
Nutritional and Physiological Properties of Thymbra spicata: In Vitro Study Using Fecal Fermentation and Intestinal Integrity Models
 , ,
, ,  , ,
, ,  , , ,
, , , 
Abstract
1. Introduction
2. Methods
2.1. Plant Collection and Extraction
2.2. Nutritional Value of Thymbra spicata
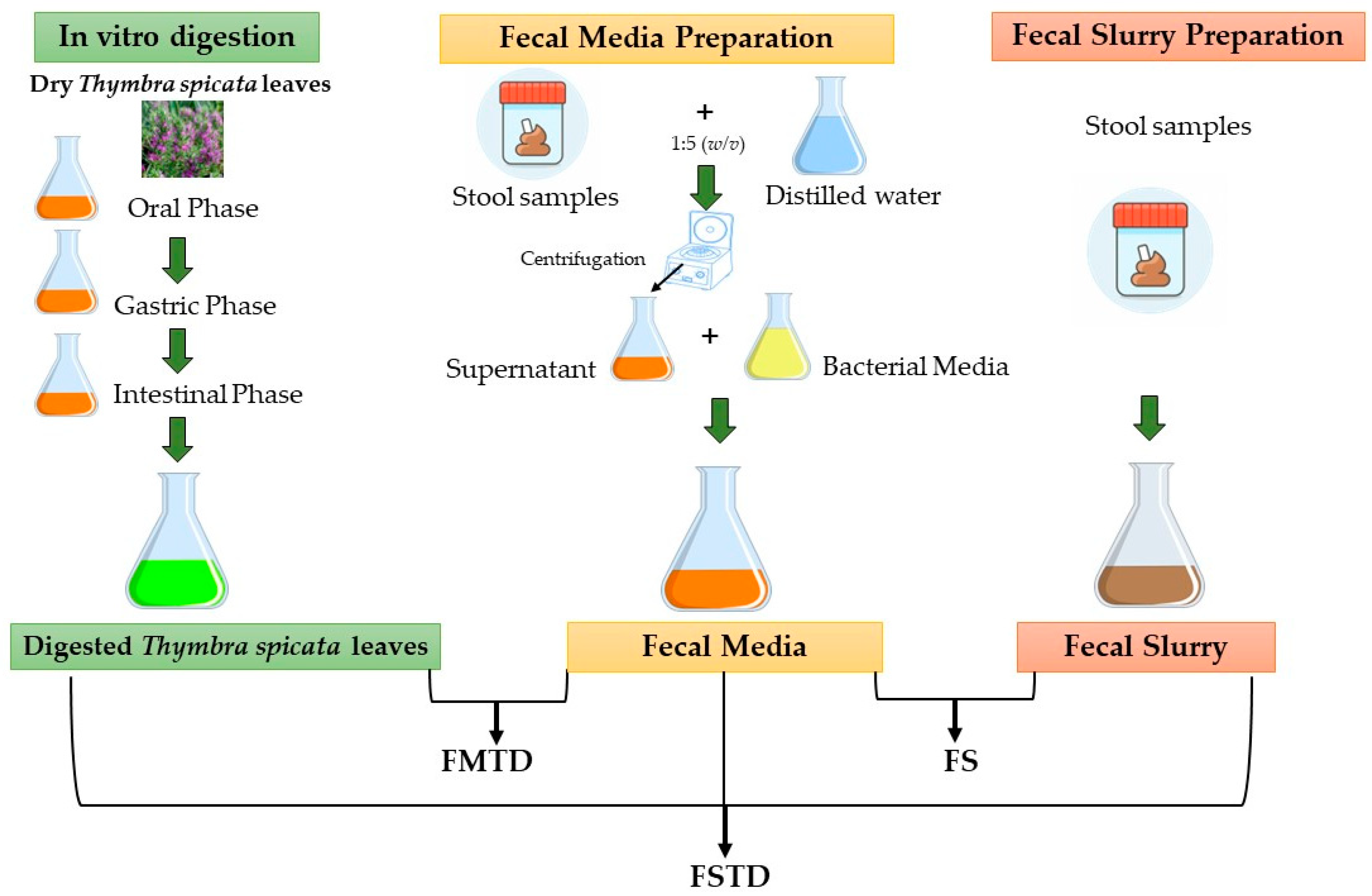
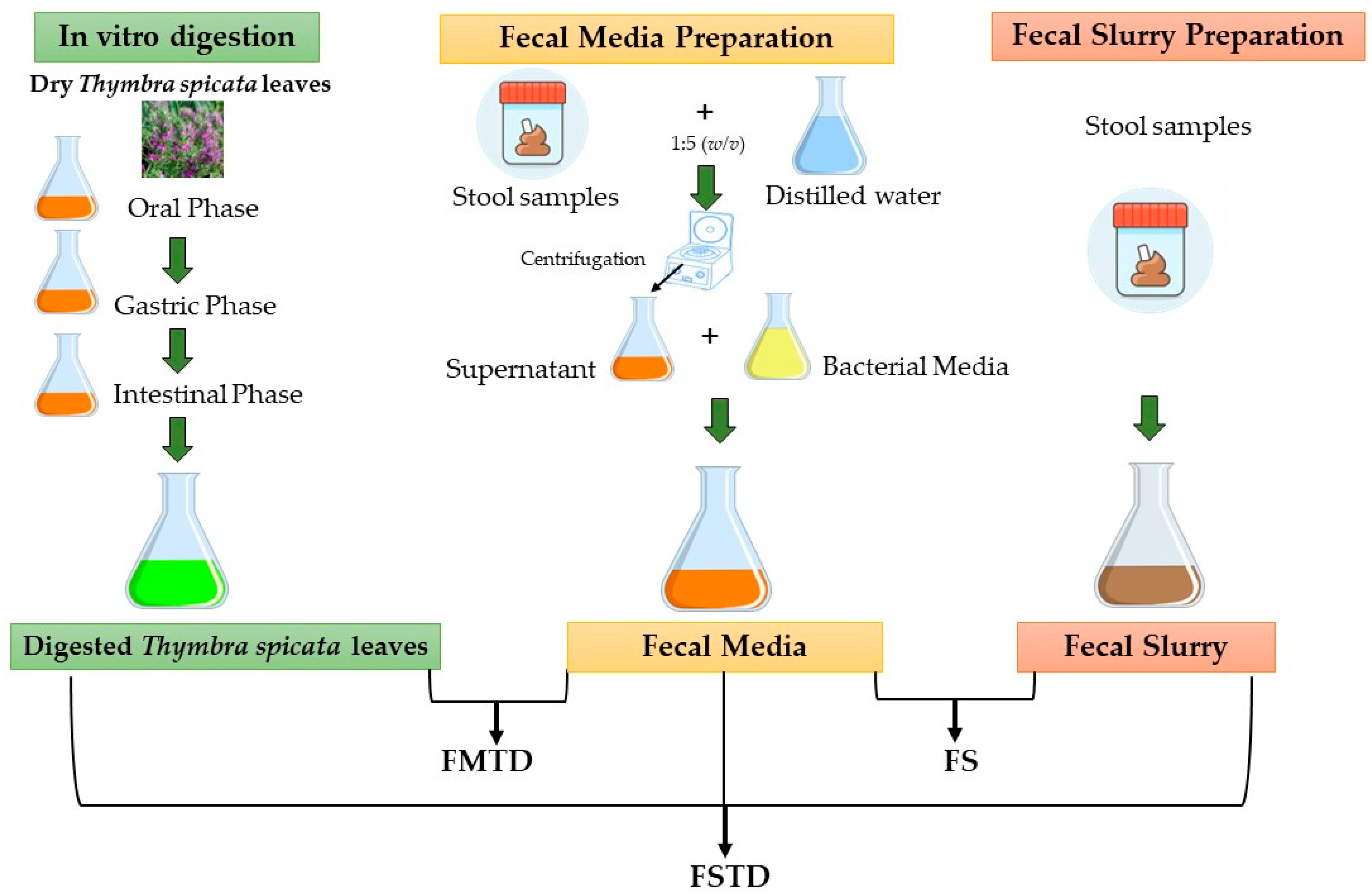
2.3. Simulated Digestion and Colonic Fermentation
