
Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic

 ,
, 
 , ,
, , 


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical and Standards
2.2. ST-65 Formulation
2.3. UHPLC-HRMS/MS Analysis
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. Exposure of SH-SY5Y to STRESSLESS (ST-65) Formulation
2.7. Sample Extraction
2.8. NMR Spectra Acquisition
2.9. NMR Spectra Analysis
3. Results
3.1. UHPLC-HRMS/MS Analysis
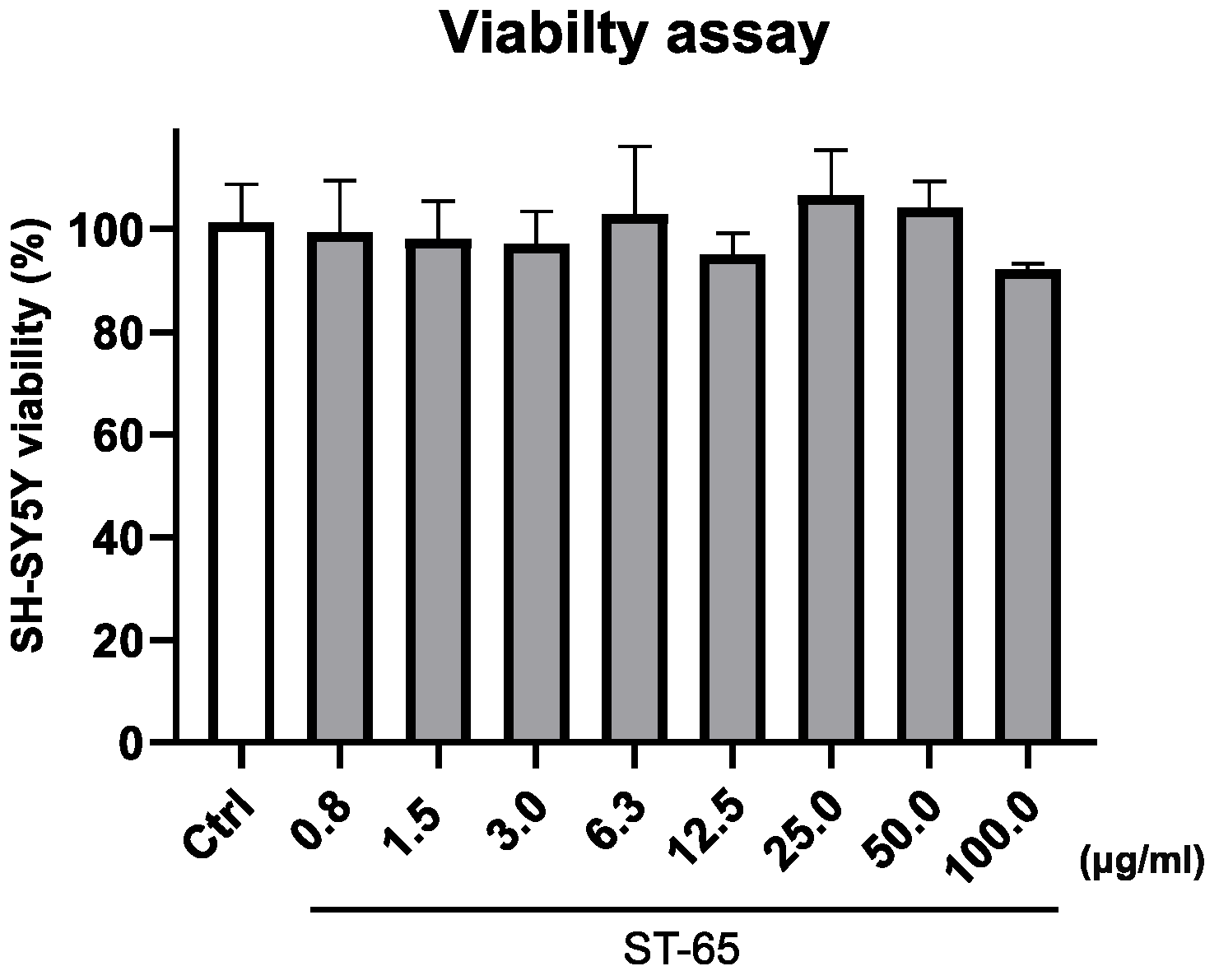
3.2. ST-65 Viability
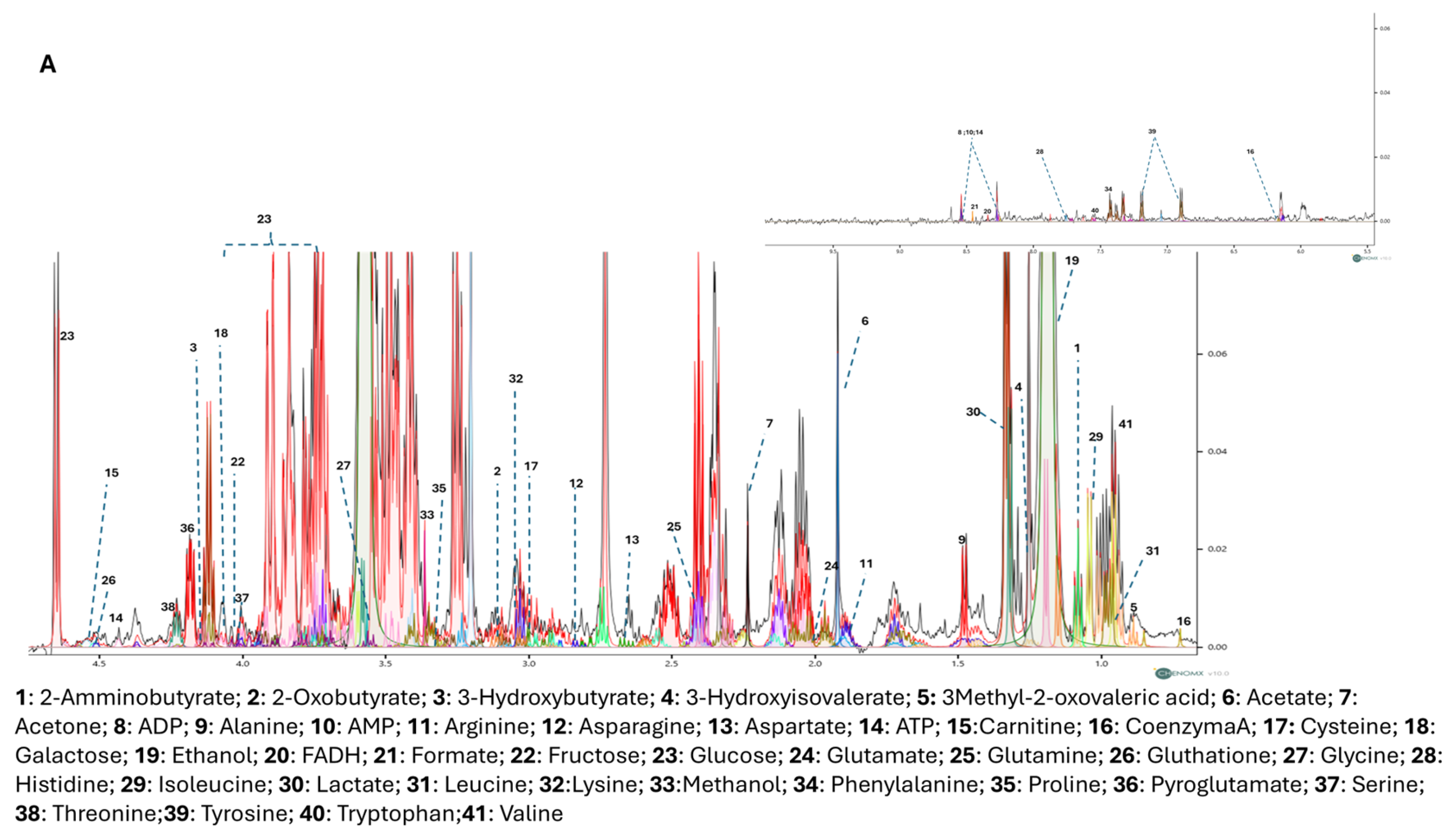
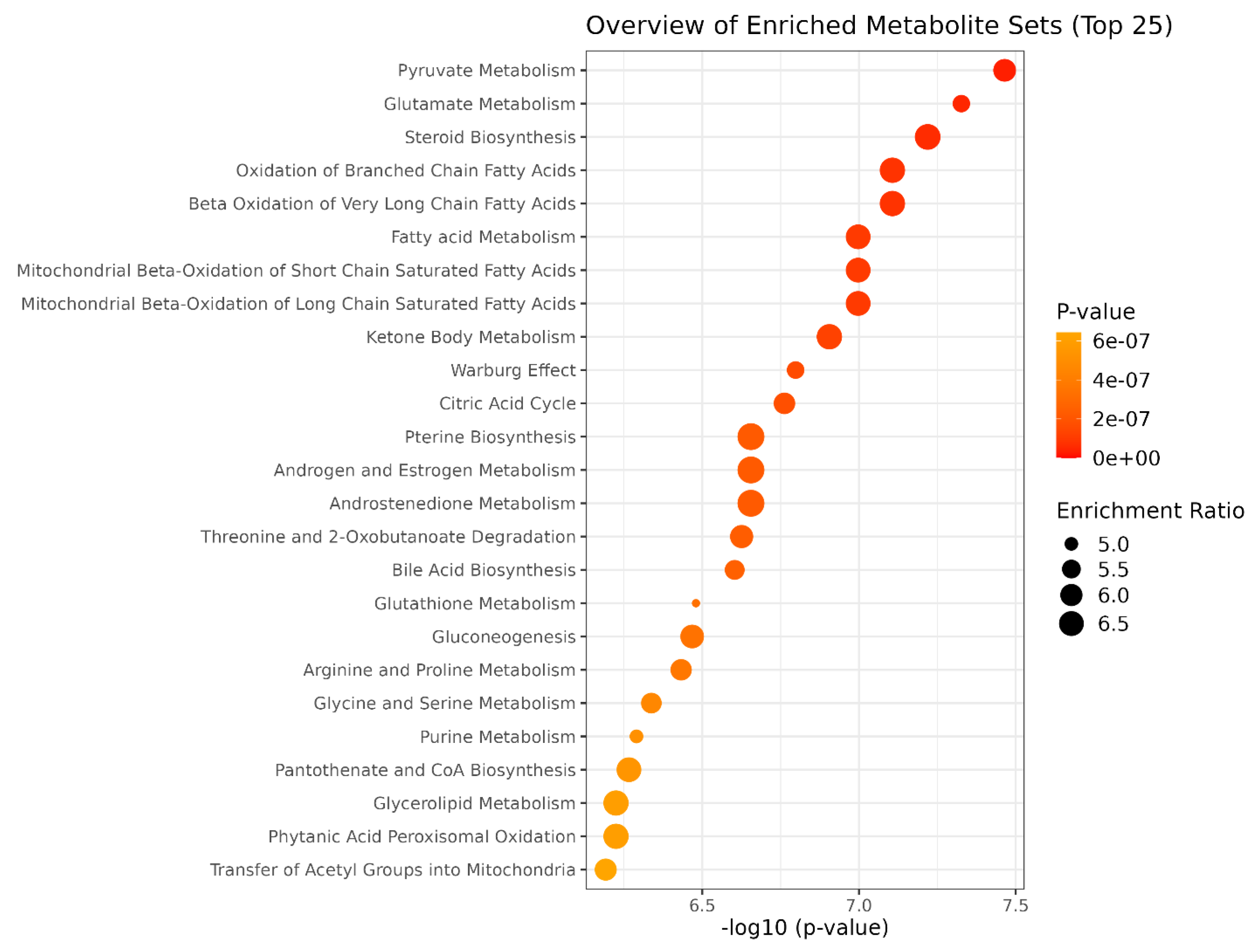
3.3. NMR Metabolomic
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The Role of Cortisol in Chronic Stress, Neurodegenerative Diseases, and Psychological Disorders. Cells 2023, 12, 2726. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.I.; Wu, A.M.S. Assessing the Role of Cortisol in Anxiety, Major Depression, and Neuroticism: A Mendelian Randomization Study Using SERPINA6/SERPINA1 Variants. Biol. Psychiatry Glob. Open Sci. 2024, 4, 100294. [Google Scholar] [CrossRef] [PubMed]
- Kuckuck, S.; van der Valk, E.S.; Scheurink, A.J.W.; van der Voorn, B.; Iyer, A.M.; Visser, J.A.; Delhanty, P.J.D.; van den Berg, S.A.A.; van Rossum, E.F.C. Glucocorticoids, stress and eating: The mediating role of appetite-regulating hormones. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2023, 24, e13539. [Google Scholar] [CrossRef] [PubMed]
- Passos, G.S.; Youngstedt, S.D.; Rozales, A.; Ferreira, W.S.; De-Assis, D.E.; De-Assis, B.P.; Santana, M.G. Insomnia Severity is Associated with Morning Cortisol and Psychological Health. Sleep Sci. 2023, 16, 92–96. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, C.; Wang, S.; Tao, S.; Liao, Y.; Shi, Z.; Tang, Z.; Wu, Y.; Liu, Y.; Yang, P. Cortisol synergizing with endoplasmic reticulum stress induces regulatory T-cell dysfunction. Immunology 2023, 170, 334–343. [Google Scholar] [CrossRef]
