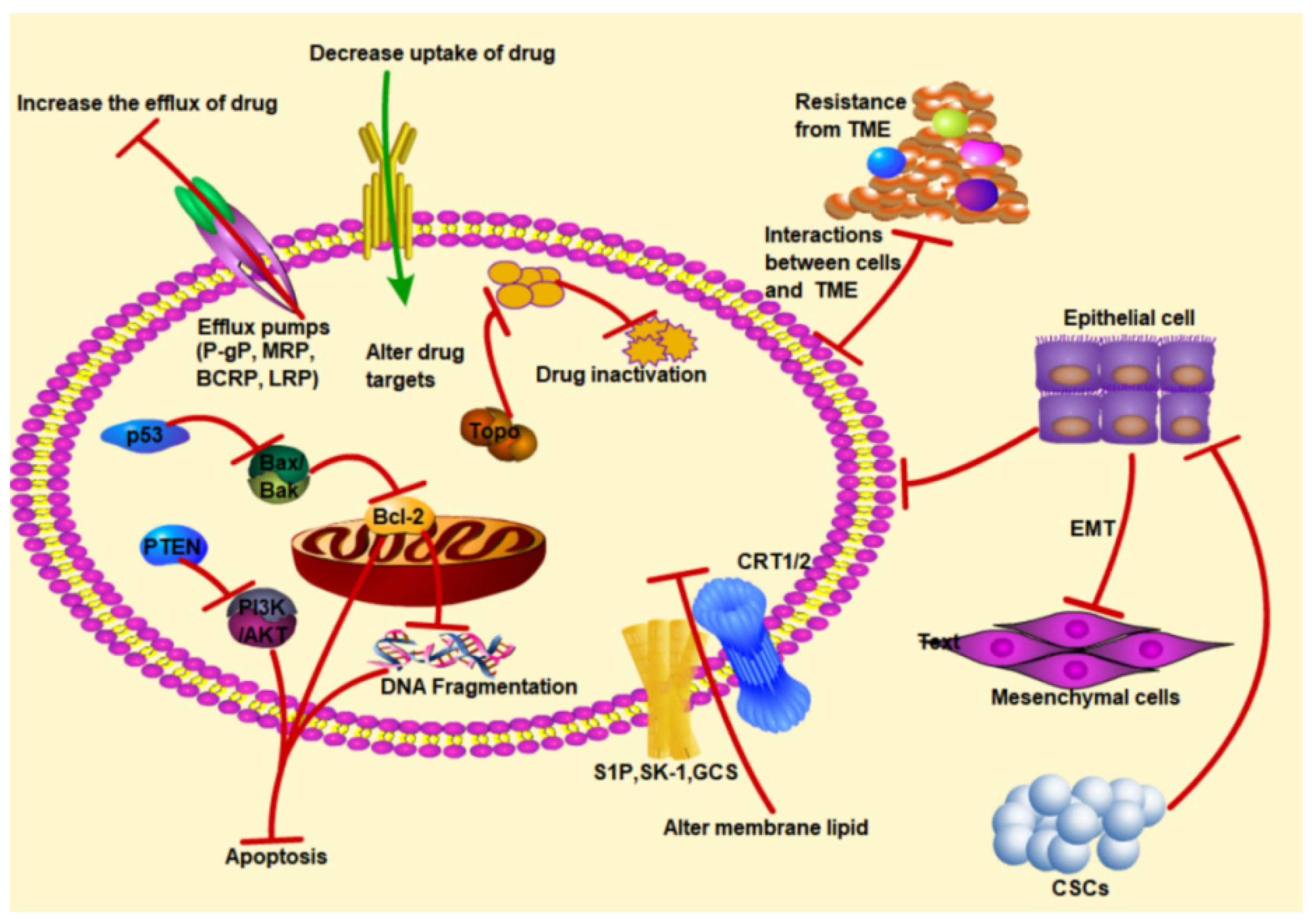
3. Molecular Mechanisms of Drug Resistance in Cancer
3.1. Drug Accumulation and Absorption Reduction
3.2. Inactivation of Drug or Alteration of Drug Target
3.3. Inhibition of Apoptosis-Related Gene Expression
3.4. Alteration in Membrane Lipid
3.5. EMT
3.6. Cancer Stem Cells
3.7. The Tumor Microenvironment
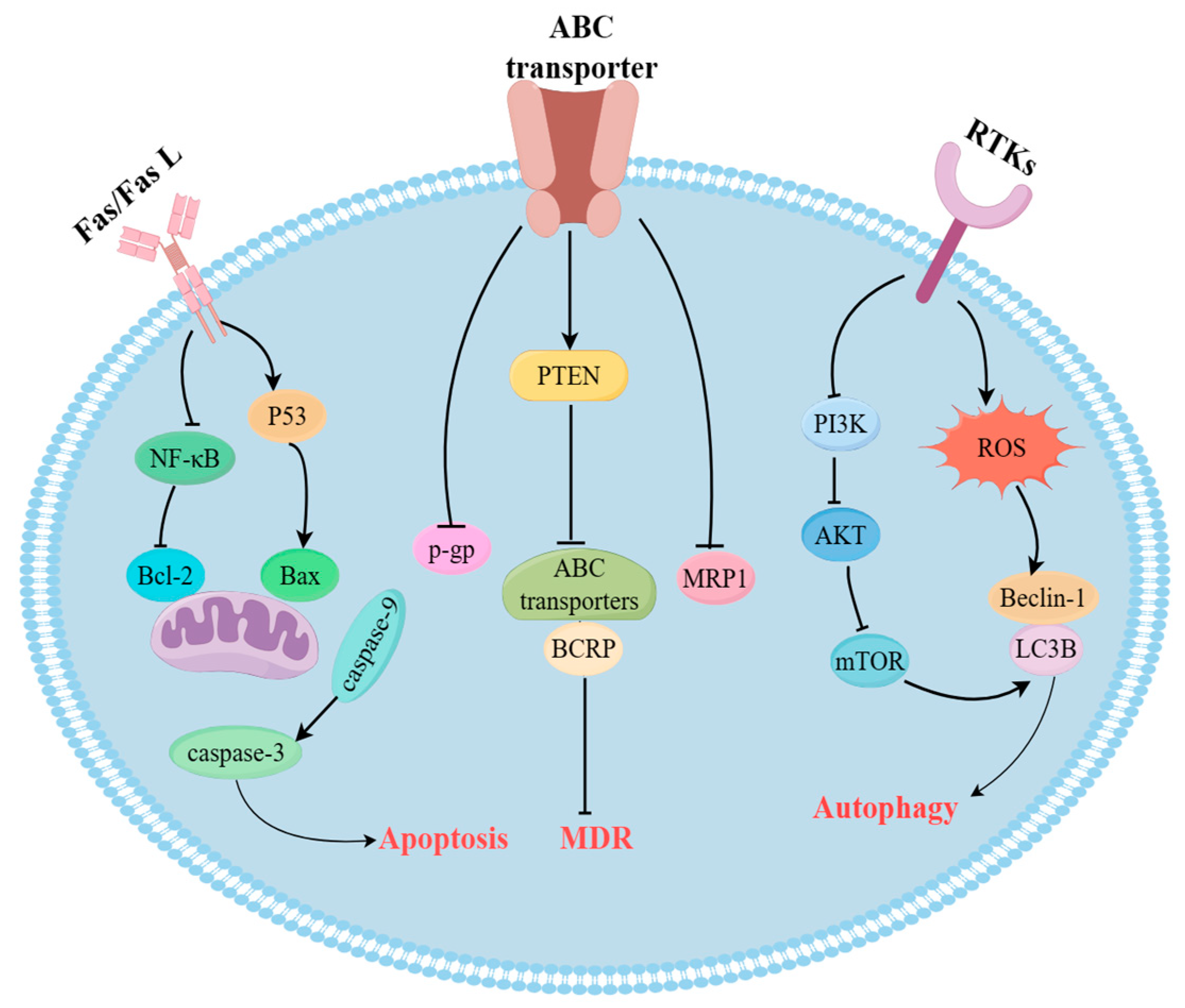
4. Potential Mechanisms of Plant-Derived Natural Products in the Treatment of Drug-Resistant Cancer
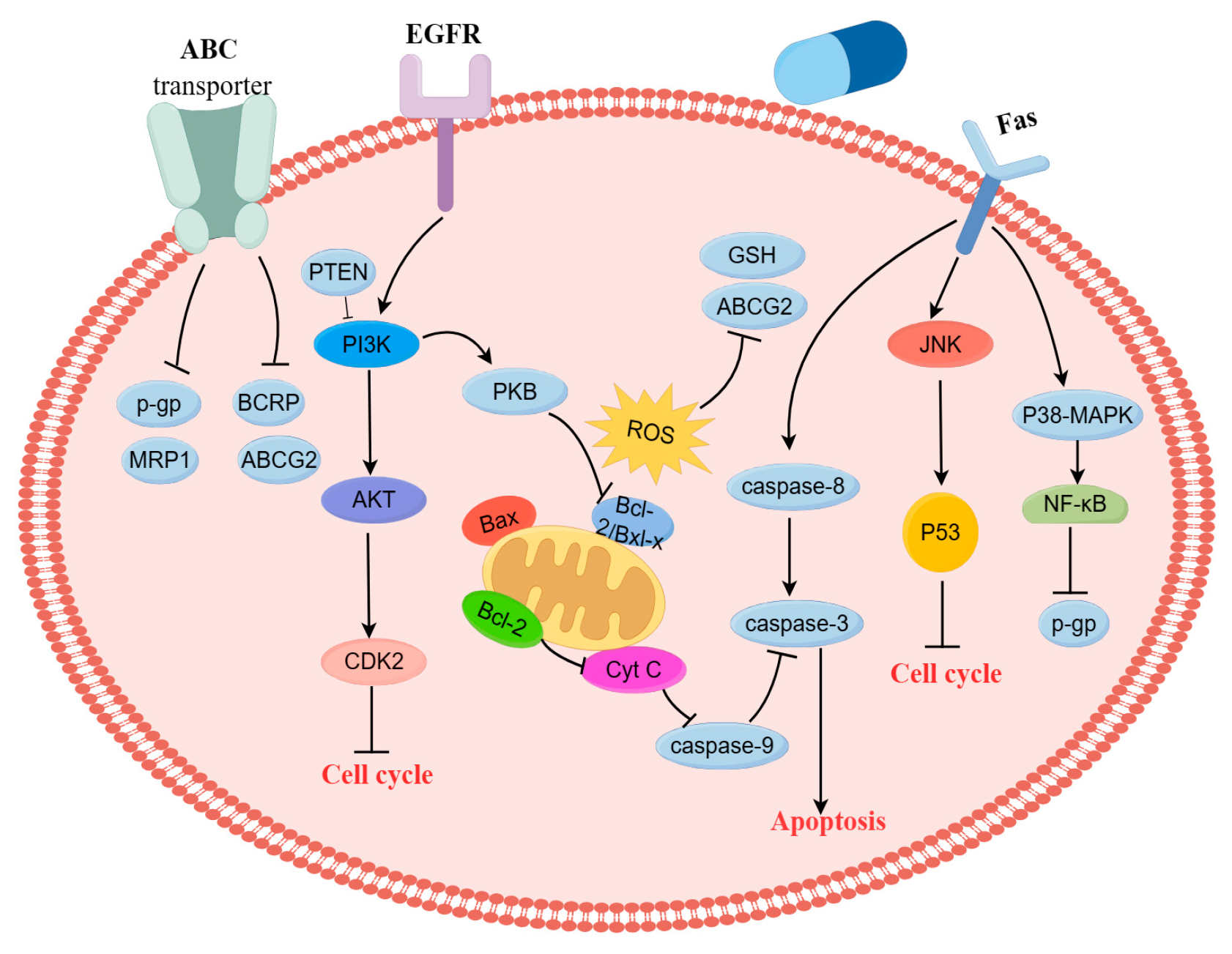
4.1. Regulation of MDR Gene-Related Proteins
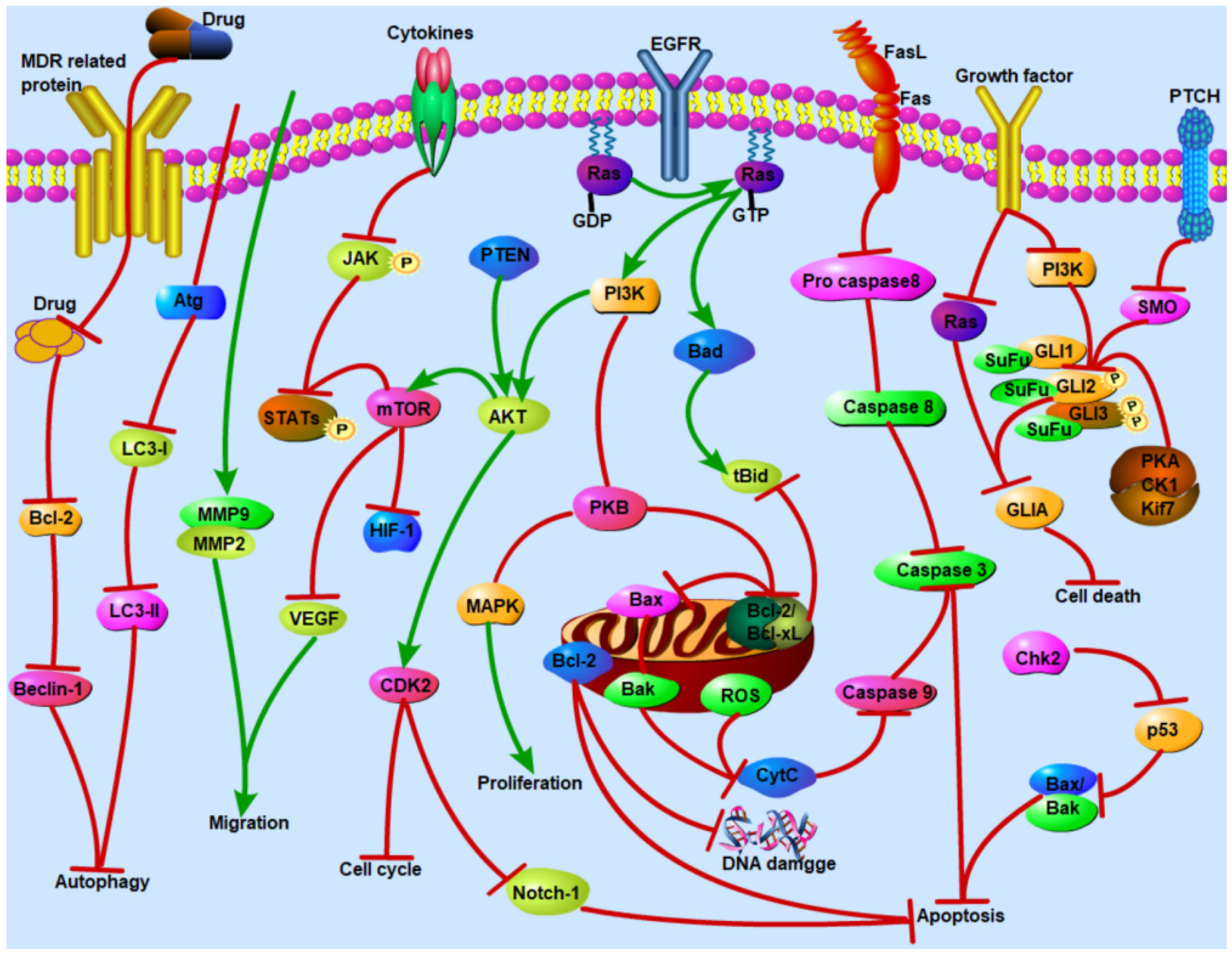
4.2. Induction of Apoptosis through PI3K/AKT Signaling Pathway
4.3. Regulation of Autophagy Pathway
4.4. Regulation of NF-κB/MAPK Signaling Pathway
4.5. Blocking Cancer Cells Cycle
4.6. Inhibiting Growth of Cancer Stem Cells
4.7. Other Signaling Pathways
5. Synergistic Effects and Combination Therapies
6. Clinical and Preclinical Research
7. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F.; Bsc, M.F.B.; Me, J.F.; Soerjomataram, M.I.; et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Germani, M.M.; Borelli, B.; Boraschi, P.; Antoniotti, C.; Ugolini, C.; Urbani, L.; Morelli, L.; Fontanini, G.; Masi, G.; Cremolini, C.; et al. The management of colorectal liver metastases amenable of surgical resection: How to shape treatment strategies according to clinical, radiological, pathological and molecular features. Cancer Treat. Rev. 2022, 106, 102382. [Google Scholar] [CrossRef] [PubMed]
- Witek, M.E.; Kimple, R.J.; Avey, G.D.; Burr, A.R.; Chandereng, T.; Yu, M.; Hu, R.; Wieland, A.M.; Labby, Z.E.; Bruce, J.Y.; et al. Prospective Study of PET/MRI Tumor Response During Chemoradiotherapy for Patients with Low-risk and Intermediate-risk p16-positive Oropharynx Cancer. Am. J. Clin. Oncol. 2022, 45, 202–207. [Google Scholar] [CrossRef]
- Zeng, Y.; Xiang, Y.; Sheng, R.; Tomás, H.; Rodrigues, J.; Gu, Z.; Zhang, H.; Gong, Q.; Luo, K. Polysaccharide-based nanomedicines for cancer immuno-therapy: A review. Bioact. Mater. 2021, 6, 3358–3382. [Google Scholar]
- Tang, Z.H.; Chen, J.L.; Liu, S.Y.; Yu, X.P.; Tong, H.J.; Quan, Z.W. Significance of combination of targeted therapy and im-munotherapy in conversion therapy of biliary tract cancer. Zhonghua Wai Ke Za Zhi 2022, 60, 343–350. [Google Scholar] [PubMed]
- Kardashian, A.; Florman, S.S.; Haydel, B.; Ruiz, R.M.; Klintmalm, G.B.; Lee, D.D.; Burcin Taner, C.; Aucejo, F.; Tevar, A.D.; Humar, A.; et al. Liver Transplantation Outcomes in a U.S. Multicenter Cohort of 789 Patients with Hepatocellular Carcinoma Presenting Beyond Milan Criteri. Hepatology 2020, 72, 2014–2028. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Wu, L.-L.; Lau, W.-Y.; Huan, H.-B.; Wen, X.-D.; Ma, K.-S.; Li, X.-W.; Bie, P. Adjuvant sorafenib after heptectomy for Barcelona Clinic Liver Cancer-stage C hepatocellular carcinoma patients. World J. Gastroenterol. 2016, 22, 5384–5392. [Google Scholar] [CrossRef]
- Chuma, M.; Terashita, K.; Sakamoto, N. New molecularly targeted therapies against advanced hepatocellular carcinoma: From molecular pathogenesis to clinical trials and future directions. Hepatol. Res. 2015, 45, E1–E11. [Google Scholar] [CrossRef]
- Ling, S.; Li, J.; Shan, Q.; Dai, H.; Lu, D.; Wen, X.; Song, P.; Xie, H.; Zhou, L.; Liu, J.; et al. USP22 mediates the multidrug resistance of hepatocellular carcinoma via the SIRT1/AKT/MRP1 signaling pathway. Mol. Oncol. 2017, 11, 682–695. [Google Scholar] [CrossRef]
- Schoenfeld, A.J.; Hellmann, M.D. Acquired Resistance to Immune Checkpoint Inhibitors. Cancer Cell 2020, 37, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Catalano, A.; Iacopetta, D.; Ceramella, J.; Scumaci, D.; Giuzio, F.; Saturnino, C.; Aquaro, S.; Rosano, C.; Sinicropi, M.S. Multidrug Resistance (MDR): A Widespread Phenomenon in Pharmacological Therapies. Molecules 2022, 27, 616. [Google Scholar] [CrossRef]
- Ramirez, M.; Rajaram, S.; Steininger, R.J.; Osipchuk, D.; Roth, M.A.; Morinishi, L.S.; Evans, L.; Ji, W.; Hsu, C.-H.; Thurley, K.; et al. Diverse drug-resistance mechanisms can emerge from drug-tolerant cancer persister cells. Nat. Commun. 2016, 7, 10690. [Google Scholar] [CrossRef]
- Chen, X.; Lu, P.; Wang, D.-D.; Yang, S.-J.; Wu, Y.; Shen, H.-Y.; Zhong, S.-L.; Zhao, J.-H.; Tang, J.-H. The role of miRNAs in drug resistance and prognosis of breast cancer formalin-fixed paraffin-embedded tissues. Gene 2016, 595, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Sun, W.; Liu, R.; Zhou, Z.; Zhang, H.; Chen, X.; Ba, Y. Plasma Exosomal miRNA Expression Profile as Oxaliplatin-Based Chemoresistant Biomarkers in Colorectal Adenocarcinoma. Front. Oncol. 2020, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, L.; Li, J.; Du, Y.; Wang, J.; Liu, J. Effects of long noncoding RNA (linc-VLDLR) existing in extracellular vesicles on the occurrence and multidrug resistance of esophageal cancer cells. Pathol. Res. Pract. 2018, 215, 470–477. [Google Scholar] [CrossRef]
- Jin, K.-T.; Lu, Z.-B.; Lv, J.-Q.; Zhang, J.-G. The Role of Long Non-Coding RNAs in Mediating Chemoresistance by Modulating Au-tophagy in Cancer. RNA Biol. 2020, 17, 1727–1740. [Google Scholar] [CrossRef]
- Cheung-Ong, K.; Giaever, G.; Nislow, C. DNA-Damaging Agents in Cancer Chemotherapy: Serendipity and Chemical Biology. Chem. Biol. 2013, 20, 648–659. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-D.; Sun, Y.; Zhou, W.-J.; Xie, X.-Z.; Zhou, Q.-M.; Lu, Y.-Y.; Su, S.-B. Resveratrol Enhances Inhibition Effects of Cisplatin on Cell Migration and Invasion and Tumor Growth in Breast Cancer MDA-MB-231 Cell Models In Vivo and In Vitro. Molecules 2021, 26, 2204. [Google Scholar] [CrossRef]
- Wei, T.; Xiaojun, X.; Peilong, C. Magnoflorine improves sensitivity to doxorubicin (DOX) of breast cancer cells via inducing apoptosis and autophagy through AKT/mTOR and p38 signaling pathways. Biomed. Pharmacother. 2020, 121, 109139. [Google Scholar] [CrossRef]
- Xin, M.; Wang, Y.; Ren, Q.; Guo, Y. Formononetin and metformin act synergistically to inhibit growth of MCF-7 breast cancer cells in vitro. Biomed. Pharmacother. 2019, 109, 2084–2089. [Google Scholar] [CrossRef]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade Drug Resist. Updates 2015, 18, 1–17. [Google Scholar] [CrossRef]
- Kang, Y.; Hu, W.; Bai, E.; Zheng, H.; Liu, Z.; Wu, J.; Jin, R.; Zhao, C.; Liang, G. Curcumin sensitizes human gastric cancer cells to 5-fluorouracil through inhibition of the NFκB survival-signaling pathway. OncoTargets Ther. 2016, 9, 7373–7384. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wu, J.; Dai, N.; Yu, H.; Si, J. Curcumin reverses chemoresistance of human gastric cancer cells by downregulating the NF-κB transcription factor. Oncol. Rep. 2011, 26, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Tan, J.; Wink, M.; Ma, Y.; Li, N.; Su, G. An isoquinoline alkaloid from the Chinese herbal plant Corydalis yanhusuo W.T. Wang inhibits P-glycoprotein and multidrug resistance-associate protein 1. Food Chem. 2013, 136, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wei, P.K.; Qin, Z.F.; Xu, L.; Zhan, L.Z.; He, J.; Li, Y.L. Experimental study on reversion effect of elemene on multidrug resistance of human gastric cancer cell line. J. Chengdu Univ. Tradit. Chin. Med. 2005, 28, 51–53. [Google Scholar]
- Song, H. Mechanism of Reversal of Invasion and Metastasis of Multidrug Resistant Gastric Cancer Cells by β-Elemene. Master’s Thesis, China Pharmaceutical University, Shenyang, China, 2019. [Google Scholar]
- Turner, N.C.; Reis-Filho, J.S. Genetic heterogeneity and cancer drug resistance. Lancet Oncol. 2012, 13, e178–e185. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Li, Z.; Gao, C.-Y.; Cho, C.H. Mechanisms of drug resistance in colon cancer and its therapeutic strategies. World J. Gastroenterol. 2016, 22, 6876–6889. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhou, L. Progress in the study of the mechanism of action of camptothecins and their drug resistance. Sichuan J. Physiol. Sci. 2007, 1, 31–33. [Google Scholar]
- Kikuchi, H.; Yuan, B.; Hu, X.; Okazaki, M. Chemopreventive and anticancer activity of flavonoids and its possibility for clinical use by combining with conventional chemotherapeutic agents. Am. J. Cancer Res. 2019, 9, 1517–1535. [Google Scholar]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef]
- Marchi, E.; O’Connor, O.A. Safety and efficacy of pralatrexate in the treatment of patients with relapsed or refractory peripheral T-cell lymphoma. Ther. Adv. Hematol. 2012, 3, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Li, L.; Ren, Y.; Xue, H.; Liu, J.; Wen, S.; Chen, J. Synthesis of N-Carbonyl Acridanes as Highly Potent Inhibitors of Tubulin Polymerization via One-Pot Copper-Catalyzed Dual Arylation of Nitriles with Cyclic Diphenyl Iodoniums. Adv. Synth. Catal. 2020, 362, 2030–2038. [Google Scholar] [CrossRef]
- Le, T.; Bhushan, V.; Sochat, M.; Chavda, Y. First Aid for the USMLE Step 1, 1st ed.; McGraw-Hill Education: New York, NY, USA, 2017; pp. 416–419. [Google Scholar]
- Nussbaumer, S.; Bonnabry, P.; Veuthey, J.-L.; Fleury-Souverain, S. Analysis of anticancer drugs: A review. Talanta 2011, 85, 2265–2289. [Google Scholar] [CrossRef] [PubMed]
- Rabik, C.A.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat. Rev. 2007, 33, 9–23. [Google Scholar] [CrossRef]
- Stordal, B.; Davey, M. Understanding cisplatin resistance using cellular models. IUBMB Life 2007, 59, 696–699. [Google Scholar] [CrossRef]
- Rabik, C.A.; Maryon, E.B.; Kasza, K.; Shafer, J.T.; Bartnik, C.M.; Dolan, M.E. Role of copper transporters in resistance to platinating agents. Cancer Chemother Pharmacol. 2009, 64, 133–142. [Google Scholar] [CrossRef]
- Zhu, Z.; Du, S.; Du, Y.; Ren, J.; Ying, G.; Yan, Z. Glutathione reductase mediates drug resistance in glioblastoma cells by regulating redox homeostasis. J. Neurochem. 2018, 144, 93–104. [Google Scholar] [CrossRef]
- Zamble, D.B.; Mu, D.; Reardon, J.T.; Sancar, A.; Lippard, S.J. Repair of Cisplatin−DNA Adducts by the Mammalian Excision Nuclease. Biochemistry 1996, 35, 10004–10013. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef]
- Dean, M.; Moitra, K.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Hum. Mutat. 2022, 43, 1162–1182. [Google Scholar] [CrossRef]
- Abdalla, A.N.; Di Stefano, M.; Poli, G.; Tuccinardi, T.; Bader, A.; Vassallo, A.; Abdallah, M.E.; El-Readi, M.Z.; Refaat, B.; Algarni, A.S.; et al. Co-Inhibition of P-gP and Hsp90 by an Isa-tin-Derived Compound Contributes to the Increase of the Chemosensitivity of MCF7/ADR-Resistant Cells to Doxorubicin. Molecules 2021, 27, 90. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, H.; Ashby, C.R.A., Jr.; Assaraf, Y.G.; Chen, Z.S.; Liu, H.M. Chemical molecular-based approach to overcome multidrug resistance in cancer by targeting P-glycoprotein (P-gp). Med. Res. Rev. 2021, 41, 525–555. [Google Scholar] [CrossRef] [PubMed]
- Elmeliegy, M.; Vourvahis, M.; Guo, C.; Wang, D.D. Effect of P-glycoprotein (P-gp) Inducers on Exposure of P-gp Substrates: Review of Clinical Drug–Drug Interaction Studies. Clin. Pharmacokinet. 2020, 59, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.X.; Sun, Y.B.; Wang, S.Q.; Duan, L.; Huo, Q.L.; Ren, F.; Li, G.F. Grape Seed Procyanidin Reversal of P-glycoprotein Associated Multi-Drug Resistance via Down-regulation of NF-κB and MAPK/ERK Mediated YB-1 Activity in A2780/T Cells. PLoS ONE 2013, 8, e71071. [Google Scholar] [CrossRef]
- Riganti, C.; Giampietro, R.; Kopecka, J.; Costamagna, C.; Abatematteo, F.S.; Contino, M.; Abate, C. MRP1-Collateral Sensitizers as a Novel Therapeutic Approach in Resistant Cancer Therapy: An In Vitro and In Vivo Study in Lung Resistant Tumor. Int. J. Mol. Sci. 2020, 21, 3333. [Google Scholar] [CrossRef]