2.3.1. In Vitro Simulation of Gastrointestinal Digestion
2.3.2. In Vitro Colonic Fermentation of Digested T. spicata
2.4. Fecal Microbiota Characterization
2.4.1. Enumeration of Cultivable Microorganisms
2.4.2. DNA Extraction
2.4.3. Real Time PCR
2.4.4. Fecal Batch Volatile Organic Compound (VOC) Analysis
2.5. Total Phenol Quantification
2.6. Total Flavonoid Quantification
2.7. DPPH Radical Scavenging Assay
2.8. Cell Culture
2.9. Ussing Chamber Studies
2.10. Statistical Analyses
3. Results
3.1. Chemical and Nutritional Value of Thymbra spicata
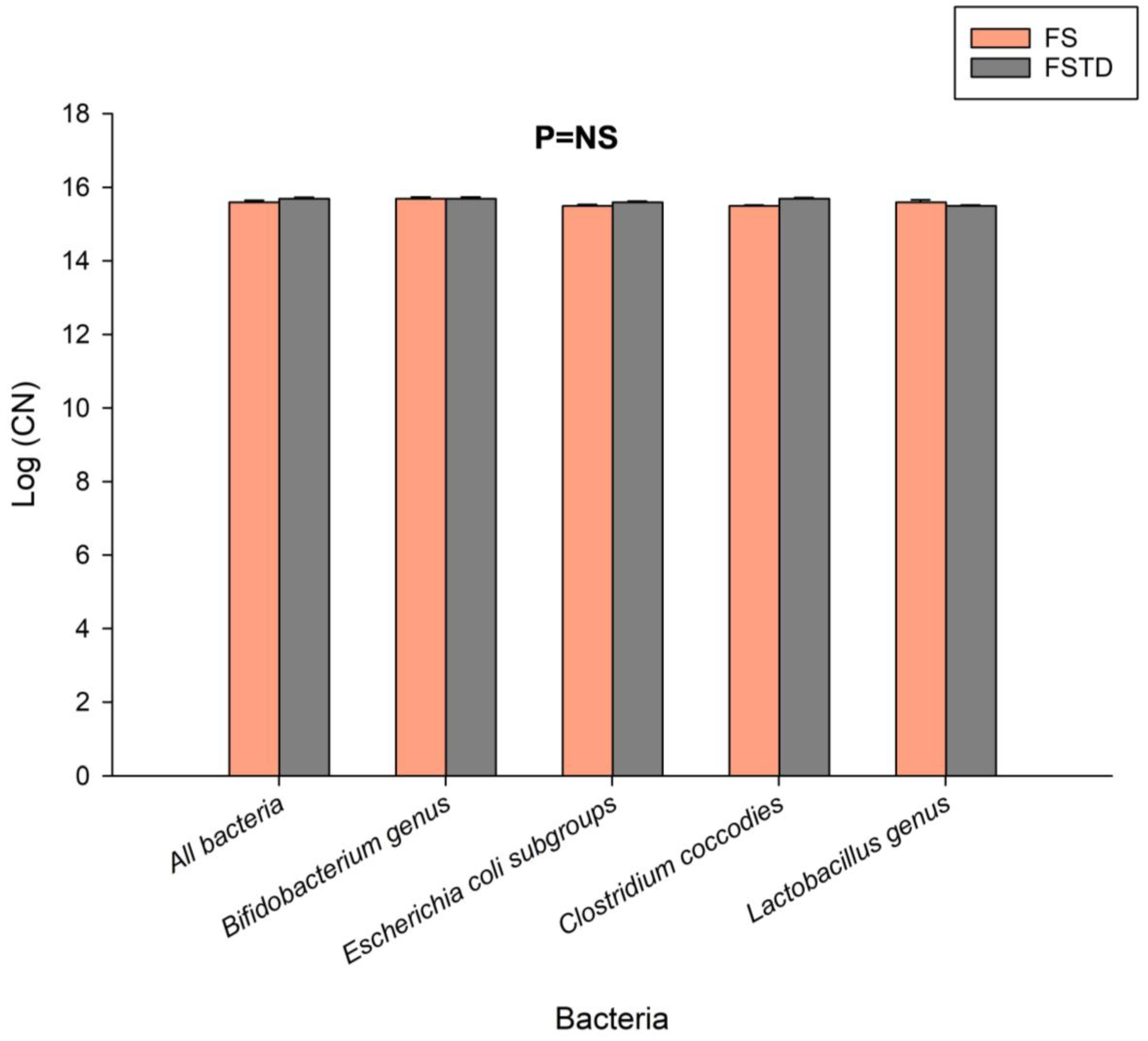
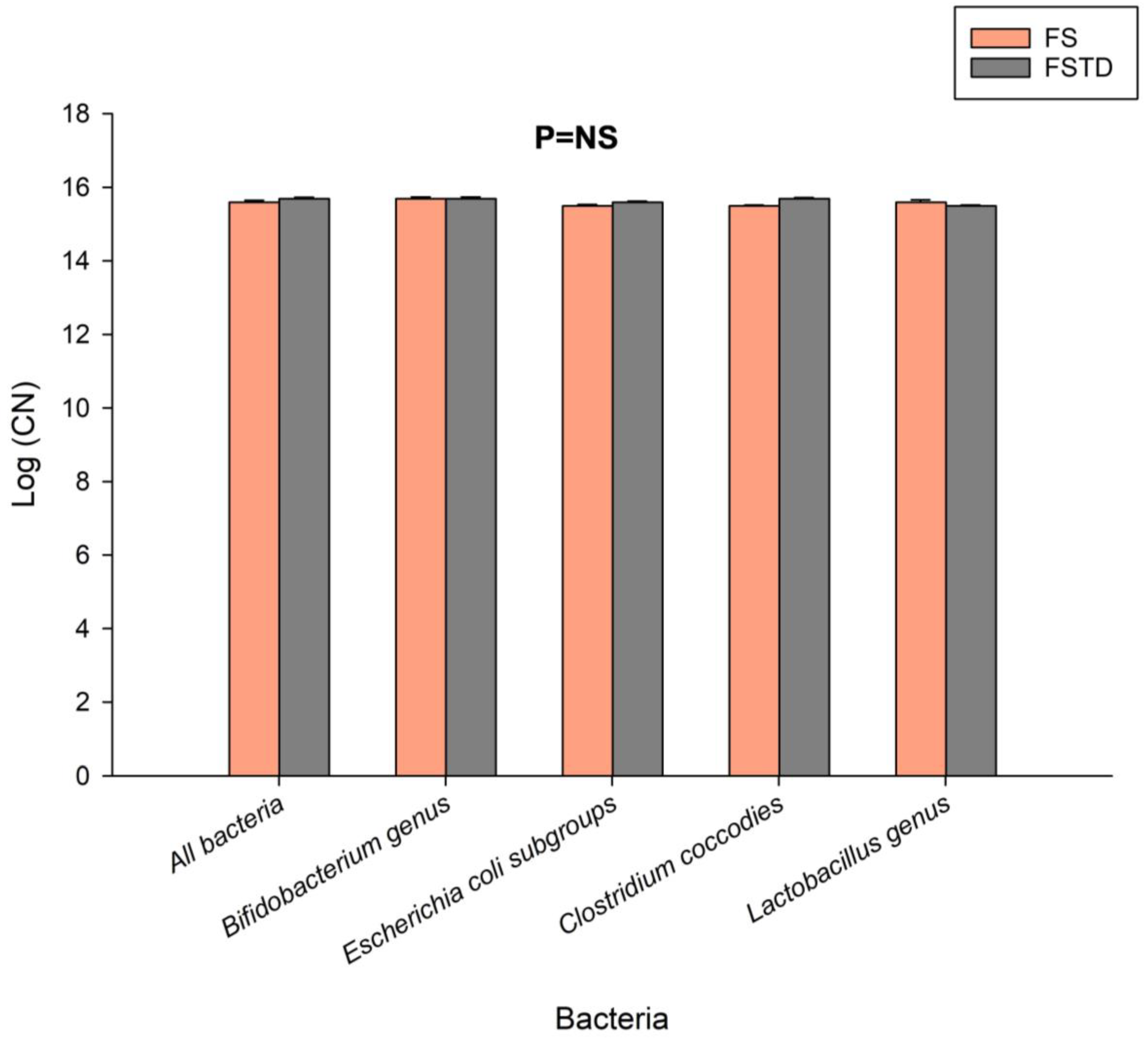
3.2. Effect of T. spicata on Fecal Microbiota after Simulated Digestion and Colonic Fermentation
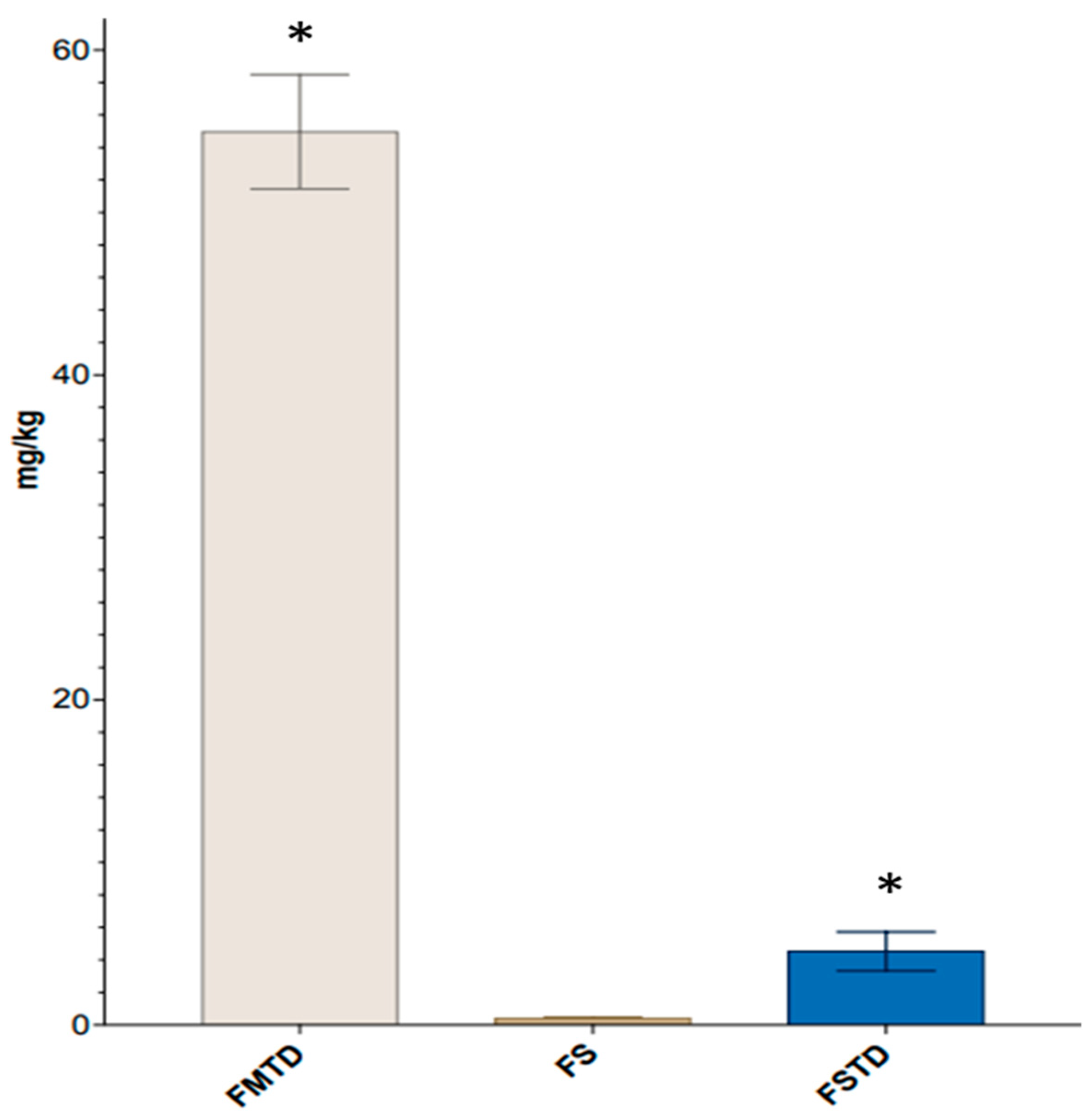
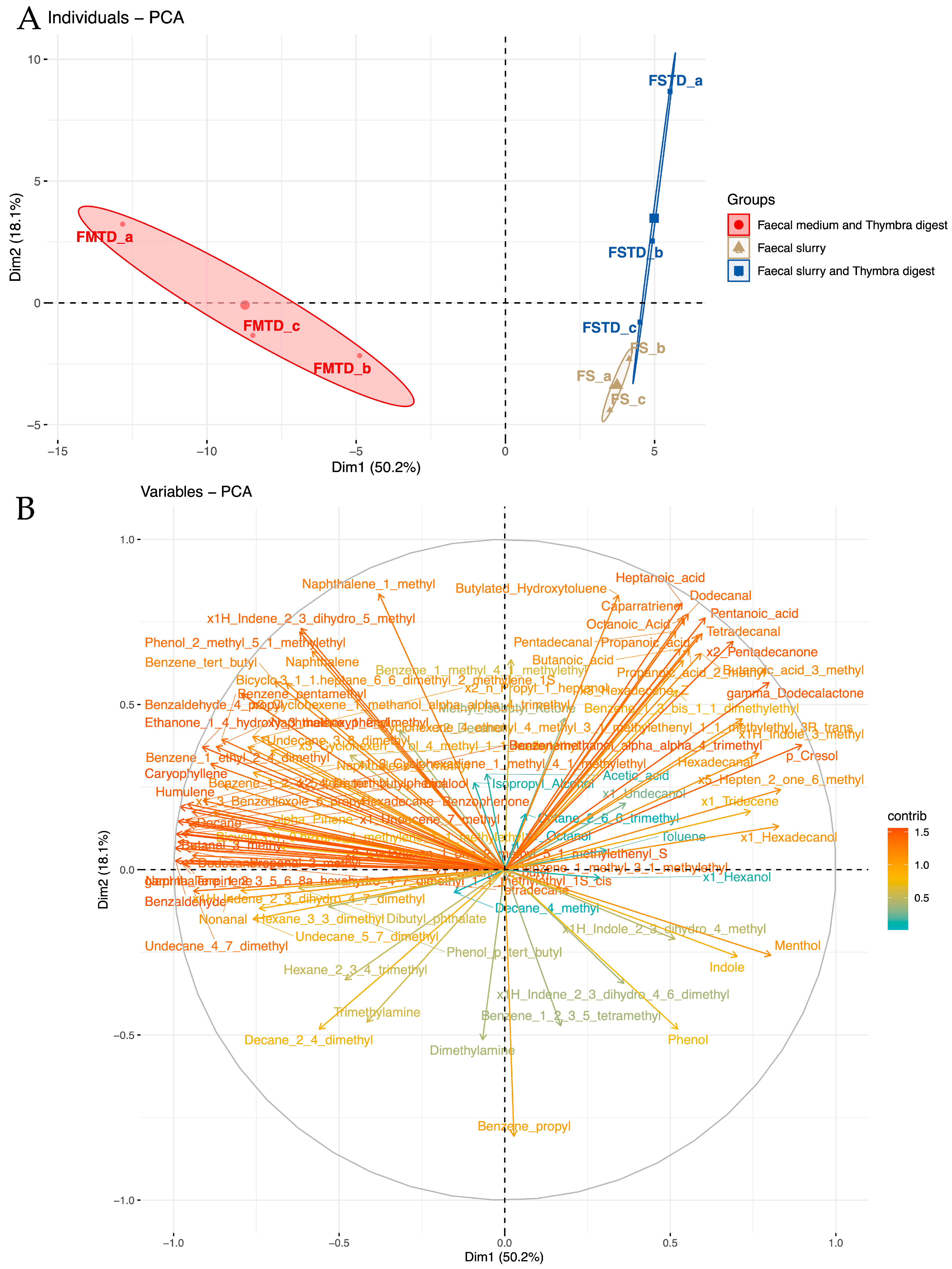