- Kip, E.; Parr-Brownlie, L.C. Healthy lifestyles and wellbeing reduce neuroinflammation and prevent neurodegenerative and psychiatric disorders. Front. Neurosci. 2023, 17, 1092537. [Google Scholar] [CrossRef]
- Mikulska, P.; Malinowska, M.; Ignacyk, M.; Szustowski, P.; Nowak, J.; Pesta, K.; Szeląg, M.; Szklanny, D.; Judasz, E.; Kaczmarek, G.; et al. Ashwagandha (Withania somnifera)-Current Research on the Health-Promoting Activities: A Narrative Review. Pharmaceutics 2023, 15, 1057. [Google Scholar] [CrossRef]
- Cheah, K.L.; Norhayati, M.N.; Husniati Yaacob, L.; Abdul Rahman, R. Effect of Ashwagandha (Withania somnifera) extract on sleep: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0257843. [Google Scholar] [CrossRef]
- Leonard, M.; Dickerson, B.; Estes, L.; Gonzalez, D.E.; Jenkins, V.; Johnson, S.; Xing, D.; Yoo, C.; Ko, J.; Purpura, M.; et al. Acute and Repeated Ashwagandha Supplementation Improves Markers of Cognitive Function and Mood. Nutrients 2024, 16, 1813. [Google Scholar] [CrossRef]
- Speers, A.B.; Cabey, K.A.; Soumyanath, A.; Wright, K.M. Effects of Withania somnifera (Ashwagandha) on Stress and the Stress- Related Neuropsychiatric Disorders Anxiety, Depression, and Insomnia. Curr. Neuropharmacol. 2021, 19, 1468–1495. [Google Scholar] [CrossRef]
- Guo, S.; Rezaei, M.J. The benefits of ashwagandha (Withania somnifera) supplements on brain function and sports performance. Front. Nutr. 2024, 11, 1439294. [Google Scholar] [CrossRef]
- Bonilla, D.A.; Moreno, Y.; Gho, C.; Petro, J.L.; Odriozola-Martínez, A.; Kreider, R.B. Effects of Ashwagandha (Withania somnifera) on Physical Performance: Systematic Review and Bayesian Meta-Analysis. J. Funct. Morphol. Kinesiol. 2021, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Pletzer, B.; Poppelaars, E.S.; Klackl, J.; Jonas, E. The gonadal response to social stress and its relationship to cortisol. Stress 2021, 24, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Lopresti, A.L.; Fairchild, T.J. Exploring the efficacy and safety of a novel standardized ashwagandha (Withania somnifera) root extract (Witholytin®) in adults experiencing high stress and fatigue in a randomized, double-blind, placebo-controlled trial. J. Psychopharmacol. 