- Mahdizadeh, S.; Karimi, G.; Behravan, J.; Arabzadeh, S.; Lage, H.; Kalalinia, F. Crocin suppresses multidrug resistance in MRP overexpressing ovarian cancer cell line. DARU J. Pharm. Sci. 2016, 24, 17. [Google Scholar] [CrossRef]
- Jackson, S.M.; Manolaridis, I.; Kowal, J.; Zechner, M.; Taylor, N.M.I.; Bause, M.; Bauer, S.; Bartholomaeus, R.; Bernhardt, G.; Koenig, B.; et al. Structural basis of small-molecule inhibition of human multidrug transporter ABCG2. Nat. Struct. Mol. Biol. 2018, 25, 333–340. [Google Scholar] [CrossRef]
- Carrillo, I.O.; Peñafiel, C.R.; Peralta, E.M.; Fuller, E.R.; Ipiña, J.J.K.; Cruz, F.C.; Guerrero, E.G.; Jaloma, J.C.; Vargas, K.N.; Tovar, A.M. Clinical significance of the ABCB1 and ABCG2 gene expression levels in acute lymphoblastic leukemia. Hematology 2016, 22, 286–291. [Google Scholar] [CrossRef]
- Zhang, K.G.; Qin, C.Y.; Wang, H.Q.; Wang, J.X.; Wang, Q.M. The Effect of TRAIL on the Expression of Multidrug Resistant Genes MDR1, LRP and GST-? in Drug-Resistant Gastric Cancer Cell SGC7901/VCR. Hepato-Gastroenterology 2012, 59, 2672–2676. [Google Scholar] [CrossRef]
- Kitazono, M.; Okumura, H.; Ikeda, R.; Sumizawa, T.; Furukawa, T.; Nagayama, S.; Seto, K.; Aikou, T.; Akiyama, S.-I. Reversal of LRP-associated drug resistance in colon carcinoma sw-620 cells. Int. J. Cancer 2000, 91, 126–131. [Google Scholar] [CrossRef]
- Haider, T.; Pandey, V.; Banjare, N.; Gupta, P.N.; Soni, V. Drug resistance in cancer: Mechanisms and tackling strategies. Pharmacol. Rep. 2020, 72, 1125–1151. [Google Scholar] [CrossRef]
- Martinho, N.; Santos, T.C.B.; Florindo, H.F.; Silva, L.C. Cisplatin-Membrane Interactions and Their Influence on Platinum Complexes Activity and Toxicity. Front. Physiol. 2019, 9, 1898. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.K.; Bortner, C.D.; Mason, R.P.; Cannon, R.E. Nitric oxide reverses drug resistance by inhibiting ATPase activity of p-glycoprotein in human multi-drug resistant cancer cells. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2806–2814. [Google Scholar] [CrossRef]
- Cai, P.-Y.; Ma, M.-L.; Zhang, Y.-F.; Zhou, Z.-X.; Wang, Y.; He, L.-P.; Wang, W. Inhibition of glutathione metabolism can limit the development of pancreatic cancer. World J. Stem Cells 2022, 14, 362–364. [Google Scholar] [CrossRef]
- Hatem, E.; El Banna, N.; Huang, M.-E. Multifaceted Roles of Glutathione and Glutathione-Based Systems in Carcinogenesis and Anticancer Drug Resistance. Antioxidants Redox Signal. 2017, 27, 1217–1234. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, C.; Lu, X.; Yan, Z.; Gao, J.; Zhao, H.; Li, S. Coniferyl ferulate, a strong inhibitor of glutathione S-transferase isolated from radix Angelicae sinensis, reverses multidrug resistance and downregulates P-glycoprotein, Evid. Based Complement. Alternat. Med. 2013, 34, 21–22. [Google Scholar]
- Pérès, B.; Nasr, R.; Zarioh, M.; Lecerf-Schmidt, F.; Pietro, A.D.; Baubichon-Cortay, H.; Boumendjiel, A. Ferrocene-embedded flavonoids targeting the Achilles heel of multidrug-resistant cancer cells through collateral sensitivity. Eur. J. Med. Chem. 2017, 130, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Tew, K.D. Glutathione-Associated Enzymes In Anticancer Drug Resistance. Cancer Res 2016, 76, 7–9. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Li, X.; Zhang, Z.; Sun, L.; Wang, Y.; He, Z.; Liu, Z.; Zhu, L.; Fu, L. Kaempferol suppression of acute colitis is regulated by the efflux transporters BCRP and MRP2. Eur. J. Pharm. Sci. 2022, 179, 106303. [Google Scholar] [CrossRef] [PubMed]
- Ganapathi, R.N.; Ganapathi, M.K. Mechanisms regulating resistance to inhibitors of topoisomerase II. Front. Pharmacol. 2013, 4, 89. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhang, G.; Lou, Z.; Xu, G.; Zhang, G. Cryptotanshinone enhances the effect of Arsenic trioxide in treating liver cancer cell by inducing apoptosis through downregulating phosphorylated- STAT3 in vitro and in vivo. BMC Complement. Altern. Med. 2017, 17, 106. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, S.-H.; Shang, X.-J.; Yu, L.-S.; Zhu, J.-W.; Zhao, A.; Zhou, Y.-F.; An, G.-H.; Zhang, Q.; Ma, B. Triptolide induces Sertoli cell apoptosis in mice via ROS/JNK-dependent activation of the mitochondrial pathway and inhibition of Nrf2-mediated antioxidant response. Acta Pharmacol. Sin. 2018, 39, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Daisuke, A.; Hiderou, Y.; Yukihiro, H. Endoplasmic Reticulum (ER) Stress and Endocrine Disorders. Int. J. Mol. Sci. 2017, 18. [Google Scholar]
- Li, Z.; Li, M.; Xia, P.; Wang, L.; Lu, Z. Targeting long non-coding RNA PVT1/TGF-β/Smad by p53 prevents glioma progression. Cancer Biol. Ther. 2022, 23, 225–233. [Google Scholar] [CrossRef]
- Zawacka-Pankau, J.E. The Role of p53 Family in Cancer. Cancers 2022, 14, 823. [Google Scholar] [CrossRef]
- Nishida, Y.; Ishizawa, J.; Ayoub, E.; Patsilevas, T.; Ostermann, L.; Montoya, R.H.; Muftuoglu, M.; Ruvolo, V.; Mak, P.Y.; Tao, W.; et al. Enhanced p53 Activation by Dual Inhibition of MDM2 and XPO1 Disrupts MYC Transcriptional Program and Restores Sensitivity to BCL-2 Inhibition in Ven/HMA Resistant AML. Blood 2021, 138, 505. [Google Scholar] [CrossRef]
- Opferman, J.T.; Kothari, A. Anti-apoptotic BCL-2 family members in development. Cell Death Differ. 2018, 25, 37–45. [Google Scholar] [CrossRef]
- García-Aranda, M.; Pérez-Ruiz, E.; Redondo, M. Bcl-2 Inhibition to Overcome Resistance to Chemo- and Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3950. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhu, D.; Lu, S.; Wang, T.; Wang, J.; Jiang, B.; Shu, Y.; Liu, P. miR-497 modulates multidrug resistance of human cancer cell lines by targeting BCL2. Med. Oncol. 2011, 29, 384–391. [Google Scholar] [CrossRef]
- Tan, W.; Zheng, T.; Wang, A.; Roacho, J.; Thao, S.; Du, P.; Jia, S.; Yu, J.; King, B.L.; Kohli, M. Dynamic changes in gene alterations during chemotherapy in metastatic castrate resistant prostate cancer. Sci. Rep. 2022, 12, 4672. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulos, A.; Varras, M.; Vasilakaki, T.; Varra, V.-K.; Tsavari, A.; Varra, F.-N.; Nonni, A.; Kavantzas, N.; Lazaris, A.C. Expression of p53 and PTEN in human primary endometrial carcinomas: Clinicopathological and immunohistochemical analysis and study of their concomitant expression. Oncol. Lett. 2019, 17, 4575–4589. [Google Scholar] [CrossRef]
- Liu, T.; Guo, J.; Zhang, X. MiR-202-5p/PTEN mediates doxorubicin-resistance of breast cancer cells via PI3K/Akt signaling pathway. Cancer Biol. Ther. 2019, 20, 989–998. [Google Scholar] [CrossRef]
- Ibrahim, H.I.M.; Ismail, M.B.; Ammar, R.B.; Ahmed, E.A. Thidiazuron suppresses breast cancer progression via targeting miR-132 and miR-202-5p/PTEN axis mediated dysregulation of PI3K/AKT signaling pathway. Biochem. Cell Biol. 2020, 99, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, B.; Sun, L.; Yan, Q.; Zhang, Y.; Zhang, Z.; Wang, C. MicroRNA-130b targets PTEN to induce resistance to cisplatin in lung cancer cells by activating Wnt/β-catenin pathway. Cell Biochem. Funct. 2018, 36, 194–202. [Google Scholar] [CrossRef]
- Li, J.; Zhou, W.; Mao, Q.; Gao, D.; Xiong, L.; Hu, X.; Zheng, Y.; Xu, X. HMGB1 Promotes Resistance to Doxorubicin in Human Hepatocellular Carcinoma Cells by Inducing Autophagy via the AMPK/mTOR Signaling Pathway. Front. Oncol. 2021, 11, 739145. [Google Scholar] [CrossRef]
- Singh, K.D.; Rawat, P.; Bonnin, J.-M. Cognitive radio for vehicular ad hoc networks (CR-VANETs): Approaches and challenges. EURASIP J. Wirel. Commun. Netw. 2014, 2014, 49. [Google Scholar] [CrossRef]
- Sun, W.L.; Lan, D.; Gan, T.Q.; Cai, Z.W. Autophagy facilitates multidrug resistance development through inhibition of apoptosis in breast cancer cells. Neoplasma 2015, 62, 199–208. [Google Scholar] [CrossRef]
- Sanchez-Jimenez, F.; Reyes-Palomares, A.; Moya-Garcia, A.A.; Ranea, J.A.; Medina, M.A. Biocomputational Resources Useful for Drug Discovery Against Compartmentalized Targets. Curr. Pharm. Des. 2014, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhu, Z.; Jia, B.; Zhang, W.; Song, J. Design of acid-activated cell-penetrating peptides with nuclear localization capacity for anticancer drug delivery. J. Pept. Sci. 2021, 27, e3354. [Google Scholar] [CrossRef]
- Raghavan, V.; Vijayaraghavalu, S.; Peetla, C.; Yamada, M.; Morisada, M.; Labhasetwar, V. Sustained epigenetic drug delivery de-pletes cholesterol–sphingomyelin rafts from resistant breast cancer cells, influencing biophysical characteristics of membrane lipids. Langmuir 2015, 31, 11564–11573. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Sun, T.; Luan, Y.; Chen, Y.; Song, J.; Ling, L.; Yuan, P.; Li, R.; Cui, K.; Ruan, Y.; et al. Berberine ameliorates erectile dysfunction in rats with streptozotocin-induced diabetes mellitus through the attenuation of apoptosis by inhibiting the SPHK1/S1P/S1PR2 and MAPK pathways. Andrology 2022, 10, 404–418. [Google Scholar] [CrossRef]
- Jung, J.H.; Taniguchi, K.; Lee, H.M.; Lee, M.Y.; Bandu, R.; Komura, K.; Lee, K.Y.; Akao, Y.; Kim, K.P. Comparative lipidomics of 5-Fluorouracil–sensitive and –resistant colorectal cancer cells reveals altered sphingomyelin and ceramide controlled by acid sphingomyelinase (SMPD1). Sci. Rep. 2020, 10, 6124. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Millner, A.; Semina, S.E.; Huggins, R.J.; Running, L.; Aga, D.S.; Tonetti, D.A.; Schiff, R.; Greene, G.L.; Atilla-Gokcumen, G.E.; et al. Endocrine Therapy-Resistant Breast Cancer Cells Are More Sensitive to Ceramide Kinase Inhibition and Elevated Ceramide Levels Than Therapy-Sensitive Breast Cancer Cells. Cancers 2022, 14, 2380. [Google Scholar] [CrossRef]
- Gao, H.; Deng, L. Sphingosine Kinase-1 Activation Causes Acquired Resistance Against Sunitinib in Renal Cell Carcinoma Cells. Cell Biochem. Biophys. 2013, 68, 419–425. [Google Scholar] [CrossRef]
- Madigan, J.P.; Robey, R.W.; Poprawski, J.E.; Huang, H.; Clarke, C.J.; Gottesman, M.M.; Cabot, M.C.; Rosenberg, D.W. A role for ceramide glycosylation in resistance to oxaliplatin in colorectal cancer. Exp. Cell Res. 2020, 388, 111860. [Google Scholar] [CrossRef]
- Zhang, J.J.; Ge, Y.L.; Wang, S.L.; Wang, Z.; Wang, Z.R. Observation on apoptosis of gastric cancer cell line SGC7901 and cispla-tin-resistant gastric cancer cell line SGC7901/DDP transfected with glucosylceramide synthase siRNA. Shandong Med. J. 2020, 60, 6–9. [Google Scholar]
- Kim, E.S.; Tang, X.; Peterson, D.R.; Kilari, D.; Chow, C.-W.; Fujimoto, J.; Kalhor, N.; Swisher, S.G.; Stewart, D.J.; Wistuba, I.I.; et al. Copper transporter CTR1 expression and tissue platinum concentration in non-small cell lung cancer. Lung Cancer 2014, 85, 88–93. [Google Scholar] [CrossRef]
- Sjostedt, N.; Holvikari, K.; Tammela, P.; Kidron, H. Inhibition of breast cancer resistance protein and multidrug resistance asso-ciated protein 2 by natural compounds and their derivatives. Mol. Pharm. 2017, 14, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Arani, H.Z.; Orouei, S.; Fallah, S.; Ghorbani, A.; Khaledabadi, M.; Kakavand, A.; Tavakolpournegari, A.; Saebfar, H.; Heidari, H.; et al. EMT mechanism in breast cancer metastasis and drug resistance: Revisiting molecular interactions and biological functions. Biomed. Pharmacother. 2022, 155, 113774. [Google Scholar] [CrossRef]
- Koker, S.C.; Yalcin, B.; Turacli, I.D. Metformin resistant MDA-MB-468 cells exhibit EMT-like phenotype and increased migration capacity. Mol. Biol. Rep. 2022, 49, 5973–5984. [Google Scholar] [CrossRef] [PubMed]
- Francou, A.; Anderson, K.V. The Epithelial-to-Mesenchymal Transition in Development and Cancer. Annu. Rev. Cancer Biol. 2020, 4, 197–220. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, L.; Wang, X.; Li, Y.; Liu, Z.; Ye, W.; Huang, W.; Lin, G.; Liu, H.; Zhang, J.; et al. Overexpression of Krüppel-Like Factor 4 Suppresses Migration and Invasion of Non-Small Cell Lung Cancer Through c-Jun-NH2-Terminal Kinase/Epithelial-Mesenchymal Transition Signaling Pathway. Front. Pharmacol. 2020, 10, 1512. [Google Scholar] [CrossRef] [PubMed]
