
High Dietary Intake of Iron Might Be Harmful to Atrial Fibrillation and Modified by Genetic Diversity: A Prospective Cohort Study

 ,
, 

Abstract
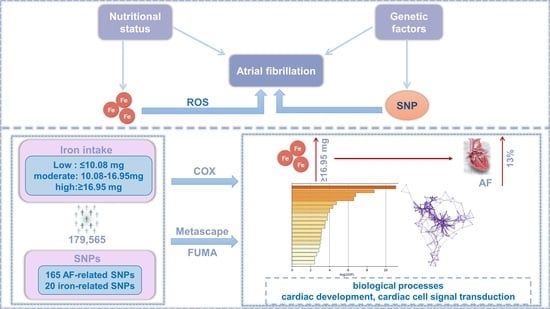
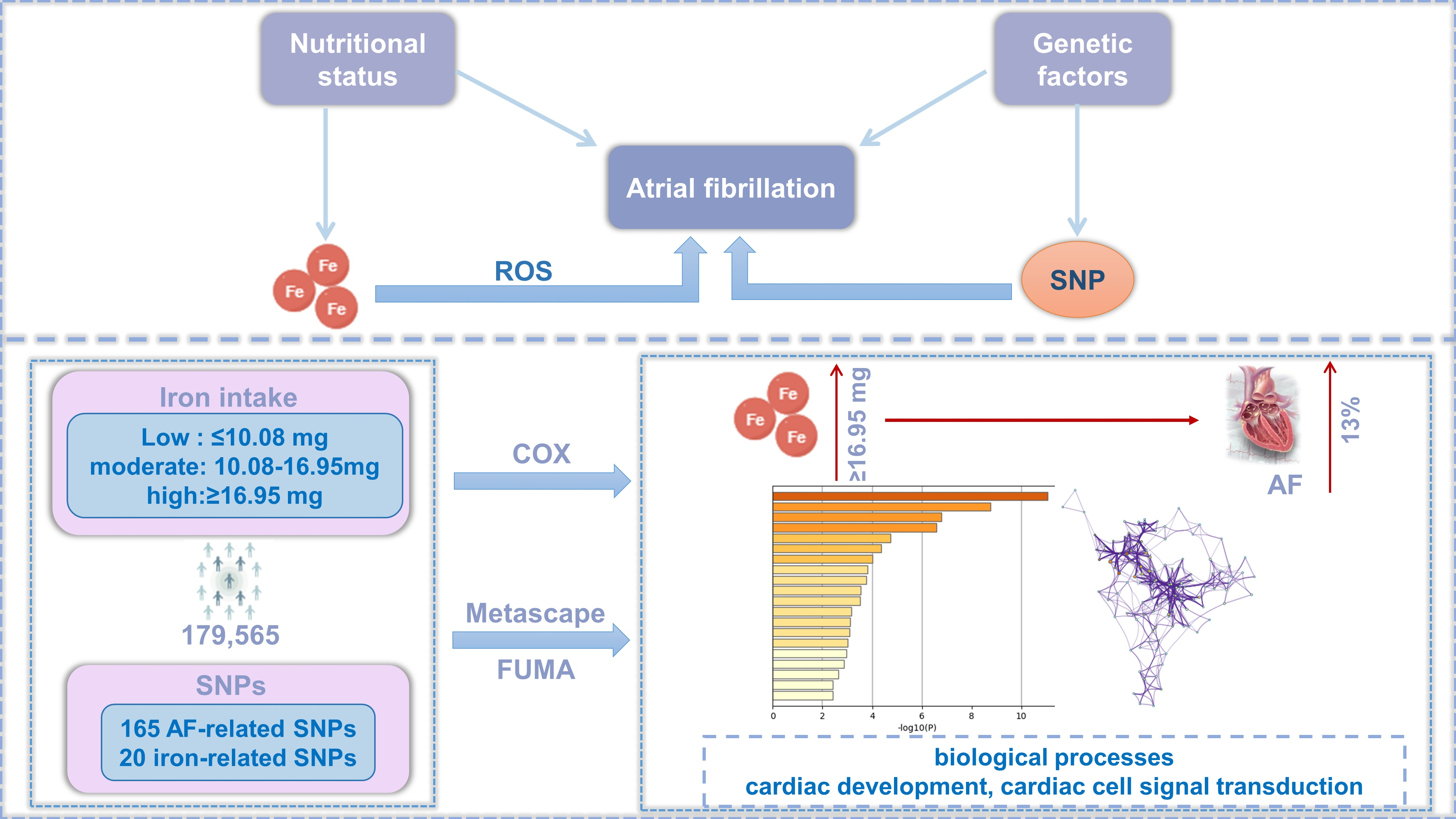
1. Introduction
2. Materials and Methods
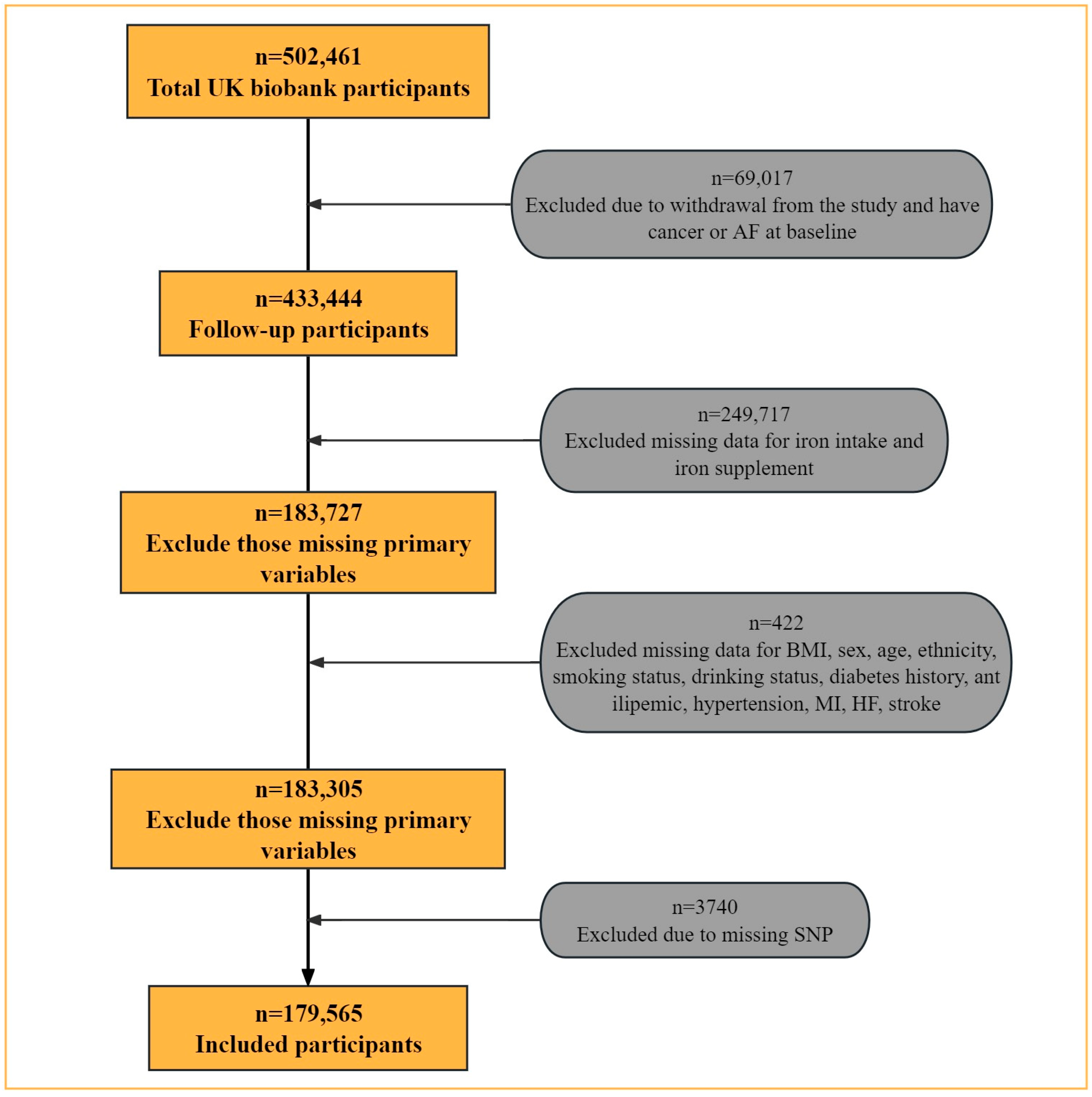
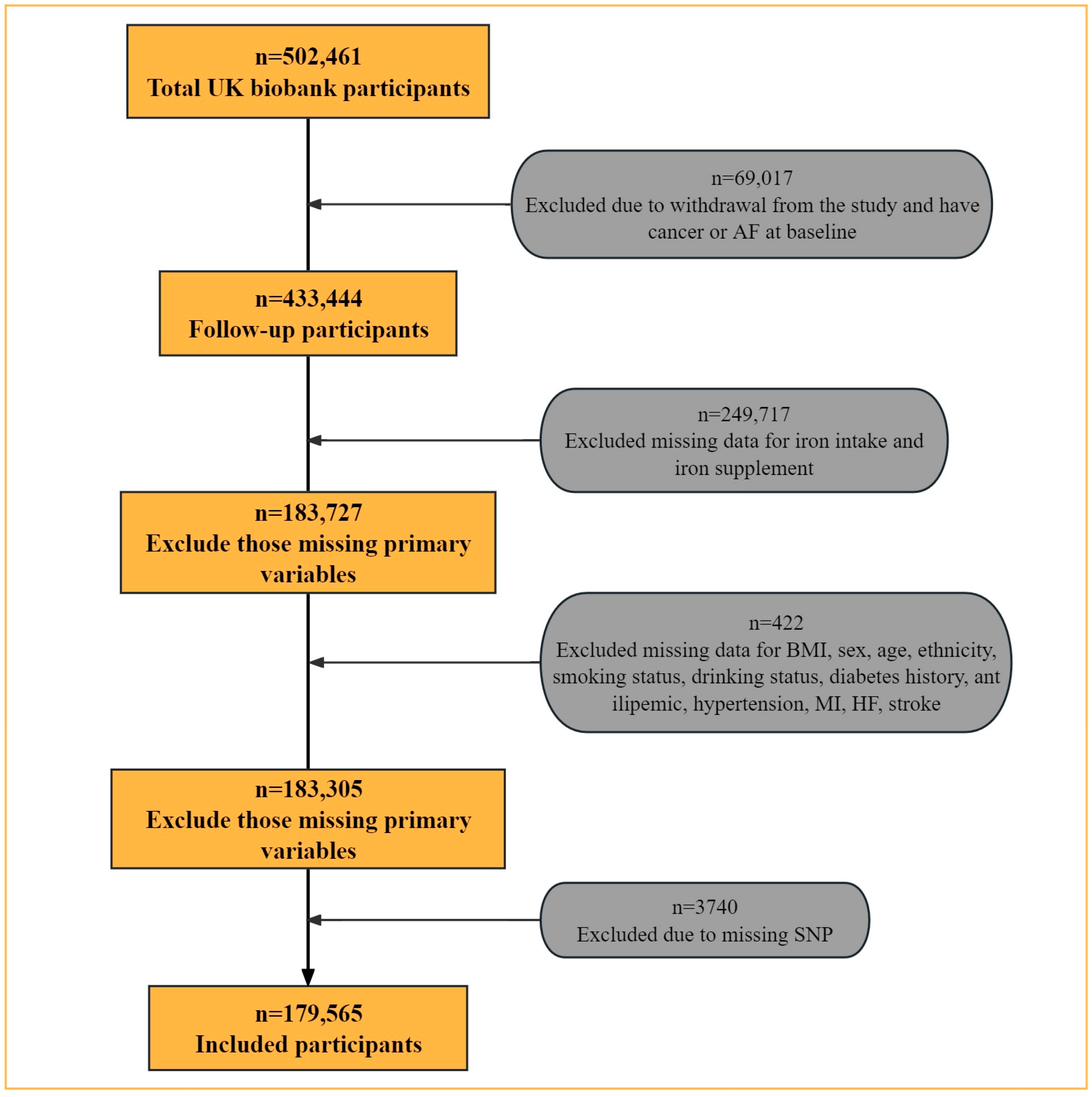
2.1. Study Design and Data Collection
2.2. Dietary Iron Intake Assessment
2.3. Follow-Up and Outcomes
2.4. Single Nucleotide Polymorphism Selection and Genotyping
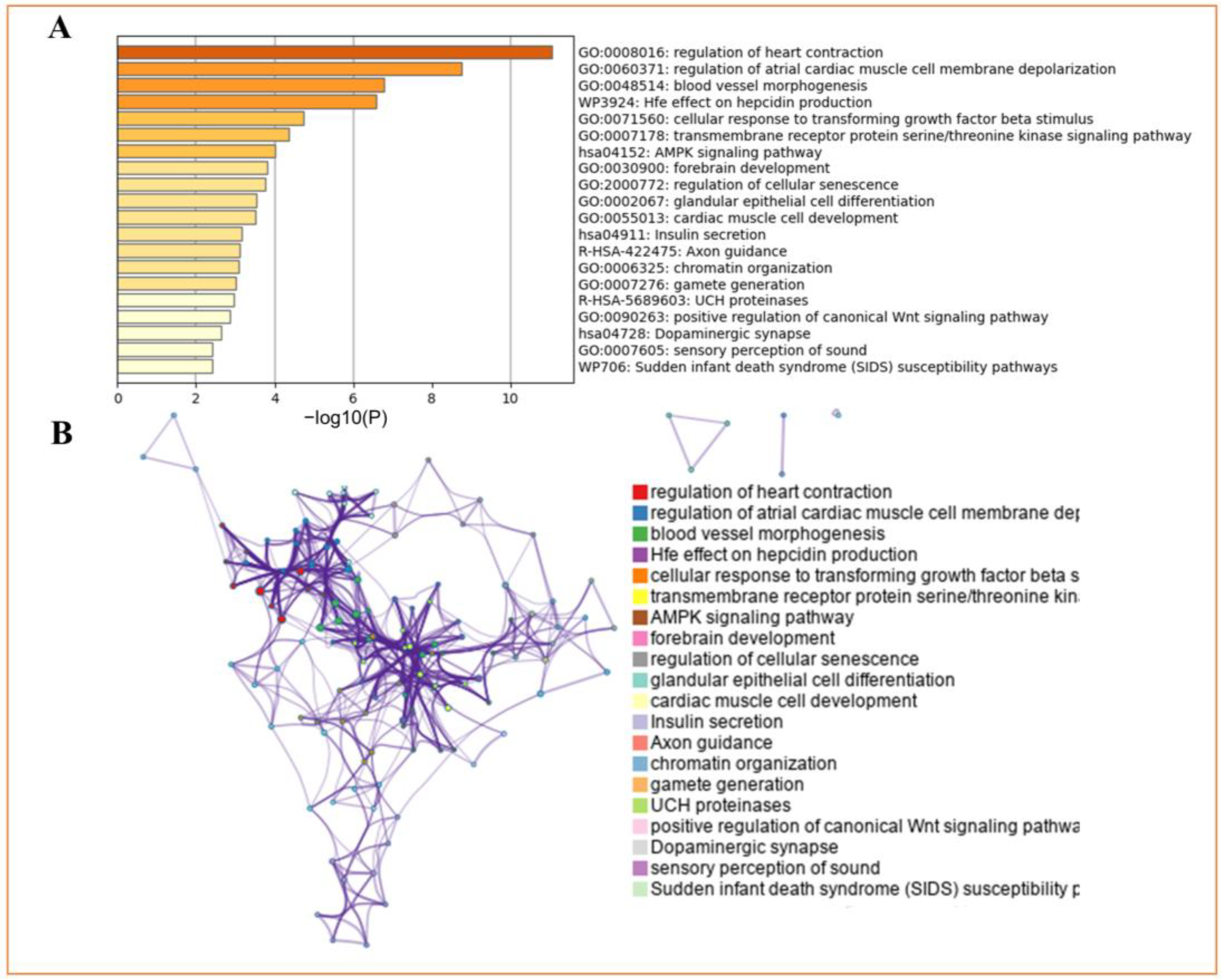
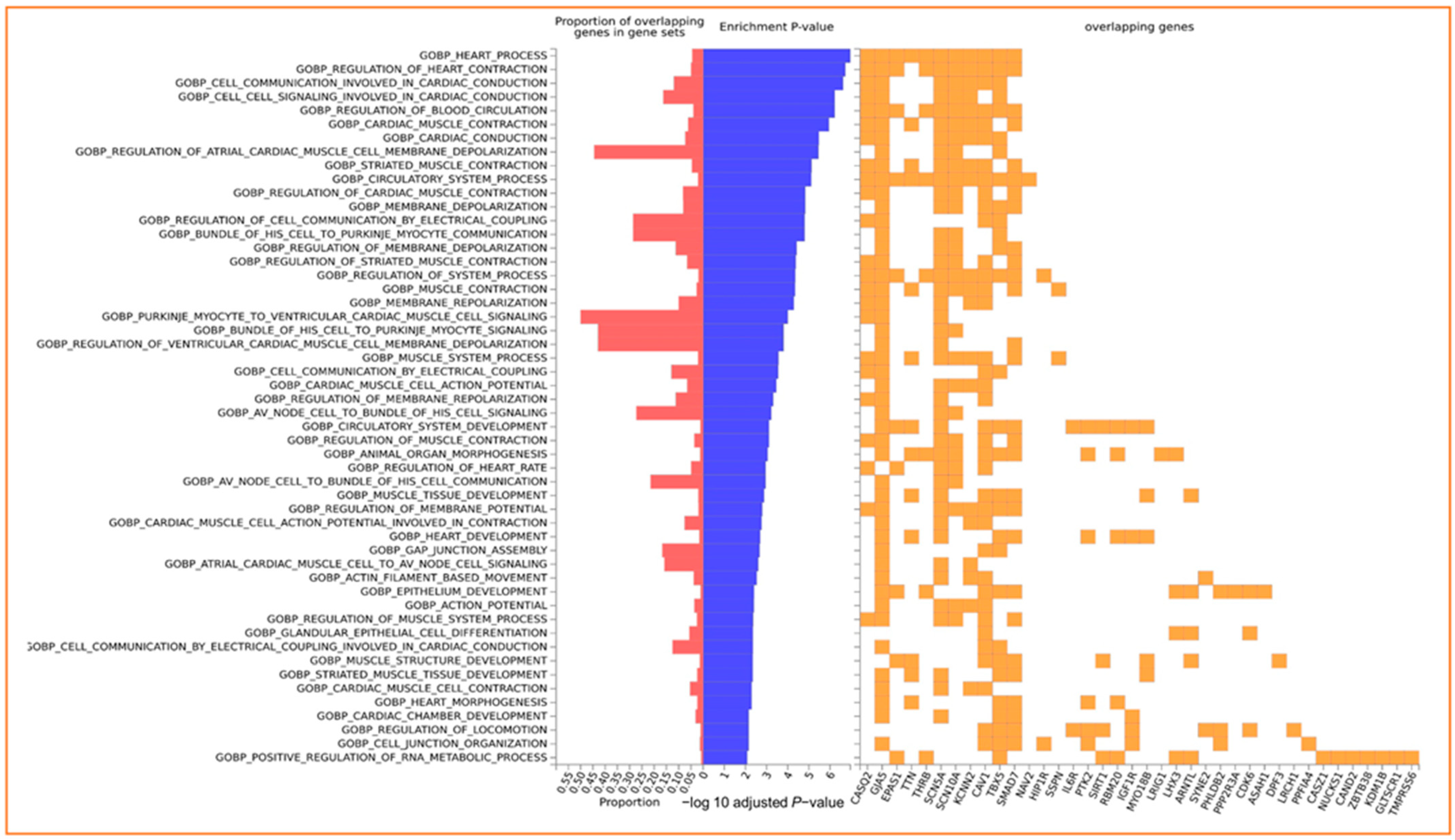
2.5. GO and Pathway Enrichment Analysis
2.6. Covariates
2.7. Statistical Analysis
2.8. Subgroup Analysis
3. Results
3.1. Baseline Characteristics
3.2. Dietary Iron Intake and the Risk of Incident Atrial Fibrillation
3.3. Incident Atrial Fibrillation according to Different SNPs and Dietary Iron Intake
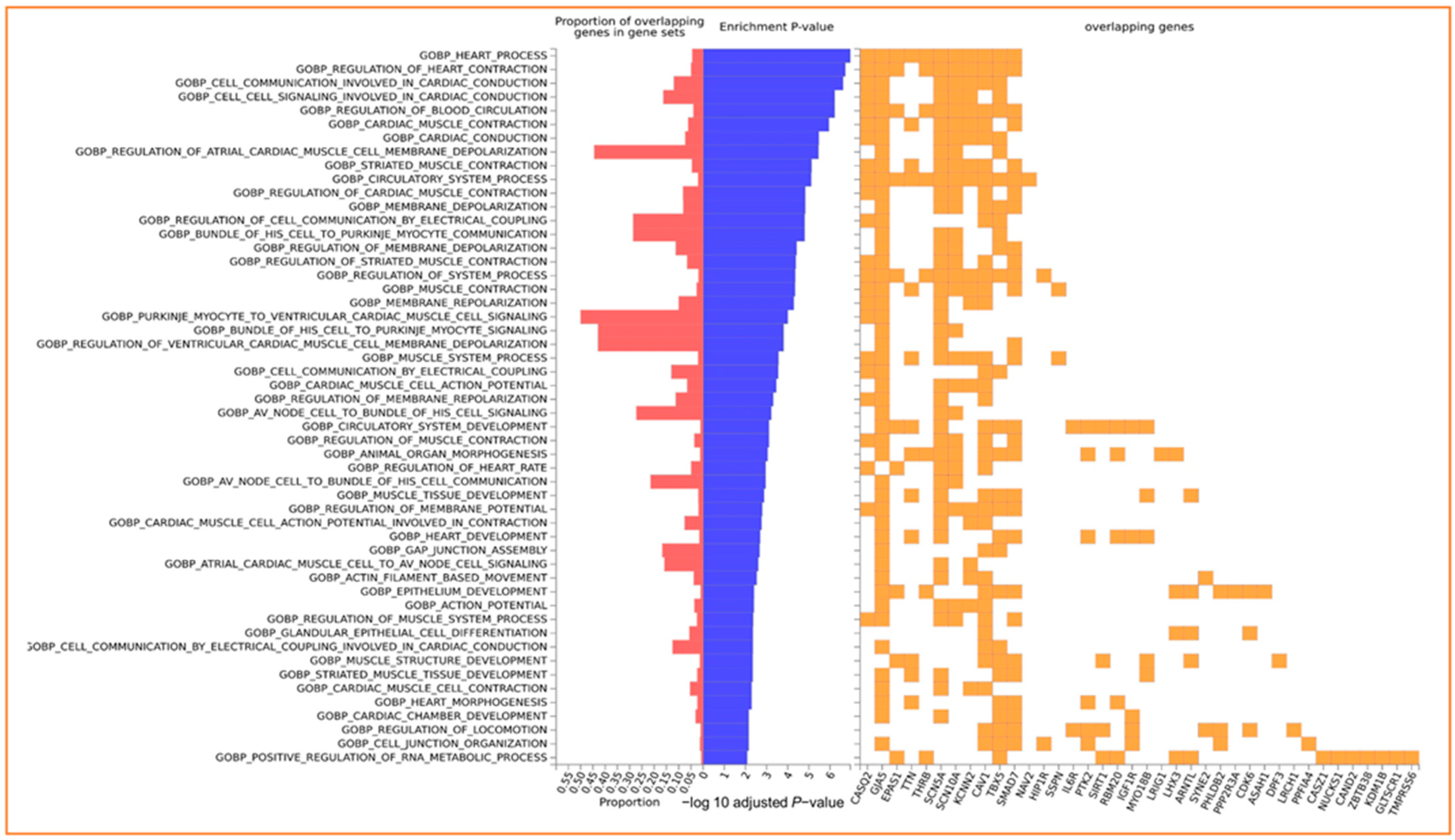
3.4. Enrichment of Input Genes in Gene Sets