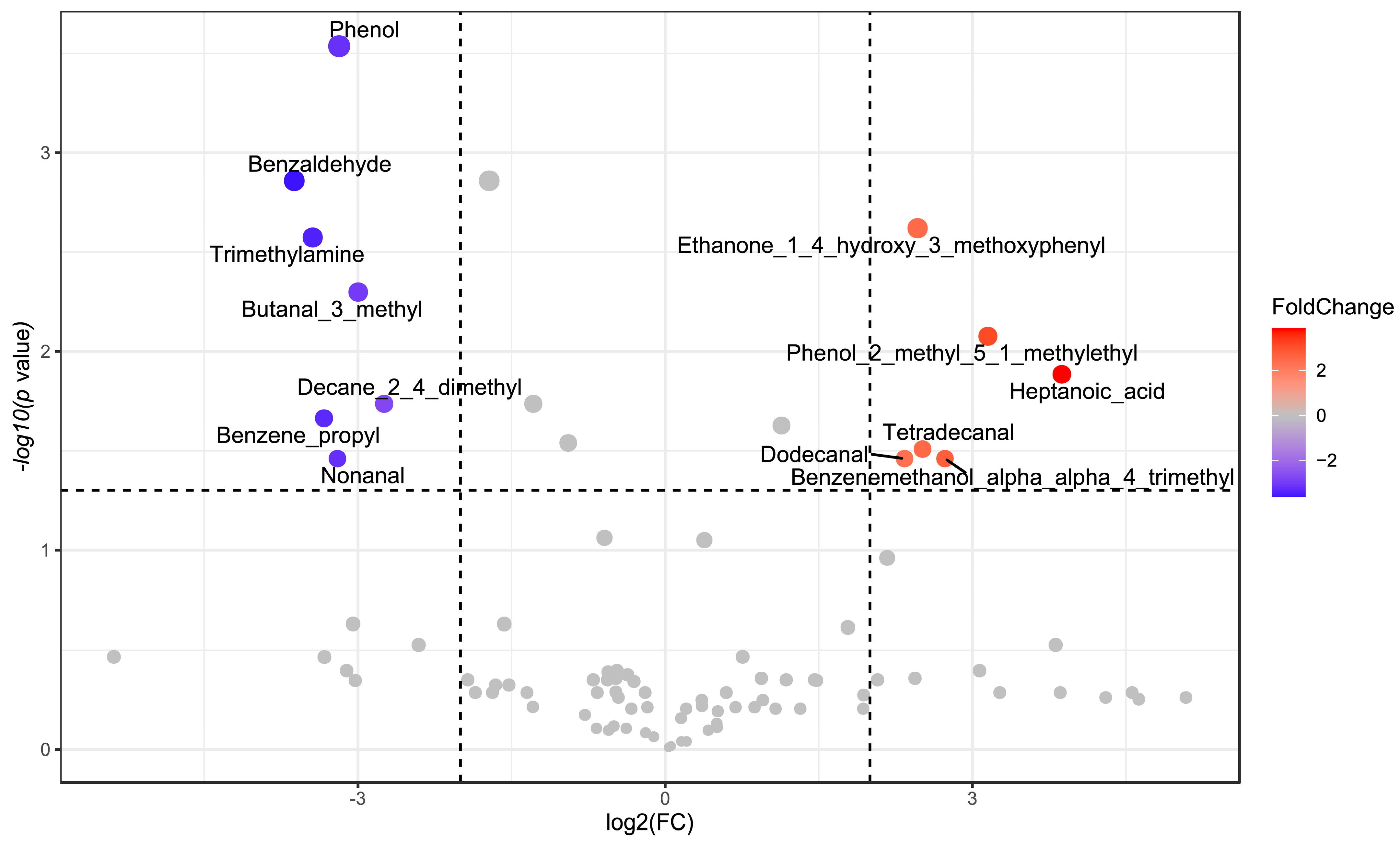
3.3. Effects of T. spicata on Fecal Volatile Organic Compounds (VOCs)
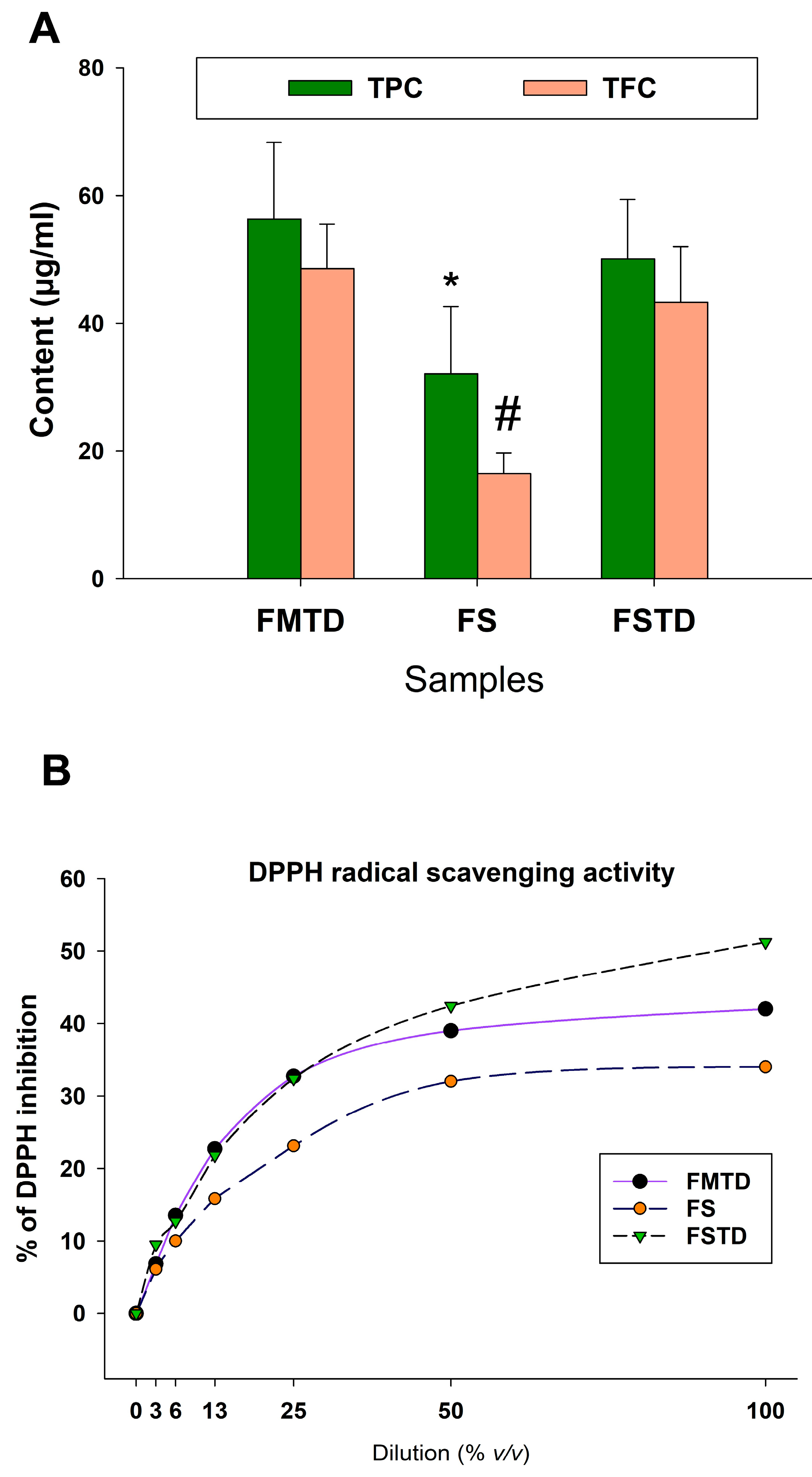
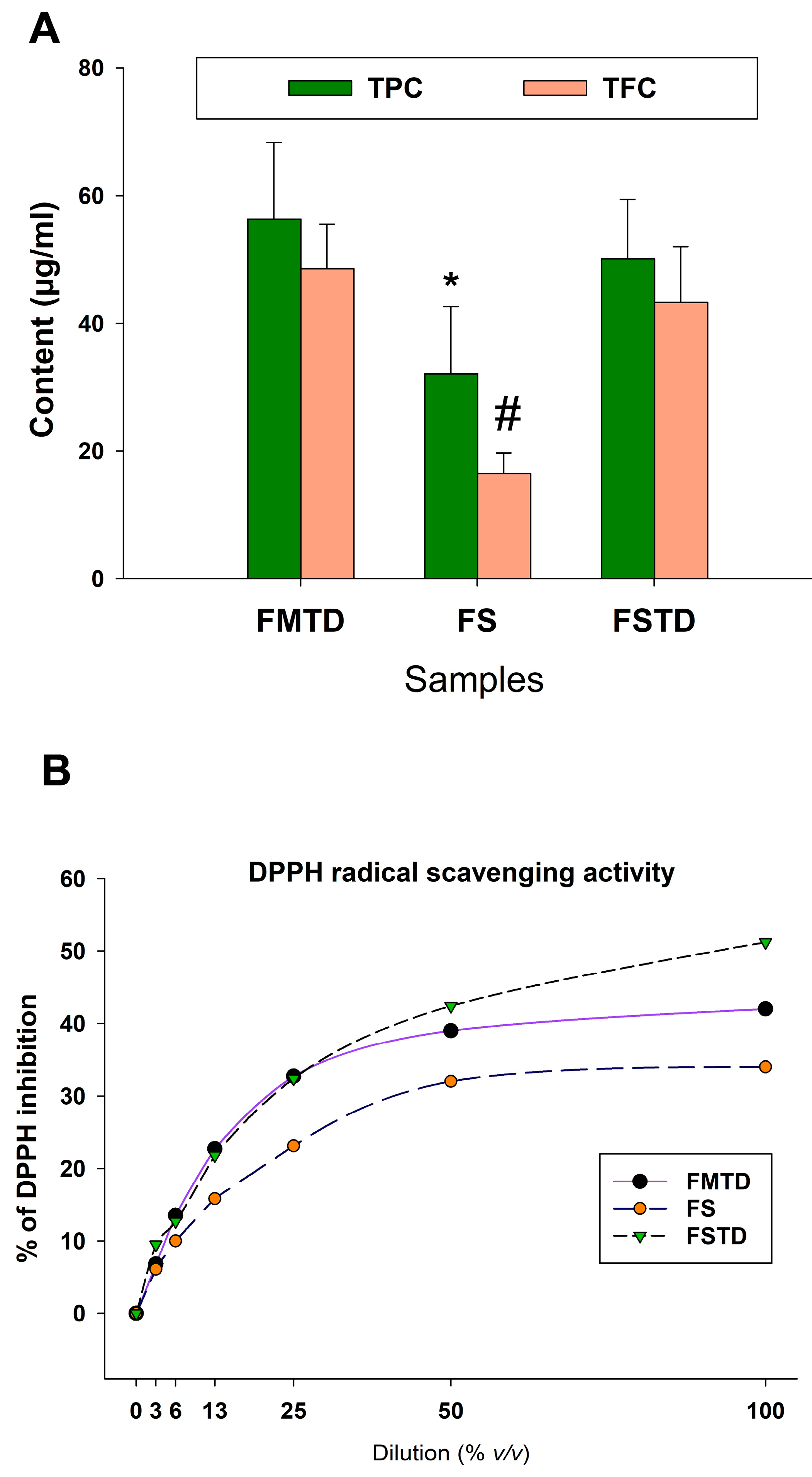
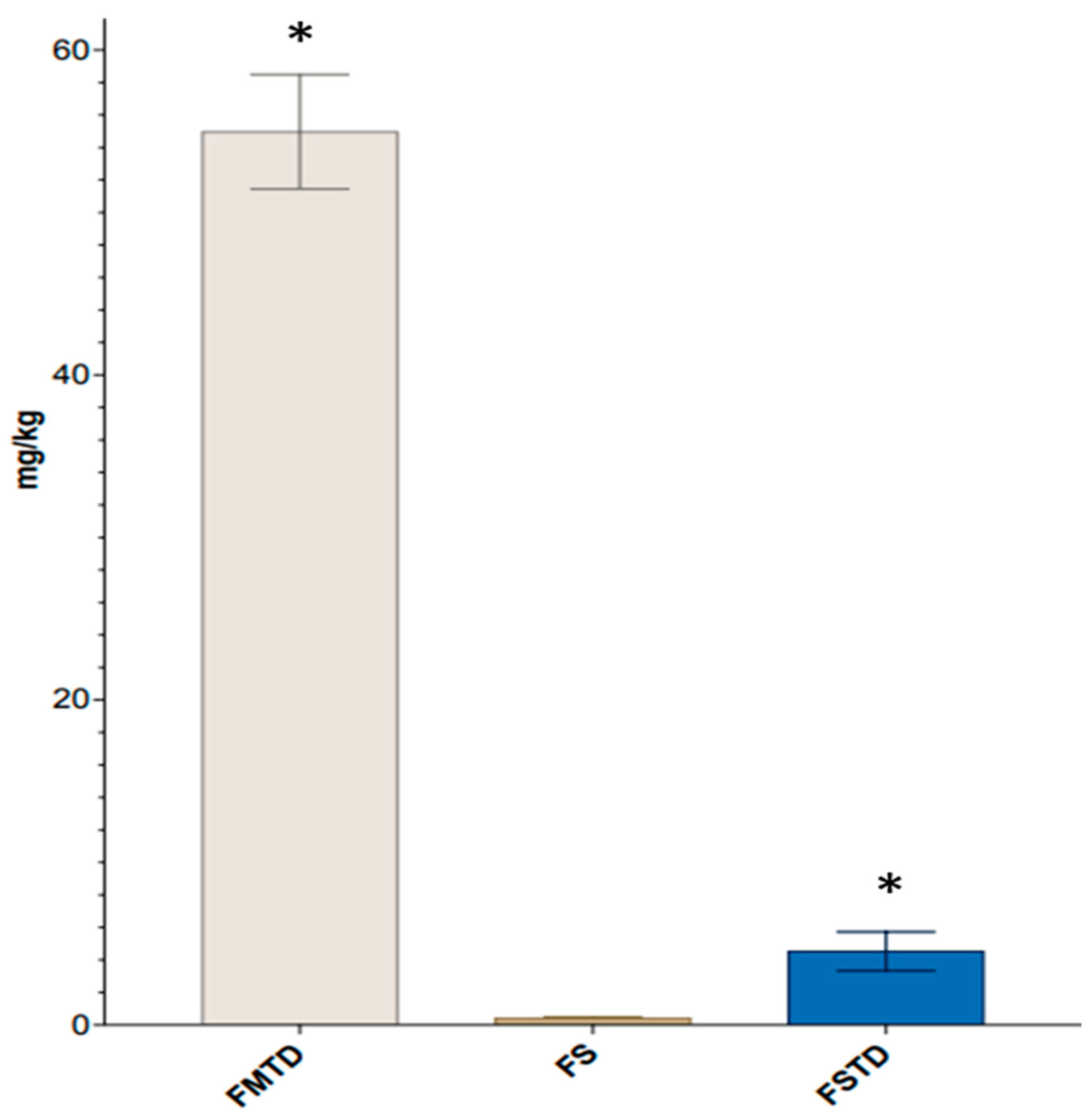
3.4. Effects of Microbial Fermentation on T. spicata Phenolic Content and Radical Scavenging Activity
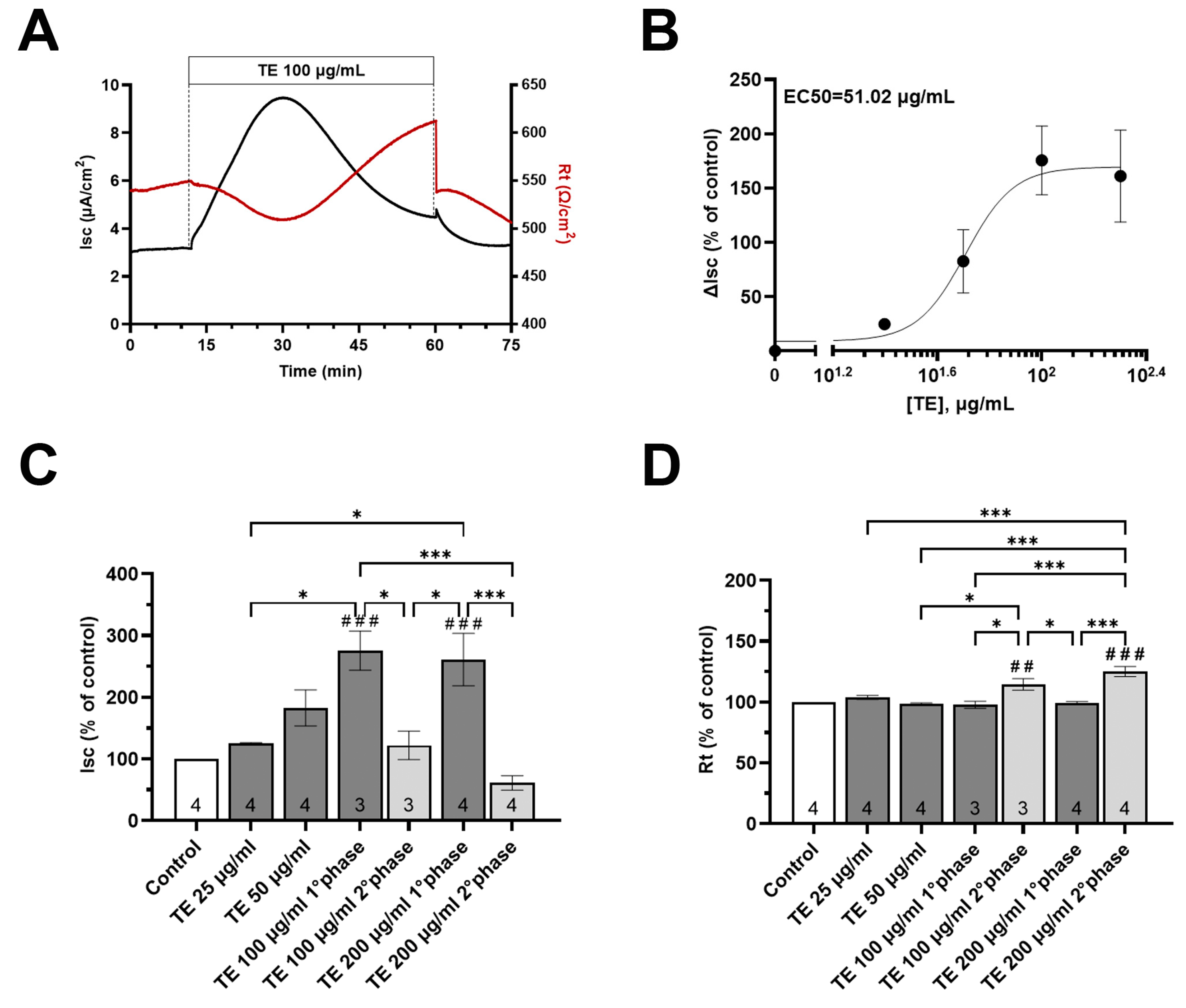
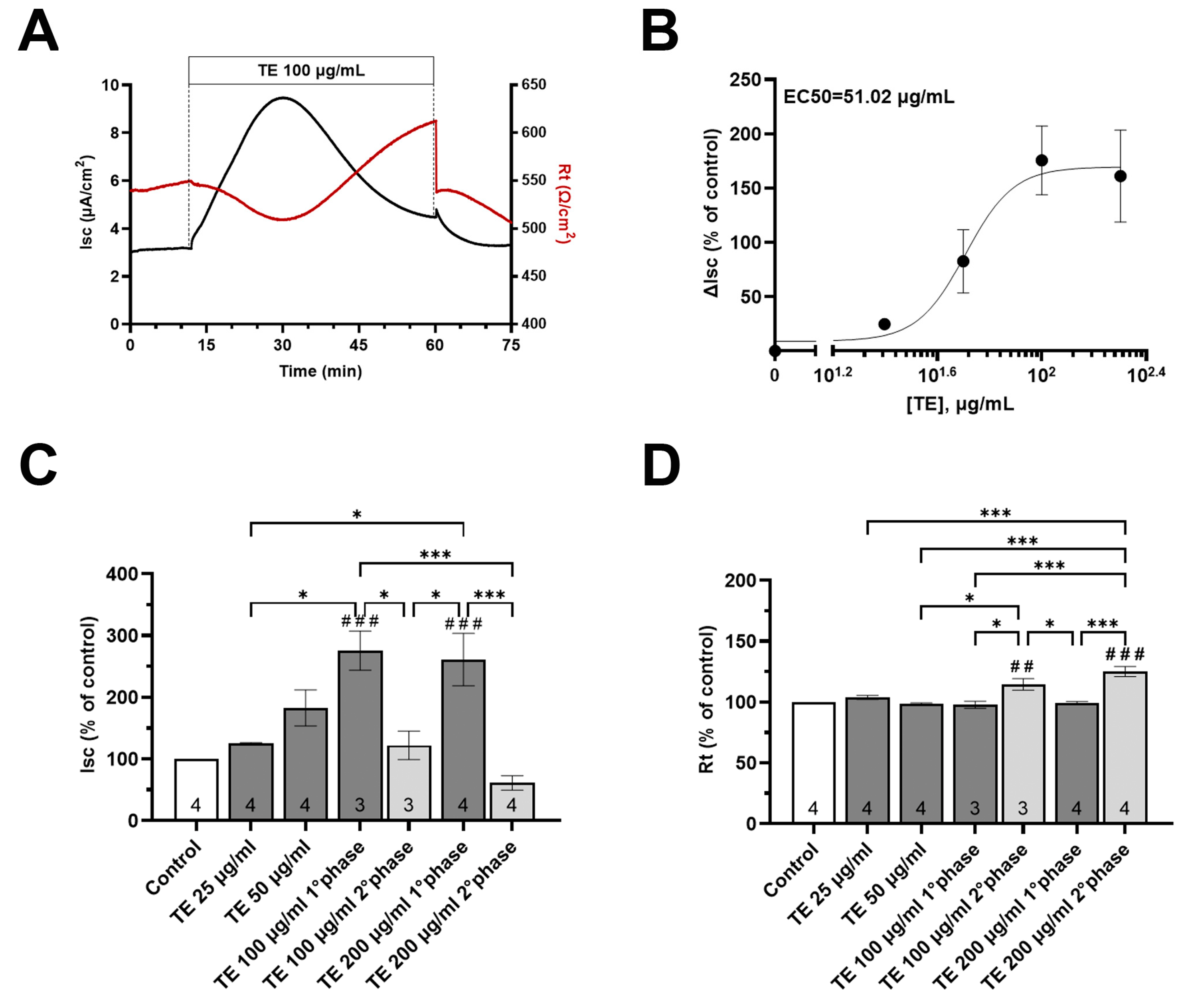