- Li, R.M.; Nai, M.M.; Duan, S.J.; Li, S.X.; Yin, B.N.; An, F.; Zhai, Y.Q.; Liu, J.; Chu, Y.R.; Yu, Y.; et al. Down-expression of GOLM1 enhances the chemo-sensitivity of cervical cancer to methotrexate through modulation of the MMP13/EMT axis. Am. J. Cancer Res. 2018, 8, 964–980. [Google Scholar]
- Xiong, Y.; Sun, F.; Dong, P.; Watari, H.; Yue, J.; Yu, M.F.; Lan, C.Y.; Wang, Y.; Ma, Z.B. iASPP induces EMT and cisplatin resistance in human cervical cancer through miR-20a-FBXL5/BTG3 signaling. J. Exp. Clin. Cancer Res. 2017, 36, 48. [Google Scholar] [CrossRef]
- Qin, Q.; Li, X.; Liang, X.; Zeng, L.; Wang, J.; Sun, L.; Zhong, D. Targeting the EMT transcription factor Snail overcomes resistance to osimertinib in EGFR-mutant non-small cell lung cancer. Thorac. Cancer 2021, 12, 1708–1715. [Google Scholar] [CrossRef]
- Zhao, J.; Tan, W.; Zhang, L.; Liu, J.; Shangguan, M.; Chen, J.; Zhao, B.; Peng, Y.; Cui, M.; Zhao, S. FGFR3 phosphorylates EGFR to promote cisplatin-resistance in ovarian cancer. Biochem. Pharmacol. 2021, 190, 114536. [Google Scholar] [CrossRef]
- Raoof, S.; Mulford, I.J.; Frisco-Cabanos, H.; Nangia, V.; Timonina, D.; Labrot, E.; Hafeez, N.; Bilton, S.J.; Drier, Y.; Ji, F.; et al. Targeting FGFR overcomes EMT-mediated resistance in EGFR mutant non-small cell lung cancer. Oncogene 2019, 38, 6399–6413. [Google Scholar] [CrossRef]
- Xu, D.; Liu, S.; Zhang, L.; Song, L. MiR-211 inhibits invasion and epithelial-to-mesenchymal transition (EMT) of cervical cancer cells via targeting MUC4. Biochem. Biophys. Res. Commun. 2017, 485, 556–562. [Google Scholar] [CrossRef]
- Jing, L.; Bo, W.; Yourong, F.; Tian, W.; Shixuan, W.; Mingfu, W. Sema4C mediates EMT inducing chemotherapeutic resistance of miR-31-3p in cervical cancer cells. Sci. Rep. 2019, 9, 17727. [Google Scholar] [CrossRef]
- Zhang, Y.B. Research on the role and mechanism of miR-4725-5p in the regulation of proliferation, migration, and cisplatin resistance in lung cancer by targeting CDH1. Master’s Thesis, Hubei University of Chinese Medicine, Wuhan, China, 2022. [Google Scholar]
- Lei, C.; Wang, Y.; Huang, Y.; Yu, H.; Huang, Y.; Wu, L.; Huang, L. Up-regulated miR155 Reverses the Epithelial-mesenchymal Transition Induced by EGF and Increases Chemo-sensitivity to Cisplatin in Human Caski Cervical Cancer Cells. PLoS ONE 2012, 7, e52310. [Google Scholar] [CrossRef] [PubMed]
- Risom, T.; Langer, E.M.; Chapman, M.P.; Rantala, J.; Fields, A.J.; Boniface, C.; Alvarez, M.J.; Kendsersky, N.D.; Pelz, C.R.; Johnson-Camacho, K.; et al. Differentiation-state plasticity is a targetable resistance mechanism in basal-like breast cancer. Nat. Commun. 2018, 9, 3815. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Diao, L.; Cuentas, E.R.P.; Denning, W.L.; Chen, L.; Fan, Y.H.; Byers, L.A.; Wang, J.; Papadimitrakopoulou, V.A.; Behrens, C.; et al. Epithelial-mesenchymal transition is associated with a distinct tumor microenvironment including elevation of inflammatory signals and multiple immune checkpoints in lung adenocarcinoma. Clin. Cancer Res. 2016, 22, 3630–3642. [Google Scholar] [CrossRef] [PubMed]
- Leon, G.; MacDonagh, L.; Finn, S.P.; Cuffe, S.; Barr, M.P. Cancer stem cells in drug resistant lung cancer: Targeting cell surface markers and signaling pathways. Pharmacol. Ther. 2016, 158, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Rofstad, E.K. Cancer stem cells (CSCs), cervical CSCs and targeted therapies. Oncotarget 2017, 8, 35351–35367. [Google Scholar] [CrossRef] [PubMed]
- Carnero, A.; Garcia-Mayea, Y.; Mir, C.; Lorente, J.; Rubio, I.; Lleonart, M. The cancer stem-cell signaling network and resistance to therapy. Cancer Treat. Rev. 2016, 49, 25–36. [Google Scholar] [CrossRef]
- Tang, Q.; Liu, S.; Shen, H.M. Relationship between cancer stem cells and drug resistant in lung cancer. Bachu Med. J. 2021, 4, 112–116. [Google Scholar]
- Di Fiore, R.; Suleiman, S.; Drago-Ferrante, R.; Subbannayya, Y.; Pentimalli, F.; Giordano, A.; Calleja-Agius, J. Cancer Stem Cells and Their Possible Implications in Cervical Cancer: A Short Review. Int. J. Mol. Sci. 2022, 23, 5167. [Google Scholar] [CrossRef]
- Mendoza-Almanza, G.; Rocha-Zavaleta, L.; Esparza-Ibarra, E.; Olmos, J.; Ortíz-Sánchez, E.; Rivas-Santiago, C. Cervical cancer stem cells and other leading factors associated with cervical cancer development (Review). Oncol. Lett. 2019, 18, 3423–3432. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Zheng, P.-S. High aldehyde dehydrogenase activity identifies cancer stem cells in human cervical cancer. Oncotarget 2013, 4, 2462–2475. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Liu, N.; Gao, Y.; Li, L. MiR-23b controls ALDH1A1 expression in cervical cancer stem cells. BMC Cancer 2017, 17, 292. [Google Scholar] [CrossRef]
- Giancotti, F.G. Mechanisms Governing Metastatic Dormancy and Reactivation. Cell 2013, 155, 750–764. [Google Scholar] [CrossRef]
- Ruíz, G.; Valencia-González, H.A.; León-Galicia, I.; García-Villa, E.; García-Carrancá, A.; Gariglio, P. Inhibition of RAD51 by siRNA and Resveratrol Sensitizes Cancer Stem Cells Derived from HeLa Cell Cultures to Apoptosis. Stem Cells Int. 2018, 2018, 2493869. [Google Scholar] [CrossRef] [PubMed]
- Heng, W.S.; Cheah, S.-C. Chelerythrine Chloride Downregulates β-Catenin and Inhibits Stem Cell Properties of Non-Small Cell Lung Carcinoma. Molecules 2020, 25, 224. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Wei, X.; Wang, Y. Embryonic stem cell markers Sox-2 and OCT4 expression and their correlation with WNT signal pathway in cervical squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 2470–2476. [Google Scholar]
- Zeltz, C.; Primac, I.; Erusappan, P.; Alam, J.; Noel, A.; Gullberg, D. Cancer-associated fibroblasts in desmoplastic tumors: Emerging role of integrins. Semin. Cancer Biol. 2019, 62, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, F.; Jiang, X.; Zhang, W.; Xiang, T.; Pan, Q.; Cai, L.; Zhao, J.; Weng, D.; Li, Y.; et al. CircITGB6 promotes ovarian cancer cisplatin resistance by resetting tumor-associated macrophage polarization toward the M2 phenotype. J. Immunother. Cancer 2022, 10, e004029. [Google Scholar] [CrossRef]
- Dijkgraaf, E.M.; Welters, M.J.; Nortier, J.W.; van der Burg, S.H.; Kroep, J.R. Interleukin-6/interleukin-6 receptor pathway as a new therapy target in epithelial ovarian cancer. Curr. Pharm. Des. 2012, 18, 3816–3827. [Google Scholar] [CrossRef] [PubMed]
- Salvagno, C.; Ciampricotti, M.; Tuit, S.; Hau, C.S.; Weverwijk, A.V.; Coffelt, S.B.; Kersten, K.; Vrijland, K.; Kos, K.; Ulas, T.; et al. Therapeutic targeting of macrophages enhances chemotherapy efficacy by unleashing type I interferon response. Nat. Cell Biol. 2019, 21, 511–521. [Google Scholar] [CrossRef]
- Stone, S.C.; Rossetti, R.A.; Lima, A.M.; Lepique, A.P. HPV associated tumor cells control tumor microenvironment and leuko-cytosis in experimental models. Immun. Inflamm. Dis. 2014, 2, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.D.; Xie, L.; Zheng, Z.R. The research of hypoxic induction factor-1 on angiogenesis and chemotherapy resistance of cervical carcinoma. Chin. J. Clin. Obstet. Gynecol. 2019, 19, 415–418. [Google Scholar]
- Zhou, B.; Yu, Y.; Yu, L.; Que, B.; Qiu, R. Sipi soup inhibits cancer-associated fibroblast activation and the inflammatory process by downregulating long non-coding RNA HIPK1-AS. Mol. Med. Rep. 2018, 18, 1361–1368. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, L.; Li, T.; Zhu, M.; Zhang, C.; Chen, L.; Zhao, P.; Zhou, H.; Yu, S.; Yang, X.; et al. The role of miR-451 in the switching between proliferation and mi-gration in malignant glioma cells: AMPK signaling, mTOR modulation and Rac1 activation required. Int. J. Oncol. 2017, 50, 1989–1999. [Google Scholar] [CrossRef]
- Wang, R.; Chen, D.Q.; Huang, J.Y.; Zhang, K.; Feng, B.; Pan, B.Z.; Chen, J.; De, W.; Chen, L.-B. Acquisition of radioresistance in docetaxel-resistant human lung adenocarcinoma cells is linked with dysregulation of miR-451/c-Myc-survivin/rad-51 signaling. Oncotarget 2014, 5, 6113–6129. [Google Scholar] [CrossRef]
- Pu, Z.; Ge, F.; Wang, Y.; Jiang, Z.; Zhu, S.; Qin, S.; Dai, Q.; Liu, H.; Hua, H. Ginsenoside-Rg3 inhibits the proliferation and invasion of hepatoma carcinoma cells via regulating long non-coding RNA HOX antisense intergenic. Bioengineered 2021, 12, 2398–2409. [Google Scholar] [CrossRef]
- Zhao, S.; Xu, K.; Jiang, R.; Li, D.Y.; Guo, X.X.; Zhou, P.; Tang, J.-F.; Li, L.-S.; Zeng, D.; Hu, L.; et al. Evodiamine inhibits proliferation and promotes apoptosis of hepatocellular carcinoma cells via the Hippo-Yes-Associated Protein signaling pathway. Life Sci. 2020, 251, 117424. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, M.Z.; Huang, Y.W.; Chen, M. Promoting effect of diosmetin on the cell cycle arrest and cell apoptosis in HepG2 cell and its mechanism. Hainan Med. J. 2016, 27, 354–357. [Google Scholar]
- Song, F.; Zhang, Y.; Pan, Z.; Zhang, Q.; Lu, X.; Huang, P. Resveratrol inhibits the migration, invasion and epithelial-mesenchymal transition in liver cancer cells through up-miR-186-5p expression. Zhejiang Da Xue Xue Bao Yi Xue Ban 2021, 50, 582–590. [Google Scholar]
- Shapira, A.; Livney, Y.D.; Broxterman, H.J.; Assaraf, Y.G. Nanomedicine for targeted cancer therapy: Towards the overcoming of drug resistance. Drug Resist. Updates 2011, 14, 150–163. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, X.Q. Research progress of P-gP inhibitory nanocarriers for the reversal of multidrug resistance in tumors. J. Modern Oncol. 2021, 29, 1808–1812. [Google Scholar]
- Zhou, Y.; Zhang, J.; Wang, K.; Han, W.; Wang, X.; Gao, M.; Wang, Z.; Sun, Y.; Yan, H.; Zhang, H.; et al. Quercetin overcomes colon cancer cells resistance to chemotherapy by inhibiting solute carrier family 1, member 5 transporter. Eur. J. Pharmacol. 2020, 881, 173185. [Google Scholar] [CrossRef]
- Iriti, M.; Kubina, R.; Cochis, A.; Sorrentino, R.; Varoni, E.M.; Kabała-Dzik, A.; Azzimonti, B.; Dziedzic, A.; Rimondini, L.; Wojtyczka, R.D. Rutin, a Quercetin Glycoside, Restores Chemosensitivity in Human Breast Cancer Cells. Phytother. Res. 2017, 31, 1529–1538. [Google Scholar] [CrossRef]
- Zheng, A.-W.; Chen, Y.-Q.; Zhao, L.-Q.; Feng, J.-G. Myricetin induces apoptosis and enhances chemosensitivity in ovarian cancer cells. Oncol. Lett. 2017, 13, 4974–4978. [Google Scholar] [CrossRef]
- Cui, M.Y.; He, H.L.; Li, P.; Mao, Y. Reversing effect of naringin on cisplatin resistance in human lung cancer A549/DDP cells. Chin. J. Pathophysiol. 2019, 35, 466–472. [Google Scholar]
- Zhu, H.; Gao, J.; Wang, L.; Qian, K.J.; Cai, L.P. In vitro study on reversal of ovarian cancer cell resistance to cisplatin by naringin via the nuclear factor-κB signaling pathway. Exp. Ther. Med. 2018, 15, 2643–2648. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Liu, J.F.; Qian, H.X. The effect and mechanism of apigenin reverse drugs resistant on A549/DDP cells. J. Nanjing Med. Univ. (Nat. Sci.) 2017, 37, 1385–1388+1394. [Google Scholar]
- Sadava, D.; Kane, S.E. Silibinin reverses drug resistance in human small-cell lung carcinoma cells. Cancer Lett. 2013, 339, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Zhang, M.Y.; Zhu, Q.S.; Lu, C.F.; Bai, X. Hyperin Enhances the Sensitivity of HCT8/VCR Colon Cancer Cell Line to Vincristine by Down-Regulating P-Glycoprotein. Clin. Lab. 2018, 64, 269–275. [Google Scholar] [CrossRef]
- Qin, J. Reversal Effect of EGCG on Cisplatin Resistance in Cervical Cancer Cell Line SiHa/DDP; University of South China: Hengyang, China, 2019. [Google Scholar]
