
Effects of Fermented Polygonum cuspidatum on the Skeletal Muscle Functions

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Sample Extraction and Fermentation
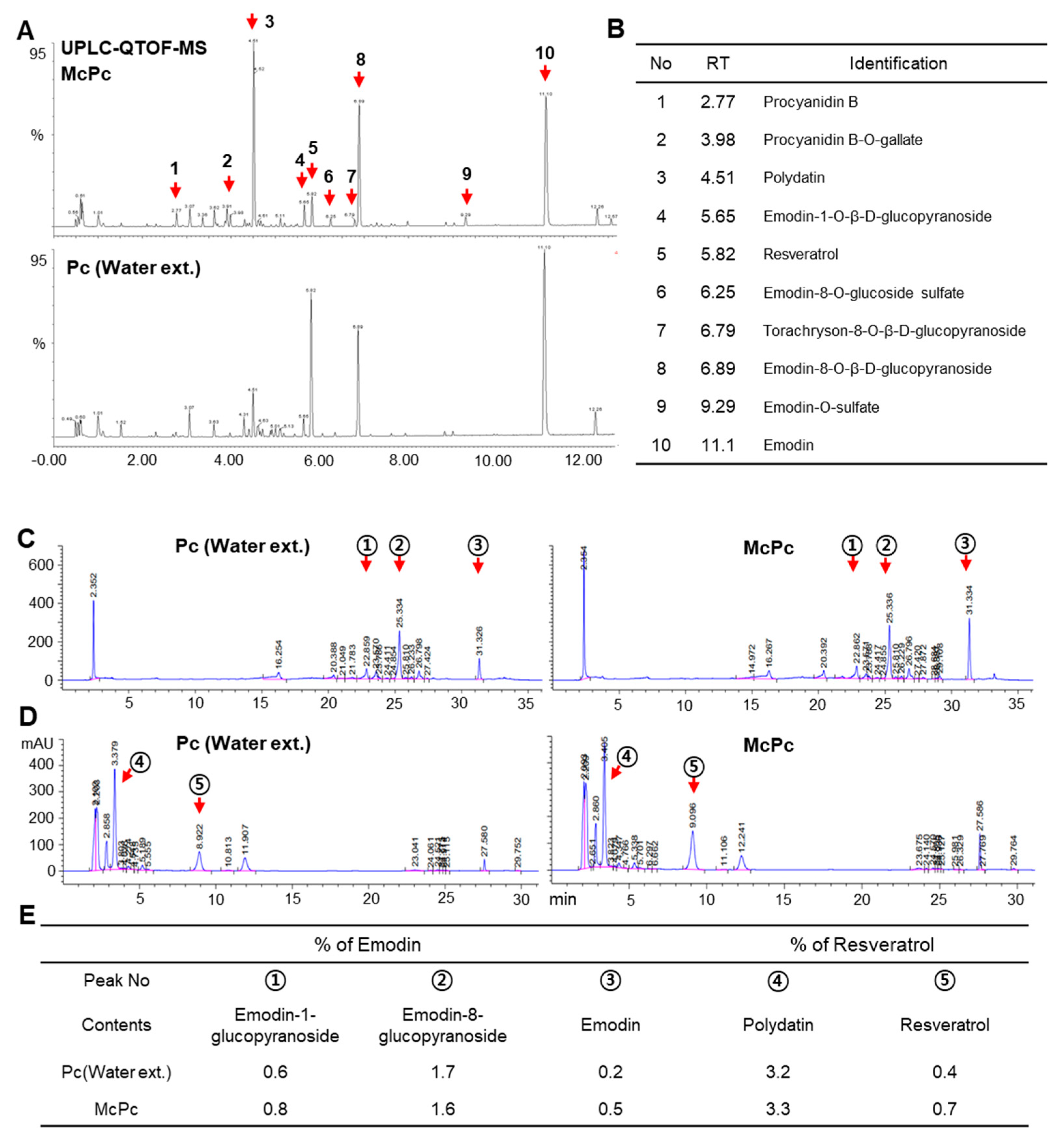
2.3. UPLC–QTOF/MS Experiments and Data Analysis
2.4. HPLC Experiments and Data Analysis
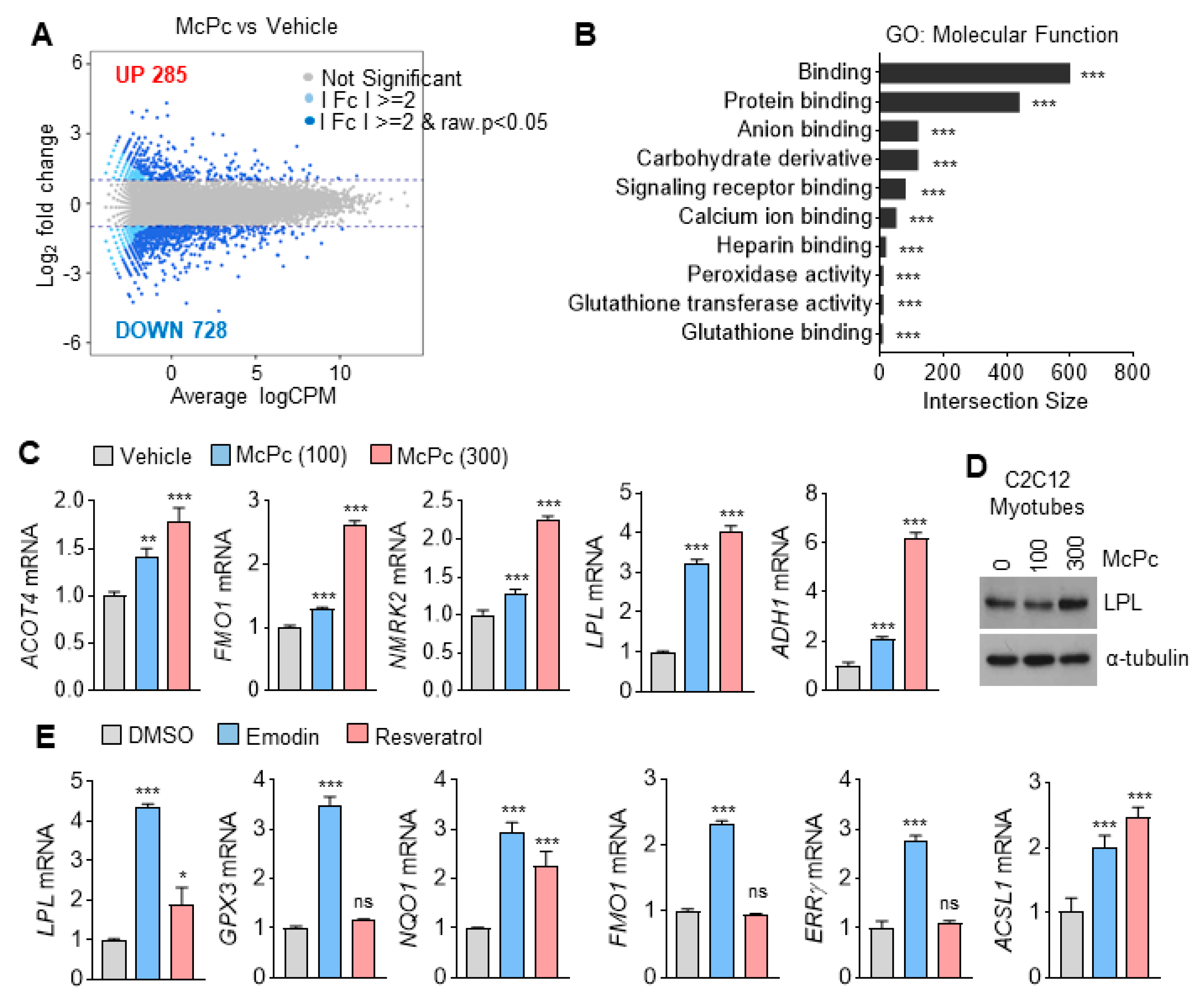
2.5. RNA Seq and Quantitative Real-Time PCR (qRT-PCR)
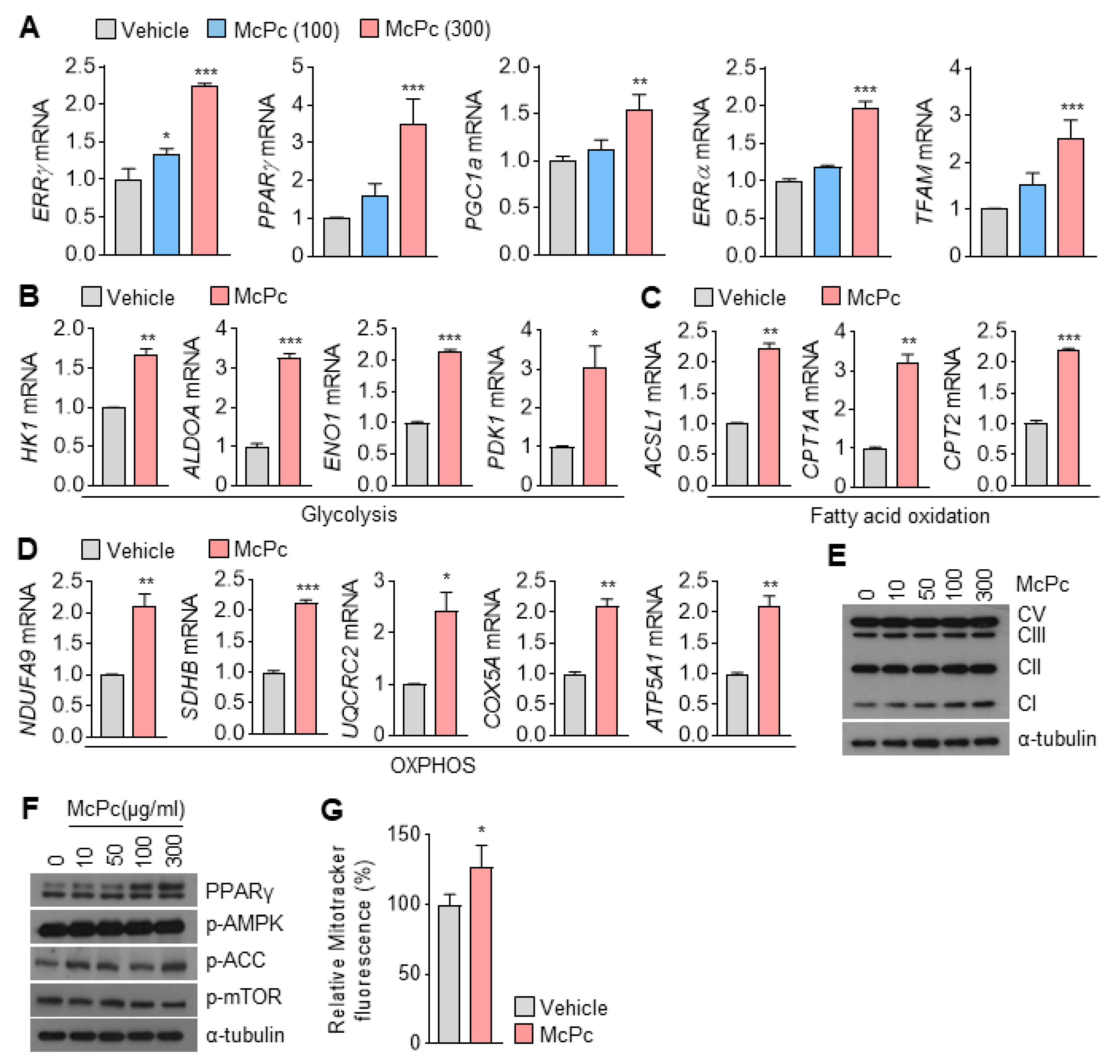
2.6. Immunoblotting
2.7. Animal Experiment
2.8. Blood Biochemistry
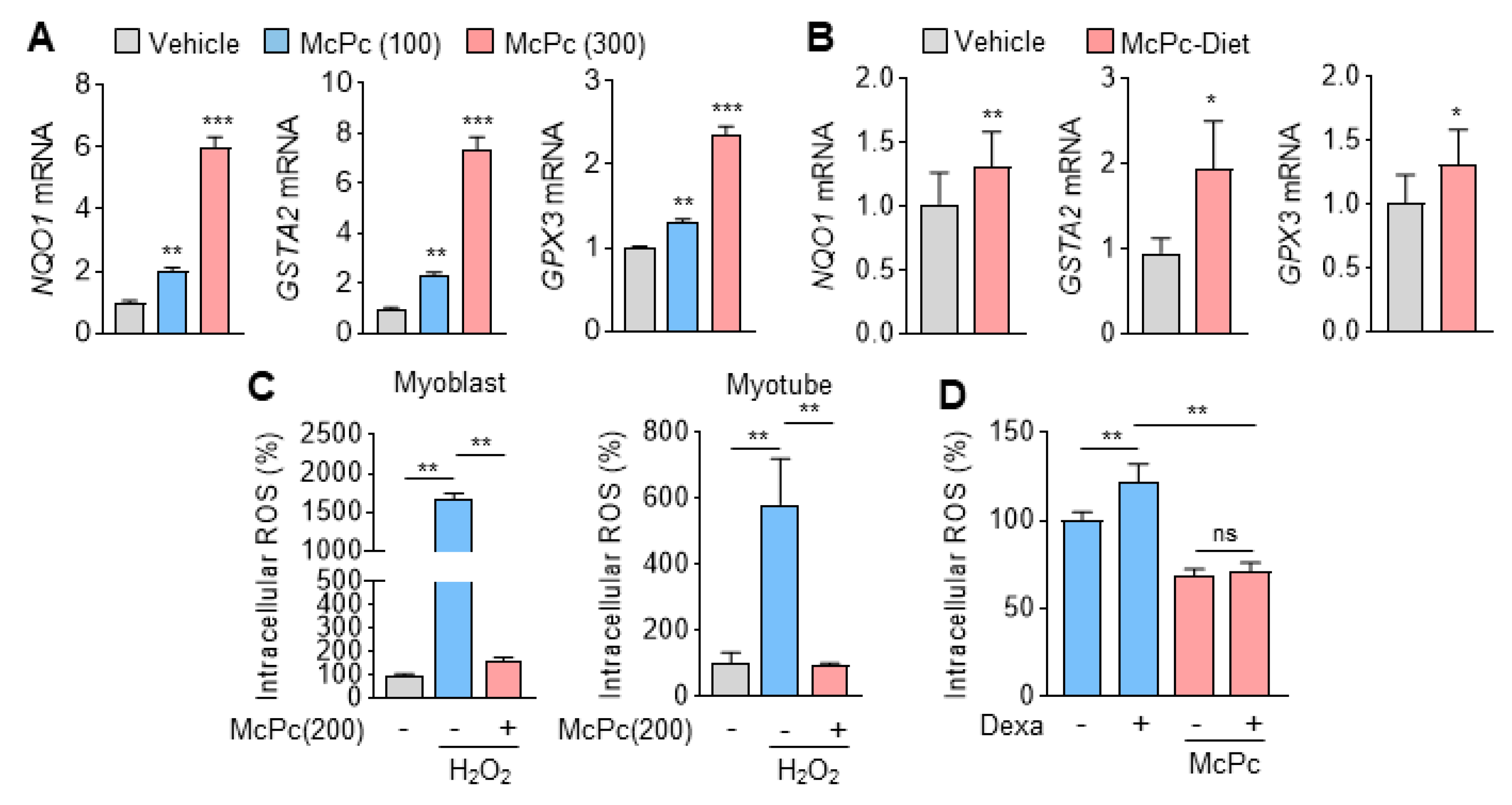
2.9. Measurement of ROS
2.10. Statistical Analysis
3. Results
3.1. Production of Emodin and Resveratrol-Enriched Polygonum cuspidatum (McPc) through Fermentation
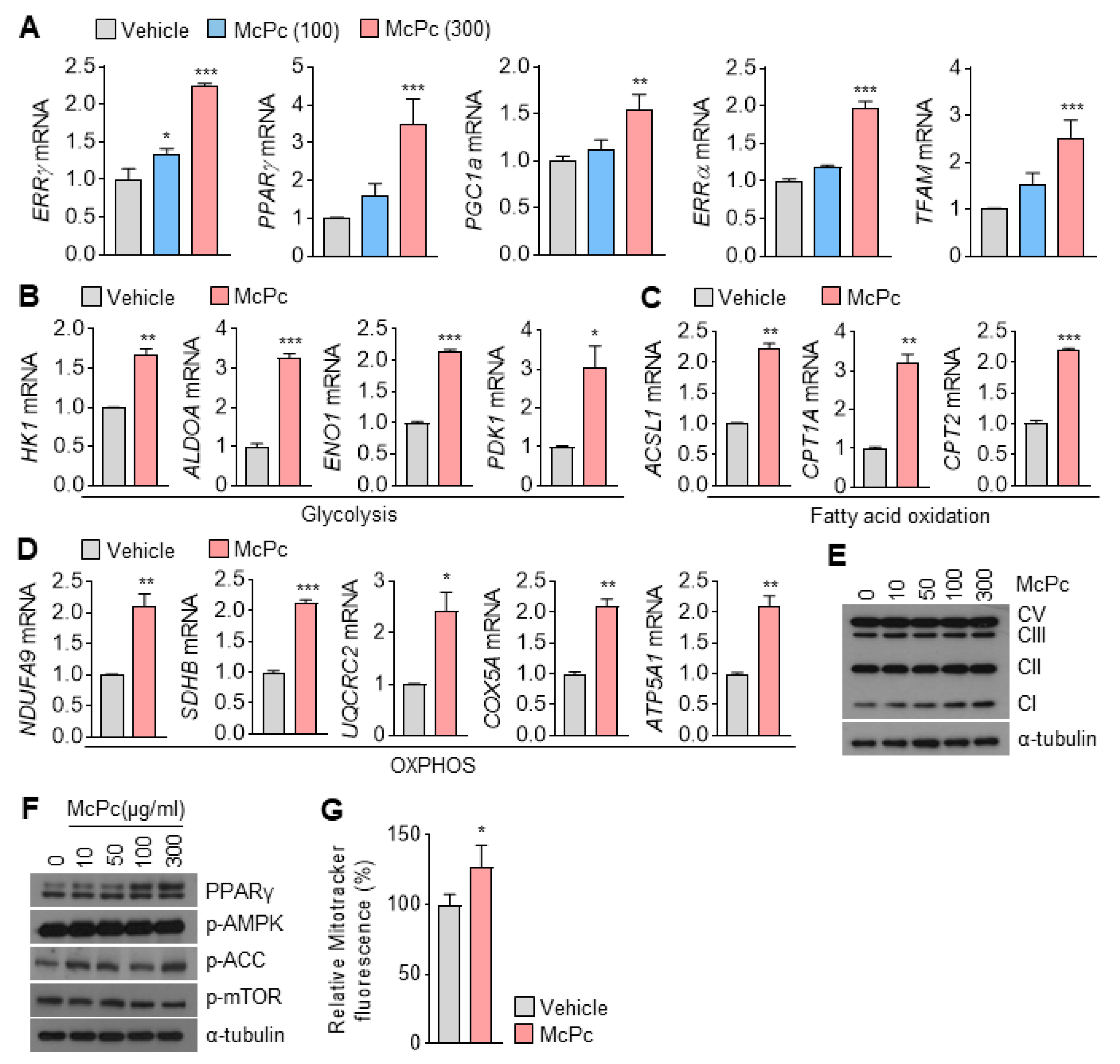
3.2. Identification of Target Genes Responsive to McPc
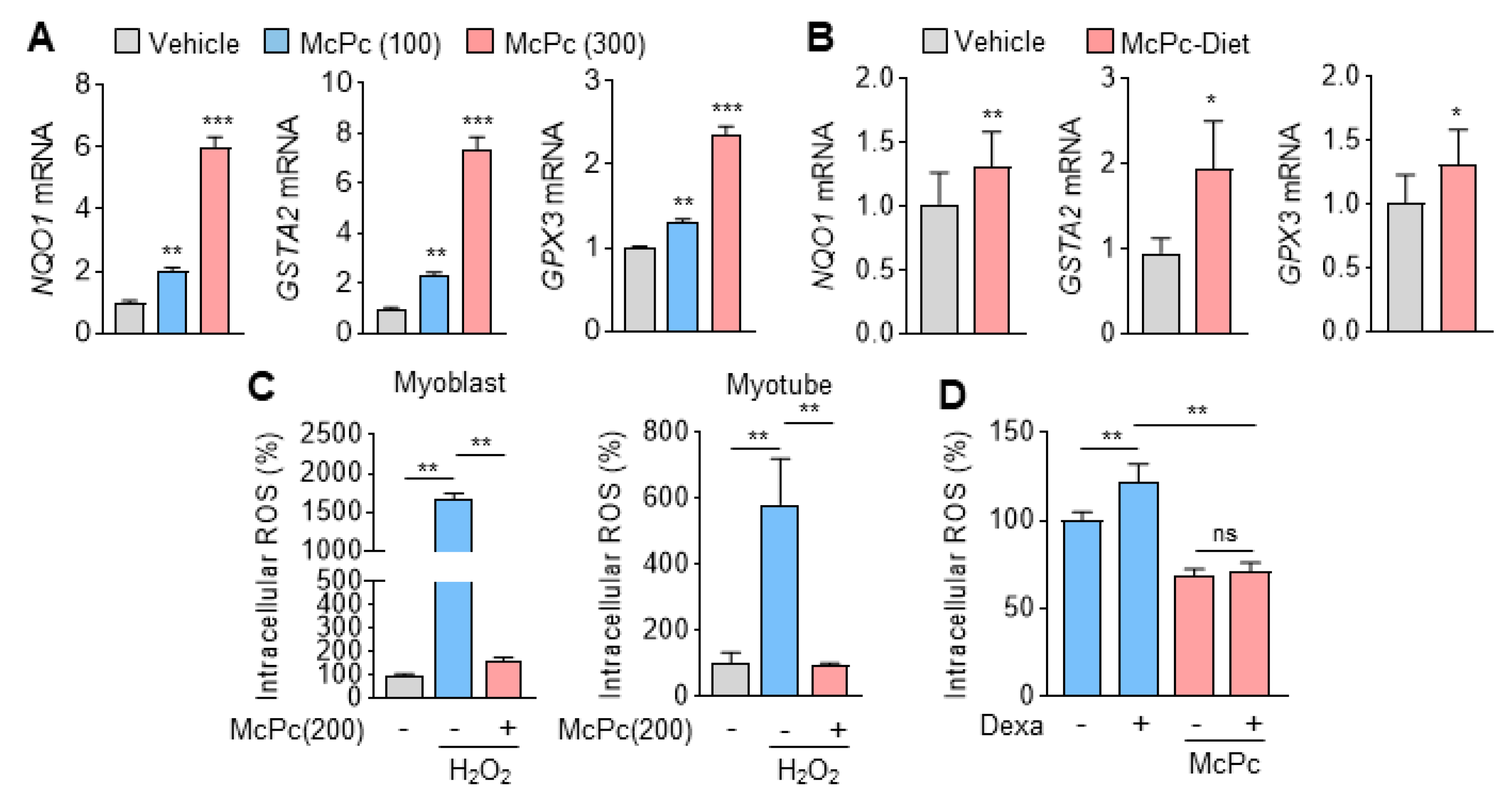
3.3. McPc Prevents Oxidative Stress In Vitro and In Vivo
3.4. McPc Enhances Expression of Mitochondrial Oxidative Capacity-Associated Genes in Cultured Myotubes
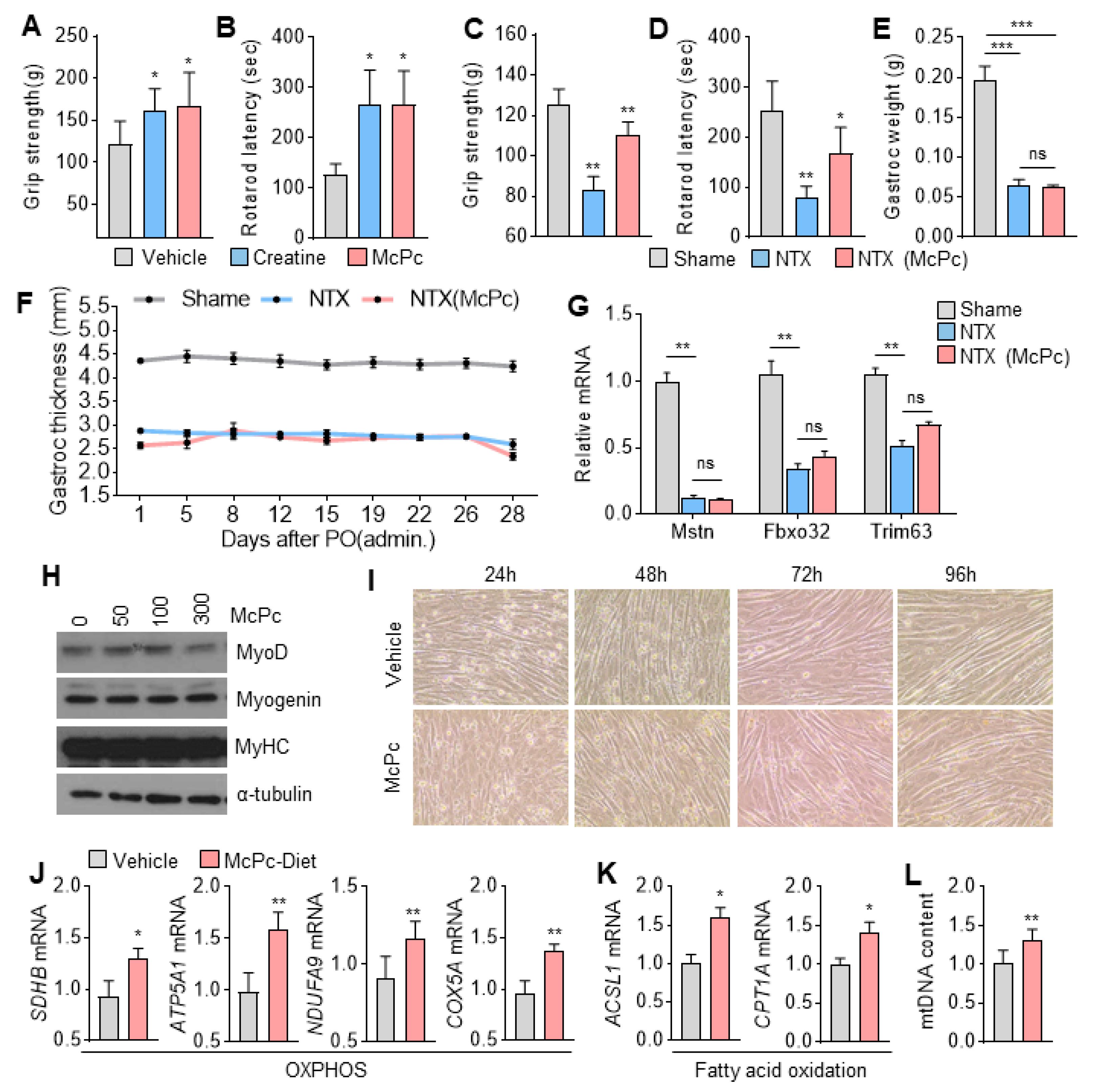
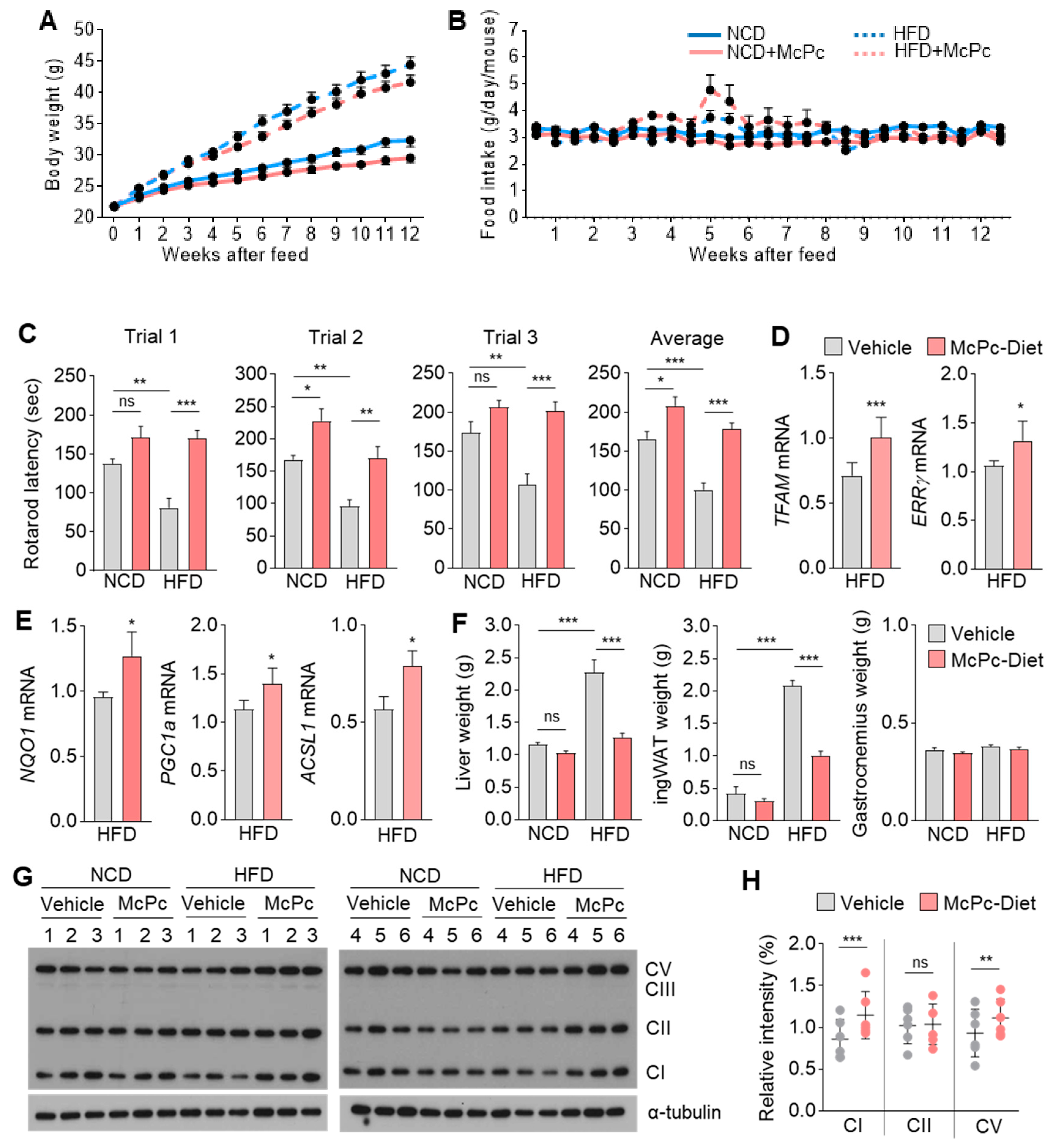
3.5. McPc Improves Skeletal Muscle Strength and Motor Function
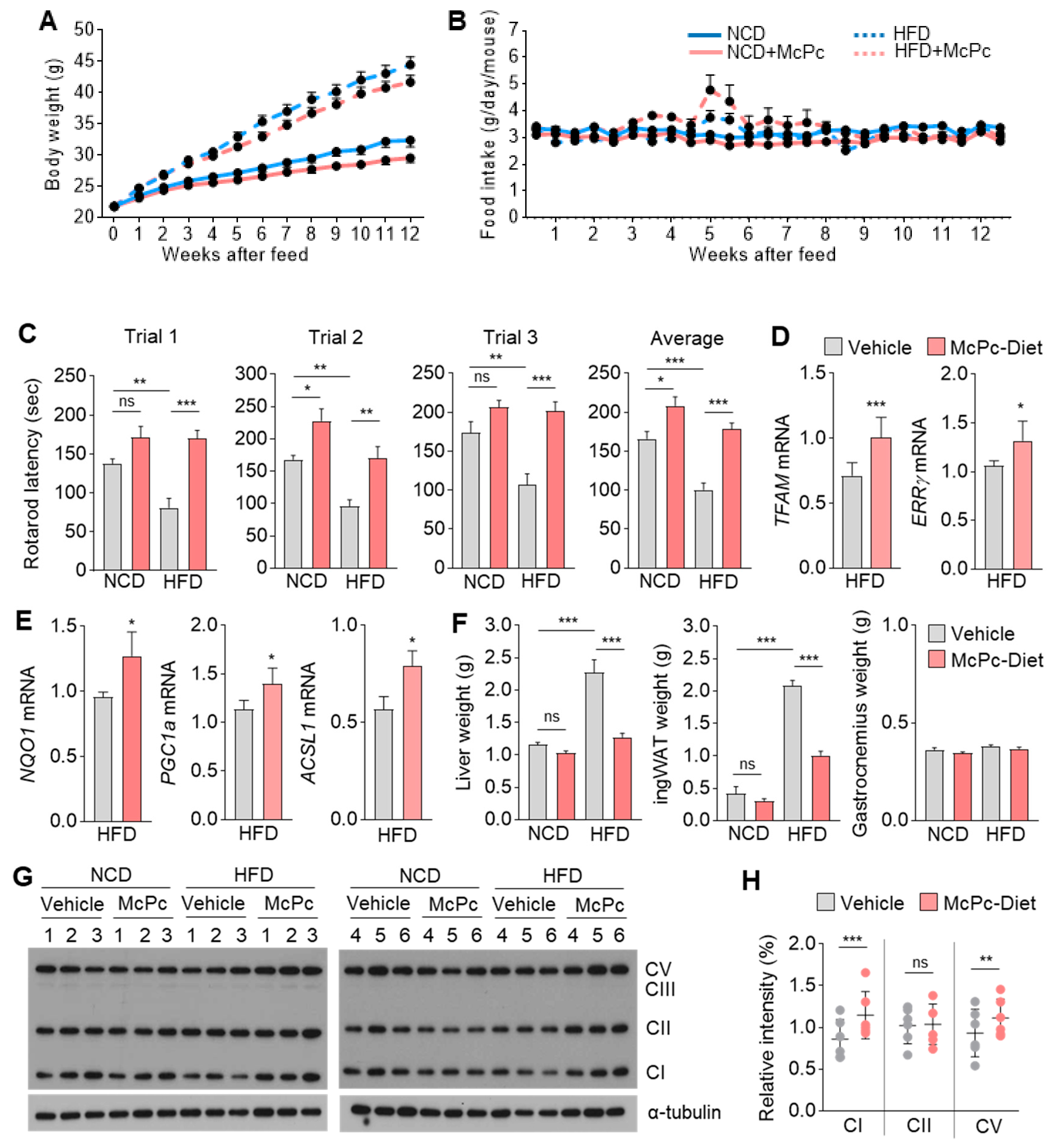
3.6. McPc Improves HFD-Induced Impaired Skeletal Muscle Strength, Fatty Liver, Hyperglycemia, and Hyperlipidemia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ke, J.; Li, M.T.; Xu, S.; Ma, J.; Liu, M.Y.; Han, Y. Advances for pharmacological activities of Polygonum cuspidatum—A review. Pharm. Biol. 2023, 61, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Geng, Q.; Wei, Q.; Wang, S.; Qi, H.; Zhu, Q.; Liu, X.; Shi, X.; Wen, S. Physcion 8-O-beta-glucopyranoside extracted from Polygonum cuspidatum exhibits anti-proliferative and anti-inflammatory effects on MH7A rheumatoid arthritis-derived fibroblast-like synoviocytes through the TGF-beta/MAPK pathway. Int. J. Mol. Med. 2018, 42, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, S.; Sui, X.; Guo, L.; Liu, X.; Li, H.; Gao, L.; Cai, S.; Li, Y.; Wang, T.; et al. Root Extract of Polygonum cuspidatum Siebold & Zucc. Ameliorates DSS-Induced Ulcerative Colitis by Affecting NF-kappaB Signaling Pathway in a Mouse Model via Synergistic Effects of Polydatin, Resveratrol, and Emodin. Front. Pharmacol. 2018, 9, 347. [Google Scholar] [CrossRef]
