
Probiotic Limosilactobacillus reuteri DSM 17938 Changes Foxp3 Deficiency-Induced Dyslipidemia and Chronic Hepatitis in Mice
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Altered Plasma Lipid Profiles in Treg-Deficiency SF Mice
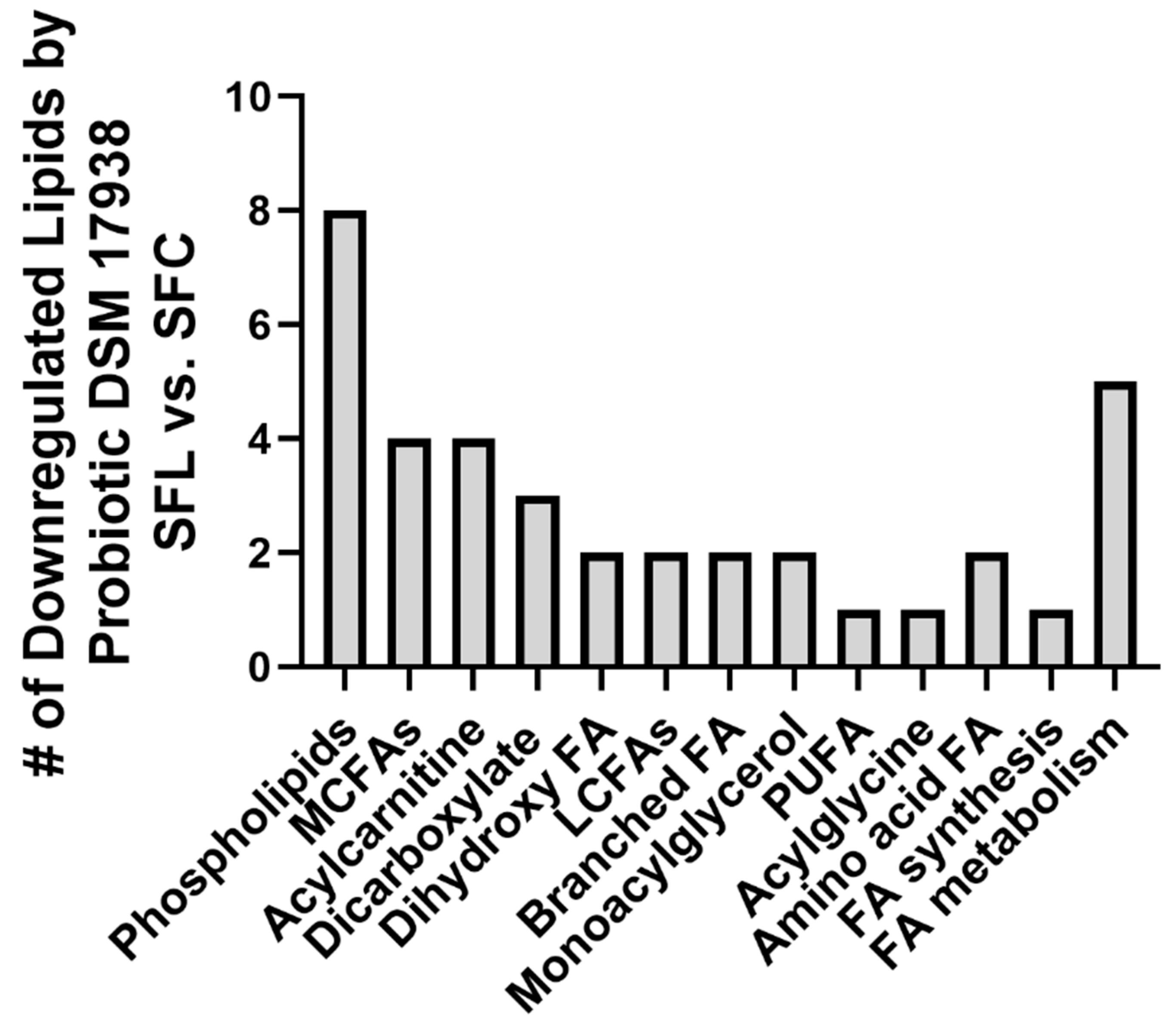
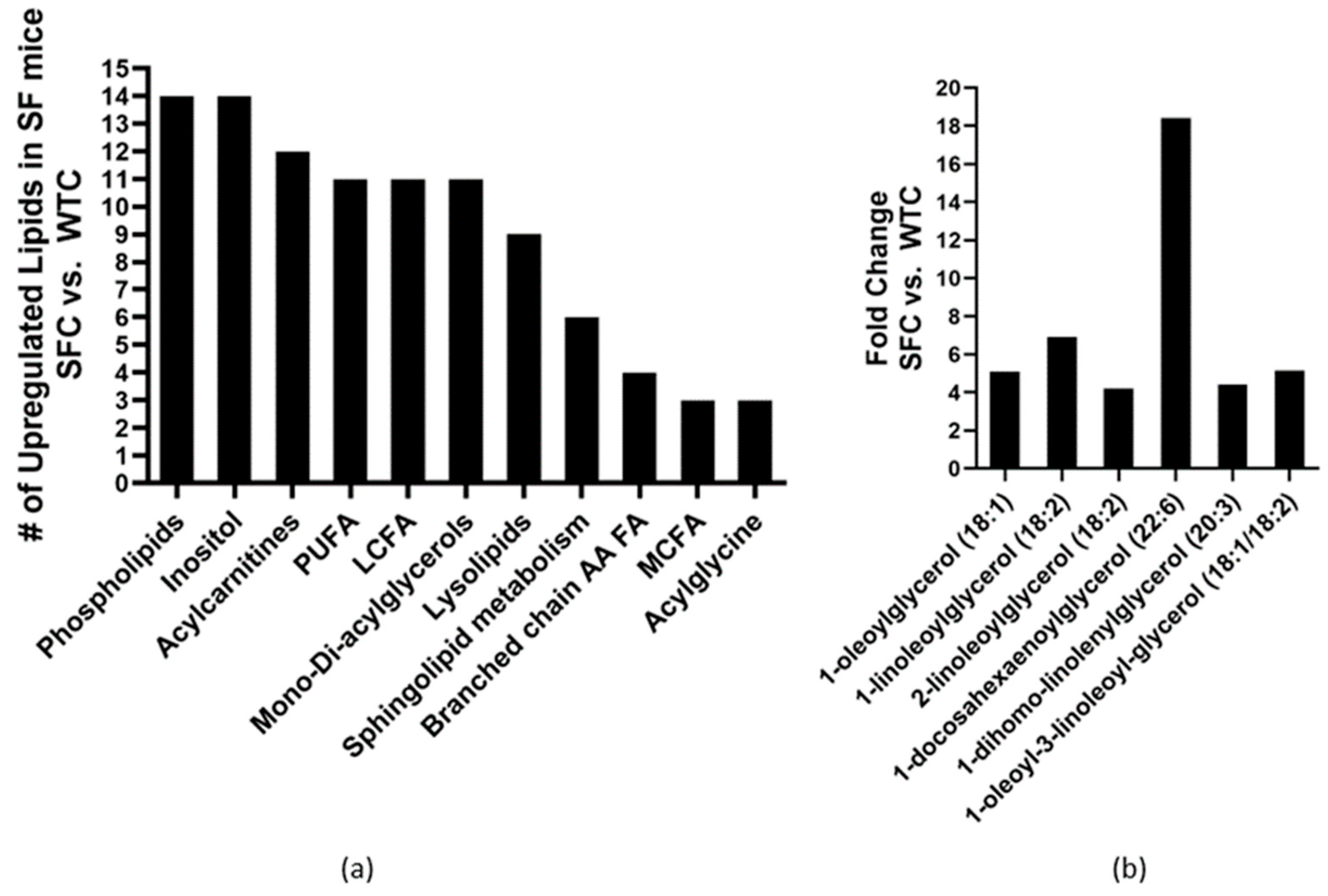
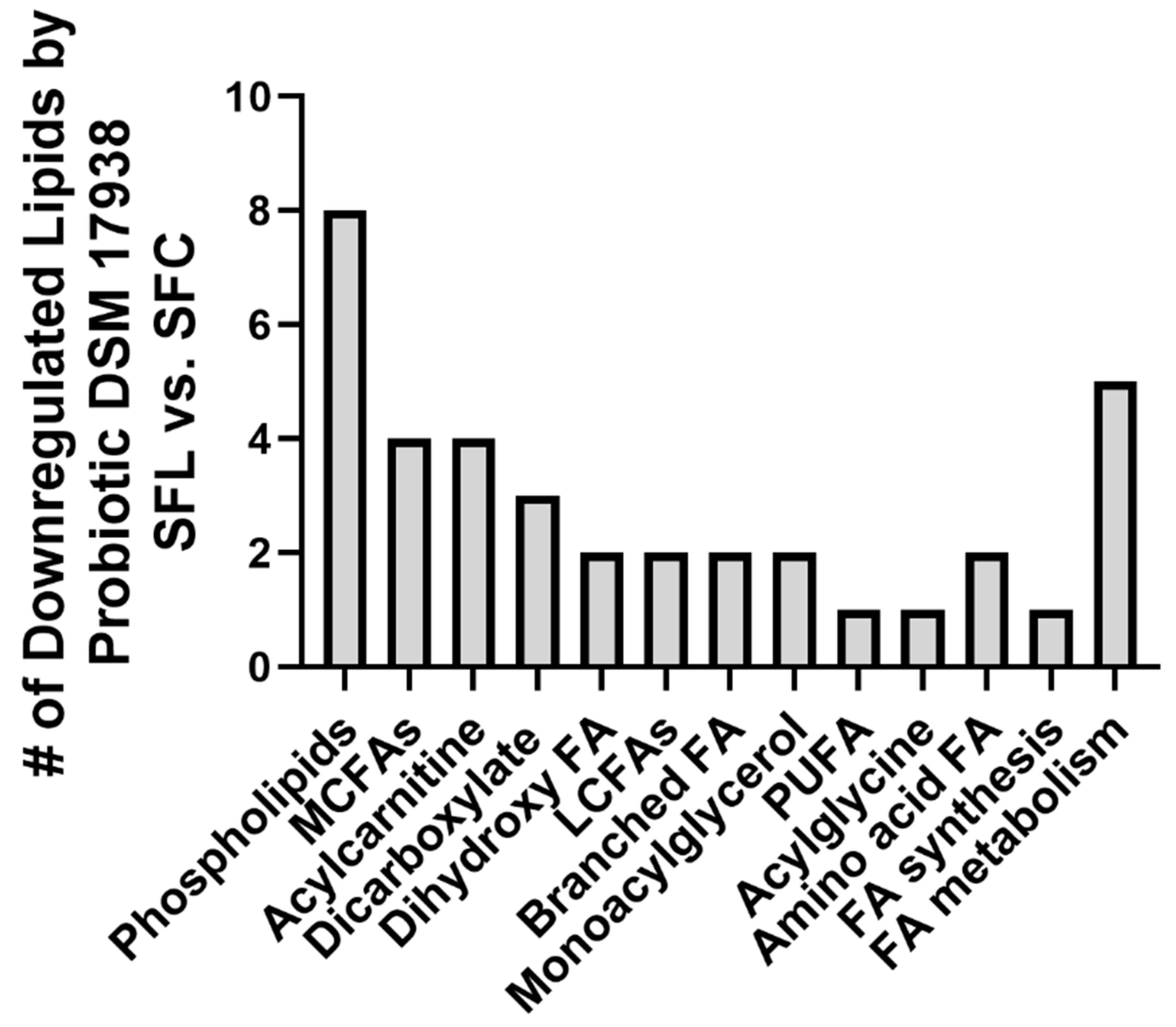
3.2. Probiotic DSM 17938 Modulates Plasma Lipid Profile in SF Mice
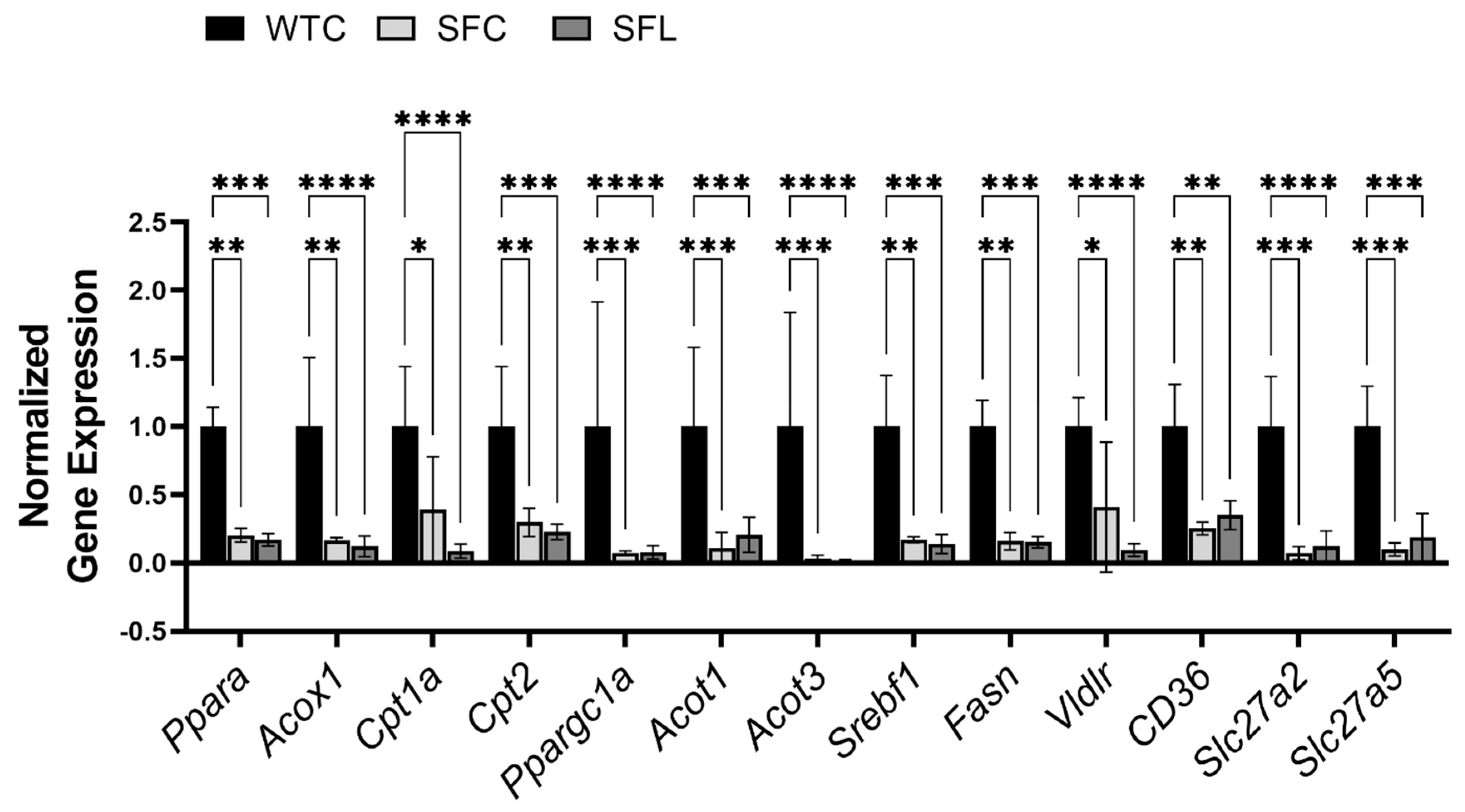
3.3. Foxp3+ Treg Deficiency Reduces Expression of Genes That Are Related to Lipid Metabolism in the Liver of SF Mice
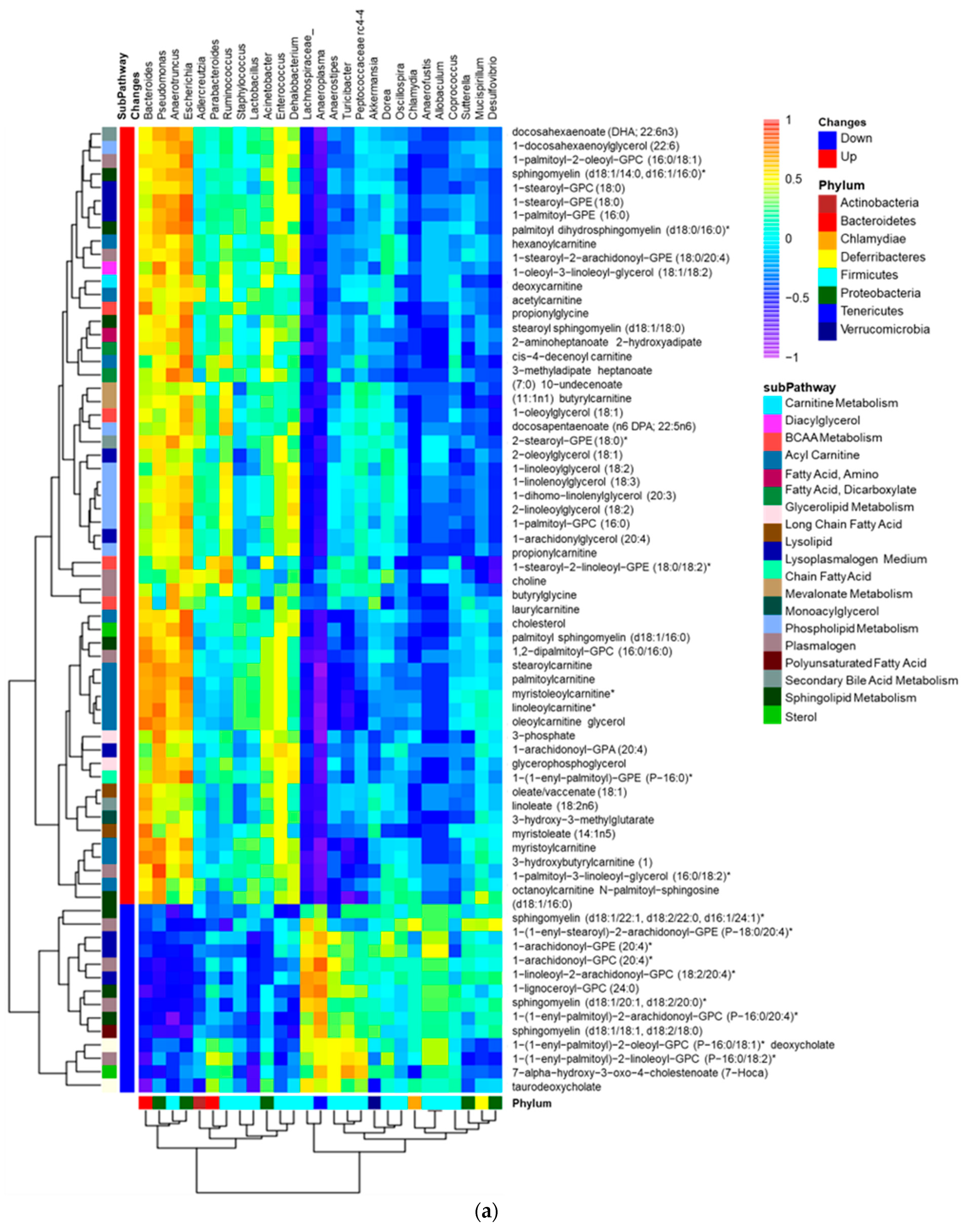
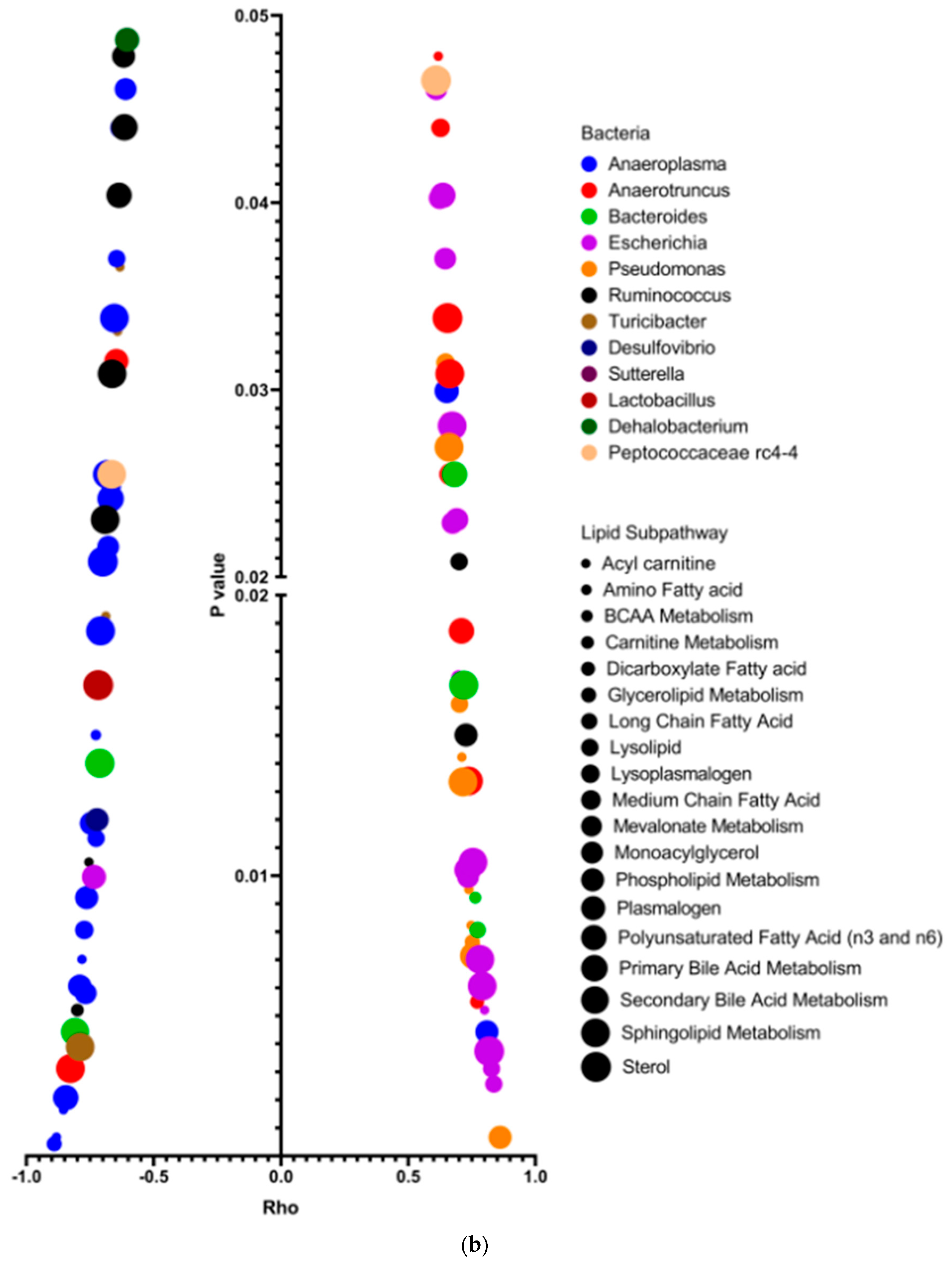
3.4. Altered Plasma Lipids Are Correlated with Gut Microbiota in SF Mice
3.5. Changed Plasma Lipids Are Positively Correlated with Plasma Th1-Associated (IFN-γ) and Th2-Associated (IL-4) Cytokine Levels
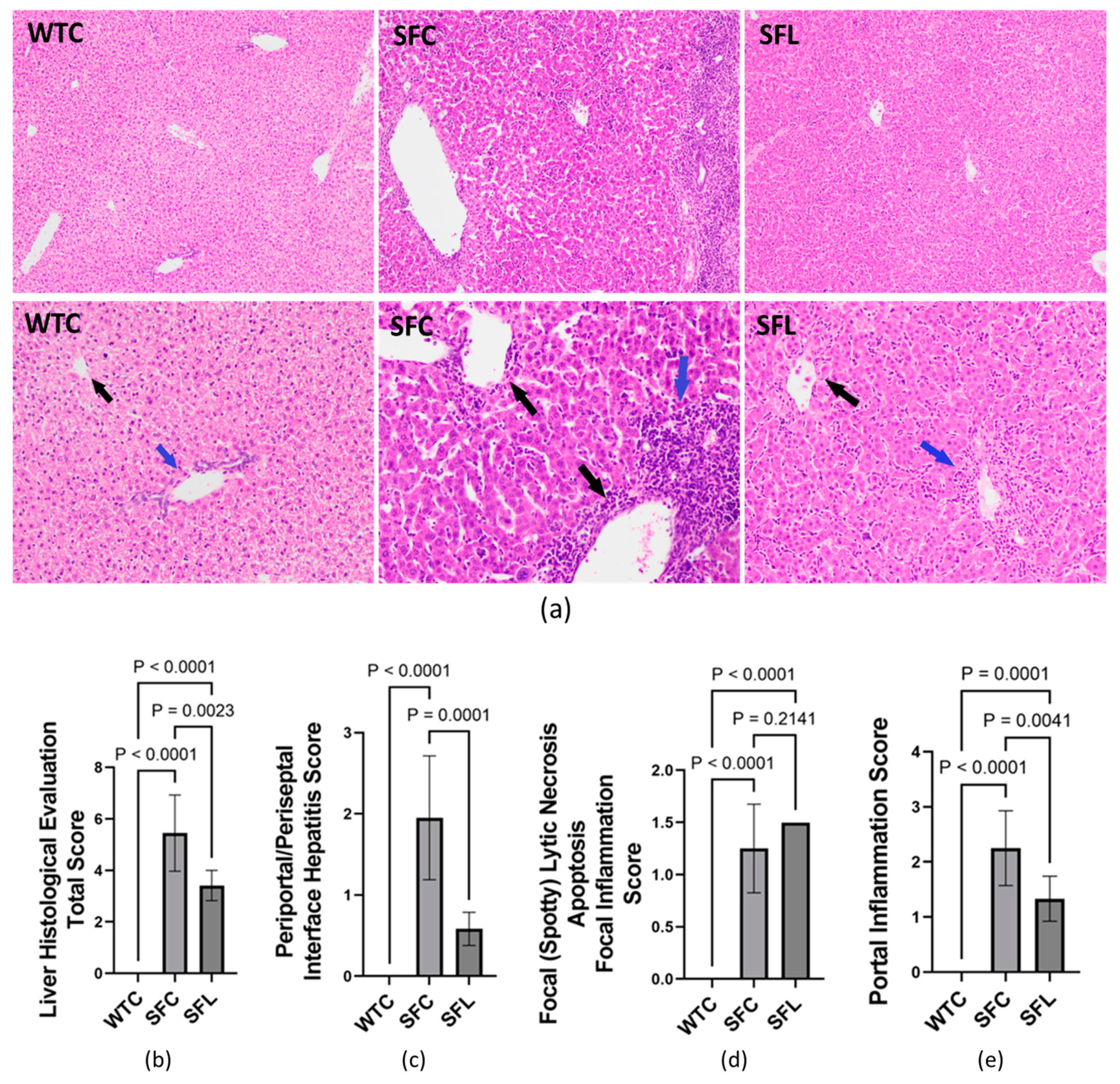
3.6. Foxp3+Treg Deficiency Induces Chronic Hepatitis and Can Be Partially Reduced through Intragastric Administration of DSM 17938
4. Discussion
4.1. Altered Lipid Metabolites in Mice and Humans with Treg-Deficiency Are Associated with Inflammation
4.2. Categories of Lipids That Are Increased in Autoimmune Diseases
4.3. Potential Role of PPARα in Dyslipidemia and Chronic Hepatitis in SF Mice
4.4. Potential Clinical Relevance of SF Mouse Chronic Hepatitis Phenotype
4.5. DSM 17938 Impacts Inflammation and Autoimmune-Associated Lipids through Modulating Gut Microbiota and Microbial-Associated Metabolites in SF Mice
4.6. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sakaguchi, S.; Mikami, N.; Wing, J.B.; Tanaka, A.; Ichiyama, K.; Ohkura, N. Regulatory T Cells and Human Disease. Annu. Rev. Immunol. 2020, 38, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Grover, P.; Goel, P.N.; Greene, M.I. Regulatory T Cells: Regulation of Identity and Function. Front. Immunol. 2021, 12, 750542. [Google Scholar] [CrossRef] [PubMed]
- Ono, M. Control of regulatory T-cell differentiation and function by T-cell receptor signalling and Foxp3 transcription factor complexes. Immunology 2020, 160, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, C.A. Transcriptional and translational control of Foxp3(+) regulatory T cell functional adaptation to inflammation. Curr. Opin. Immunol. 2020, 67, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Wildin, R.S.; Freitas, A. IPEX and FOXP3: Clinical and research perspectives. J. Autoimmun. 2005, 25, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Rasmussen, J.P.; Rudensky, A.Y. Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat. Immunol. 2007, 8, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Grover, P.; Goel, P.N.; Piccirillo, C.A.; Greene, M.I. FOXP3 and Tip60 Structural Interactions Relevant to IPEX Development Lead to Potential Therapeutics to Increase FOXP3 Dependent Suppressor T Cell Functions. Front. Pediatr. 2021, 9, 607292. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, R.; Roncarolo, M.G. IPEX syndrome from diagnosis to cure, learning along the way. J. Allergy Clin. Immunol. 2023; in press. [Google Scholar] [CrossRef]