3.5. Subgroup Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kornej, J.; Borschel, C.S.; Benjamin, E.J.; Schnabel, R.B. Epidemiology of Atrial Fibrillation in the 21st Century: Novel Methods and New Insights. Circ. Res. 2020, 127, 4–20. [Google Scholar] [CrossRef]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Al-Mubarak, A.A.; Grote Beverborg, N.; Zwartkruis, V.; van Deutekom, C.; de Borst, M.H.; Gansevoort, R.T.; Bakker, S.J.L.; Touw, D.J.; de Boer, R.A.; van der Meer, P.; et al. Micronutrient deficiencies and new-onset atrial fibrillation in a community-based cohort: Data from PREVEND. Clin. Res. Cardiol. 2023, 43, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cheng, J.; Wang, Y. Genetic support of a causal relationship between iron status and atrial fibrillation: A Mendelian randomization study. Genes Nutr. 2022, 17, 8. [Google Scholar] [CrossRef]
- Gujja, P.; Rosing, D.R.; Tripodi, D.J.; Shizukuda, Y. Iron overload cardiomyopathy: Better understanding of an increasing disorder. J. Am. Coll. Cardiol. 2010, 56, 1001–1012. [Google Scholar] [CrossRef]
- Roselli, C.; Rienstra, M.; Ellinor, P.T. Genetics of Atrial Fibrillation in 2020. Circ. Res. 2020, 127, 21–33. [Google Scholar] [CrossRef]