- Zeng, H.; Wang, Y.; Gu, Y.; Wang, J.; Zhang, H.; Gao, H.; Jin, Q.; Zhao, L. Polydatin attenuates reactive oxygen species-induced airway remodeling by promoting Nrf2-mediated antioxidant signaling in asthma mouse model. Life Sci. 2019, 218, 25–30. [Google Scholar] [CrossRef]
- Ding, W.; Dong, M.; Deng, J.; Yan, D.; Liu, Y.; Xu, T.; Liu, J. Polydatin attenuates cardiac hypertrophy through modulation of cardiac Ca2+ handling and calcineurin-NFAT signaling pathway. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H792–H802. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.Q.; Kim, Y.S.; Lee, Y.M.; Lee, M.; Lim, W.J.; Yim, W.J.; Han, M.W.; Lim, J.H. Polygonum cuspidatum Extract (Pc-Ex) Containing Emodin Suppresses Lung Cancer-Induced Cachexia by Suppressing TCF4/TWIST1 Complex-Induced PTHrP Expression. Nutrients 2022, 14, 1508. [Google Scholar] [CrossRef]
- Zhao, Y.S.; Eweys, A.S.; Zhang, J.Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.B.; Xiao, X. Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components. Antioxidants 2021, 10, 2004. [Google Scholar] [CrossRef]
- Mazloom, K.; Siddiqi, I.; Covasa, M. Probiotics: How Effective Are They in the Fight against Obesity? Nutrients 2019, 11, 258. [Google Scholar] [CrossRef]
- Mallet, J.F.; Shahbazi, R.; Alsadi, N.; Matar, C. Polyphenol-Enriched Blueberry Preparation Controls Breast Cancer Stem Cells by Targeting FOXO1 and miR-145. Molecules 2021, 26, 4330. [Google Scholar] [CrossRef]
- Shahbazi, R.; Sharifzad, F.; Bagheri, R.; Alsadi, N.; Yasavoli-Sharahi, H.; Matar, C. Anti-Inflammatory and Immunomodulatory Properties of Fermented Plant Foods. Nutrients 2021, 13, 1516. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Piasecki, M.; Atherton, P.J. The age-related loss of skeletal muscle mass and function: Measurement and physiology of muscle fibre atrophy and muscle fibre loss in humans. Ageing Res. Rev. 2018, 47, 123–132. [Google Scholar] [CrossRef]