- Yang, L.; Li, D.; Tang, P.; Zuo, Y. Curcumin increases the sensitivity of K562/DOX cells to doxorubicin by targeting S100 calci-um-binding protein A8 and P-glycoprotein. Oncol. Lett. 2020, 19, 83–92. [Google Scholar] [PubMed]
- Zhang, F.Y.; Du, G.J.; Zhang, L.; Zhang, C.L.; Lu, W.L.; Liang, W. Naringenin Enhances the Anti-Tumor Effect of Doxorubicin Through Selectively Inhibiting the Activity of Multidrug Resistance-Associated Proteins but not P-glycoprotein. Pharm. Res. 2009, 26, 914–925. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chen, W.; Qu, L.; Wu, J.; Si, J. Icaritin reverses multidrug resistance of HepG2/ADR human hepatoma cells via downreg-ulation of MDR1 and P-glycoprotein expression. Mol. Med. Rep. 2013, 8, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Niu, M.; Zhang, W.; Yan, J.; Li, J.; Tan, X.; Li, B.; Su, M.; Di, B.; Yan, F. Emodin reverses leukemia multidrug resistance by competitive inhibition and downregulation of P-glycoprotein. PLoS ONE 2017, 12, e0187971. [Google Scholar] [CrossRef]
- Guo, H.; Liu, F.; Yang, S.; Xue, T. Emodin alleviates gemcitabine resistance in pancreatic cancer by inhibiting MDR1/Pglycoprotein and MRPs expression. Oncol. Lett. 2020, 20, 167. [Google Scholar] [CrossRef]
- Ma, J.; Yang, J.; Wang, C.; Zhang, N.; Dong, Y.; Wang, C.; Wang, Y.; Lin, X. Emodin Augments Cisplatin Cytotoxicity in Platinum-Resistant Ovarian Cancer Cells via ROS-Dependent MRP1 Downregulation. BioMed Res. Int. 2014, 2014, 107671. [Google Scholar] [CrossRef]
- Chen, X.R.; Zhou, S.H.; Wang, R.C.; Xu, L. Effect and mechanism of β-elemene on SKOV3/DDP resistant ovarian cancer cells. Pharm. Clin. Res. 2021, 29, 81–85. [Google Scholar]
- Zhang, Y.; Qu, X.J.; Liu, Y.P.; Yang, X.H.; Hou, K.Z.; Teng, Y.E.; Zhang, J.D. Mechanism of synergistic anti-tumor effect of β-elemene with daunorubicin on human leukemia cells. J. Xi’an Jiaotong Univ. (Med. Sci.) 2011, 32, 664–667. [Google Scholar]
- Amerigos Daddy, J.C.K.; Chen, M.; Raza, F.; Xiao, Y.; Su, Z.; Ping, Q. Co-Encapsulation of Mitoxantrone and β-Elemene in Solid Lipid Nanoparticles to Overcome Multidrug Resistance in Leukemia. Pharmaceutics 2020, 12, 191. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, L.; Lu, X.; Wu, K.; Zeng, J.; Gao, Y.; Shi, Q.; Wang, X.; Chang, L.S.; He, D. Sinomenine reverses multidrug resistance in bladder cancer cells via P-glycoprotein-dependent and independent manners. Pharmazie 2014, 69, 48–54. [Google Scholar]
- Zuo, L.J.; Zhang, G.X.; Duan, J.M. Effects of Matrine on Multidrug Resistance of Human Bladder Cancer BIU-87/ADM Cells. Chin. J. Inf. Tradit. Chin. Med. 2016, 23, 73–75. [Google Scholar]
- Wang, L.; Liu, L.; Chen, Y.; Du, Y.; Wang, J.; Liu, J. Correlation between adenosine triphosphate (ATP)-binding cassette transporter G2 (ABCG2) and drug resistance of esophageal cancer and reversal of drug resistance by artesunate. Pathol. Res. Pract. 2018, 214, 1467–1473. [Google Scholar] [CrossRef]
- Kou, Y.; Tong, B.; Wu, W.; Liao, X.; Zhao, M. Berberine Improves Chemo-Sensitivity to Cisplatin by Enhancing Cell Apoptosis and Repressing PI3K/AKT/mTOR Signaling Pathway in Gastric Cancer. Front. Pharmacol. 2020, 11, 616251. [Google Scholar] [CrossRef]
- Qian, K.; Tang, C.Y.; Chen, L.Y.; Zheng, S.; Zhao, Y.; Ma, L.-S.; Xu, L.; Fan, L.-H.; Yu, J.-D.; Tan, H.-S.; et al. Berberine Reverses Breast Cancer Multidrug Resistance Based on Fluorescence Pharmacokinetics In Vitro and In Vivo. ACS Omega 2021, 6, 10645–10654. [Google Scholar] [CrossRef]
- Sui, H.; Liu, X.; Jin, B.H.; Pan, S.F.; Zhou, L.H.; Yu, N.A.; Li, Q. Zuo Jin Wan, a Traditional Chinese Herbal Formula, Reverses P-gp-Mediated MDR In Vitro and In Vivo. Evid. Based Complement. Altern. Med. 2013, 2013, 957078. [Google Scholar] [CrossRef] [PubMed]
- Gil, E.M.C. Targeting the PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer. Cancer Treat. Rev. 2014, 40, 862–871. [Google Scholar] [CrossRef]
- Best, S.A.; De Souza, D.P.; Kersbergen, A.; Ritchie, M.E.; McConville, M.J.; Sutherland, K.D.; Policheni, A.N.; Dayalan, S.; Tull, D. Synergy between the KEAP1/NRF2 and PI3K pathways drives non-small-cell lung cancer with an altered immune microenvironment. Cell Metab. 2018, 27, 935–943. [Google Scholar] [CrossRef]
- Zheng, J.; Asakawa, T.; Chen, Y.; Zheng, Z.; Chen, B.; Lin, M.; Liu, T.; Hu, J. Synergistic effect of baicalin and adriamycin in resistant HL-60/ADM leukaemia cells. Cell. Physiol. Biochem. 2017, 43, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, L.; Zhu, L.T.; Wang, Y.; Pan, D.; Yao, J.; You, Q.D.; Guo, Q.L. Wogonin reverses hypoxia resistance of human colon cancer HCT116 cells via downregulation of HIF-1α and glycolysis, by inhibiting PI3K/Akt signaling pathway. Mol. Carcinog. 2014, 53 (Suppl. S1), E107–E118. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, W.; Zhao, Q.; Wu, C.; Fan, C.; Lai, H.; Li, S. Combination of tanshinone IIA and doxorubicin possesses synergism and attenuation effects on doxorubicin in the treatment of breast cancer. Phytother. Res. 2019, 33, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Jia, Y.; Liu, Z.; Shu, X.; Liu, K. Resveratrol increases anti-proliferative activity of bestatin through downregu-lating P-glycoprotein expression via inhibiting PI3K/Akt/mTOR pathway in K562/ADR cells. J. Cell. Biochem. 2016, 117, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Kim, C.G.; Lim, Y.; Shin, S.Y.; Lee, Y.H. Curcumin down-regulates the multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NFκB pathway. Cancer Lett. 2008, 259, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Feng, S.; Yao, X.; Yuan, Z.; Liu, L.; Xie, Y. Nobiletin enhances the efficacy of chemotherapeutic agents in ABCB1 overex-pression cancer cells. Sci. Rep. 2015, 5, 18789. [Google Scholar] [CrossRef]
- Moon, J.Y.; Hung, L.V.M.; Unno, T.; Cho, S.K. Nobiletin Enhances Chemosensitivity to Adriamycin through Modulation of the Akt/GSK3β/β–Catenin/MYCN/MRP1 Signaling Pathway in A549 Human Non-Small-Cell Lung Cancer Cells. Nutrients 2018, 10, 1829. [Google Scholar] [CrossRef]
- Li, T.; Si, W.; Zhu, J.; Yin, L.; Zhong, C. Emodin reverses 5-Fu resistance in human colorectal cancer via downregulation of PI3K/Akt signaling pathway. Am. J. Transl. Res. 2020, 12, 1851–1861. [Google Scholar]
- Du, C.S.; Ma, Y.N.; Wang, S.; Zhang, F.; Zhang, J.; Sang, G.J. Study on reversal effects of levoshikonin on cisplatin resistance of human cervical carcinoma HeLa cells. China Pharm. 2020, 31, 1867–1873. [Google Scholar]
- Portilho, A.J.d.S.; da Silva, E.L.; Bezerra, E.C.A.; Gomes, C.B.d.S.M.R.; Ferreira, V.; de Moraes, M.E.A.; da Rocha, D.R.; Burbano, R.M.R.; Moreira-Nunes, C.A.; Montenegro, R.C. 1,4-Naphthoquinone (CNN1) Induces Apoptosis through DNA Damage and Promotes Upregulation of H2AFX in Leukemia Multidrug Resistant Cell Line. Int. J. Mol. Sci. 2022, 23, 8105. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Q.; Li, Y.; Liu, X.X.; Mao, X.G. Effects of triptolide combined with cisplatin on growth, apoptosis and invasion of human cervical cancer cell cisplatin-resistance line Hela/DDP. Acad. J. Chin. PLA Med. School 2020, 41, 518–522. [Google Scholar]
- Yuan, R.; Fan, Q.; Liang, X.; Han, S.; He, J.; Wang, Q.-Q.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits TGF-β1-induced epithelial–mesenchymal transition (EMT) in NSCLC through regulating ROS and PI3K/Akt/mTOR pathways. Chin. Med. 2022, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Z.; Liu, Z.Q.; Zhao, Y.X.; Sun, T.Z. Study on the improvement effect and mechanism of elemene on multidrug resistance of eisplatin-resistant human lung cancer A549 cell line. J. Zhejiang Chin. Med. Univ. 2021, 45, 16–22. [Google Scholar]
- Feng, L.; Yang, J.M.; Yang, R.; Sun, L.; Dong, L.L.; Zhang, Q.W. Study on reversal effect of quercetin on human cervical squamous carcinoma cisplatin-resistant cell line SiHa/DDP. China Pharm. 2021, 32, 2875–2879. [Google Scholar]
- Xu, Y.N.; Tang, J.; Ren, S.F.; Yi, Y.L. Luteolin reverses doxorubicin resistance in cervical cancer by regulating PI3K/Akt signaling pathway. Anti-Tumor Pharm. 2022, 12, 183–190. [Google Scholar]
- Zhou, B.G.; Wei, C.S.; Zhang, S.; Zhang, Z.; Gao, H.M. Matrine reversed multidrug resistance of breast cancer MCF-7/ADR cells through PI3K/AKT signaling pathway. J. Cell. Biochem. 2018, 119, 3885–3891. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Cai, J.B.; Wang, P. Effect of imperatorin on cisplatin resistance of cervical cancer cells. Zhejiang Pract. Med. 2016, 21, 391–393+397. [Google Scholar]
- Wang, X. Metformin Inhibits Cisplatin Resistance in Endometrial Cancer and Its Possible Related Mechanisms; University of South China: Hengyang, China, 2021. [Google Scholar]
- Niu, Y.J.; Shi, H.R.; Xie, Y. Study on the improvement effect of metformin on progestin-resistance of endometrial cancer. Mater. Child Health Care China 2016, 31, 605–607. [Google Scholar]
- Yang, Y.; Chen, Y.; Bai, J. Effect of β-elemene on growth of multidrug-resistance gastric cancer cells xenografts in nude mice. Prog. Anat. Sci. 2019, 25, 229–231+236. [Google Scholar]
- Wang, Z.H. Study on the Mechanism of Sijunzi Tang Improving gefitinib Resistance by Regulatig Glutamine Metabolism; Peking Union Medical College: Beijing, China, 2022. [Google Scholar]
- Lv, P.; Dong, Q.; Liu, Y.; Li, R.B.; Fan, Q.Y.; Chen, X.Y.; Hou, L. Effects of compound Zhebei Extract (CABE) on the expression of apoptotic protein and xenograft tumors of human colon cancer oxaliplatin-resistant cells (HCT-116/L-OHP) in nude mice: An experimental study. J. Beijing Univ. Tradit. Chin. Med. 2020, 43, 834–840. [Google Scholar]
- Lv, P.; Zhao, H.; Shi, F.Q.; Chen, X.Y.; Hou, L. Tumor suppression of the compound zhebei granules and cisplatin in the mice with L1210 /CDDP transplanted tumor and the impacts on relevant apoptotic proteins. World J. Integr. Tradit. West. Med. 2019, 14, 894–897+901. [Google Scholar]
- Wu, L.H. Correlation Analysis of Hepatitis Virus X Gene Mutation with Autophagy-Related Proteins (Beclin-1, LC3B and p62) on Hepatocellular Carcinoma; Guangxi Medical University: Nanning, China, 2019. [Google Scholar]
- Fernández, Á.F.; Sebti, S.; Wei, Y.; Zou, Z.; Shi, M.; McMillan, K.L.; He, C.; Ting, T.; Liu, Y.; Chiang, W.-C.; et al. Disruption of the beclin 1–BCL2 autophagy regulatory complex promotes longevity in mice. Nature 2018, 558, 136–140. [Google Scholar] [CrossRef]
- Lin, L.C.; Wu, C.H.; Shieh, T.M.; Chen, H.Y.; Huang, T.C.; Hsia, S.M. The licorice dietary component isoliquiritigenin chemosensi-tizes human uterine sarcoma cells to doxorubicin and inhibits cell growth by inducing apoptosis and autophagy via inhibition of m-TOR signaling. J. Funct. Foods 2017, 33, 332–344. [Google Scholar] [CrossRef]
- Li, J.; Duan, B.; Guo, Y.; Zhou, R.; Sun, J.; Bie, B.; Yang, S.; Huang, C.; Yang, J.; Li, Z. Baicalein sensitizes hepatocellular carcinoma cells to 5-FU and Epirubicin by activating apoptosis and ameliorating P-glycoprotein activity. Biomed. Pharmacother. 2018, 98, 806–812. [Google Scholar] [CrossRef]
- Choi, B.Y.; Joo, J.C.; Lee, Y.K.; Jang, I.-S.; Park, S.J.; Park, Y.J. Anti-cancer effect of Scutellaria baicalensis in combination with cisplatin in human ovarian cancer cell. BMC Complement. Altern. Med. 2017, 17, 277. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Luan, L.P. Effects of Baicalin on the sensitivity of C-33A/cis cells in cervical cancer by enhancing autophagy mediated cisplatin. Pharmacol. Clin. Chin. Materia Med. 2018, 34, 44–47. [Google Scholar]
- Peng, L.Q.; Wu, X.X.; Chen, G.; Cai, H.Z.; Tang, Y.P.; Chen, Q.Y.; Chen, X.Y. Effects of Wenyang Zhenshuai Granules on the Expression of Key Mitochondrial Autophagy Proteins in the Doxorubicin-Induced Model of H9c2 Cardiomyocyte Injury. Bull. Exp. Biol. Med. 2022, 173, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, C.; Meng, Q.; Wu, J.; Sun, H.; Sun, P.; Ma, X.; Huo, X.; Liu, K. Puerarin sensitized K562/ADR cells by inhibiting NF-κB pathway and inducing autophagy. Phytother. Res. 2021, 35, 1658–1668. [Google Scholar] [CrossRef]