- Patel, K.K.; Venkatesan, C.; Abdelhalim, H.; Zeeshan, S.; Arima, Y.; Linna-Kuosmanen, S.; Ahmed, Z. Genomic approaches to identify and investigate genes associated with atrial fibrillation and heart failure susceptibility. Hum. Genom. 2023, 17, 47. [Google Scholar] [CrossRef] [PubMed]
- Galesloot, T.E.; Geurts-Moespot, A.J.; den Heijer, M.; Sweep, F.C.; Fleming, R.E.; Kiemeney, L.A.; Vermeulen, S.H.; Swinkels, D.W. Associations of common variants in HFE and TMPRSS6 with iron parameters are independent of serum hepcidin in a general population: A replication study. J. Med. Genet. 2013, 50, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.B.; Thorolfsdottir, R.B.; Fritsche, L.G.; Zhou, W.; Skov, M.W.; Graham, S.E.; Herron, T.J.; McCarthy, S.; Schmidt, E.M.; Sveinbjornsson, G.; et al. Biobank-driven genomic discovery yields new insight into atrial fibrillation biology. Nat. Genet. 2018, 50, 1234–1239. [Google Scholar] [CrossRef]
- Jallow, M.W.; Campino, S.; Prentice, A.M.; Cerami, C. Association of common TMPRSS6 and TF gene variants with hepcidin and iron status in healthy rural Gambians. Sci. Rep. 2021, 11, 8075. [Google Scholar] [CrossRef]