2023, 37, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Pons, D.G.; Sastre-Serra, J.; Oliver, J.; Roca, P. Sexual hormones regulate the redox status and mitochondrial function in the brain. Pathological implications. Redox Biol. 2020, 31, 101505. [Google Scholar] [CrossRef]
- Narayanan, A.K.; Nagegowda, D.A. Biosynthesis of the triterpenoid withanolides in Withaniasomnifera. Curr. Opin. Plant Biol. 2024, 81, 102576. [Google Scholar] [CrossRef]
- Anju, T.R.; Smijin, S.; Jobin, M.; Paulose, C.S. Altered muscarinic receptor expression in the cerebral cortex of epileptic rats: Restorative role of Withania somnifera. Biochem. Cell Biol. 2018, 96, 433–440. [Google Scholar] [CrossRef]
- Zhu, J.; Park, S.; Jeong, K.H.; Kim, W.J. Withanolide-A treatment exerts a neuroprotective effect via inhibiting neuroinflammation in the hippocampus after pilocarpine-induced status epilepticus. Epilepsy Res. 2020, 165, 106394. [Google Scholar] [CrossRef]
- Khan, S.A.; Khan, S.B.; Shah, Z.; Asiri, A.M. Withanolides: Biologically Active Constituents in the Treatment of Alzheimer’s Disease. Med. Chem. 2016, 12, 238–256. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Kaul, A.; Li, K.; Priyandoko, D.; Kaul, S.C.; Wadhwa, R. Effect of Ashwagandha Withanolides on Muscle Cell Differentiation. Biomolecules 2021, 11, 1454. [Google Scholar] [CrossRef]
- Brimson, J.M.; Brimson, S.; Prasanth, M.I.; Thitilertdecha, P.; Malar, D.S.; Tencomnao, T. The effectiveness of Bacopa monnieri (Linn.) Wettst. as a nootropic, neuroprotective, or antidepressant supplement: Analysis of the available clinical data. Sci. Rep. 2021, 11, 596. [Google Scholar] [CrossRef] [PubMed]