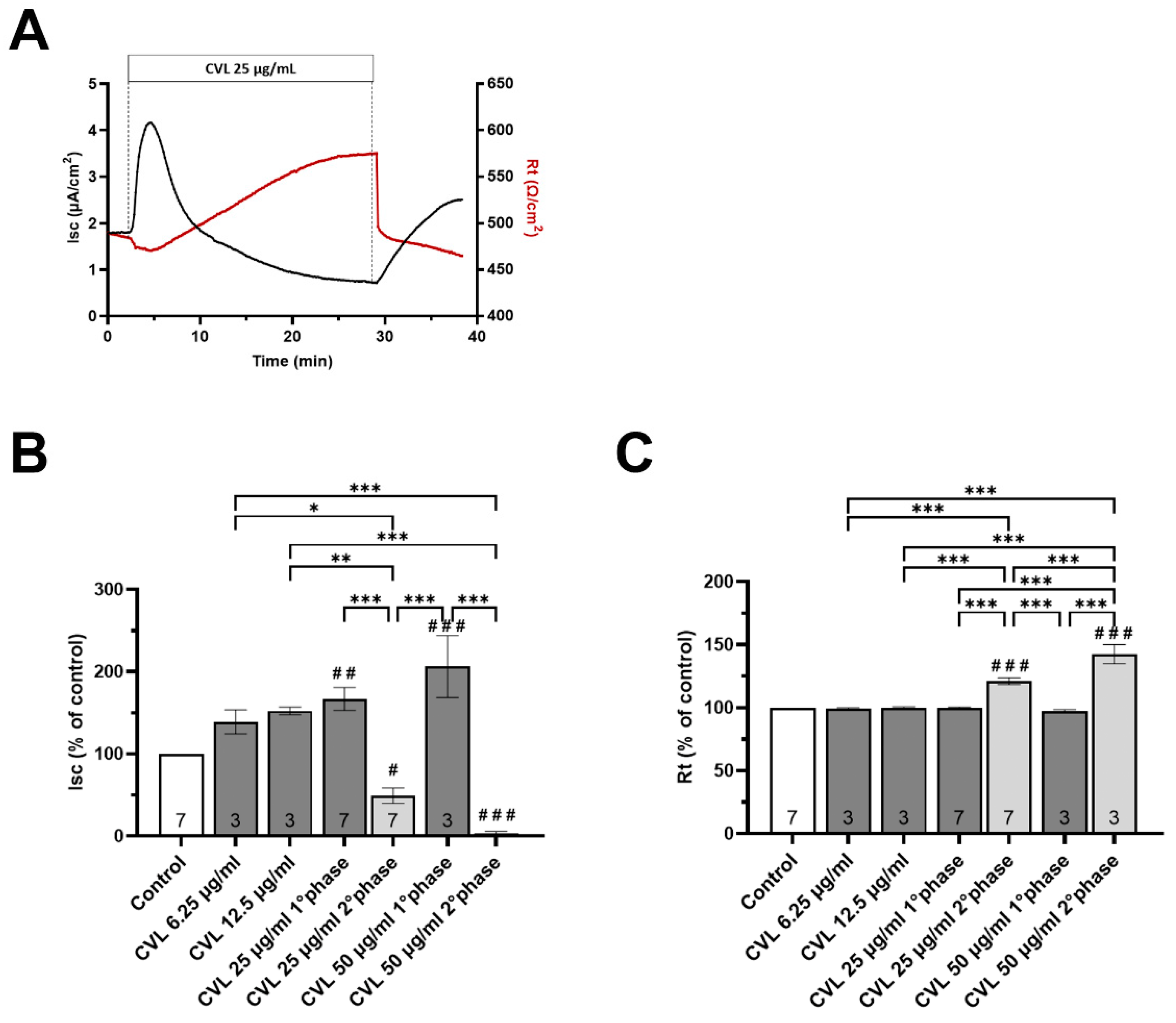
3.5. Effects of T. spicata Ethanolic Extract and Carvacrol on the Intestinal Barrier Function
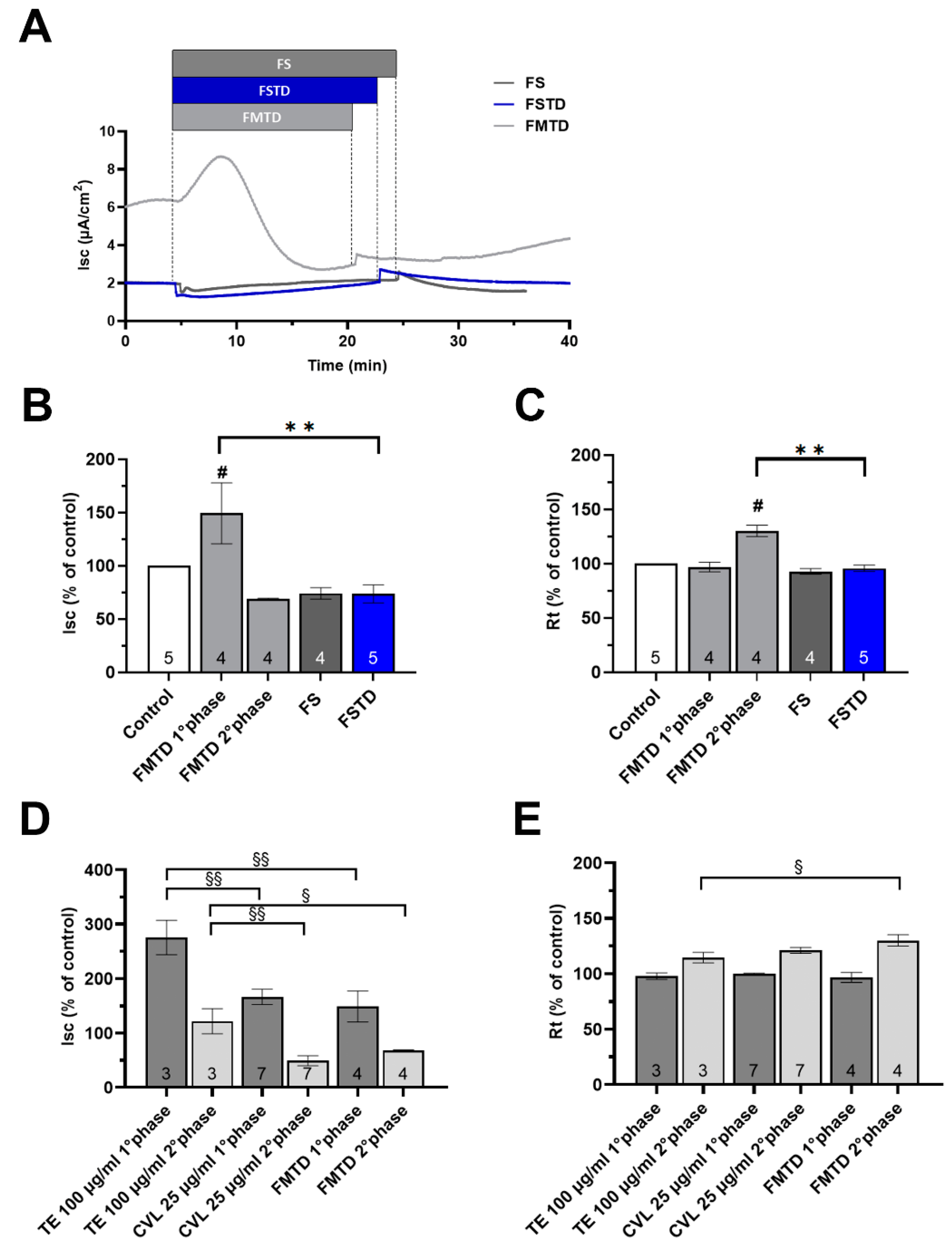
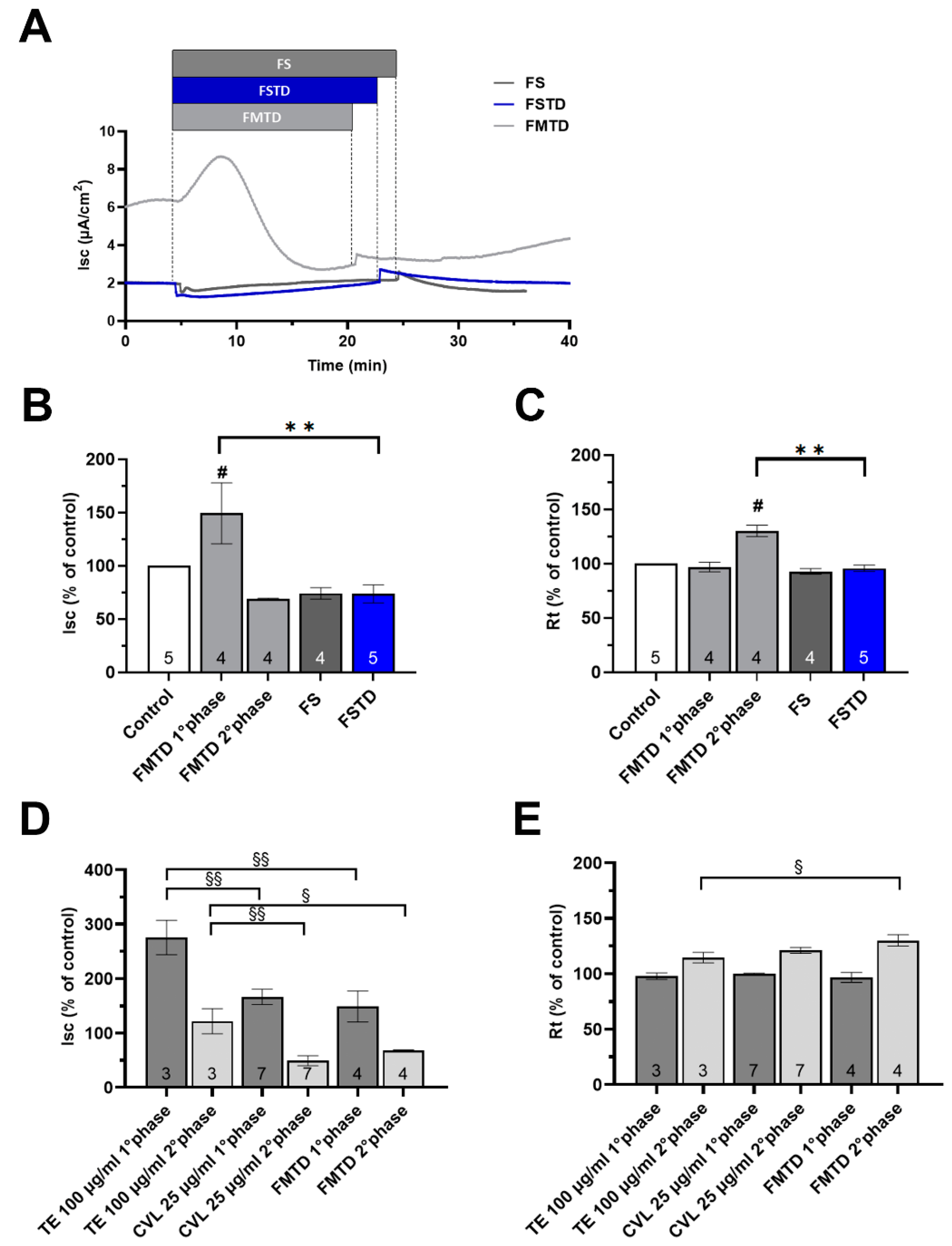
3.6. In Vitro Digestion and Microbial Fermentation Influences the Effect of T. spicata on the Intestinal Barrier
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sotos-Prieto, M.; Bhupathiraju, S.N.; Mattei, J.; Fung, T.T.; Li, Y.; Pan, A.; Willett, W.C.; Rimm, E.B.; Hu, F.B. Association of Changes in Diet Quality with Total and Cause-Specific Mortality. N. Engl. J. Med. 2017, 377, 143–153. [Google Scholar] [CrossRef]
- World Health Organization. Healthy Diet. 2023. Available online: https://www.who.int/en/news-room/fact-sheets/detail/healthy-diet (accessed on 15 December 2023).