- Benyamin, B.; Ferreira, M.A.; Willemsen, G.; Gordon, S.; Middelberg, R.P.; McEvoy, B.P.; Hottenga, J.J.; Henders, A.K.; Campbell, M.J.; Wallace, L.; et al. Common variants in TMPRSS6 are associated with iron status and erythrocyte volume. Nat. Genet. 2009, 41, 1173–1175. [Google Scholar] [CrossRef]
- Galesloot, T.E.; Janss, L.L.; Burgess, S.; Kiemeney, L.A.L.M.; Den Heijer, M.; De Graaf, J.; Holewijn, S.; Benyamin, B.; Whitfield, J.B.; Swinkels, D.W.; et al. Iron and hepcidin as risk factors in atherosclerosis: What do the genes say? BMC Genet. 2015, 16, 79. [Google Scholar] [CrossRef]
- De Tayrac, M.; Roth, M.-P.; Jouanolle, A.-M.; Coppin, H.; Le Gac, G.; Piperno, A.; Férec, C.; Pelucchi, S.; Scotet, V.; Bardou-Jacquet, E.; et al. Genome-wide association study identifies TF as a significant modifier gene of iron metabolism in HFE hemochromatosis. J. Hepatol. 2015, 62, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Flower, R.; Hyland, C.; Saiepour, N.; Faddy, H. Genetic factors associated with iron storage in Australian blood donors. Blood Transfus. 2018, 16, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ye, Y.; Tu, H.; Hildebrandt, M.A.; Zhao, L.; Heymach, J.V.; Roth, J.A.; Wu, X. MicroRNA-related genetic variants in iron regulatory genes, dietary iron intake, microRNAs and lung cancer risk. Ann. Oncol. 2017, 28, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.; Del Greco, M.F.; Walker, A.P.; Srai, S.K.S.; Laffan, M.A.; Minelli, C. The Effect of Iron Status on Risk of Coronary Artery Disease: A Mendelian Randomization Study-Brief Report. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1788–1792. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.J. UK Biobank: Bank on it. Lancet 2007, 369, 1980–1982. [Google Scholar] [CrossRef] [PubMed]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M.; et al. UK Biobank: An Open Access Resource for Identifying the Causes of a Wide Range of Complex Diseases of Middle and Old Age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef]
- Collins, R. UK Biobank: Protocol for a Large-Scale Prospective Epidemiological Resource. UK Biobank. 2007. Available online: https://policycommons.net/artifacts/1774896/uk-biobank/2506542/ (accessed on 1 October 2023).