- Sushma; Sahu, M.R.; Murugan, N.A.; Mondal, A.C. Amelioration of Amyloid-β Induced Alzheimer’s Disease by Bacopa monnieri through Modulation of Mitochondrial Dysfunction and GSK-3β/Wnt/β-Catenin Signaling. Mol. Nutr. Food Res. 2024, 68, e2300245. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L.; Smith, S.J.; Ali, S.; Metse, A.P.; Kalns, J.; Drummond, P.D. Effects of a Bacopa monnieri extract (Bacognize®) on stress, fatigue, quality of life and sleep in adults with self-reported poor sleep: A randomised, double-blind, placebo-controlled study. J. Funct. Foods 2021, 85, 104671. [Google Scholar] [CrossRef]
- Palollathil, A.; Najar, M.A.; Amrutha, S.; Pervaje, R.; Modi, P.K.; Prasad, T.S.K. Bacopa monnieri confers neuroprotection by influencing signaling pathways associated with interleukin 4, 13 and extracellular matrix organization in Alzheimer’s disease: A proteomics-based perspective. Neurochem. Int. 2024, 180, 105864. [Google Scholar] [CrossRef]
- Gupta, V.; Prasad, S. Differential Alterations in the Expression of AMPA Receptor and Its Trafficking Proteins in the Hippocampus Are Associated with Recognition Memory Impairment in the Rotenone-Parkinson’s Disease Mouse Model: Neuroprotective Role of Bacopa monnieri Extract CDRI 08. Mol. Neurobiol. 2024, 1–19. [Google Scholar] [CrossRef]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; Dos Santos Buglio, D.; Barbalho Lamas, C.; Direito, R.; Fornari Laurindo, L.; Tanaka, M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants 2024, 13, 393. [Google Scholar] [CrossRef]
- Bhandari, P.; Sendri, N.; Devidas, S.B. Dammarane triterpenoid glycosides in Bacopa monnieri: A review on chemical diversity and bioactivity. Phytochemistry 2020, 172, 112276. [Google Scholar] [CrossRef]
- Castaldo, G.; Pagano, I.; Grimaldi, M.; Marino, C.; Molettieri, P.; Santoro, A.; Stillitano, I.; Romano, R.; Montoro, P.; D’Ursi, A.M.; et al. Effect of Very-Low-Calorie Ketogenic Diet on Psoriasis Patients: A Nuclear Magnetic Resonance-Based Metabolomic Study. J. Proteome Res. 2021, 20, 1509–1521. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef]