- López, S.I.; Ciocca, M.; Oleastro, M.; Cuarterolo, M.L.; Rocca, A.; de Dávila, M.T.; Roy, A.; Fernández, M.C.; Nievas, E.; Bosaleh, A.; et al. Autoimmune hepatitis type 2 in a child with IPEX syndrome. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 690–693. [Google Scholar] [CrossRef]
- Suscovich, T.J.; Perdue, N.R.; Campbell, D.J. Type-1 immunity drives early lethality in scurfy mice. Eur. J. Immunol. 2012, 42, 2305–2310. [Google Scholar] [CrossRef]
- Haeberle, S.; Raker, V.; Haub, J.; Kim, Y.O.; Weng, S.Y.; Yilmaz, O.K.; Enk, A.; Steinbrink, K.; Schuppan, D.; Hadaschik, E.N. Regulatory T cell deficient scurfy mice exhibit a Th2/M2-like inflammatory response in the skin. J. Dermatol. Sci. 2017, 87, 285–291. [Google Scholar] [CrossRef]
- Vicari, E.; Haeberle, S.; Bolduan, V.; Ramcke, T.; Vorobyev, A.; Goletz, S.; Iwata, H.; Ludwig, R.J.; Schmidt, E.; Enk, A.H.; et al. Pathogenic Autoantibody Derived from Regulatory T Cell-Deficient Scurfy Mice Targets Type VII Collagen and Leads to Epidermolysis Bullosa Acquisita-Like Blistering Disease. J. Investig. Dermatol. 2022, 142, 980–984.e4. [Google Scholar] [CrossRef]
- Sharma, R.; Sharma, P.R.; Kim, Y.C.; Leitinger, N.; Lee, J.K.; Fu, S.M.; Ju, S.T. IL-2-controlled expression of multiple T cell trafficking genes and Th2 cytokines in the regulatory T cell-deficient scurfy mice: Implication to multiorgan inflammation and control of skin and lung inflammation. J. Immunol. 2011, 186, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, F.; Ziegler, S.F. FOXP3 and scurfy: How it all began. Nat. Rev. Immunol. 2014, 14, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Hadaschik, E.N.; Wei, X.; Leiss, H.; Heckmann, B.; Niederreiter, B.; Steiner, G.; Ulrich, W.; Enk, A.H.; Smolen, J.S.; Stummvoll, G.H. Regulatory T cell-deficient scurfy mice develop systemic autoimmune features resembling lupus-like disease. Arthritis Res. Ther. 2015, 17, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sharma, R.; Ju, S.T.; He, X.S.; Tao, Y.; Tsuneyama, K.; Tian, Z.; Lian, Z.X.; Fu, S.M.; Gershwin, M.E. Deficiency in regulatory T cells results in development of antimitochondrial antibodies and autoimmune cholangitis. Hepatology 2009, 49, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, K.; Haeberle, S.; Kim, Y.O.; Fritzler, M.J.; Weng, S.Y.; Goeppert, B.; Raker, V.K.; Steinbrink, K.; Schuppan, D.; Enk, A.; et al. Regulatory T-cell deficiency leads to features of autoimmune liver disease overlap syndrome in scurfy mice. Front. Immunol. 2023, 14, 1253649. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Hoang, T.K.; Wang, T.; Ferris, M.; Taylor, C.M.; Tian, X.; Luo, M.; Tran, D.Q.; Zhou, J.; Tatevian, N.; et al. Resetting microbiota by Lactobacillus reuteri inhibits T reg deficiency-induced autoimmunity via adenosine A2A receptors. J. Exp. Med. 2017, 214, 107–123. [Google Scholar] [CrossRef]
- Rosander, A.; Connolly, E.; Roos, S. Removal of antibiotic resistance gene-carrying plasmids from Lactobacillus reuteri ATCC 55730 and characterization of the resulting daughter strain, L. reuteri DSM 17938. Appl. Environ. Microbiol. 2008, 74, 6032–6040. [Google Scholar] [CrossRef]