- Wang, Y.Q. Hesperetin Reverses Drug Resistance and Mechanism of Cisplatin-Resistant Ovarian Adenocarcinoma Cell Lines A2780/DDP by Autophagy and Apoptosis; Fujian Medical University: Fuzhou, China, 2021. [Google Scholar]
- Li, Z.; Wang, N.; Yue, T.; Liu, L. Matrine reverses the drug resistance of K562/ADM cells to ADM and VCR via promoting au-tophagy. Transl. Cancer Res. 2020, 9, 786–794. [Google Scholar] [CrossRef]
- Chen, Y.Q. Effect of Lycorine on Proliferation, Autophagy and Apoptosis of K562 Cells Resistant to Imatinib and Its Mechanism; Lanzhou University: Lanzhou, China, 2022. [Google Scholar]
- Liu, R.; Xing, S.; Wang, L.; Zhang, Y.; Xu, B.H. Cepharanthine reverses icotinib resistance in human non-small cell lung cancer cells by regulating p53 signaling-mediated autophagy. J. Clin. Pulmonary Med. 2022, 27, 744–750+772. [Google Scholar]
- Zhou, K.; Yan, Y.; Li, B.; Liu, X. Reverse effect of olene on drug resistance of SPC-A-1/cisplatin in lung adenocarcinoma and its mechanism. Chin. J. Exp. Sur. 2018, 35, 119–122. [Google Scholar]
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; et al. The mechanism of m6A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by β-elemene. Cell Death Dis. 2020, 11, 969. [Google Scholar] [CrossRef] [PubMed]
- Le, F.Y. A Preliminary Study on Triptolide Inhibiting the Invasion and Migration of Drug-Resistant Ovarian Cancer Cells by Inducing Autophagy; Nanchang University: Nanchang, China, 2021. [Google Scholar]
- Jia, Y.L.; Li, J.F.; Liang, G.F. Effect of naringenin on cisplatin resistance of cervical cancer HeLa/DDP cells via AMPK pathway mediated autophagy. Drugs Clin. 2021, 36, 878–884. [Google Scholar]
- Pan, P.Y.; Zhou, H.; Xu, M.; Liu, C.Y.; Wang, C. Study on Kang’ai injection regulating Beclin 1-dependent autophagy-apoptosis interaction and improving cisplatin resistance in A549/DDP cells. Acad. J. Shanghai Univ. Tradit. Chin. Med. 2022, 36, 41–51. [Google Scholar]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. Berberine Reverses Doxorubicin Resistance by Inhibiting Autophagy Through the PTEN/Akt/mTOR Signaling Pathway in Breast Cancer. OncoTargets Ther. 2020, 13, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xu, J.; Guo, J.; Shang, Y.; Wang, J.; Wang, T. The enhancement of Tetrandrine to gemcitabine-resistant PANC-1 cytochemical sensitivity involves the promotion of PI3K/Akt/mTOR-mediated apoptosis and AMPK-regulated autophagy. Acta Histochem. 2021, 123, 151769. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Ling, X.; Chen, Y.; Wu, X.; Zhao, Z.; Wang, W.; Wang, S.; Lai, G.; Yu, Z. Hesperetin reverses P-glycoprotein-mediated cisplatin resistance in DDP-resistant human lung cancer cells via modulation of the nuclear factor-κB signaling pathway. Int. J. Mol. Med. 2020, 45, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; NishimuraI, N.; Okamoto, T.; Wada, K.; Naora, K. Sophora alopecuroides molecular mechanism of Matrine from in the reversing effect of multi-anticancer drug resistance in K562/ADR Cells. BioMed Res. Int. 2019, 2019, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.F.; Yu, H.B.; Wang, C.M.; Qiang, W.A.; Wang, S.P.; Zhang, J.M.; Yu, H.; Cui, L.; Wu, T.; Li, D.Q. Furanodiene induces extrinsic and intrinsic apoptosis in doxorubicin-resistant MCF-7 breast cancer cells via NF-κB-independent mechanism. Front. Pharmacol. 2017, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Dong, X.P.; Zhang, S.L.; You, Q.Y.; Jiang, X.T.; Zhao, X.G. Triptolide reverses the Taxol resistance of lung adenocarcinoma by inhibiting the NF-κB signaling pathway and the expression of NF-κB-regulated drug-resistant genes. Mol. Med. Rep. 2016, 13, 153–159. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, Z.-L.; Jiang, Q.-C.; Wang, S.-R. Schisandrin A reverses doxorubicin-resistant human breast cancer cell line by the inhibition of P65 and Stat3 phosphorylation. Breast Cancer 2018, 25, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, H.; Jiang, K.; Song, X.; Wang, X.; Yang, Y.; Liu, H.; Ji, Q.; Yu, X.; Liu, Y.; et al. Cudraxanthone L inhibits gastric cancer by regulating the MAPK signalling and promoting FAS-mediated pathway. Biomed. Pharmacother. 2021, 141, 111876. [Google Scholar] [CrossRef]
- He, Y.; Cao, X.; Kong, Y.; Wang, S.; Xia, Y.; Bi, R.; Liu, J. Apoptosis-promoting and migration-suppressing effect of alantolactone on gastric cancer cell lines BGC-823 and SGC-7901 via regulating p38MAPK and NF-κB pathways. Hum. Exp. Toxicol. 2019, 38, 1132–1144. [Google Scholar] [CrossRef]
- Han, Z.; Zhao, X.; Zhang, E.; Ma, J.; Zhang, H.; Li, J.; Xie, W.; Li, X. Resistomycin Induced Apoptosis and Cycle Arrest in Human Hepatocellular Carcinoma Cells by Activating p38 MAPK Pathway In Vitro and In Vivo. Pharmaceuticals 2021, 14, 958. [Google Scholar] [CrossRef]
- Zhu, Q.; Guo, Y.; Chen, S.; Fu, D.; Li, Y.; Li, Z.; Ni, C. Irinotecan Induces Autophagy-Dependent Apoptosis and Positively Regulates ROS-Related JNK- and P38-MAPK Pathways in Gastric Cancer Cells. OncoTargets Ther. 2020, 13, 2807–2817. [Google Scholar] [CrossRef]
- Li, B.; Zhou, M.; Wang, J.; Xu, H.; Yang, M. Suppressing ERK Pathway Impairs Glycochenodeoxycholate-Mediated Survival and Drug-Resistance in Hepatocellular Carcinoma Cells. Front. Oncol. 2021, 11, 663944. [Google Scholar] [CrossRef] [PubMed]
- Bonney, E.A. Mapping out p38MAPK. Am. J. Reproduct. Immunol. 2017, 77, e12652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lu, M.; Zhang, Z.; Tian, X.; Wang, S.; Lv, D. Resveratrol reverses P-glycoprotein-mediated multidrug resistance of U2OS/ADR cells by suppressing the activation of the NF-κB and p38 MAPK signaling pathways. Oncol. Lett. 2016, 12, 4147–4154. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Gao, Y.; Sun, L.; Liu, J.; Chen, H.; Yu, L.; Chen, Z.; Chen, W.; Lin, L. Rosmarinic acid reverses non-small cell lung cancer cisplatin resistance by activating the MAPK signaling pathway. Phytother. Res. 2020, 34, 1142–1153. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Mei, J.; Tan, Y. Baicalin attenuates DDP (cisplatin) resistance in lung cancer by downregulating MARK2 and p-Akt. Int. J. Oncol. 2016, 50, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.B.; Zhang, L.X.; Luo, Q.; Ren, H.Y.; Niu, S.R. Curcumin reverses the drug resistance of esophageal cancer Eca-109/VCR cells. Chin. J. Clin. Pharmacol. 2021, 37, 136–139. [Google Scholar]
- Cao, C.; Huang, L.J.; Huang, Z.S.; Hu, G.Y.; Deng, Z.H.; Li, G.Z.; Lu, W.Q.; Zhong, Q.H. Effect of curcumin on proliferation and adriamycin-resistance of drug-resistant hepatocarcinoma cell HepG2/ADM. Guangxi Med. J. 2020, 42, 976–980. [Google Scholar]
- Li, M.; Cai, O.; Yu, Y.; Tan, S. Paeonol inhibits the malignancy of Apatinib-resistant gastric cancer cells via LINC00665/miR-665/MAPK1 axis. Phytomedicine 2022, 96, 153903. [Google Scholar] [CrossRef]
- Zhong, S.S.; Yuan, Y.P.; Xin, L.Y.; Chen, Y.J.; Zhang, L.Q. Reversal of drug resistance in K562/ADM cells caused by RA and the related mechanisms. J. Exp. Hematol. 2021, 29, 1704–1709. [Google Scholar]
- Kwak, A.-W.; Park, J.W.; Lee, S.-O.; Lee, J.-Y.; Seo, J.-H.; Yoon, G.; Lee, M.-H.; Choi, J.-S.; Shim, J.-H. Isolinderalactone sensitizes oxaliplatin-resistance colorectal cancer cells through JNK/p38 MAPK signaling pathways. Phytomedicine 2022, 105, 154383. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Jiang, X.X.; Wang, Y.; Ding, X.; Li, Z.Q.; Gao, N. Tetrandrine sensitizes vincristine to induce apoptosis in SGC-7901/VCR cells through MAPK signaling pathway. J. Army Med. Univ. 2022, 44, 691–699. [Google Scholar]
- Lu, P.; Fan, J.J.; Luo, X. The effect of Muscone on lung cancer cells resistance to cisplatin and tumor growth in mice. J. Guangxi Med. Univ. 2020, 37, 1948–1953. [Google Scholar]
- Ye, K.; Wei, Q.; Gong, Z.; Huang, Y.; Liu, H.; Li, Y.; Peng, X. Effect of norcantharidin on the proliferation, apoptosis, and cell cycle of human mesangial cells. Ren. Fail. 2017, 39, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Cyclin-dependent protein kinase inhibitors including palbociclib as anticancer drugs. Pharmacol. Res. 2016, 107, 249–275. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Liang, H.; Qu, B.; Li, Y.; Liu, J.; Chen, C.; Zhang, D.; Zhang, X.; Gao, A. Moxidectin inhibits glioma cell viability by inducing G0/G1 cell cycle arrest and apoptosis. Oncol. Rep. 2018, 40, 1348–1358. [Google Scholar] [CrossRef]
- Lohberger, B.; Leithner, A.; Stuendl, N.; Kaltenegger, H.; Kullich, W.; Steinecker-Frohnwieser, B. Diacerein retards cell growth of chondrosarcoma cells at the G2/M cell cycle checkpoint via cyclin B1/CDK1 and CDK2 downregulation. BMC Cancer 2015, 15, 891–991. [Google Scholar] [CrossRef]
- Xu, J.; Guo, Z.; Wang, Q.Y.; Liang, D.X. Reversal effect of shikonin on cisplatin resistance of ovarian cancer SKOV3/DDP cells. Chin. J. Pathophysiol. 2018, 34, 1616–1621. [Google Scholar]
- Giacomini, I.; Cocetta, V.; Carrara, M.; Ragazzi, E.; Montopoli, M. Plumbagin Induces Cell Cycle Arrest and Apoptosis in A431 Cisplatin-Resistant Cancer Cells. Nat. Prod. Commun. 2020, 15, 1934578X20921627. [Google Scholar] [CrossRef]
- Wang, S.; Lei, T.; Zhang, M. The Reversal Effect and Its Mechanisms of Tetramethylpyrazine on Multidrug Resistance in Human Bladder Cancer. PLoS ONE 2016, 11, e0157759. [Google Scholar] [CrossRef]
- Huang, S.Y. Reversal Effect of HTT on Drug Resistance of Vemurafenib Melanoma Cells; Chongqing Medical University: Chongqing, China, 2021. [Google Scholar]
- Yang, J.; Wang, D.D.; Liang, Y.Q.; Ding, L.N.; Zhuo, J.R.; Liang, G.P. Antitumor activities of triptolide succinate monoester on A549/DDP cells. Chem. Rea. 2022, 44, 1128–1135. [Google Scholar]
- Wang, X.Y.; Pei, X.D.; He, Z.L.; Lin, J.W.; Li, Y.Z.; Chen, W.Q.; Liu, X.L.; Jinag, L.H. In vitro activity of Kamebakaurin on hepatocellular carcinoma resistant strain HepG2/ADM cells. Anhui Med. Pharm. J. 2021, 25, 2131–2135. [Google Scholar]
- Liu, G.-W.; Liu, Y.-H.; Jiang, G.-S.; Ren, W.-D. The reversal effect of Ginsenoside Rh2 on drug resistance in human colorectal carcinoma cells and its mechanism. Hum. Cell 2018, 31, 189–198. [Google Scholar] [CrossRef]
- Li, G.; Fang, S.; Shao, X.; Li, Y.; Tong, Q.; Kong, B.; Chen, L.; Wang, Y.; Yang, J.; Yu, H.; et al. Curcumin Reverses NNMT-Induced 5-Fluorouracil Resistance via Increasing ROS and Cell Cycle Arrest in Colorectal Cancer Cells. Biomolecules 2021, 11, 1295. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.N.; Guo, H.Y.; Wang, T.; Zhang, Y.D.; Guo, W.J.; Jiang, B.; Su, H.X. Synergistic inhibitory effect of isoxanthohumol combined with cisplatin on cisplatin resistant A549/DDP cells in lung cancer. Chin. Pharmacol. Bull. 2021, 37, 1429–1434. [Google Scholar]
- Sun, Y.C.; Jin, E.H. Reversal effect of metformin on drug resistance of oxaliplatin in human gastric cancer SGC-7901/L-OHP cells. Chin. J. Clin. Res. 2018, 31, 499–504. [Google Scholar]
- Hu, J.N.; Song, Z.B.; Fu, D.L.; Luo, P.P.; Xu, Y.Y. Effect and mechanism of buzhong yiqi decoction on enhancing chemosensitivity of cisplatin-resistant human gastric cancer cells line. Chin. J. Integr. Tradit. West. Med. Dig. 2022, 30, 90–95. [Google Scholar]
- Lei, Y.; Gan, H.; Huang, Y.; Chen, Y.; Chen, L.; Shan, A.; Zhao, H.; Wu, M.; Li, X.; Ma, Q.; et al. Digitoxin inhibits proliferation of multidrug-resistant HepG2 cells through G2/M cell cycle arrest and apoptosis. Oncol. Lett. 2020, 20, 71. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shi, P.; Zhao, G.; Xu, J.; Peng, W.; Zhang, J.; Zhang, G.; Wang, X.; Dong, Z.; Chen, F.; et al. Targeting Cancer Stem Cell Pathways for Cancer Therapy. Signal Transduct. Target. Ther. 2020, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L. The Effect and Mechanism Study of Luteolin on the Stemness Property in Resistant Esophageal Cancer Cells; Zhengzhou University: Zhengzhou, China, 2020. [Google Scholar]