- McKay, R.T. How the 1D-NOESY suppresses solvent signal in metabonomics NMR spectroscopy: An examination of the pulse sequence components and evolution. Concepts Magn. Reson. Part A 2011, 38A, 197–220. [Google Scholar] [CrossRef]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Spigelman, A.F.; MacDonald, P.E.; Wishart, D.S.; Li, S.; et al. MetaboAnalyst 6.0: Towards a unified platform for metabolomics data processing, analysis and interpretation. Nucleic Acids Res. 2024, 52, W398–W406. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Hoque, M.A.; Sugimoto, M. Robust volcano plot: Identification of differential metabolites in the presence of outliers. BMC Bioinform. 2018, 19, 128. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef]
- Abdelwahed, M.T.; Hegazy, M.A.; Mohamed, E.H. Major biochemical constituents of Withania somnifera (ashwagandha) extract: A review of chemical analysis. Rev. Anal. Chem. 2023, 42. [Google Scholar] [CrossRef]
- Nuengchamnong, N.; Sookying, S.; Ingkaninan, K. LC-ESI-QTOF-MS based screening and identification of isomeric jujubogenin and pseudojujubogenin aglycones in Bacopa monnieri extract. J. Pharm. Biomed. Anal. 2016, 129, 121–134. [Google Scholar] [CrossRef]
- Wang, K.C.; Wang, S.Y.; Kuo, C.H.; Tseng, Y.J. Distribution-based classification method for baseline correction of metabolomic 1D proton nuclear magnetic resonance spectra. Anal. Chem. 2013, 85, 1231–1239. [Google Scholar] [CrossRef]
- Lin, C.Y.; Wu, H.; Tjeerdema, R.S.; Viant, M.R. Evaluation of metabolite extraction strategies from tissue samples using NMR metabolomics. J. Metabolomics 2007, 3, 55–67. [Google Scholar] [CrossRef]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics Off. J. Metabolomic Soc. 2017, 13, 36. [Google Scholar] [CrossRef]
- Worley, B.; Halouska, S.; Powers, R. Utilities for quantifying separation in PCA/PLS-DA scores plots. Anal. Biochem. 2013, 433, 102–104. [Google Scholar] [CrossRef]
- Ruiz-Perez, D.; Guan, H.; Madhivanan, P.; Mathee, K.; Narasimhan, G. So you think you can PLS-DA? BMC Bioinform. 2020, 21, 2. [Google Scholar] [CrossRef]