- Gutierrez-Castrellon, P.; Indrio, F.; Bolio-Galvis, A.; Jimenez-Gutierrez, C.; Jimenez-Escobar, I.; Lopez-Velazquez, G. Efficacy of Lactobacillus reuteri DSM 17938 for infantile colic: Systematic review with network meta-analysis. Medicine 2017, 96, e9375. [Google Scholar] [CrossRef]
- Savino, F.; Cordisco, L.; Tarasco, V.; Palumeri, E.; Calabrese, R.; Oggero, R.; Roos, S.; Matteuzzi, D. Lactobacillus reuteri DSM 17938 in infantile colic: A randomized, double-blind, placebo-controlled trial. Pediatrics 2010, 126, e526–e533. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, A.; Kesavelu, D.; Kumar, K.; Chatterjee, P.; Jog, P.; Gopalan, S.; Paul, R.; Veligandla, K.C.; Mehta, S.; Mane, A.; et al. Role of Lactobacillus reuteri DSM 17938 on Crying Time Reduction in Infantile Colic and Its Impact on Maternal Depression: A Real-Life Clinic-Based Study. Clin Pract. 2022, 12, 37–45. [Google Scholar] [CrossRef]
- Gutiérrez Escárate, C.; Bustos Medina, L.; Caniulao Ríos, K.; Taito Antivil, C.; Gallegos Casanova, Y.; Silva Beltrán, C. Probiotic intervention to prevent necrotizing enterocolitis in extremely preterm infants born before 32 weeks of gestation or with a birth weight of less than 1500 g. Arch. Argent Pediatr. 2021, 119, 185–191. [Google Scholar] [PubMed]
- Hoang, T.K.; He, B.; Wang, T.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Protective effect of Lactobacillus reuteri DSM 17938 against experimental necrotizing enterocolitis is mediated by Toll-like receptor 2. Am. J. Physiol.-Gastrointest. Liver Physiol. 2018, 315, G231–G240. [Google Scholar] [CrossRef]
- Liu, Y.; Hoang, T.K.; Taylor, C.M.; Park, E.S.; Freeborn, J.; Luo, M.; Roos, S.; Rhoads, J.M. Limosilactobacillus reuteri and Lacticaseibacillus rhamnosus GG differentially affect gut microbes and metabolites in mice with Treg deficiency. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 320, G969–G981. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, X.; Daniel, R.C.; Okeugo, B.; Armbrister, S.A.; Luo, M.; Taylor, C.M.; Wu, G.; Rhoads, J.M. Impact of probiotic Limosilactobacillus reuteri DSM 17938 on amino acid metabolism in the healthy newborn mouse. Amino Acids 2022, 54, 1383–1401. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Hoang, T.K.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Adenosine A2A Receptor Deletion Blocks the Beneficial Effects of Lactobacillus reuteri in Regulatory T-Deficient Scurfy Mice. Front. Immunol. 2017, 8, 1680. [Google Scholar] [CrossRef] [PubMed]
- Parhofer, K.G. Interaction between Glucose and Lipid Metabolism: More than Diabetic Dyslipidemia. Diabetes Metab. J. 2015, 39, 353–362. [Google Scholar] [CrossRef]
- Jorgensen, S.F.; Macpherson, M.E.; Skarpengland, T.; Berge, R.K.; Fevang, B.; Halvorsen, B.; Aukrust, P. Disturbed lipid profile in common variable immunodeficiency—A pathogenic loop of inflammation and metabolic disturbances. Front. Immunol. 2023, 14, 1199727. [Google Scholar] [CrossRef]