- Roh, E.; Choi, K.M. Health Consequences of Sarcopenic Obesity: A Narrative Review. Front. Endocrinol. 2020, 11, 332. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.D.; Fan, S.D.; Chen, X.Y.; Yan, X.L.; Zhang, X.Z.; Ma, B.W.; Yu, D.Y.; Xiao, W.Y.; Zhuang, C.L.; Yu, Z. Nrf2 deficiency exacerbates frailty and sarcopenia by impairing skeletal muscle mitochondrial biogenesis and dynamics in an age-dependent manner. Exp. Gerontol. 2019, 119, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Garnier, A.; Fortin, D.; Delomenie, C.; Momken, I.; Veksler, V.; Ventura-Clapier, R. Depressed mitochondrial transcription factors and oxidative capacity in rat failing cardiac and skeletal muscles. J. Physiol. 2003, 551, 491–501. [Google Scholar] [CrossRef]
- Taylor, D.F.; Bishop, D.J. Transcription Factor Movement and Exercise-Induced Mitochondrial Biogenesis in Human Skeletal Muscle: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 1517. [Google Scholar] [CrossRef]
- Fan, W.; Evans, R. PPARs and ERRs: Molecular mediators of mitochondrial metabolism. Curr. Opin. Cell Biol. 2015, 33, 49–54. [Google Scholar] [CrossRef]
- Murakami, T.; Shimomura, Y.; Yoshimura, A.; Sokabe, M.; Fujitsuka, N. Induction of nuclear respiratory factor-1 expression by an acute bout of exercise in rat muscle. Biochim. Biophys. Acta 1998, 1381, 113–122. [Google Scholar] [CrossRef]
- Baar, K.; Song, Z.; Semenkovich, C.F.; Jones, T.E.; Han, D.H.; Nolte, L.A.; Ojuka, E.O.; Chen, M.; Holloszy, J.O. Skeletal muscle overexpression of nuclear respiratory factor 1 increases glucose transport capacity. FASEB J. 2003, 17, 1666–1673. [Google Scholar] [CrossRef]
- Ramachandran, B.; Yu, G.; Gulick, T. Nuclear respiratory factor 1 controls myocyte enhancer factor 2A transcription to provide a mechanism for coordinate expression of respiratory chain subunits. J. Biol. Chem. 2008, 283, 11935–11946. [Google Scholar] [CrossRef] [PubMed]