- Akarachantachote, N.; Chadcham, S.; Saithanu, K. Cutoff threshold of variable importance in projection for variable selection. J. Int. J. Pure Appl. Math. 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Aquili, L. The Role of Tryptophan and Tyrosine in Executive Function and Reward Processing. Int. J. Tryptophan Res. IJTR 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Fendt, S.M.; Verstreken, P. Neurons eat glutamate to stay alive. J. Cell Biol. 2017, 216, 863–865. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Lemasters, J.J. ATP/ADP ratio, the missed connection between mitochondria and the Warburg effect. Mitochondrion 2014, 19 Pt A, 78–84. [Google Scholar] [CrossRef]
- Gómez Afonso, A.; Fernandez-Lazaro, D.; Adams, D.P.; Monserdà-Vilaró, A.; Fernandez-Lazaro, C.I. Effects of Withania somnifera (Ashwagandha) on Hematological and Biochemical Markers, Hormonal Behavior, and Oxidant Response in Healthy Adults: A Systematic Review. Curr. Nutr. Rep. 2023, 12, 465–477. [Google Scholar] [CrossRef]
- Moon, J.Y.; Choi, M.H.; Kim, J. Metabolic profiling of cholesterol and sex steroid hormones to monitor urological diseases. Endocr.-Relat. Cancer 2016, 23, R455–R467. [Google Scholar] [CrossRef]
- Lamarre, S.G.; Morrow, G.; Macmillan, L.; Brosnan, M.E.; Brosnan, J.T. Formate: An essential metabolite, a biomarker, or more? Clin. Chem. Lab. Med. 2013, 51, 571–578. [Google Scholar] [CrossRef]
- Leonardi, R.; Zhang, Y.M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Prog. Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef]
- Li, S.; Sheng, Z.H. Oligodendrocyte-derived transcellular signaling regulates axonal energy metabolism. Curr. Opin. Neurobiol. 2023, 80, 102722. [Google Scholar] [CrossRef]
- Moiseenok, A.G.; Kanunnikova, N.P. Brain CoA and Acetyl CoA Metabolism in Mechanisms of Neurodegeneration. Biochem. Biokhimiia 2023, 88, 466–480. [Google Scholar] [CrossRef]