- Cas, M.D.; Roda, G.; Li, F.; Secundo, F. Functional Lipids in Autoimmune Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 3074. [Google Scholar] [CrossRef]
- Ko, C.W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Poggioli, R.; Hirani, K.; Jogani, V.G.; Ricordi, C. Modulation of inflammation and immunity by omega-3 fatty acids: A possible role for prevention and to halt disease progression in autoimmune, viral, and age-related disorders. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 7380–7400. [Google Scholar] [PubMed]
- Wang, Z.; Koonen, D.; Hofker, M.; Fu, J. Gut microbiome and lipid metabolism: From associations to mechanisms. Curr. Opin. Lipidol. 2016, 27, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Clardy, J.; Xavier, R.J. Gut microbiome lipid metabolism and its impact on host physiology. Cell Host Microbe 2023, 31, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Freeborn, J.; Armbrister, S.A.; Tran, D.Q.; Rhoads, J.M. Treg-associated monogenic autoimmune disorders and gut microbial dysbiosis. Pediatr. Res. 2022, 91, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Skarpengland, T.; Macpherson, M.E.; Hov, J.R.; Kong, X.Y.; Bohov, P.; Halvorsen, B.; Fevang, B.; Berge, R.K.; Aukrust, P.; Jørgensen, S.F. Altered Plasma Fatty Acids Associate with Gut Microbial Composition in Common Variable Immunodeficiency. J. Clin. Immunol. 2022, 42, 146–157. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Liu, Y.; Hoang, T.K.; Wang, T.; He, B.; Tran, D.Q.; Zhou, J.; Tatevian, N.; Rhoads, J.M. Circulating L-selectin expressing-T cell subsets correlate with the severity of Foxp3 deficiency autoimmune disease. Int. J. Clin. Exp. Pathol. 2016, 9, 899–909. [Google Scholar]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Lactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-kappaB signaling in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G608–G617. [Google Scholar] [CrossRef]
- Sung, V.; D’Amico, F.; Cabana, M.D.; Chau, K.; Koren, G.; Savino, F.; Szajewska, H.; Deshpande, G.; Dupont, C.; Indrio, F.; et al. Lactobacillus reuteri to Treat Infant Colic: A Meta-analysis. Pediatrics 2018, 141, e20171811. [Google Scholar] [CrossRef]
- Savino, F.; Garro, M.; Montanari, P.; Galliano, I.; Bergallo, M. Crying Time and RORγ/FOXP3 Expression in Lactobacillus reuteri DSM17938-Treated Infants with Colic: A Randomized Trial. J. Pediatr. 2018, 192, 171–177.e1. [Google Scholar] [CrossRef]
- Liu, Y.; Tran, D.Q.; Fatheree, N.Y.; Marc Rhoads, J. Lactobacillus reuteri DSM 17938 differentially modulates effector memory T cells and Foxp3+ regulatory T cells in a mouse model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G177–G186. [Google Scholar] [CrossRef]
- Liu, Y.; Armbrister, S.A.; Okeugo, B.; Mills, T.W.; Daniel, R.C.; Oh, J.H.; van Pijkeren, J.P.; Park, E.S.; Saleh, Z.M.; Lahiri, S.; et al. Probiotic-Derived Ecto-5′-Nucleotidase Produces Anti-Inflammatory Adenosine Metabolites in Treg-Deficient Scurfy Mice. Probiotics Antimicrob. Proteins 2023, 15, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glockner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Nakaji, M.; Hayashi, Y.; Ninomiya, T.; Yano, Y.; Yoon, S.; Seo, Y.; Nagano, H.; Komori, H.; Hashimoto, K.; Orino, A.; et al. Histological grading and staging in chronic hepatitis: Its practical correlation. Pathol. Int. 2002, 52, 683–690. [Google Scholar] [CrossRef]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.W.; Staels, B.; De Bosscher, K. Molecular Actions of PPARα in Lipid Metabolism and Inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [PubMed]