- Rangwala, S.M.; Wang, X.; Calvo, J.A.; Lindsley, L.; Zhang, Y.; Deyneko, G.; Beaulieu, V.; Gao, J.; Turner, G.; Markovits, J. Estrogen-related receptor gamma is a key regulator of muscle mitochondrial activity and oxidative capacity. J. Biol. Chem. 2010, 285, 22619–22629. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Atkins, A.R.; Yu, R.T.; Downes, M.; Evans, R.M. Road to exercise mimetics: Targeting nuclear receptors in skeletal muscle. J. Mol. Endocrinol. 2013, 51, T87–T100. [Google Scholar] [CrossRef]
- Fan, W.; Waizenegger, W.; Lin, C.S.; Sorrentino, V.; He, M.X.; Wall, C.E.; Li, H.; Liddle, C.; Yu, R.T.; Atkins, A.R.; et al. PPARdelta Promotes Running Endurance by Preserving Glucose. Cell Metab. 2017, 25, 1186–1193.e4. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Lee, Y.H.; Chang, Y.U.; Yi, H.K. PPARgamma regulates inflammatory reaction by inhibiting the MAPK/NF-kappaB pathway in C2C12 skeletal muscle cells. J. Physiol. Biochem. 2017, 73, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Schrager, M. Redox signaling regulates skeletal muscle remodeling in response to exercise and prolonged inactivity. Redox Biol. 2022, 54, 102374. [Google Scholar] [CrossRef]
- Jia, W.; Wang, C.; Wang, Y.; Pan, G.; Jiang, M.; Li, Z.; Zhu, Y. Qualitative and quantitative analysis of the major constituents in Chinese medical preparation Lianhua-Qingwen capsule by UPLC-DAD-QTOF-MS. Sci. World J. 2015, 2015, 731765. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, X.; Chen, M.; Hu, H.; Cao, J.; Chai, T.; Wang, H. A Study on Tissue-Specific Metabolite Variations in Polygonum cuspidatum by High-Resolution Mass Spectrometry-Based Metabolic Profiling. Molecules 2019, 24, 1058. [Google Scholar] [CrossRef]
- Wang, X.; Hu, H.; Wu, Z.; Fan, H.; Wang, G.; Chai, T.; Wang, H. Tissue-specific transcriptome analyses reveal candidate genes for stilbene, flavonoid and anthraquinone biosynthesis in the medicinal plant Polygonum cuspidatum. BMC Genom. 2021, 22, 353. [Google Scholar] [CrossRef]