- World Cancer Research Fund/American Institute for Cancer Research. Cancer Prevention Recommendations. 2023. Available online: https://www.wcrf.org/dietandcancer/cancer-prevention-recommendations (accessed on 15 December 2023).
- US Department of Agriculture. Dietary Guidelines for Americans: 2020–2025. US Department of Health and Human Services 2020. 2023. Available online: https://www.dietaryguidelines.gov/sites/default/files/2020-12/Dietary_Guidelines_for_Americans_2020-2025.pdf (accessed on 15 December 2023).
- Di Salvo, E.; Gangemi, S.; Genovese, C.; Cicero, N.; Casciaro, M. Polyphenols from Mediterranean Plants: Biological Activities for Skin Photoprotection in Atopic Dermatitis, Psoriasis, and Chronic Urticaria. Plants 2023, 12, 3579. [Google Scholar] [CrossRef]
- Eran Nagar, E.; Okun, Z.; Shpigelman, A. Digestive fate of polyphenols: Updated view of the influence of chemical structure and the presence of cell wall material. Curr. Opin. Food Sci. 2020, 31, 38–46. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary polyphenol impact on gut health and microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 690–711. [Google Scholar] [CrossRef]
- Scott, M.B.; Styring, A.K.; McCullagh, J.S.O. Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals. Pathogens 2022, 11, 770. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Wu, J.; Liu, J.; Zhou, Y.; Tan, Y.; Feng, W.; Peng, C. Interactions between gut microbiota and polyphenols: A mechanistic and metabolomic review. Phytomed. Int. J. Phytother. Phytopharm. 2023, 119, 154979. [Google Scholar] [CrossRef]
- Khalil, M.; Rita Caponio, G.; Diab, F.; Shanmugam, H.; Di Ciaula, A.; Khalifeh, H.; Vergani, L.; Calasso, M.; De Angelis, M.; Portincasa, P. Unraveling the beneficial effects of herbal Lebanese mixture “Za’atar”. History, studies, and properties of a potential healthy food ingredient. J. Funct. Foods 2022, 90, 104993. [Google Scholar] [CrossRef]
- Khalil, M.; Abdallah, H.; Razuka-Ebela, D.; Calasso, M.; De Angelis, M.; Portincasa, P. The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications. Antioxidants 2023, 12, 426. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Salis, A.; Damonte, G.; Daher, A.; Voci, A.; Vergani, L. Antisteatotic and antioxidant activities of Thymbra spicata L. extracts in hepatic and endothelial cells as in vitro models of non-alcoholic fatty liver disease. J. Ethnopharmacol. 2019, 239, 111919. [Google Scholar] [CrossRef] [PubMed]
- Akkol, E.K.; Avci, G.; Küçükkurt, I.; Keleş, H.; Tamer, U.; Ince, S.; Yesilada, E. Cholesterol-reducer, antioxidant and liver protective effects of Thymbra spicata L. var. spicata. J. Ethnopharmacol. 2009, 126, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Avci, G.; Kupeli, E.; Eryavuz, A.; Yesilada, E.; Kucukkurt, I. Antihypercholesterolaemic and antioxidant activity assessment of some plants used as remedy in Turkish folk medicine. J. Ethnopharmacol. 2006, 107, 418–423. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Serale, N.; Parodi, A.; Voci, A.; Vergani, L.; Daher, A. Antitumor Activity of Ethanolic Extract from Thymbra spicata L. aerial Parts: Effects on Cell Viability and Proliferation, Apoptosis Induction, STAT3, and NF-kB Signaling. Nutr. Cancer 2021, 73, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Toker, E.B.; Yeşilbağ, K. In vitro antiviral activity of Thymbra spicata L. extract on bovine respiratory viruses (BCoV, BPIV-3, BRSV, BVDV and BoHV-1). J. Appl. Microbiol. 2022, 132, 2625–2632. [Google Scholar] [CrossRef] [PubMed]
- Gedikoğlu, A.; Sökmen, M.; Çivit, A. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Sci. Nutr. 2019, 7, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Haroun, M.F.; Al-Kayali, R.S. Synergistic effect of Thymbra spicata L. extracts with antibiotics against multidrug-resistant Staphylococcus aureus and Klebsiella pneumoniae strains. Iran. J. Basic Med. Sci. 2016, 19, 1193–1200. [Google Scholar] [PubMed]
- Kerem, S.; Koşar, N.; Tekin, F.; Güreser, A.S.; Özbek, Ö. Investigation of antimicrobial activities and molecular characterization of the species belong to Origanum, Thymus and Thymbra genera by ISSR. Mol. Biol. Rep. 2023, 50, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hoca, M.; Becer, E.; Vatansever, H.S. Carvacrol is potential molecule for diabetes treatment. Arch. Physiol. Biochem. 2023, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mączka, W.; Twardawska, M.; Grabarczyk, M.; Wińska, K. Carvacrol-A Natural Phenolic Compound with Antimicrobial Properties. Antibiotics 2023, 12, 824. [Google Scholar] [CrossRef]
- Imran, M.; Aslam, M.; Alsagaby, S.A.; Saeed, F.; Ahmad, I.; Afzaal, M.; Arshad, M.U.; Abdelgawad, M.A.; El-Ghorab, A.H.; Khames, A.; et al. Therapeutic application of carvacrol: A comprehensive review. Food Sci. Nutr. 2022, 10, 3544–3561. [Google Scholar] [CrossRef]
- Al-Tawalbeh, D.; Alkhawaldeh, Y.; Abu Sawan, H.; Al-Mamoori, F.; Al-Samydai, A.; Mayyas, A. Assessment of carvacrol-antibiotic combinations’ antimicrobial activity against methicillin-resistant Staphylococcus aureus. Front. Microbiol. 2023, 14, 1349550. [Google Scholar] [CrossRef] [PubMed]
- Şimşek, H.; Gür, C.; Küçükler, S.; İleritürk, M.; Akaras, N.; Öz, M.; Kandemir, F.M. Carvacrol Reduces Mercuric Chloride-Induced Testicular Toxicity by Regulating Oxidative Stress, Inflammation, Apoptosis, Autophagy, and Histopathological Changes. Biol. Trace Elem. Res. 2023. [Google Scholar] [CrossRef]
- Zhao, W.; Deng, C.; Han, Q.; Xu, H.; Chen, Y. [Corrigendum] Carvacrol may alleviate vascular inflammation in diabetic db/db mice. Int. J. Mol. Med. 2023, 52, 90. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.; Serale, N.; Diab, F.; Baldini, F.; Portincasa, P.; Lupidi, G.; Vergani, L. Beneficial Effects of Carvacrol on In Vitro Models of Metabolically-Associated Liver Steatosis and Endothelial Dysfunction: A Role for Fatty Acids in Interfering with Carvacrol Binding to Serum Albumin. Curr. Med. Chem. 2022, 29, 5113–5129. [Google Scholar] [CrossRef]
- Diab, F.; Khalil, M.; Lupidi, G.; Zbeeb, H.; Salis, A.; Damonte, G.; Bramucci, M.; Portincasa, P.; Vergani, L. Influence of Simulated In Vitro Gastrointestinal Digestion on the Phenolic Profile, Antioxidant, and Biological Activity of Thymbra spicata L. Extracts. Antioxidants 2022, 11, 1778. [Google Scholar] [CrossRef] [PubMed]
- Kwok, L.Y.; Zhang, J.; Guo, Z.; Gesudu, Q.; Zheng, Y.; Qiao, J.; Huo, D.; Zhang, H. Characterization of fecal microbiota across seven Chinese ethnic groups by quantitative polymerase chain reaction. PLoS ONE 2014, 9, e93631. [Google Scholar] [CrossRef]
- Jian, C.; Luukkonen, P.; Yki-Järvinen, H.; Salonen, A.; Korpela, K. Quantitative PCR provides a simple and accessible method for quantitative microbiota profiling. PLoS ONE 2020, 15, e0227285. [Google Scholar] [CrossRef]
- Piccapane, F.; Gerbino, A.; Carmosino, M.; Milano, S.; Arduini, A.; Debellis, L.; Svelto, M.; Caroppo, R.; Procino, G. Aquaporin-1 Facilitates Transmesothelial Water Permeability: In Vitro and Ex Vivo Evidence and Possible Implications in Peritoneal Dialysis. Int. J. Mol. Sci. 2021, 22, 12535. [Google Scholar] [CrossRef]
- Farias-Pereira, R.; Zuk, J.B.; Khavaran, H. Plant bioactive compounds from Mediterranean diet improve risk factors for metabolic syndrome. Int. J. Food Sci. Nutr. 2023, 74, 403–423. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Front. Nutr. 2019, 6, 188. [Google Scholar] [CrossRef]
- Diab, F.; Zbeeb, H.; Baldini, F.; Portincasa, P.; Khalil, M.; Vergani, L. The Potential of Lamiaceae Herbs for Mitigation of Overweight, Obesity, and Fatty Liver: Studies and Perspectives. Molecules 2022, 27, 5043. [Google Scholar] [CrossRef] [PubMed]