- Aregger, M.; Lawson, K.A.; Billmann, M.; Costanzo, M.; Tong, A.H.Y.; Chan, K.; Rahman, M.; Brown, K.R.; Ross, C.; Usaj, M.; et al. Systematic mapping of genetic interactions for de novo fatty acid synthesis identifies C12orf49 as a regulator of lipid metabolism. Nat. Metab. 2020, 2, 499–513. [Google Scholar] [CrossRef]
- Son, N.H.; Basu, D.; Samovski, D.; Pietka, T.A.; Peche, V.S.; Willecke, F.; Fang, X.; Yu, S.Q.; Scerbo, D.; Chang, H.R.; et al. Endothelial cell CD36 optimizes tissue fatty acid uptake. J. Clin. Investig. 2018, 128, 4329–4342. [Google Scholar] [CrossRef]
- Samovski, D.; Jacome-Sosa, M.; Abumrad, N.A. Fatty Acid Transport and Signaling: Mechanisms and Physiological Implications. Annu. Rev. Physiol. 2023, 85, 317–337. [Google Scholar] [CrossRef]
- Preidis, G.A.; Kim, K.H.; Moore, D.D. Nutrient-sensing nuclear receptors PPARα and FXR control liver energy balance. J. Clin. Investig. 2017, 127, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive lipids, inflammation and chronic diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ochoa, Á.; Brunius, C.; Borrás-Linares, I.; Quirantes-Piné, R.; Cádiz-Gurrea, M.L.; Precisesads Clinical, C.; Alarcón Riquelme, M.E.; Segura-Carretero, A. Metabolic Disturbances in Urinary and Plasma Samples from Seven Different Systemic Autoimmune Diseases Detected by HPLC-ESI-QTOF-MS. J. Proteome Res. 2020, 19, 3220–3229. [Google Scholar] [CrossRef]
- Ben-Skowronek, I. IPEX Syndrome: Genetics and Treatment Options. Genes 2021, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Gorczyca, D.; Szponar, B.; Paściak, M.; Czajkowska, A.; Szmyrka, M. Serum levels of n-3 and n-6 polyunsaturated fatty acids in patients with systemic lupus erythematosus and their association with disease activity: A pilot study. Scand. J. Rheumatol. 2022, 51, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bi, X.; Wang, S.; Zhang, Z.; Li, F.; Zhao, A.Z. Therapeutic Potential of ω-3 Polyunsaturated Fatty Acids in Human Autoimmune Diseases. Front. Immunol. 2019, 10, 2241. [Google Scholar] [CrossRef] [PubMed]
- Dambrova, M.; Makrecka-Kuka, M.; Kuka, J.; Vilskersts, R.; Nordberg, D.; Attwood, M.M.; Smesny, S.; Sen, Z.D.; Guo, A.C.; Oler, E.; et al. Acylcarnitines: Nomenclature, Biomarkers, Therapeutic Potential, Drug Targets, and Clinical Trials. Pharmacol. Rev. 2022, 74, 506–551. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, F.; Guo, Y.; Lu, Y.; Ji, W.; Lin, L.; Chen, W.; Xu, T.; Kong, D.; Shen, Q.; et al. Untargeted lipidomics reveals specific lipid abnormalities in systemic lupus erythematosus. Clin. Exp. Rheumatol. 2022, 40, 1011–1018. [Google Scholar] [CrossRef]