- Oh, T.I.; Lee, M.; Lee, Y.M.; Kim, G.H.; Lee, D.; You, J.S.; Kim, S.H.; Choi, M.; Jang, H.; Park, Y.M.; et al. PGC1alpha Loss Promotes Lung Cancer Metastasis through Epithelial-Mesenchymal Transition. Cancers 2021, 13, 1772. [Google Scholar] [CrossRef]
- Ogawa, T.; Nikawa, T.; Furochi, H.; Kosyoji, M.; Hirasaka, K.; Suzue, N.; Sairyo, K.; Nakano, S.; Yamaoka, T.; Itakura, M.; et al. Osteoactivin upregulates expression of MMP-3 and MMP-9 in fibroblasts infiltrated into denervated skeletal muscle in mice. Am. J. Physiol. Cell Physiol. 2005, 289, C697–C707. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, P.; Plant, P.J.; Correa, J.; Bain, A.; Takeda, M.; Kawabe, H.; Rotin, D.; Bain, J.R.; Batt, J.A. The ubiquitin ligase Nedd4-1 participates in denervation-induced skeletal muscle atrophy in mice. PLoS ONE 2012, 7, e46427. [Google Scholar] [CrossRef]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2’,7’-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Abdul Hakim, B.N.; Xuan, N.J.; Oslan, S.N.H. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, R.; Li, W. Emodin in cardiovascular disease: The role and therapeutic potential. Front. Pharmacol. 2022, 13, 1070567. [Google Scholar] [CrossRef] [PubMed]
- Monisha, B.A.; Kumar, N.; Tiku, A.B. Emodin and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 47–73. [Google Scholar] [CrossRef]
- Ulla, A.; Uchida, T.; Miki, Y.; Sugiura, K.; Higashitani, A.; Kobayashi, T.; Ohno, A.; Nakao, R.; Hirasaka, K.; Sakakibara, I.; et al. Morin attenuates dexamethasone-mediated oxidative stress and atrophy in mouse C2C12 skeletal myotubes. Arch. Biochem. Biophys. 2021, 704, 108873. [Google Scholar] [CrossRef]
- Axelrod, C.L.; Dantas, W.S.; Kirwan, J.P. Sarcopenic obesity: Emerging mechanisms and therapeutic potential. Metabolism 2023, 146, 155639. [Google Scholar] [CrossRef]
- Zahedi, H.S.; Jazayeri, S.; Ghiasvand, R.; Djalali, M.; Eshraghian, M.R. Effects of Polygonum cuspidatum containing resveratrol on inflammation in male professional basketball players. Int. J. Prev. Med. 2013, 4, S1–S4. [Google Scholar]