- Dirican, E.; Turkez, H.; Toğar, B. Modulatory effects of Thymbra spicata L. different extracts against the mercury induced genotoxicity in human lymphocytes in vitro. Cytotechnology 2012, 64, 181–186. [Google Scholar] [CrossRef]
- Omar, G.; Abdallah, L.; Barakat, A.; Othman, R.; Bourinee, H. In vitro haemostatic efficacy of aqueous, methanol and ethanol plant extracts of three medicinal plant species in Palestine. Braz. J. Biol. 2020, 80, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Wang, H.; Chen, C.; Hu, W.; Ai, C.; Chen, L.; Teng, H. Flavonoids and gastrointestinal health: Single molecule for multiple roles. Crit. Rev. Food Sci. Nutr. 2023, 1–19. [Google Scholar] [CrossRef]
- Kiliç, T. Analysis of essential oil composition of Thymbra spicata var. spicata: Antifungal, antibacterial and antimycobacterial activities. Z. Naturforschung. C J. Biosci. 2006, 61, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, X.; Ma, X.; Geng, S.; Jiang, X.; Huang, Q.; Hu, C.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [Google Scholar] [CrossRef]
- Hall, H.N.; Wilkinson, D.J.; Le Bon, M. Oregano essential oil improves piglet health and performance through maternal feeding and is associated with changes in the gut microbiota. Anim. Microbiome 2021, 3, 2. [Google Scholar] [CrossRef]
- Kelly, C.; Gundogdu, O.; Pircalabioru, G.; Cean, A.; Scates, P.; Linton, M.; Pinkerton, L.; Magowan, E.; Stef, L.; Simiz, E.; et al. The In Vitro and In Vivo Effect of Carvacrol in Preventing Campylobacter Infection, Colonization and in Improving Productivity of Chicken Broilers. Foodborne Pathog. Dis. 2017, 14, 341–349. [Google Scholar] [CrossRef]
- Mooyottu, S.; Flock, G.; Upadhyay, A.; Upadhyaya, I.; Maas, K.; Venkitanarayanan, K. Protective Effect of Carvacrol against Gut Dysbiosis and Clostridium difficile Associated Disease in a Mouse Model. Front. Microbiol. 2017, 8, 625. [Google Scholar] [CrossRef]
- Ran, C.; Hu, J.; Liu, W.; Liu, Z.; He, S.; Dan, B.C.; Diem, N.N.; Ooi, E.L.; Zhou, Z. Thymol and Carvacrol Affect Hybrid Tilapia through the Combination of Direct Stimulation and an Intestinal Microbiota-Mediated Effect: Insights from a Germ-Free Zebrafish Model. J. Nutr. 2016, 146, 1132–1140. [Google Scholar] [CrossRef]
- Upadhyay, A.; Arsi, K.; Wagle, B.R.; Upadhyaya, I.; Shrestha, S.; Donoghue, A.M.; Donoghue, D.J. Trans-Cinnamaldehyde, Carvacrol, and Eugenol Reduce Campylobacter jejuni Colonization Factors and Expression of Virulence Genes In Vitro. Front. Microbiol. 2017, 8, 713. [Google Scholar] [CrossRef]
- Mooyottu, S.; Kollanoor-Johny, A.; Flock, G.; Bouillaut, L.; Upadhyay, A.; Sonenshein, A.L.; Venkitanarayanan, K. Carvacrol and trans-cinnamaldehyde reduce Clostridium difficile toxin production and cytotoxicity in vitro. Int. J. Mol. Sci. 2014, 15, 4415–4430. [Google Scholar] [CrossRef]
- Burt, S.A.; Adolfse, S.J.; Ahad, D.S.; Tersteeg-Zijderveld, M.H.; Jongerius-Gortemaker, B.G.; Post, J.A.; Brüggemann, H.; Santos, R.R. Cinnamaldehyde, Carvacrol and Organic Acids Affect Gene Expression of Selected Oxidative Stress and Inflammation Markers in IPEC-J2 Cells Exposed to Salmonella typhimurium. Phytother. Res. 2016, 30, 1988–2000. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Horváthová, E.; Sramková, M.; Lábaj, J.; Slamenová, D. Study of cytotoxic, genotoxic and DNA-protective effects of selected plant essential oils on human cells cultured in vitro. Neuro Endocrinol. Lett. 2006, 27 (Suppl. S2), 44–47. [Google Scholar] [PubMed]
- Slamenová, D.; Horváthová, E.; Sramková, M.; Marsálková, L. DNA-protective effects of two components of essential plant oils carvacrol and thymol on mammalian cells cultured in vitro. Neoplasma 2007, 54, 108–112. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primers | Primer Sequence (5′–3′) | Gene | Product Size (bp) | T Annealing (°C) | Reference |
|---|---|---|---|---|---|---|
| All Bacteria | Uni331F | TCCTACGGGAGGCAGCAGT | 16S rRNA | 466 | 58 | [29] |
| Uni797R | GGACTACCAGGGTATCTATCCTGTT | |||||
| Lactobacillus genus | Lp-F | AAAATCATGCGTGCGGGTAC | 16S rRNA | 341 | 55 | [29] |
| Lp-R | ATGTTGCGTTGGCTTCGTCT | |||||
| Bifidobacterium genus | Bifid-F | CTCCTGGAAACGGGTGG | 16S rRNA | 550 | 55 | [29] |
| Bifid-R | GGTGTTCTTCCCGATATCTACA | |||||
| Escherichia coli subgroup (E. coli, Hafnia alvei, Shigella spp.) | Eco-F | GTTAATACCTTTGCTCATTGA | V3-V4 hypervariable regions | 340 | 60 | [30] |
| Eco-R | ACCAGGGTATCTAATCCTGTT | |||||
| Clostridium coccoides group | Ccoc-F Ccoc-R | AAATGACGGTACCTGACTAA CTTTGAGTTTCATTCTTGCGAA | 16S rRNA | 440 | 50 | [29] |
| Compounds | Quantity | Compounds | Quantity |
|---|---|---|---|
| Total fat (g/100 g) | 4.3 | Total carbohydrates (g/100 g) | 52.7 |
| Saturated fatty acids (g/100 g) | 0.9 | Total dietary fiber (g/100 g) | 12.3 |
| Fatty acid methyl esters (% of fat content) | Simple sugars (g/100 g) | ||
| C12:0—Lauric acid | <0.1 | Fructose | 0.03 |
| C14:0—Myristic acid | <0.1 | Galactose | n.d |
| C16:0—Palmitic acid | 17.5 | Glucose | 1.69 |
| C16:1—Palmitoleic acid | <0.1 | Lactose | n.d |
| C17:0—Heptadecanoic acid | <0.1 | Maltose | n.d |
| C17:1—Heptadecenoic acid | <0.1 | Sucrose | 0.02 |
| C18:0—Stearic acid | 2.6 | Sum sugars | 1.74 |
| C18:1—Oleic acid | 40.6 | Nitrogen (g/100 g) | 0.99 |
| C18:2—Linoleic acid | 35.5 | Total protein (g/100 g) | 6.19 |
| C20:0—Arachic acid | 2.6 | Moisture (g/100 g) | 17.0 |
| C18:3—Linolenic acid | 0.5 | Salt (g/100 g) | 0.009 |
| C20:1—Eicosenoco acid | <0.1 | Ash (g/100 g) | 9.11 |
| C22:0—Beenic acid | <0.1 | Energy value (Kcal/100 g) | 324 |
| C22:1—Erucic acid | <0.1 | Energy value (KJ/100 g) | 1367 |
| C24:0—Lignoceric acid | <0.1 |
| Viale Microbes (Log UFC/mL) | FS (Control) | FSTD | p-Value |
|---|---|---|---|
| Total aerobes | 9.3 ± 0.3 | 9.0 ± 0.4 | 0.61 |
| Total anaerobes | 8.0 ± 0.5 | 8.0 ± 0.4 | 0.99 |
| Lactobacilli | 5.0 ± 1.0 | 4.4 ± 1.0 | 0.69 |
| Lactococci | 7.8 ± 0.4 | 7.6 ± 0.5 | 0.81 |
| Bifiobacteria | 7.1 ± 0.7 | 6.9 ± 0.5 | 0.86 |
| Enterobacteriaceae | 8.8 ± 0.2 | 8.2 ± 0.6 | 0.39 |
| Enterococci | 4.9 ± 0.1 | 3.5 ± 1.4 | 0.46 |
| Clostridi | 8.8 ± 0.1 | 8.3 ± 0.2 | 0.09 |
| SCFAs (ppm) | FMTD | FS | FSTD |
|---|---|---|---|
| Acetic acid | n.d. | 10.7 ± 2.3 | 10.1 ± 1.2 |
| Propanoic acid | n.d. | 4.8 ± 2.1 * | 8.5 ± 2.2 * |
| Isobutyric acid | n.d. | 3.2 ± 0.9 | 1.8 ± 1.2 |
| Butanoic acid | n.d. | 3.6 ± 1.2 | 4.9 ± 1.1 |
| Isovaleric acid | n.d. | 1.5 ± 1.30 | 2.7 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, M.; Piccapane, F.; Vacca, M.; Celano, G.; Mahdi, L.; Perniola, V.; Apa, C.A.; Annunziato, A.; Iacobellis, I.; Procino, G.; et al. Nutritional and Physiological Properties of Thymbra spicata: In Vitro Study Using Fecal Fermentation and Intestinal Integrity Models. Nutrients 2024, 16, 588. https://doi.org/10.3390/nu16050588
Khalil M, Piccapane F, Vacca M, Celano G, Mahdi L, Perniola V, Apa CA, Annunziato A, Iacobellis I, Procino G, et al. Nutritional and Physiological Properties of Thymbra spicata: In Vitro Study Using Fecal Fermentation and Intestinal Integrity Models. Nutrients. 2024; 16(5):588. https://doi.org/10.3390/nu16050588
Chicago/Turabian StyleKhalil, Mohamad, Francesca Piccapane, Mirco Vacca, Giuseppe Celano, Laura Mahdi, Valeria Perniola, Carmen Aurora Apa, Alessandro Annunziato, Ilaria Iacobellis, Giuseppe Procino, and et al. 2024. "Nutritional and Physiological Properties of Thymbra spicata: In Vitro Study Using Fecal Fermentation and Intestinal Integrity Models" Nutrients 16, no. 5: 588. https://doi.org/10.3390/nu16050588
APA StyleKhalil, M., Piccapane, F., Vacca, M., Celano, G., Mahdi, L., Perniola, V., Apa, C. A., Annunziato, A., Iacobellis, I., Procino, G., Calasso, M., De Angelis, M., Caroppo, R., & Portincasa, P. (2024). Nutritional and Physiological Properties of Thymbra spicata: In Vitro Study Using Fecal Fermentation and Intestinal Integrity Models. Nutrients, 16(5), 588. https://doi.org/10.3390/nu16050588