- Fernández-Ochoa, Á.; Quirantes-Piné, R.; Borrás-Linares, I.; Gemperline, D.; Alarcón Riquelme, M.E.; Beretta, L.; Segura-Carretero, A. Urinary and plasma metabolite differences detected by HPLC-ESI-QTOF-MS in systemic sclerosis patients. J. Pharm. Biomed. Anal. 2019, 162, 82–90. [Google Scholar] [CrossRef]
- Alexandropoulou, I.; Grammatikopoulou, M.G.; Gkouskou, K.K.; Pritsa, A.A.; Vassilakou, T.; Rigopoulou, E.; Lindqvist, H.M.; Bogdanos, D.P. Ceramides in Autoimmune Rheumatic Diseases: Existing Evidence and Therapeutic Considerations for Diet as an Anticeramide Treatment. Nutrients 2023, 15, 229. [Google Scholar] [CrossRef]
- Pedersen, K.; Ipsen, D.H.; Skat-Rørdam, J.; Lykkesfeldt, J.; Tveden-Nyborg, P. Dietary Long-Chain Fatty Acids Accelerate Metabolic Dysfunction in Guinea Pigs with Non-Alcoholic Steatohepatitis. Nutrients 2023, 15, 2445. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, N.S.; Nazli, R.; Zafar, H.; Fatima, S. Effects of lipid based Multiple Micronutrients Supplement on the birth outcome of underweight pre-eclamptic women: A randomized clinical trial. Pak. J. Med. Sci. 2022, 38, 219–226. [Google Scholar] [PubMed]
- He, B.; Hoang, T.K.; Tian, X.; Taylor, C.M.; Blanchard, E.; Luo, M.; Bhattacharjee, M.B.; Freeborn, J.; Park, S.; Couturier, J.; et al. Lactobacillus reuteri Reduces the Severity of Experimental Autoimmune Encephalomyelitis in Mice by Modulating Gut Microbiota. Front. Immunol. 2019, 10, 385. [Google Scholar] [CrossRef]
- Mirza, A.Z.; Althagafi, I.I.; Shamshad, H. Role of PPAR receptor in different diseases and their ligands: Physiological importance and clinical implications. Eur. J. Med. Chem. 2019, 166, 502–513. [Google Scholar] [CrossRef]
- Tahri-Joutey, M.; Andreoletti, P.; Surapureddi, S.; Nasser, B.; Cherkaoui-Malki, M.; Latruffe, N. Mechanisms Mediating the Regulation of Peroxisomal Fatty Acid Beta-Oxidation by PPARα. Int. J. Mol. Sci. 2021, 22, 8969. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.E.; Gordon, D.M.; Hipp, J.A.; Hong, S.; Mitchell, Z.L.; Franco, N.R.; Robison, J.W.; Anderson, C.D.; Stec, D.F.; Hinds, T.D., Jr. Loss of hepatic PPARα promotes inflammation and serum hyperlipidemia in diet-induced obesity. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2019, 317, R733–R745. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; Lopez-Gambero, A.J.; Serrano, A.; Pavon, F.J.; Baixeras, E.; Rodriguez de Fonseca, F.; Suarez, J. Peroxisome Proliferator-Activated Receptors: Experimental Targeting for the Treatment of Inflammatory Bowel Diseases. Front. Pharmacol. 2020, 11, 730. [Google Scholar] [CrossRef]
- Grau, R.; Punzón, C.; Fresno, M.; Iñiguez, M.A. Peroxisome-proliferator-activated receptor alpha agonists inhibit cyclo-oxygenase 2 and vascular endothelial growth factor transcriptional activation in human colorectal carcinoma cells via inhibition of activator protein-1. Biochem. J. 2006, 395, 81–88. [Google Scholar] [CrossRef]
- Chang, H.; Zhao, F.; Xie, X.; Liao, Y.; Song, Y.; Liu, C.; Wu, Y.; Wang, Y.; Liu, D.; Wang, Y.; et al. PPARα suppresses Th17 cell differentiation through IL-6/STAT3/RORγt pathway in experimental autoimmune myocarditis. Exp. Cell Res. 2019, 375, 22–30. [Google Scholar] [CrossRef]