- Perez-Cornago, A.; Pollard, Z.; Young, H.; Van Uden, M.; Andrews, C.; Piernas, C.; Key, T.J.; Mulligan, A.; Lentjes, M. Description of the updated nutrition calculation of the Oxford WebQ questionnaire and comparison with the previous version among 207,144 participants in UK Biobank. Eur. J. Nutr. 2021, 60, 4019–4030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-B.; Chen, C.; Pan, X.-F.; Guo, J.; Li, Y.; Franco, O.H.; Liu, G.; Pan, A. Associations of healthy lifestyle and socioeconomic status with mortality and incident cardiovascular disease: Two prospective cohort studies. BMJ 2021, 373, n604. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Watanabe, K.; Taskesen, E.; Van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [PubMed]
- Mayanja, R.; Machipisa, T.; Soremekun, O.; Kamiza, A.B.; Kintu, C.; Kalungi, A.; Kalyesubula, R.; Sande, O.J.; Jjingo, D.; Fabian, J.; et al. Genome-wide association analysis of cystatin-C kidney function in continental Africa. eBioMedicine 2023, 95, 104775. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, M.; Santema, B.T.; Roselli, C.; Nelson, C.P.; Koekemoer, A.; Romaine, S.P.R.; Van Gelder, I.C.; Lam, C.S.P.; Artola, V.A.; Lang, C.C.; et al. Genetic risk and atrial fibrillation in patients with heart failure. Eur. J. Heart Fail. 2020, 22, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.L.; Aribas, E.; Geurts, S.; van Lennep, J.E.R.; Ikram, M.A.; Bos, M.M.; de Groot, N.M.S.; Kavousi, M. Association Between Sex-Specific Risk Factors and Risk of New-Onset Atrial Fibrillation Among Women. JAMA Netw. Open 2022, 5, e2229716. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, Y.; Barnes, M.E.; Gersh, B.J.; Cha, S.S.; Bailey, K.R.; Abhayaratna, W.P.; Seward, J.B.; Tsang, T.S.M. Secular Trends in Incidence of Atrial Fibrillation in Olmsted County, Minnesota, 1980 to 2000, and Implications on the Projections for Future Prevalence. Circulation 2006, 114, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, L.F.; Nordestgaard, B.G.; Schnohr, P.; Ellervik, C. Increased Ferritin Concentration and Risk of Atrial Fibrillation and Heart Failure in Men and Women: Three Studies of the Danish General Population Including 35,799 Individuals. Clin. Chem. 2019, 65, 180–188. [Google Scholar] [CrossRef]
- Witkowska, A.M.; Waśkiewicz, A.; Zujko, M.E.; Szcześniewska, D.; Stepaniak, U.; Pająk, A.; Drygas, W. Are Total and Individual Dietary Lignans Related to Cardiovascular Disease and Its Risk Factors in Postmenopausal Women? A Nationwide Study. Nutrients 2018, 10, 865. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Kong, B.; Shuai, W.; Zhang, J.J.; Huang, H. Shensong Yangxin attenuates metabolic syndrome-induced atrial fibrillation via inhibition of ferroportin-mediated intracellular iron overload. Phytomedicine 2022, 101, 154086. [Google Scholar] [CrossRef]
- Anderson, G.J.; Frazer, D.M. Current understanding of iron homeostasis. Am. J. Clin. Nutr. 2017, 106, 1559S–1566S. [Google Scholar] [CrossRef]
- Benyamin, B.; Esko, T.; Ried, J.S.; Radhakrishnan, A.; Vermeulen, S.H.; Traglia, M.; Gögele, M.; Anderson, D.; Broer, L.; Podmore, C.; et al. Novel loci affecting iron homeostasis and their effects in individuals at risk for hemochromatosis. Nat. Commun. 2014, 5, 4926. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Roy, C.N.; Yao, W.; Matteini, A.; Semba, R.D.; Arking, D.; Walston, J.D.; Fried, L.P.; Singleton, A.; Guralnik, J.; et al. A genome-wide association analysis of serum iron concentrations. Blood 2010, 115, 94–96. [Google Scholar] [CrossRef]
- Nauffal, V.; Di Achille, P.; Klarqvist, M.D.R.; Cunningham, J.W.; Hill, M.C.; Pirruccello, J.P.; Weng, L.C.; Morrill, V.N.; Choi, S.H.; Khurshid, S.; et al. Genetics of myocardial interstitial fibrosis in the human heart and association with disease. Nat. Genet. 2023, 55, 777–786. [Google Scholar] [CrossRef] [PubMed]