- Ghanim, H.; Sia, C.L.; Abuaysheh, S.; Korzeniewski, K.; Patnaik, P.; Marumganti, A.; Chaudhuri, A.; Dandona, P. An antiinflammatory and reactive oxygen species suppressive effects of an extract of Polygonum cuspidatum containing resveratrol. J. Clin Endocrinol. Metab. 2010, 95, E1–E8. [Google Scholar] [CrossRef] [PubMed]
- Bahramzadeh, A.; Bolandnazar, K.; Meshkani, R. Resveratrol as a potential protective compound against skeletal muscle insulin resistance. Heliyon 2023, 9, e21305. [Google Scholar] [CrossRef]
- Delmas, D.; Cornebise, C.; Courtaut, F.; Xiao, J.; Aires, V. New Highlights of Resveratrol: A Review of Properties against Ocular Diseases. Int. J. Mol. Sci. 2021, 22, 1295. [Google Scholar] [CrossRef]
- Jhanji, M.; Rao, C.N.; Massey, J.C.; Hope, M.C., 3rd; Zhou, X.; Keene, C.D.; Ma, T.; Wyatt, M.D.; Stewart, J.A.; Sajish, M. Cis- and trans-resveratrol have opposite effects on histone serine-ADP-ribosylation and tyrosine induced neurodegeneration. Nat. Commun. 2022, 13, 3244. [Google Scholar] [CrossRef] [PubMed]
- Lauretti, E.; Di Meco, A.; Merali, S.; Pratico, D. Chronic behavioral stress exaggerates motor deficit and neuroinflammation in the MPTP mouse model of Parkinson’s disease. Transl. Psychiatry 2016, 6, e733. [Google Scholar] [CrossRef]
- Schwerzmann, K.; Hoppeler, H.; Kayar, S.R.; Weibel, E.R. Oxidative capacity of muscle and mitochondria: Correlation of physiological, biochemical, and morphometric characteristics. Proc. Natl. Acad. Sci. USA 1989, 86, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of mitochondrial biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.P.; Li, Y.Q.; Li, Y.J.; Zi, L.; Tao, Y.X.; Hao, J.J.; Zhang, M.; Gu, W.; Zhang, F.; Yu, J.; et al. In vivo identification of the pharmacodynamic ingredients of Polygonum cuspidatum for remedying the mitochondria to alleviate metabolic dysfunction-associated fatty liver disease. Biomed. Pharmacother. 2022, 156, 113849. [Google Scholar] [CrossRef]