- Depommier, C.; Vitale, R.M.; Iannotti, F.A.; Silvestri, C.; Flamand, N.; Druart, C.; Everard, A.; Pelicaen, R.; Maiter, D.; Thissen, J.P.; et al. Beneficial Effects of Akkermansia muciniphila Are Not Associated with Major Changes in the Circulating Endocannabinoidome but Linked to Higher Mono-Palmitoyl-Glycerol Levels as New PPARα Agonists. Cells 2021, 10, 185. [Google Scholar] [CrossRef]
- Chen, J.; Liu, W.; Zhu, W. Foxp3(+) Treg Cells Are Associated with Pathological Process of Autoimmune Hepatitis by Activating Methylation Modification in Autoimmune Hepatitis Patients. Med. Sci. Monit. 2019, 25, 6204–6212. [Google Scholar] [CrossRef] [PubMed]
- Agakidis, C.; Agakidou, E.; Sarafidis, K.; Papoulidis, I.; Xinias, I.; Farmaki, E. Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome Associated with a Novel Mutation of FOXP3 Gene. Front. Pediatr. 2019, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Chung, H.S.; Lee, K.; Lee, H.; Kim, M.; Bae, H. Curcumin attenuates the scurfy-induced immune disorder, a model of IPEX syndrome, with inhibiting Th1/Th2/Th17 responses in mice. Phytomedicine 2017, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Belvoncikova, P.; Maronek, M.; Gardlik, R. Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 10729. [Google Scholar] [CrossRef] [PubMed]
- De Luca, F.; Shoenfeld, Y. The microbiome in autoimmune diseases. Clin. Exp. Immunol. 2019, 195, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Alookaran, J.J.; Rhoads, J.M. Probiotics in Autoimmune and Inflammatory Disorders. Nutrients 2018, 10, 1537. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tran, D.Q.; Rhoads, J.M. Probiotics in Disease Prevention and Treatment. J. Clin. Pharmacol. 2018, 58 (Suppl. 10), S164–S179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hoang, T.K.; Park, E.S.; Freeborn, J.; Okeugo, B.; Tran, D.Q.; Rhoads, J.M. Probiotic-educated Tregs are more potent than naive Tregs for immune tolerance in stressed new-born mice. Benef. Microbes 2023, 14, 73–84. [Google Scholar] [CrossRef]
- Da, M.; Chen, L.; Enk, A.; Ring, S.; Mahnke, K. The Multifaceted Actions of CD73 During Development and Suppressive Actions of Regulatory T Cells. Front. Immunol. 2022, 13, 914799. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, X.; He, B.; Hoang, T.K.; Taylor, C.M.; Blanchard, E.; Freeborn, J.; Park, S.; Luo, M.; Couturier, J.; et al. Lactobacillus reuteri DSM 17938 feeding of healthy newborn mice regulates immune responses while modulating gut microbiota and boosting beneficial metabolites. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G824–G838. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef]
- Liu, X.; Jin, G.; Tang, Q.; Huang, S.; Zhang, Y.; Sun, Y.; Liu, T.; Guo, Z.; Yang, C.; Wang, B.; et al. Early life Lactobacillus rhamnosus GG colonisation inhibits intestinal tumour formation. Br. J. Cancer 2022, 126, 1421–1431. [Google Scholar] [CrossRef]