- International Consortium for Blood Pressure Genome-Wide Association Studies; Ehret, G.B.; Munroe, P.B.; Rice, K.M.; Bochud, M.; Johnson, A.D.; Chasman, D.I.; Smith, A.V.; Tobin, M.D.; Verwoert, G.C.; et al. Genetic variants in novel pathways influence blood pressure and cardiovascular disease risk. Nature 2011, 478, 103–109. [Google Scholar] [CrossRef]
- Schroor, M.M.; Plat, J.; Mensink, R.P. Relation between single nucleotide polymorphisms in circadian clock relevant genes and cholesterol metabolism. Mol. Genet. Metab. 2023, 138, 107561. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Overall (n = 179,565) | Low (n = 35,967) | Moderate (n = 107,724) | High (n = 35,874) | p-Value |
|---|---|---|---|---|---|
| Atrial fibrillation (%) | |||||
| No | 172,872 (96.3) | 34,788 (96.7) | 103,781(96.3) | 34,303 (95.6) | <0.001 |
| Yes | 6693 (3.7) | 1179 (3.3) | 3943 (3.7) | 1571 (4.4) | |
| Sex (%) | |||||
| Female | 98,429 (54.8) | 23,019 (64.0) | 60,741 (56.4) | 14,669 (40.9) | <0.001 |
| Male | 81,136 (45.2) | 12,948 (36.0) | 46,983 (43.6) | 21,205 (59.1) | |
| Age (%) | |||||
| ≤65 years | 153,666 (85.6) | 31,579 (87.8) | 91,902 (85.3) | 30,185 (84.1) | <0.001 |
| >65 years | 25,899 (14.4) | 4388 (12.2) | 15,822 (14.7) | 5689 (15.9) | |
| Ethnicity (%) | |||||
| Nonwhite | 7958 (4.4) | 2885 (8.0) | 3754 (3.5) | 1319 (3.7) | <0.001 |
| White | 171,607 (95.6) | 33,082 (92.0) | 103,970 (96.5) | 34,555 (96.3) | |
| Setting (%) | |||||
| Urban | 150,958 (84.9) | 30,870 (86.8) | 90,099 (84.4) | 29,989 (84.5) | <0.001 |
| Rural | 26,787 (15.1) | 4676 (13.2) | 16,606 (15.6) | 5505 (15.5) | |
| BMI (%) | |||||
| Normal | 66,637 (37.1) | 12,143 (33.8) | 41,050 (38.1) | 13,444 (37.5) | <0.001 |
| Underweight | 960 (0.5) | 200 (0.6) | 552 (0.5) | 208 (0.6) | |
| Overweight | 74,597 (41.5) | 14,716 (40.9) | 44,632 (41.4) | 15,249 (42.5) | |
| Obese | 37,371 (20.8) | 8908 (24.8) | 21,490 (19.9) | 6973 (19.4) | |
| Iron supplement (%) | |||||
| No | 173,391 (96.6) | 34,600 (96.2) | 104,192 (96.7) | 34,599 (96.4) | <0.001 |
| Yes | 6174 (3.4) | 1367 (3.8) | 3532 (3.3) | 1275 (3.6) | |
| TPI (mean (SD)) | −1.57 (2.88) | −1.22 (3.04) | −1.68 (2.82) | −1.59 (2.87) | <0.001 |
| Hypertension (%) | |||||
| No | 148,867 (82.9) | 29,703 (82.6) | 89,697 (83.3) | 29,467 (82.1) | <0.001 |
| Yes | 30,698 (17.1) | 6264 (17.4) | 18,027 (16.7) | 6407 (17.9) | |
| Smoking status (%) | |||||
| Never | 102,830 (57.3) | 20,464 (56.9) | 62,395 (57.9) | 19,971 (55.7) | <0.001 |
| Previous | 62,569 (34.8) | 11,570 (32.2) | 37,702 (35.0) | 13,297 (37.1) | |
| Current | 14,166 (7.9) | 3933 (10.9) | 7627 (7.1) | 2606 (7.3) | |
| Drinking status (%) | |||||
| Never | 5814 (3.2) | 1834 (5.1) | 3082 (2.9) | 898 (2.5) | <0.001 |
| Previous | 5311 (3.0) | 1492 (4.1) | 2880 (2.7) | 939 (2.6) | |
| Current | 168,440 (93.8) | 32,641 (90.8) | 101,762 (94.5) | 34,037 (94.9) | |
| PA (%) | |||||
| Low | 27,931 (18.3) | 6493 (21.9) | 16,701 (18.2) | 4737 (15.1) | <0.001 |
| Moderate | 64,486 (42.3) | 12,269 (41.3) | 39,392 (43.0) | 12,825 (40.9) | |
| High | 60,134 (39.4) | 10,917 (36.8) | 35,437 (38.7) | 13,780 (44.0) | |
| History of diabetes (%) | |||||
| No | 172,522 (96.1) | 34,396 (95.6) | 103,712 (96.3) | 34,414 (95.9) | <0.001 |
| Yes | 7043 (3.9) | 1571 (4.4) | 4012 (3.7) | 1460 (4.1) | |
| History of obesity (%) | |||||
| No | 175,658 (97.8) | 35,026 (97.4) | 105,491 (97.9) | 35,141 (98.0) | <0.001 |
| Yes | 3907 (2.2) | 941 (2.6) | 2233 (2.1) | 733 (2.0) | |
| Antidiabetics (%) | |||||
| No | 174,519 (97.2) | 34,798 (96.7) | 104,878 (97.4) | 34,843 (97.1) | <0.001 |
| Yes | 5046 (2.8) | 1169 (3.3) | 2846 (2.6) | 1031 (2.9) | |
| Antilipemic (%) | |||||
| No | 155,360 (86.5) | 31,104 (86.5) | 93,481 (86.8) | 30,775 (85.8) | <0.001 |