- Du, Y.; Ko, K.M. Effects of emodin treatment on mitochondrial ATP generation capacity and antioxidant components as well as susceptibility to ischemia-reperfusion injury in rat hearts: Single versus multiple doses and gender difference. Life Sci. 2005, 77, 2770–2782. [Google Scholar] [CrossRef]
- Song, P.; Kim, J.H.; Ghim, J.; Yoon, J.H.; Lee, A.; Kwon, Y.; Hyun, H.; Moon, H.Y.; Choi, H.S.; Berggren, P.O.; et al. Emodin regulates glucose utilization by activating AMP-activated protein kinase. J. Biol. Chem. 2013, 288, 5732–5742. [Google Scholar] [CrossRef]
- Lian, D.; Chen, M.M.; Wu, H.; Deng, S.; Hu, X. The Role of Oxidative Stress in Skeletal Muscle Myogenesis and Muscle Disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- El Assar, M.; Alvarez-Bustos, A.; Sosa, P.; Angulo, J.; Rodriguez-Manas, L. Effect of Physical Activity/Exercise on Oxidative Stress and Inflammation in Muscle and Vascular Aging. Int. J. Mol. Sci. 2022, 23, 8713. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Bian, A.L.; Hu, H.Y.; Rong, Y.D.; Wang, J.; Wang, J.X.; Zhou, X.Z. A study on relationship between elderly sarcopenia and inflammatory factors IL-6 and TNF-alpha. Eur. J. Med. Res. 2017, 22, 25. [Google Scholar] [CrossRef] [PubMed]
- Nelke, C.; Dziewas, R.; Minnerup, J.; Meuth, S.G.; Ruck, T. Skeletal muscle as potential central link between sarcopenia and immune senescence. EBioMedicine 2019, 49, 381–388. [Google Scholar] [CrossRef] [PubMed]
- White, J.P. IL-6, cancer and cachexia: Metabolic dysfunction creates the perfect storm. Transl. Cancer Res. 2017, 6, S280–S285. [Google Scholar] [CrossRef]
- Tsoli, M.; Robertson, G. Cancer cachexia: Malignant inflammation, tumorkines, and metabolic mayhem. Trends Endocrinol. Metab. 2013, 24, 174–183. [Google Scholar] [CrossRef]
- Tao, T.; Zhang, Q.; Liu, Z.; Zhang, T.; Wang, L.; Liu, J.; He, T.; Chen, Y.; Feng, J.; Chen, Y. Polygonum cuspidatum Extract Exerts Antihyperlipidemic Effects by Regulation of PI3K/AKT/FOXO3 Signaling Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 3830671. [Google Scholar] [CrossRef]