- Zhao, J.; Li, L.; Wang, Z.; Li, L.; He, M.; Han, S.; Dong, Y.; Liu, X.; Zhao, W.; Ke, Y.; et al. Luteolin attenuates cancer cell stemness in PTX-resistant oesophageal cancer cells through mediating SOX2 protein stability. Pharmacol. Res. 2021, 174, 105939. [Google Scholar] [CrossRef]
- Wang, X.Y.; Zhao, Q.; Wang, B.; Yuan, S.; Li, K. Mechanism of Quercetin reversing drug resistance of human breast cancer MCF-7 cells to doxorubicin. Mod. Prevent. Med. 2018, 45, 1844–1849+1859. [Google Scholar]
- Su, J.W.; Zhou, X.H.; Ye, Y.X.; Jiang, Q. Effect of matrine on reversing chemotherapy resistance of human colon cancer stem cells and its related mechanism. Curr. Immunol. 2020, 40, 454–459+464. [Google Scholar]
- Manogaran, P.; Somasundaram, B.; Viswanadha, V.P. Reversal of cisplatin resistance by neferine/isoliensinine and their combi-natorial regimens with cisplatin-induced apoptosis in cisplatin-resistant colon cancer stem cells (CSCs). J. Biochem. Mol. Toxicol. 2022, 36, e22967. [Google Scholar] [CrossRef]
- Wang, W.J.; Wu, S.F.; Guo, P.T.; Lu, Y.Y.; Wan, Y.T.; Zeng, H.; Qi, C. Reversing effects of ursolic acid on drug resistance in nude mice bearing ovarian cancer stem cells. Shanghai J. Tradit. Chin. Med. 2016, 50, 70–76. [Google Scholar]
- Li, Y.L.; Lei, F.F.; Fan, X.; Zhu, M.; Chi, X.Y.; Li, G. The Inhibitory Effect of Curcumin on Chemoresistance of Colon Cancer Cells. J. Hubei Univ. Med. 2021, 40, 338–341+348. [Google Scholar]
- Huang, W.; Wen, F.; Gu, P.; Liu, J.; Xia, Y.; Li, Y.; Zhou, J.; Song, S.; Ruan, S.; Gu, S.; et al. The inhibitory effect and mechanism of Yi-qi-hua-yu-jie-du decoction on the drug resistance of gastric cancer stem cells based on ABC transporters. Chin. Med. 2022, 17, 93. [Google Scholar] [CrossRef]
- Mordhorst, L.B.; Ahlin, C.; Sorbe, B. Prognostic impact of the expression of Hedgehog proteins in cervical carcinoma FIGO stages I–IV treated with radiotherapy or chemoradiotherapy. Gynecol. Oncol. 2014, 135, 305–311. [Google Scholar] [CrossRef]
- Regan, J.L.; Schumacher, D.; Staudte, S.; Steffen, A.; Haybaeck, J.; Keilholz, U.; Schweiger, C.; Golob-Schwarzl, N.; Mumberg, D.; Henderson, D.; et al. Non-Canonical Hedgehog Signaling Is a Positive Regulator of the WNT Pathway and Is Required for the Survival of Colon Cancer Stem Cells. Cell Rep. 2017, 21, 2813–2828. [Google Scholar] [CrossRef]
- De Sousa e Melo, F.; Kurtova, A.V.; Harnoss, J.M.; Kljavin, N.; Hoeck, J.D.; Hung, J.; Anderson, J.E.; Storm, E.E.; Modrusan, Z.; Koeppen, H.; et al. A distinct role for Lgr5+ stem cells in primary and metastatic colon cancer. Nature 2017, 543, 676–680. [Google Scholar] [CrossRef]
- Li, C.; Tang, C.N.; He, G.S. Inhibition of genistein on characteristics of liver cancer stem-like cells and its effect on fluoroura-cil-resistance. Hainan Med. J. 2015, 26, 1108–1111. [Google Scholar]
- Qian, Q.H.; Chen, R.F.; Li, J.; An, C.P.; Zhu, Y.; Cao, Q. Effects of norcantharidin on cancer stem cell activity of drug resistant ovarian cancer cells via Hedgehog signaling pathway. China J. Tradit. Chin. Med. Pharm. 2021, 36, 2926–2930. [Google Scholar]
- Fath, M.K.; Ebrahimi, M.; Nourbakhsh, E.; Hazara, A.Z.; Mirzaei, A.; Shafieyari, S.; Salehi, A.; Hoseinzadeh, M.; Payandeh, Z.; Barati, G. PI3K/Akt/mTOR signaling pathway in cancer stem cells. Pathol.-Res. Pract. 2022, 237, 154010. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.P.; Zhuang, Y.F.; Cheng, J.; Li, Q. Curcumin increased the sensitivity of gastric cancer cells to cisplatin by inhibiting the Wnt signaling pathway. J. Region. Anatomy Operat. Surg. 2021, 30, 749–753. [Google Scholar]
- Wang, A.; Ku, H.; Liang, Y.; Chen, Y.; Hwu, Y.; Yeh, T. The autonomous notch signal pathway is activated by baicalin and baicalein but is suppressed by niclosamide in K562 cells. J. Cell. Biochem. 2009, 106, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Wang, Z.H.; Ma, G. Effects of sinomenine combined with cisplatin on proliferation and apoptosis of cisplatin- resistant cell lines. Med. J. Chin. People’s Armed Police Force 2022, 33, 158–162. [Google Scholar]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.C.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.B.; Chen, J.Y.; Tong, W.; Tian, F. Research progress in drug resistance and countermeasures of targeted therapy in non-small cell lung cancer. Oncol. Progr. 2020, 18, 1625–1627+1667. [Google Scholar]
- Elaskalani, O.; Razak, N.B.A.; Falasca, M.; Metharom, P. Epithelial-mesenchymal transition as a therapeutic target for overcoming chemoresistance in pancreatic cancer. World J. Gastrointest. Oncol. 2017, 9, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.Y.; Yuan, G.L.; Ma, S.S.; Luo, L.M.; Xu, Y. Study on the mechanism of quercetin improving gemcitabine resistance in NSCLC through the EMT pathway mediated by miR-101/EZH2 axis. Tianjin Med. J. 2022, 50, 125–130. [Google Scholar]
- Zhang, Y.; Wang, C.; Yu, D.; Gao, Y.; Jing, H.; Wang, Y.; Liu, C.Y. Astragalus polysaccharides improve cisplatin resistance by inhibiting EMT of lung adenocarcinoma A549/DDP cells transplanted into nude mice. Chin. J. Exp. Tradit. Med. Formulae 2022, 28, 79–85. [Google Scholar]
- Wu, J.; Mo, S.X.; Wei, Y.T.; Feng, Z.Q.; Liang, L. Reversal of Cisplatin Resistance in Lung Cancer A549 Cells by Matrine-mediated Cell Epithelial-mesenchymal Transition. Herald Med. 2021, 40, 1312–1317. [Google Scholar]
- Hsieh, M.J.; Wang, C.W.; Lin, J.T.; Chuang, Y.C.; Hsi, Y.; Lo, Y.S.; Lin, C.C.; Chen, M.K. Celastrol, a plant-derived triterpene, induces cisplatin-resistance nasopharyngeal carcinoma cancer cell apoptosis though ERK1/2 and p38 MAPK signaling pathway. Phytomedicine 2019, 58, 152805. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.B.; Qiu, H.P.; Jin, Z.; Jiang, L.Y.; Yao, S.H. Reverse effect of Rubesin on fluvestrin resistance in breast cancer MCF-7 cells and its mechanism. Chin. J. Pharmacol. Toxicol. 2021, 35, 809–815. [Google Scholar]
- Yao, C.; Jiang, J.; Tu, Y.; Du, H.; Zhang, Y. β-elemene reverses the drug resistance of A 549/DDP lung cancer cells by activating intracellular redox system, decreasing mitochondrial membrane potential and P-glycoprotein expression, and inducing apop-tosis. Thorac. Cancer 2014, 5, 304–312. [Google Scholar] [CrossRef]
- Wang, Z.R.; Fan, H.F.; Zhang, Q.; Guo, J.; Li, D.H. Blockade of JAK2-STAT3 Signaling Pathway by β-elemene Promoting Prolif-eration and Apoptosis of Lung Cancer Cells by Paclitaxel. Chin. Archives Tradit. Chin. Med. 2019, 37, 1600–1604. [Google Scholar]
- Shilnikova, K.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Park, J.E.; Hyun, Y.J.; Zhen, A.X.; Jeong, Y.J.; Jung, U.; Kim, I.G.; et al. Shikonin induces mitochondria-mediated apoptosis and attenuates epithelial-mesenchymal transition in cisplatin-resistant human ovarian cancer cells. Oncol. Lett. 2018, 15, 5417–5424. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, F.; Nan, Y.; Qu, L.; Na, W.; Jia, C.; Chen, X. PKM2 Inhibitor Shikonin Overcomes the Cisplatin Resistance in Bladder Cancer by Inducing Necroptosis. Int. J. Biol. Sci. 2018, 14, 1883–1891. [Google Scholar] [CrossRef]
- Zheng, F.; Li, J.; Ma, C.; Tang, X.; Tang, Q.; Wu, J.; Chai, X.; Xie, J.; Yang, X.; Hann, S.S. Novel regulation of miR-34a-5p and HOTAIR by the combination of berberine and gefitinib leading to inhibition of EMT in human lung cancer. J. Cell. Mol. Med. 2020, 24, 5578–5592. [Google Scholar] [CrossRef]
- Deng, M.; Liu, B.; Song, H.; Yu, R.; Zhou, D.; Chen, Y.; Ma, Y.; Lv, F.; Xu, L.; Zhang, Z.; et al. β-Elemene inhibits the metastasis of multidrug-resistant gastric cancer cells through miR-1323/Cbl-b/EGFR pathway. Phytomedicine 2020, 69, 153184. [Google Scholar] [CrossRef]
- Wang, Y.; Mou, Q.R.; Liu, C.Y.; Yuan, G. Effects of Buzhong Yiqi Decoction combined with Cisplatin on expression of β-catenin mRNA and protein in transplantation tumor of nude mice with human lung cancer. China J. Tradit. Chin. Med. Pharm. 2022, 37, 2191–2194. [Google Scholar]
- Wang, W.; Deng, T.H.; Jin, C.H.; Zhao, S.L.; Fu, Y.; Xiao, X.F.; Tan, L.; Rui, M.; Fan, F.Y. Discuss the Intervention effect of Jiedu Fuzheng prescription on a mouse model of lung cancer with acquired drug-resistance to molecular-targeted therapy. Hunan J. Tradit. Chin. Med. 2020, 36, 160–164. [Google Scholar]
- Palmer, A.C.; Sorger, P.K. Combination cancer therapy can confer benefit via patient-to-patient variability without drug additivity or synergy. Cell 2017, 171, 1678–1691.e13. [Google Scholar] [CrossRef]
- Wu, L.; Gao, J.; Zhang, Y.; Sui, B.; Wen, Y.; Wu, Q.; Liu, K.; He, S.; Bo, X. A hybrid deep forest-based method for predicting synergistic drug combinations. Cell Rep. Methods 2023, 3, 100411. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Jiang, P.; Wang, P.Q.; Yang, C.S.; Wang, X.R.; Feng, Q. EGCG enhances cisplatin sensitivity by regulating expression of the copper and cisplatin influx transporter CTR1 in ovary cancer. PLoS ONE 2015, 10, e0125402. [Google Scholar]
- Zhou, D.-H.; Wang, X.; Yang, M.; Shi, X.; Huang, W.; Feng, Q. Combination of Low Concentration of (−)-Epigallocatechin Gallate (EGCG) and Curcumin Strongly Suppresses the Growth of Non-Small Cell Lung Cancer in Vitro and in Vivo through Causing Cell Cycle Arrest. Int. J. Mol. Sci. 2013, 14, 12023–12036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, S.-X.; Ma, J.-W.; Li, H.-Y.; Ye, J.-C.; Xie, S.-M.; Du, B.; Zhong, X.-Y. EGCG inhibits properties of glioma stem-like cells and synergizes with temozolomide through downregulation of P-glycoprotein inhibition. J. Neuro-Oncol. 2015, 121, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wei, F.; Wang, Y.; Wu, B.; Fang, Y.; Xiong, B. EGCG synergizes the therapeutic effect of cisplatin and oxaliplatin through au-tophagic pathway in human colorectal cancer cells. J. Pharmacol. Sci. 2015, 128, 27–34. [Google Scholar] [CrossRef]
- Shin, D.M.; Amin, A.R.M.; Wang, D.; Rahman, M.A.; Nannapaneni, S.; Khuri, F.R.; Chen, Z.G. 1187 molecular mechanism of synergistic anti-tumor activity by the combination of natural compounds Green tea (−)epigallocathetin-3-gallate (EGCG) and resveratrol for potential chemoprevention in head and neck cancer (HNC). Eur. J. Cancer 2012, 48, S286. [Google Scholar] [CrossRef]
- Wang, W.; Chen, D.; Zhu, K. SOX2OT variant 7 contributes to the synergistic interaction between EGCG and Doxorubicin to kill osteosarcoma via autophagy and stemness inhibition. J. Exp. Clin. Cancer Res. 2018, 37, 37. [Google Scholar] [CrossRef]
- Kumazoe, M.; Tsukamoto, S.; Lesnick, C.; Kay, N.E.; Yamada, K.; Shanafelt, T.D.; Tachibana, H. Vardenafil, a clinically available phosphodiesterase inhibitor, potentiates the killing effect of EGCG on CLL cells. Br. J. Haematol. 2015, 168, 610–613. [Google Scholar] [CrossRef]
- Haque, A.; Rahman, M.A.; Chen, Z.G.; Saba, N.F.; Khuri, F.R.; Shin, D.M.; Amin, A.R.M.R. Combination of erlotinib and EGCG induces apoptosis of head and neck cancers through posttranscriptional regulation of Bim and Bcl-2. Apoptosis 2015, 20, 986–995. [Google Scholar] [CrossRef]
- Zhao, C.; Gao, W.; Chen, T. Synergistic induction of apoptosis in A549 cells by dihydroartemisinin and gemcitabine. Apoptosis 2014, 19, 668–681. [Google Scholar] [CrossRef]
- Feng, X.; Li, L.; Jiang, H.; Jiang, K.; Jin, Y.; Zheng, J. Dihydroartemisinin potentiates the anticancer effect of cisplatin via mTOR inhibition in cisplatin-resistant ovarian cancer cells: Involvement of apoptosis and autophagy. Biochem. Biophys. Res. Commun. 2014, 444, 376–381. [Google Scholar] [CrossRef]