| Yes | 24,205 (13.5) | 4863 (13.5) | 14,243 (13.2) | 5099 (14.2) | |
| Baseline CVD | |||||
| No | 172,998 (96.3) | 34,623 (96.3) | 103,937 (96.5) | 34,438 (96.0) | <0.001 |
| Yes | 6567 (3.7) | 1344 (3.7) | 3787 (3.5) | 1436 (4.0) | |
| Events n (%) | HR (95% CI) | p-Value | Adjusted HR (95% CI) a | p-Value | |
|---|---|---|---|---|---|
| Low iron intake (n = 35,967) | 1179 (3.28) | Reference | Reference | ||
| Moderate iron intake (n = 107,724) | 3943 (3.66) | 1.11 (1.04, 1.18) | <0.001 | 1.05 (0.98, 1.12) | 0.18 |
| High iron intake (n = 35,874) | 1571 (4.38) | 1.33 (1.24, 1.44) | <0.001 | 1.13 (1.05, 1.22) | <0.001 |
| GO | Category | Description | Count | % | Log10 (p) | Log10 (q) |
|---|---|---|---|---|---|---|
| GO:0008016 | GO Biological Processes | regulation of heart contraction | 10 | 16.67 | −11.06 | −6.72 |
| GO:0060371 | GO Biological Processes | regulation of atrial cardiac muscle cell membrane depolarization | 4 | 6.67 | −8.76 | −5.13 |
| GO:0048514 | GO Biological Processes | blood vessel morphogenesis | 9 | 15 | −6.79 | −3.59 |
| WP3924 | WikiPathways | Hfe effect on hepcidin production | 3 | 5 | −6.59 | −3.45 |
| GO:0071560 | GO Biological Processes | cellular response to transforming growth factor beta stimulus | 5 | 8.33 | −4.74 | −1.97 |
| GO:0007178 | GO Biological Processes | transmembrane receptor protein serine/threonine kinase signaling pathway | 5 | 8.33 | −4.36 | −1.65 |
| hsa04152 | KEGG Pathway | AMPK signaling pathway | 4 | 6.67 | −4 | −1.44 |
| GO:0030900 | GO Biological Processes | forebrain development | 6 | 10 | −3.82 | −1.3 |
| GO:2000772 | GO Biological Processes | regulation of cellular senescence | 3 | 5 | −3.77 | −1.26 |
| GO:0002067 | GO Biological Processes | glandular epithelial cell differentiation | 3 | 5 | −3.53 | −1.08 |
| GO:0055013 | GO Biological Processes | cardiac muscle cell development | 3 | 5 | −3.51 | −1.06 |
| hsa04911 | KEGG Pathway | Insulin secretion | 3 | 5 | −3.17 | −0.83 |
| R-HAS-422475 | Reactome Gene Sets | Axon guidance | 6 | 10 | −3.11 | −0.79 |
| GO:0006325 | GO Biological Processes | chromatin organization | 7 | 11.67 | −3.09 | −0.79 |
| GO:0007276 | GO Biological Processes | gamete generation | 7 | 11.67 | −3.01 | −0.74 |
| R-HSA-5689603 | Reactome Gene Sets | UCH proteinases | 3 | 5 | −2.96 | −0.72 |
| GO:0090263 | GO Biological Processes | positive regulation of canonical Wnt signaling pathway | 3 | 5 | −2.86 | −0.67 |
| hsa04728 | KEGG Pathway | Dopaminergic synapse | 3 | 5 | −2.64 | −0.51 |
| GO:0007605 | GO Biological Processes | sensory perception of sound | 3 | 5 | −2.42 | −0.36 |
| WP706 | WikiPathways | Sudden infant death syndrome (SIDS) susceptibility pathways | 3 | 5 | −2.4 | −0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habudele, Z.; Chen, G.; Qian, S.E.; Vaughn, M.G.; Zhang, J.; Lin, H. High Dietary Intake of Iron Might Be Harmful to Atrial Fibrillation and Modified by Genetic Diversity: A Prospective Cohort Study. Nutrients 2024, 16, 593. https://doi.org/10.3390/nu16050593
Habudele Z, Chen G, Qian SE, Vaughn MG, Zhang J, Lin H. High Dietary Intake of Iron Might Be Harmful to Atrial Fibrillation and Modified by Genetic Diversity: A Prospective Cohort Study. Nutrients. 2024; 16(5):593. https://doi.org/10.3390/nu16050593
Chicago/Turabian StyleHabudele, Zierdi, Ge Chen, Samantha E. Qian, Michael G. Vaughn, Junguo Zhang, and Hualiang Lin. 2024. "High Dietary Intake of Iron Might Be Harmful to Atrial Fibrillation and Modified by Genetic Diversity: A Prospective Cohort Study" Nutrients 16, no. 5: 593. https://doi.org/10.3390/nu16050593
APA StyleHabudele, Z., Chen, G., Qian, S. E., Vaughn, M. G., Zhang, J., & Lin, H. (2024). High Dietary Intake of Iron Might Be Harmful to Atrial Fibrillation and Modified by Genetic Diversity: A Prospective Cohort Study. Nutrients, 16(5), 593. https://doi.org/10.3390/nu16050593