- Szrok-Jurga, S.; Turyn, J.; Hebanowska, A.; Swierczynski, J.; Czumaj, A.; Sledzinski, T.; Stelmanska, E. The Role of Acyl-CoA β-Oxidation in Brain Metabolism and Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 13977. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective Effect of Antioxidants in the Brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef] [PubMed]
- Ronowska, A.; Szutowicz, A.; Bielarczyk, H.; Gul-Hinc, S.; Klimaszewska-Łata, J.; Dyś, A.; Zyśk, M.; Jankowska-Kulawy, A. The Regulatory Effects of Acetyl-CoA Distribution in the Healthy and Diseased Brain. Front. Cell. Neurosci. 2018, 12, 169. [Google Scholar] [CrossRef]
- Sang, C.; Philbert, S.A.; Hartland, D.; Unwin, R.D.; Dowsey, A.W.; Xu, J.; Cooper, G.J.S. Coenzyme A-Dependent Tricarboxylic Acid Cycle Enzymes Are Decreased in Alzheimer’s Disease Consistent with Cerebral Pantothenate Deficiency. Front. Aging Neurosci. 2022, 14, 893159. [Google Scholar] [CrossRef]
- Kalaiselvi, T.; Panneerselvam, C. Effect of l-carnitine on the status of lipid peroxidation and antioxidants in aging rats. J. Nutr. Biochem. 1998, 9, 575–581. [Google Scholar] [CrossRef]
- Virmani, M.A.; Cirulli, M. The Role of l-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. Int. J. Mol. Sci. 2022, 23, 2717. [Google Scholar] [CrossRef]
- Maldonado, C.; Vázquez, M.; Fagiolino, P. Potential Therapeutic Role of Carnitine and Acetylcarnitine in Neurological Disorders. Curr. Pharm. Des. 2020, 26, 1277–1285. [Google Scholar] [CrossRef]
- Jones, L.L.; McDonald, D.A.; Borum, P.R. Acylcarnitines: Role in brain. Prog. Lipid Res. 2010, 49, 61–75. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. a | RT [min] | m/z | Formula | ppm | MS/MS | Name |
|---|---|---|---|---|---|---|
| 1 | 6.8 | 224.1644 | C13H22O2N | −0.47 | 124, 93 | unknown |
| 2 | 9.9 | 226.1801 | C13H24O2N | −0.334 | 144, 126, 84 | unknown |
| 3 | 10.9 | 611.1597 | C27H31O16 | −1.524 | 465, 303 | Rutin b |
| 4 | 12.71 | 595.1658 | C27H31O15 | 0.241 | 449, 287 | Kaempferol hexose-deoxyhexose |
| 5 | 13.1 | 611.1959 | C28H35O15 | −1.745 | 447, 303 | Hesperidin b |