- Zong, L.; Cheng, G.; Liu, S.; Pi, Z.; Liu, Z.; Song, F. Reversal of multidrug resistance in breast cancer cells by a combination of ursolic acid with doxorubicin. J. Pharm. Biomed. Anal. 2019, 165, 268–275. [Google Scholar] [CrossRef]
- Liu, Y.; Ni, J.; Liu, L.; Li, F.; Yin, X. Tanshinone IIA inhibits adriamycin resistance in gastric cancer cells. Chin. J. Physiol. 2019, 35, 2208–2214. [Google Scholar]
- Xu, Z.; Chen, L.; Xiao, Z.; Zhu, Y.; Hui, J.; Yan, J.; Cheng, G.; Yilai, W.; Lin, W.; Wen, Z.; et al. Potentiation of the anticancer effect of doxorubicin in drug-resistant gastric cancer cells by tanshinone IIA. Phytomedicine 2018, 51, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, P.; Huang, W.; Liu, L. Experimental study on effects of shengmai injection: Enhancing 5-FU anti-tumor efficacy and reducing its toxicity. J. Chin. Integr Med. 2005, 3, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Yan, M. Study on Mechanism of Shengmai Injection Reversing Multidrug Resistance in Gastric Cancer Based on JNK Signaling Pathway. Master’s Thesis, Shanxi Traditional Chinese Medical Hospital, Taiyuan, China, 2017. [Google Scholar]
- Li, W.; Li, Y.; Yu, C.; He, Y. Reversal of multidrug resistance by the Chinese medicine Yiqi Jianpi Huaji Decoction and the mechanism of action in human gastric cancer SGC7901/VCR cells. Evid. Based Complement. Altern. Med. 2015, 2015, 390812. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-L.; Shen, H.-L.; Ao, Z.-F.; Chen, B.-A.; Xia, W.; Gao, F.; Zhang, Y.-N. Combination of tetrandrine as a potential-reversing agent with daunorubicin, etoposide and cytarabine for the treatment of refractory and relapsed acute myelogenous leukemia. Leuk. Res. 2006, 30, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Sun, H.; Wang, F. H1, a novel derivative of tetrandrine reverse P-glycoprotein- mediated multidrug resistance by in-hibiting transport function and expression of P-glycoprotein. Cancer Chemother. Pharmacol. 2011, 67, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.-L.; Wang, W.; Ho, C.-T. 7,3′,4′-Trihydroxyisoflavone modulates multidrug resistance transporters and induces apoptosis via production of reactive oxygen species. Toxicology 2012, 302, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mu, X.-D.; Li, E.-Z.; Luo, Y.; Song, N.; Qu, X.-J.; Hu, X.-J.; Liu, Y.-P. The Role of E3 Ubiquitin Ligase Cbl Proteins in β-Elemene Reversing Multi-Drug Resistance of Human Gastric Adenocarcinoma Cells. Int. J. Mol. Sci. 2013, 14, 10075–10089. [Google Scholar] [CrossRef] [PubMed]
- Bin, T.E.; Asif, S.; Rafi, A.M.; Rahman, T.; Abir, M.H.; Faijanur-Rob Siddiquee, M.; Ahmed, H.; Rahman, N.; Nainu, F.; Wahyudin, E.; et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches#13. Front. Oncol. 2022, 12, 891652. [Google Scholar]
- Abdur, R.; Tareq, A.; Ahmed, A.K.; Imran, M.; Shah, Z.A.; Emran, T.B.; Mitra, S.; Khan, Z.; Alhumaydi, F.A.; Aljohani, A.S.M.; et al. Berberine as a Potential Anticancer Agent: A Comprehensive Review. Molecules 2021, 26, 7368. [Google Scholar]
- Gu, K.J.; Li, G. An Overview of Cancer Prevention: Chemoprevention and Immunoprevention. J. Cancer Prev. 2020, 25, 127–135. [Google Scholar] [CrossRef]
- Umar, A. Cancer Immunoprevention: A New Approach to Intercept Cancer Early. Cancer Prev. Res. 2014, 7, 1067–1071. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line and In Vivo Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Flavonoids | Quercetin | 33 μmol/L | Doxorubicin resistant human colon cancer | SW620/Ad300 | Increased: accumulation of drug, apoptosis | [137] |
| Rutin | 20 μmol/L | Human breast cancer | MDA-MB-231 MCF-7 | Decreased: P-gP | [138] | |
| Myricetin | 25 μmol/L | Ovarian cancer | A2780 OVCAR3 | Increased: Bax, cleaved caspase 3 Decreased: Bcl-2, MDR1 | [139] | |
| Naringin | 129.77 μmol/L | Cisplatin resistant human lung cancer | A549/DDP | Increased: Bax, cleaved caspase 3 Decreased: Bcl-2, P-gP, MRP1, p-AKT, CXCR4 | [140] | |
| 10–40 μmol/L | Cisplatin resistant ovarian cancer | SKOV3/CDDP | Decreased: NF-κB, P-gP | [141] | ||
| Apigenin | 5–80 mol/L | Cisplatin resistant human lung cancer | A549/DDP | Decreased: LRP, P-gP | [142] | |
| Silibinin | 60 μmol/L | Multidrug resistant small-cell lung carcinoma | VPA17 | Decreased: drug efflux | [143] | |
| Polyphenols | Hyperin | 12.5 μmol/L | Vincristine resistant colorectal cancer | HCT8/VCR | Decreased: P-gP | [144] |
| (-)-Epigallocatechin 3-gallate | 15.81 µg/ml | Cisplatin resistant cervical cancer | SiHa/DDP | Decreased: P-gP, ABCG2 | [145] | |
| Curcumin | 2.5 µmol/L | Doxorubicin resistant human leukemia | K562/DOX | Increased: intracellular DOX Decreased: S100A8, P-gP | [146] | |
| Naringenin | 100 μmol/L | Doxorubicin resistant human breast cancer | MCF-7/DOX | Increased: uptake of 5-CFDA, doxorubicin | [147] | |
| Icaritin | 0–30 μmol/L | Adriamycin resistant human hepatoma | HepG2/ADR | Decreased: MDR gene, P-gP | [148] | |
| Quinones | Emodin | 0–20 μmol/L | Adriamycin resistant human myelogenous myeloid leukemia | K562/ADM | Increased: intracellular accumulation of rhodamine 123 Decreased: P-gP | [149] |
| 40 mg/kg | Gemcitabine resistant human pancreatic cancer mice model | gemcitabine-resistant PANC-1 cell | Decreased: MDR1, P-gP, MRP1, MRP5 | [150] | ||
| 50 µmol/L | Platinum resistant ovarian cancer | COC1/DDP | Decreased: P-gP | [151] | ||
| artesunate | 25, 50 mg/kg | drug-resistant esophageal cancer | Eca109/ABCG2, Xenografts of Eca109/ABCG2 in BALB/c nu/nu mice | Increased: apoptosis Decreased: Tumor volume, ABCG2 | [157] | |
| Terpenoids | β-elemene | 20–320 µmol/L | Cisplatin resistant ovarian cancer | SKOV3/DDP | Decreased: ABCB1, LRP, P-gP | [152] |
| 31.92 µg/mL | Daunorubicin resistant on human leukemia | K562/DNR | Decreased: P-gP | [153] | ||
| 22.02 µg/mL | Doxorubicin resistant on human leukemia | K562/DOX | Decreased: P-gP | [154] | ||
| Alkaloids | Sinomenine | 100, 200 µg/mL | Doxorubicin resistant human bladder cancer | 253J/DOX | Increased: cleaved PARP, Bax Decreased: P-gP | [155] |
| Matrine | 32.5 µg/mL | Adriamycin resistant human bladder cancer | BIU-87/ADM | Decreased: P-gP, adriamycin efflux | [156] | |
| Berberine | BGC-823/DDP: 549.6 µmol/L SGC-7901/DDP: 562.1 µmol/L | Cisplatin resistant gastric cancer | BGC-823/DDP, SGC-7901/DDP, Xenografts of SGC-7901/DDP in BALB/c-nu mice | Increased: c-caspase 3, c-caspase9, Bax Decreased: MDR1, MRP1, and PI3K/AKT/mTOR signaling pathway related protein | [158] | |
| 20 μg/mL | doxorubicin resistant breast cancer | MCF-7/DoxFluc, Xenografts of MCF-7/DoxFluc in BALB/c nude mice | Decreased: P-gP/ABCB1, MRP1/ABCC1 | [159] | ||
| Compounds | Zuojinwan | 1027.5–4110 mg/kg | drug-resistant cancer | HCT116/L-OHP, SGC7901/DDP, Bel/Fu, Xenografts of HCT116/L-OHP in male athymic nude mice | Decreased: P-gP | [160] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line and In Vivo Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Polyphenols | Grape seed procyanidin | 10–40 µmol/L | Paclitaxel-resistant ovarian cancer | A2780/T | Increased: accumulation of Rho-123, p65, YB-1 Decreased: P-gP, phospho-AKT, phospho-IκBα, p-ERK1/2 | [49] |
| Baicalin | 5, 10 μmol/L | Adriamycin-resistant leukemia cell | HL-60/ADM | Increased: cleaved PARP, cleaved caspase 3 Decreased: MRP1, LRP, Bcl2, p-AKT | [163] | |
| Wogonin | 20–80 μmol/L | Human colon cancer under hypoxic conditions | HCT116 | Decreased: HIF-1α, HKII, PDHK1, LDHA, p-PI3K, p-AKT | [164] | |
| Tanshinone IIA | 20 µg/mL | Doxorubicin-resistant human breast cancer | MCF-7/DOX | Decreased: P-gP, BCRP, MRP1 | [165] | |
| Resveratrol | 45.9 µmol/L | Adriamycin-resistant human leukemia | K562/ADR | Increased: caspase 3 and caspase 8 Decreased: P-gP, MDR1, p-AKT | [166] | |
| Curcumin | 0–45 µmol/L | Adriamycin-resistant mouse leukemia cell | L210/ADR | Decreased: P-gP, p-AKT at Ser-473, p-GSK-3b at Ser-9 | [167] | |
| Nobiletin | 31.62 µmol/L | Paclitaxel-resistant ovarian cancer | A2780/T | Increased: P53 Decreased: AKT, p-AKT, p-ERK1/2, Nrf2 | [168] | |
| 50 µmol/L | Adriamycin-resistant human non-small-cell lung cancer | A549/ADR | Decreased: MRP1, MYCN, β-catenin, p-GSK-3β, Bcl-2, survivin | [169] | ||
| Quinones | Emodin | 9 μmol/L | 5-fuorouracil-resistant human colorectal carcinoma | SW480/5-Fu | Increased: cleaved caspase 3, Bax Decreased: Bcl-2, p-ERK, p-AKT | [170] |
| Levoshikonin | 3.92 μmol/L | Cisplatin-resistant cervical cancer | Hela/DDP | Increased: Bax, Cleaved caspase 3 Decreased: Bcl-2 | [171] | |
| 1,4-Naphthoquinone | 0.1 µmol/L | Vincristine-resistant chronic myeloid leukemia Daunorubicin-resistant chronic myeloid leukemia | K-562-Lucena1 FEPS | Increased: H2AFX, ABCB1 | [172] | |
| Terpenoids | Triptolide | 10–160 nmol/L | Cisplatin-resistant cervical cancer | Hela/DDP | Decreased: Bcl-2, XIAP | [173] |
| Cucurbitacin B | 15 nmol/L | Gefitinib-resistant lung cancer | A549-GR | Increased: E-cadherin, ROS Decreased: EGFR, N-cadherin, Vimentin, p-PI3K, p-AKT, p-mTOR | [174] | |
| β-elemene | 20, 40 µg/mL | Cisplatin-resistant human lung cancer | A549/DDP | Decreased: MDR1, LRP, p-PI3K, p-AKT | [175] | |
| 5–100 µg/mL | Adriamycin-resistant gastric cancer | SGC7901/Adr, Xenografts of SGC7901/Adr in BALB/c nude mice | Increased: caspase 3 | [182] | ||
| Flavonoids | Quercetin | 0.005–0.15 µg/mL | Cisplatin-resistant cervical cancer | SiHa/DDP | Decreased: AKT, mTOR, p70S6K, P-gP | [176] |
| Luteolin | 10–200 mg/L | Doxorubicin-resistant cervical cancer | HeLa/DOX | Increased: PI3K, Cleaved caspase 3 Decreased: p-AKT, p-mTOR, p70S6K, Ki67 | [177] | |
| Alkaloids | Matrine | 0.15–1.2 mg/mL | Adriamycin-resistant human breast cancer | MCF-7/ADR | Increased: PTEN, Bax, cleaved caspase 3 Decrease: MDR1, MRP1, p-AKT, Bcl-2 | [178] |
| Furanocoumarin | Imperatorin | 2 µmol/L | Cisplatin-resistant cervical cancer | Hela/R | Increased: Bim, Bak, Bax, release of cytochrome c, activation of caspase 3 | [179] |
| Compound | Metformin | 0.25–8 mmol/L | Cisplatin-resistant endometrial cancer | Ishikawa/DDP | Increased: Bax Decreased: Bcl-2, AKT1 | [180] |
| 10–80 mmol/L | Progestin-resistant endometrial cancer | MPA-R-Ishikawa | Decreased: proliferation | [181] | ||
| Sijunzi Tang | 4.806 mg/mL | Gefitinib-resistant non-small cell lung cancer | PG9-GR, Xenografts of PC9/GR in BALB/c nu/nu mice | Increased: Bax, Decreased: Bcl-2 | [183] | |
| Compound Zhebei | 2.5–10 g/kg | Oxaliplatin-resistant colon cancer | Xenografts of HCT-116/L-OHP in BALB/c nude mice | Increased: Bax, Decreased: Bcl-2 | [184] | |
| Compound Zhebei | 2.5–10 g/kg | Cisplatin-resistant acute lymphoblastic leukemia | Xenografts of L1210 in DBA/2 nude mice | Increased: Bax, Decreased: Bcl-2 | [185] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line and In Vivo Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Flavonoids | Isoliquiritigenin | 0–100 µmol/L | Doxorubicin resistant human uterine sarcoma | MES-SA/Dx5 MES-SA/Dx5-R | Increased: cleaved PARP, cleaved caspase 7, LC3BI, LC3BII Decreased: PARP, Bcl-2, caspase 7, p-mTOR, SQSTM1/p62 | [188] |
| Baicalein | 0–10 µg/mL | 5-Fluorouracil resistant hepatocellular carcinoma | Bel7402/5-Fu | Decreased: P-gP, Bcl-xL | [189] | |
| Baicalin | 20–40 µmol/L | Cisplatin resistant cervical cancer | C-33A/Cis | Increased: ratio of MAPLC3II/MAPLC3I, Beclin-1, Atg5, Atg12 | [191] | |
| Puerarin | 25 µmol/L | Adriamycin resistant human leukemia | K562/ADR | Increased: LC3-II, Beclin-1, caspase 3, accumulation of adriamycin Decreased: MDR1, NF-κB activity, cyclingB1, p-AKT, p-JNK | [193] | |
| Hesperetin | 50–200 µmol/L | Cisplatin resistant ovarian adenocarcinoma | A2780/DDP | Increased: p62, p53, Bax, Caspase 3 Decreased: LC3, Beclin1, AMPK, mTOR, Bcl-2 | [194] | |
| Alkaloids | Matrine | 0.2 mg/mL | Adriamycin resistant human leukemia | K562/ADM | Increased: LC3II Decreased: p62 | [195] |