- Chan, Y.K.; Brar, M.S.; Kirjavainen, P.V.; Chen, Y.; Peng, J.; Li, D.; Leung, F.C.; El-Nezami, H. High fat diet induced atherosclerosis is accompanied with low colonic bacterial diversity and altered abundances that correlates with plaque size, plasma A-FABP and cholesterol: A pilot study of high fat diet and its intervention with Lactobacillus rhamnosus GG (LGG) or telmisartan in ApoE(-/-) mice. BMC Microbiol. 2016, 16, 264. [Google Scholar]
- Wang, T.; Sternes, P.R.; Guo, X.K.; Zhao, H.; Xu, C.; Xu, H. Autoimmune diseases exhibit shared alterations in the gut microbiota. Rheumatology 2023, Kead364. [Google Scholar] [CrossRef] [PubMed]
- Suciu, A.; Abenavoli, L.; Pellicano, R.; Luzza, F.; Dumitrascu, D.L. Transaminases: Oldies but goldies. A narrative review. Minerva Gastroenterol. Dietol. 2020, 66, 246–251. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Pathway | Lipid Biochemical | R Value | p Value |
|---|---|---|---|
| Lysolipid | 1-linoleoyl-GPA (18:2) | 0.94 | 0.015 |
| Medium-Chain Fatty Acid | caprylate (8:0) | 0.89 | 0.033 |
| Sphingolipid metabolism | stearoyl sphingomyelin (d18:1/18:0) | 0.89 | 0.033 |
| Primary Bile Acid Metabolism | taurochenodeoxycholate | 0.94 | 0.017 |
| Secondary Bile Acid Metabolism | taurohyodeoxycholic acid | 0.93 | 0.007 |
| Lipid Pathway | Lipid Biochemical | R Value | p Value |
|---|---|---|---|
| Lysolipid | 1-(1-enyl-oleoyl)-GPE (P-18:1) | 0.94 | 0.017 |
| 1-stearoyl-GPE (18:0) | 0.89 | 0.033 | |
| 1-arachidonoyl-GPA (20:4) | 0.99 | 0.0003 | |
| Phospholipid Metabolism | 1-palmitoyl-2-oleoyl-GPC (16:0/18:1) | 0.89 | 0.033 |
| Long-Chain Fatty Acid | eicosenoate (20:1) | 0.94 | 0.017 |
| erucate (22:1n9) | 0.94 | 0.017 | |
| Polyunsaturated Fatty Acid | adrenate (22:4n6) | 0.94 | 0.017 |
| Lysoplasmalogen | 1-(1-enyl-palmitoyl)-GPE (P-16:0) | 0.94 | 0.017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nessim Kostandy, E.; Suh, J.H.; Tian, X.; Okeugo, B.; Rubin, E.; Shirai, S.; Luo, M.; Taylor, C.M.; Kim, K.H.; Rhoads, J.M.; et al. Probiotic Limosilactobacillus reuteri DSM 17938 Changes Foxp3 Deficiency-Induced Dyslipidemia and Chronic Hepatitis in Mice. Nutrients 2024, 16, 511. https://doi.org/10.3390/nu16040511
Nessim Kostandy E, Suh JH, Tian X, Okeugo B, Rubin E, Shirai S, Luo M, Taylor CM, Kim KH, Rhoads JM, et al. Probiotic Limosilactobacillus reuteri DSM 17938 Changes Foxp3 Deficiency-Induced Dyslipidemia and Chronic Hepatitis in Mice. Nutrients. 2024; 16(4):511. https://doi.org/10.3390/nu16040511
Chicago/Turabian StyleNessim Kostandy, Erini, Ji Ho Suh, Xiangjun Tian, Beanna Okeugo, Erin Rubin, Sara Shirai, Meng Luo, Christopher M. Taylor, Kang Ho Kim, J. Marc Rhoads, and et al. 2024. "Probiotic Limosilactobacillus reuteri DSM 17938 Changes Foxp3 Deficiency-Induced Dyslipidemia and Chronic Hepatitis in Mice" Nutrients 16, no. 4: 511. https://doi.org/10.3390/nu16040511
APA StyleNessim Kostandy, E., Suh, J. H., Tian, X., Okeugo, B., Rubin, E., Shirai, S., Luo, M., Taylor, C. M., Kim, K. H., Rhoads, J. M., & Liu, Y. (2024). Probiotic Limosilactobacillus reuteri DSM 17938 Changes Foxp3 Deficiency-Induced Dyslipidemia and Chronic Hepatitis in Mice. Nutrients, 16(4), 511. https://doi.org/10.3390/nu16040511