| 6 | 16.3 | 303.0497 | C15H11O7 | −0.723 | 153 | Quercetin b |
| 7 | 17.4 | 783.4157 | C40H63O15 | −0.482 | 459, 441, 423 | Withanoside IV/Withanoside X |
| 8 | 17.6 | 489.2847 | C28H41O7 | 0.225 | 317, 299, 281 | Vicosalactone B isomer |
| 9 | 17.8 | 489.2847 | C28H41O7 | 0.163 | 317, 299, 281 | Vicosalactone B isomer |
| 10 | 17.8 | 783.4156 | C40H63O15 | −0.635 | 459, 441, 423 | Withanoside IV/Withanoside X |
| 11 | 18.1 | 621.363 | C34H53O10 | −0.457 | 459, 441, 423 | Coagulin Q |
| 12 | 18.7 | 621.3632 | C34H53O10 | −0.264 | 459, 441, 423 | Coagulin Q isomer |
| 13 | 20.8 | 471.2737 | C28H39O6 | −0.733 | 435, 341, 299, 281 | Whitanolide A isomer |
| 14 | 21.8 | 471.2739 | C28H39O6 | −0.457 | 435, 341, 299, 281 | Whitanolide A isomer |
| 15 | 22.8 | 471.2737 | C28H39O6 | −0.797 | 435, 341, 299, 281 | Whitanolide A isomer |
| 16 | 22.8 | 767.4209 | C40H63O14 | −0.434 | 443, 425, 407, 389 | Withanoside V |
| 17 | 28.0 | 797.4688 | C42H69O14 | 0.861 | 599, 441, 423 | Bacopaside N |
| 18 | 31.4 | 931.5315 | C40H83O23 | −0.51 | 477 | unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Elia, M.; Marino, C.; Celano, R.; Napolitano, E.; D’Ursi, A.M.; Russo, M.; Rastrelli, L. Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic. Nutrients 2024, 16, 4096. https://doi.org/10.3390/nu16234096
D’Elia M, Marino C, Celano R, Napolitano E, D’Ursi AM, Russo M, Rastrelli L. Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic. Nutrients. 2024; 16(23):4096. https://doi.org/10.3390/nu16234096
Chicago/Turabian StyleD’Elia, Maria, Carmen Marino, Rita Celano, Enza Napolitano, Anna Maria D’Ursi, Mariateresa Russo, and Luca Rastrelli. 2024. "Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic" Nutrients 16, no. 23: 4096. https://doi.org/10.3390/nu16234096
APA StyleD’Elia, M., Marino, C., Celano, R., Napolitano, E., D’Ursi, A. M., Russo, M., & Rastrelli, L. (2024). Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic. Nutrients, 16(23), 4096. https://doi.org/10.3390/nu16234096