| Lycorine | 6 µmol/L | Imatinib resistant human leukemia | K562/IM | Increased: p62 Decreased: Beclin-1, Atg5, LC3-II, P-gP | [196] | |
| Cepharanthine | 8 µmol/L | Icotinib resistant non-small cell lung cancer | PC-9/IR | Increased: p53, Beclin-1, ratio of LC3-II/LC3-I Decreased: ratio of p-mTOR/mTOR | [197] | |
| Berberine | 100 µmol/L | Doxorubicin resistant breast cancer | MCF-7/ADR, Xenografts of MCF-7/ADR in BALB/c nude mice | Increased: p62, Akt, mTOR; Decreased: MDR1, LC3-II/I, PTEN | [203] | |
| Terpenoids | β-elemene | 15 µg/mL | Cisplatin resistant human lung cancer | SPC-A-1/DDP | Increased: Beclin-1 Decreased: MRP-1, P-gP | [198] |
| 120 µg/mL | Gefitinib resistant non-small cell lung cancer | PC9GR HCC827GR | Increased: LC3B-II, SQSTM1 Decreased: METTL3 | [199] | ||
| Triptolide | 0–100 nmol/L | Cisplatin resistant ovarian cancer | SKOV3/DDP | Increased: E-cadherin, Beclin1, LC3, ratio of LC3II/LC3-I Decreased: N-cadherin, MMP9 | [200] | |
| Polyphenols | Naringenin | 15–240 mg/L | Cisplatin resistant cervical cancer | Hela/DDP | Increased: ratio of p-AMPK/AMPK, Bax, Beclin1, LC3II Decreased: LC3I | [201] |
| Compound | Extract of Scutellaria baicalensis | 0–400 µg/mL | Cisplatin resistant Ovarian cancer | A2780Cis | Increased: Atg12 Decreased: p53, DRAM, Beclin 1, Atg5 | [190] |
| Kang’ai injection | 100.07 g/L | Cisplatin resistant Human Lung Cancer | A549/DDP | Increased: Atg7, LC3I, LC3II, Beclin 1, ratio of Bax/Bcl-2, cleaved caspase 3 | [202] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line and In Vivo Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Polyphenols | Hesperetin | 0.6–160 µmol/L | Cisplatin resistant human lung cancer | A549/DDP | Decreased: P-gP, ratio of p-P65/t-P65, ratio of P-IκB/IκB | [205] |
| Resveratrol | 100 µmol/L | Adriamycin resistant human Osteosarcoma | U2OS/ADR | Decreased: P-gP, acetylated p65, p-p38 | [216] | |
| Rosmarinic acid | 46.47 µg/mL | Cisplatin resistant non-small cell lung cancer | A549/DDP | Increased: cleaved caspase 3, Bax, p53, p21, p-C-Jun Decreased: Bcl-2, caspase 3, P-gP | [217] | |
| Baicalin | 8 µg/mL | Cisplatin resistant human lung cancer | A549/DDP | Decreased: MARK2, p-AKT | [218] | |
| Curcumin | 20 μmol/L | Vincristine resistant esophageal carcinoma | Eca-109/VCR | Decreased: p38MAPK, p-p38MAPK, ERCC1, P-gp | [219] | |
| 0–60 µg/mL | Adriamycin resistant hepatocarcinoma | HepG2/ADM | Decreased: p38MAPK | [220] | ||
| Paeonol | BGC-823/AP: 53.64 mg/L MGC-803/AP: 56.83 mg/L | Apatinib resistant gastric cancer | BGC-823/AP MGC-803/AP | Decreased: HK2, GLUT1, LDHB, MAPK1 | [221] | |
| Lignans | Schisandrin A | 20 µmol/L | Doxorubicin resistant human breast cancer | MCF-7/DOX | Increased: Cleaved caspase 9 PARP Decreased: P-gP | [209] |
| Lignans | Muscone | 0–16 µmol/L | cisplatin resistant lung cancer | A549/DDP, Xenografts of A549/DDP in BALB/c-nu mice | Decreased: MRP1, P-gP, p50, p52, p65 | [225] |
| Alkaloids | Matrine | 0–300 µmol/L | Adriamycin resistant human leukemia | K562/ADR | Increased: cleaved caspase 9 Decreased: ABCB1, ABCC1, ABCG2, phosphorylation of NF-κB, survivin, Bcl-xL | [206] |
| Tetrandrine | 0–8 μmol/L | Vincristine resistant gastric cancer | SGC-7901/VCR | Increased: cleaved-caspase 9, cleaved-caspase 3, p-p38, p-JNK, activation of PARP Decreased: Bcl2, p-ERK, p-CDC2 | [224] | |
| Terpenoids | Furanodiene | 0.200 µmol/L | Doxorubicin resistant breast cancer | MCF-7/DOX | Increased: Bad, caspase 3/7/8, PARP, p65, IKKα/β Decreased: Bcl-xl | [207] |
| Triptolide | 0.03–3 µmol/L | Taxol resistant human lung adenocarcinoma | A549/Taxol | Decreased: NF-κB, P62, FLIP, XIAP, Bcl-2, Bcl-xL, COX-2 | [208] | |
| Rotundioic acid | 0–4 µg/mL | Adriamycin resistant human leukemia | K562/ADM | Increased: p-p38,p-ERK1/2 Decreased: MDR1, P-gP | [222] | |
| Isolinderalactone | HCT116-OxR: 5.13 μmol/L HT29-OxR: 9.46 μmol/L | Oxaliplatin resistant colorectal cancer | HCT116-OxR HT29-OxR | Increased: p21, p27, GRP78, CHOP, DR4, DR5, p-JNK, p-p38, Bim, Bax Decreased: cyclinB1, CDC2, Bid, Bcl-2, Bcl-xL | [223] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Quinones | Shikonin | 4 μmol/L | Cisplatin-resistant ovarian cancer | SKOV3/DDP | Increased: P18, Bax, cleaved caspase 3 Decreased: cyclin D1, CDK2, ratio of p-Rb/Rb, Bcl-2 | [230] |
| Plumbagin | 90.8 μmol/L | Cisplatin-resistant human cervix squamous carcinoma | A431/Pt | Increased: ROS, p-Rb, sub G0 phase | [231] | |
| Alkaloids | Tetramethypyrazine | 73 µmol/L | Multidrug-resistant human bladder cancer | Pumc-91/ADM T24/DDP | Increased: Topo-II, S phase Decreased: MRP1, GST, BCL-2 | [232] |
| Homoharringtonine | 18.57 ng/mL | Vemurafenib-resistant melanoma | A375R | Increased: G0/G1 phase Decreased: Cyclin E1, CDK2, IRS4, PI3K, p-Akt, p-ERK | [233] | |
| Terpenoids | Triptolide succinate monoester | 33.32 µmol/L | Cisplatin-resistant human lung adenocarcinoma | A549/DDP | Increased: G2/M, apoptosis | [234] |
| Kamebakaurin | 62.97 μmol/L | Adriamycin-resistant hepatocellular carcinoma | HepG2/ADM | Increased: Bax, p-PTEN, G2 phase Decreased: Bcl-2, MDR1, p-AKT | [235] | |
| Saponins | Ginsenoside Rh2 | 20 µmol/L | 5-fuorouracil-resistant human colorectal carcinoma | HCT-8/5-Fu LoVo/5-Fu | Increased: cleaved-caspase 3, p-IκB, E-cadherin, G1 phase Decreased: cyclin D1, CDK2, p-Rb, Bcl-2, N-cadherin, Vimentin, MMP9, MRP1, MDR1, LRP, GST | [236] |
| Polyphenols | Curcumin | 13.69 μmol/L | NNMT-related resistant colorectal cancer | SW480/NNMT | Increased: p-Rb, G2/M Phase, ROS Decreased: p-STAT3, CDK1, CDK2 | [237] |
| Flavonoids | Isoxanthohumol | 42.1 μmol/L | Cisplatin-resistant human lung adenocarcinoma | A549/DDP | Increased: G0/G1 phase Decreased: P-gP, LRP, MRP, PI3K, p-AKt | [238] |
| Compound | Metformin | 28. 02 μg/ml | Oxaliplatin-resistant human gastric cancer | SGC-7901/L-OHP | Decreased: cyclin D1 | [239] |
| Buzhong yiqi decoction | 0.25 g/mL | Cisplatin-resistant human gastric cancer | SGC-7901/CDDP | Increased: G1/S phase Decreased: survivin, Bcl-2, NF-κB | [240] | |
| Digitoxin | 132.65 nmol/L | Adriamycin-resistant human liver cancer | HepG2/ADM | Increased: cyclin B1, numbers of punctuate γH2AX foci, p-CHK2, p-ATR(Ser428), Bax, G2/M phase Decreased: p-CDK1 (Thr14), CDC25C, Bcl-2 | [241] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Flavonoids | Luteolin | 20 μmol/L | Paclitaxel-resistant esophageal cancer | TE-1/PTX EC109/PTX | Increased: E-cadherin Decreased: Slug, N-cadherin, Vimentin | [243] |
| 0–40 μmol/L | Paclitaxel-resistant esophageal cancer | TE-1/PTX EC109/PTX | Increased: p-PI3K, p-AKT, E-cadherin Decreased: SOX2, OCT4, NANOG | [244] | ||
| Quercetin | 0.7 µmol/L | Doxorubicin-resistant human breast cancer | MCF-7/dox | Increased: PTEN Decreased: CD44+/CD24−/low, p-AKT | [245] | |
| Genistein | 10 μmol/L | Multidrug-resistant liver cancer | SMMC-7721 | Decreased: CD133, CD44, Gli1, ABCG2 | [254] | |
| Alkaloids | Matrine | 10 mg/mL | 5-Fuorouracil-resistant colon cancer stem cells | HCT116/5-Fu | Increased: apoptosis Decreased: ABCB1, p-PI3K, p-Akt, p-mTOR, P-gP | [246] |
| Neferine/isoliensinine | Neferine: 6.5 μmol/L Isoliensinine: 12.5 μmol/L | Cisplatin-resistant colon cancer stem cells | HCT-15/DDP-CSCs | Increased: ROS, Bax, cytochrome c, caspase 3, PARP, sub G0 phase Decreased: PI3K, p-AKT, mTOR, BCl-2 | [247] | |
| Terpenoids | Ursolic acid | 9–32 μg/g | Drug resistance in nude mice bearing ovarian cancer stem cells | Skov3-sp | Decreased: ABCG2 | [248] |
| Norcantharidin | A2780Cis: 1.46 μg/mL SKOV3Pac: 1.8 μg/ml | Drug-resistant ovarian cancer | A2780Cis SKOV3Pac | Increased: Ptch1 Decreased: Sox2, Nanog, Bmi1, Shh | [255] | |
| Polyphenols | Curcumin | 10, 20 μmol/L | Drug-resistant colon cancer | HCT116/OX HCT8/5-Fu | Decreased: CD133, ALDH1, CD24 | [249] |
| 10 μmol/L | 5-Fuorouracil-resistant gastric cancer | BGC-823/5-Fu | Increased: E-cadherin, GSK-3β Decreased: Wnt2, β-catenin, vimentin | [257] | ||
| Compound | Yi-qi-hua-yu-jie-du decoction | 2.5 g/mL | 5-Fuorouracil-resistant gastric cancer | BGC823/5-Fu | Decreased: P-gP, MRP1, Nrf2, p-PI3K, p-AKT | [250] |
| Chemical Family | Molecule Name | Concentration | Resistant Cancer | Cell Line and In Vivo Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Polyphenols | Baicalin | 0–100 µmol/L | Human erythroleukemia | K562 | Increased: cleaved Notch1 | [258] |
| Alkaloids | Sinomenine | 40 µmol/L | Cisplatin-resistant cervical cancer | Hela/DDP | Decreased: Notch1, STAT3, Hes1 | [259] |
| Matrine | 1–32 μmol/L | Cisplatin-resistant lung cancer | A549/DDP | Increased: E-cadherin Decreased: Vimentin, Slug, p-p65 | [265] | |
| Berberine | 25 µmol/L | EGFR-TKI-resistant lung cancer | A549,H1975 | Increased: E-cadherin, Decreased: vimentin, Snail | [272] | |
| Flavonoids | Quercetin | 128 μmol/L | Gemcitabine-resistant non-small-cell lung cancer | A549/Gb | Increased: E-cadherin, miR-101 Decreased: N-cadherin, Vimentin, EZH2, TGF-β1, ratio of p-SMAD4/SMAD4 | [263] |
| Polysaccharide | Astragalus polysaccharides | 0.3 g/kg/d | Cisplatin-resistant lung adenocarcinoma BALB/c nude mice | A549/DDP | Increased: α-catenin Decreased: quality of tumor, N-cadherin, LRP, MRP, P-gP | [264] |
| Terpenoids | Celastrol | 0–4 μmol/L | Cisplatin-resistant nasopharyngeal carcinoma | Cis-039 Cis-BM | Increased: cleaved caspase 3, cleaved caspase 8, cleaved caspase 9, PARP, Bax, Bim S, p-p38, p-ERK1/2, p-JNK1/2, Decreased: Bcl-xL, Bcl-2 | [266] |
| Oridonin | 5 μmol/L | Fulvestran-resistant breast cancer | MCF-7/Ful | Increased: γ-H2AX, Bax, caspase 3, caspase 9, ratio of p-CDC2/CDC2, ratio of LC3-II/I, G0/G1 phase Decreased: Bcl-2, cyclin D1 | [267] | |
| Terpenoids | β-Elemene | 20 μg/mL | Cisplatin-resistant human lung adenocarcinoma | A549/DDP | Increased: apoptosis, ROS, released of GSH, accumulation of intracellular Rh-123 Decreased: the mitochondrial membrane potential (ΔΨm), P-gP | [268] |
| 108.5 μg/mL | Paclitaxel-resistant lung cancer | A549/Taxol | Increased: Bax, caspase 3 Decreased: JAK2, STAT3, p-STAT3, Bcl-2 | [269] | ||
| 1.5 µg/mL | Adriamycin-resistant gastric cancer | SGC7901/ADR, Xenografts of SGC7901/Adr in BALB/c-nu/nu mice | Increased: E-cadherin. Cb1-b, Decreased: Vimentin, MMP2, MMP9, ZEB1, ZEB2, EGFR, p-EGFR, p-AKT, p-ERK | [273] | ||
| Quinones | Shikonin | 9 µmol/L | Cisplatin-resistant human ovarian cancer | A2780-CR | Increased: Bax, p-ERK1/2, ERK2, p-JNK1/2, JNK1/2, p-p38, p38, E-cadherin Decreased: Bcl-2, N-cadherin | [270] |
| 0.4 µmol/L | Cisplatin-resistant bladder cancer | T24R | Increased: p-RIP3, PUMA, Bax Decreased: Bcl-2 | [271] | ||
| Compound | Buzhong Yiqi Decoction | 1.46–5.82 g/kg | Cisplatin-resistant lung cancer | A549/DDP, Xenografts of A549/DDP in BALB/c nu/nu mice | Decreased: β-catenin | [274] |
| Jiedu Fuzheng prescription | 52, 104 g/kg | Gefitinib-resistant non-small cell lung cancer | Xenografts of PC9/GR in BALB/c nu/nu mice | Decreased: p-ERK/ERK, Nrf2 | [275] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Wang, Y.; Xia, L.; Li, J. Research Progress of Plant-Derived Natural Products against Drug-Resistant Cancer. Nutrients 2024, 16, 797. https://doi.org/10.3390/nu16060797
Liu W, Wang Y, Xia L, Li J. Research Progress of Plant-Derived Natural Products against Drug-Resistant Cancer. Nutrients. 2024; 16(6):797. https://doi.org/10.3390/nu16060797
Chicago/Turabian StyleLiu, Wenli, Yuqin Wang, Lijie Xia, and Jinyao Li. 2024. "Research Progress of Plant-Derived Natural Products against Drug-Resistant Cancer" Nutrients 16, no. 6: 797. https://doi.org/10.3390/nu16060797
APA StyleLiu, W., Wang, Y., Xia, L., & Li, J. (2024). Research Progress of Plant-Derived Natural Products against Drug-Resistant Cancer. Nutrients, 16(6), 797. https://doi.org/10.3390/nu16060797