- Li, J.; Ding, L.; Song, B.; Xiao, X.; Qi, M.; Yang, Q.; Yang, Q.; Tang, X.; Wang, Z.; Yang, L. Emodin improves lipid and glucose metabolism in high fat diet-induced obese mice through regulating SREBP pathway. Eur. J. Pharmacol. 2016, 770, 99–109. [Google Scholar] [CrossRef]
- Hao, J.; Chen, C.; Huang, K.; Huang, J.; Li, J.; Liu, P.; Huang, H. Polydatin improves glucose and lipid metabolism in experimental diabetes through activating the Akt signaling pathway. Eur. J. Pharmacol. 2014, 745, 152–165. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequences (5′-3′) | Reverse Sequences (5′-3′) |
|---|---|---|
| LPL | GCCCGAGGTTTCCACAAATA | GCTGAAGTAGGAGTCGCTTATC |
| GPX3 | ATCCTGCCTTCTGTCCCTGCTC | TGGTGAGGGCTCCATACTCGTA |
| NQO1 | GACAACGGTCCTTTCCAGAATA | CTCTGAATCGGCCAGAGAATG |
| ERRγ | CAGAAGTACAAGCGCAGAATAGA | CACCAACAAATGCGAGACAATC |
| ADH1 | GAAGAAGTCTACAAGGACCCATC | CACCGCAGCTTTGCATTT |
| NMRK2 | GGCCCATGTACCAGAAGTATAG | CCTCCAGAACTTGATGGAAGAG |
| ACOT4 | GAAGAAGCAGTGCGGTACAT | GCCATGATCAGACAGACATCAG |
| GSTA2 | ATACAGAGTCCGGAAGATTTGG | GGTGGCGATGTAGTTGAGAA |
| PPARγ | CAGGCTTCCACTATGGAGTTC | GGCAGTTAAGATCACACCTATCA |
| PGC1α | AAACTGACTTCGAGCTGTACTT | CCCATGAGGTATTGACCATCTC |
| ERRα | TGCTCAGCTCTCTACCCAAAC | GGACAGCTGTACTCGATGCTC |
| TFAM | GGAATGTGGAGCGTGCTAAAA | GCTGGAAAAACACTTCGGAATA |
| HK1 | AGGGCGCATTACTCCAGAG | CCCTGTGGGTGTCTTGTGTG |
| ALDOA | CGTGTGAATCCCTGCATTGG | CAGCCCCTGGGTAGTTGTC |
| ENO1 | TGCGTCCACTGGCATCTAC | CAGAGCAGGCGCAATAGTTTTA |
| PDK1 | GGACTTCGGGTCAGTGAATGC | TCCTGAGAAGATTGTCGGGGA |
| ACSL1 | GCTTGTGGATGTGGAAGAAATG | TCTTGCTGGGTCTTTCAAGTAG |
| CPT1A | CTCTGCTGCATGGTAGATGTT | GCTCTGCGTTTATGCCTATCT |
| CPT2 | CCTGCTCGCTCAGGATAAACA | GTGTCTTCAGAAACCGCACTG |
| NDUFA9 | GTCCGCTTTCGGGTTGTTAGA | CCTCCTTTCCCGTGAGGTA |
| SDHB | AATTTGCCATTTACCGATGGGA | AGCATCCAACACCATAGGTCC |
| UQCRC2 | AAAGTTGCCCCGAAGGTTAAA | GAGCATAGTTTTCCAGAGAAGCA |
| COX5A | GCCGCTGTCTGTTCCATTC | GCATCAATGTCTGGCTTGTTGAA |
| ATP5A1 | TCTCCATGCCTCTAACACTCG | CCAGGTCAACAGACGTGTCAG |
| Mstn | CAGGAGAAGATGGGCTGAATC | AGTGCTCATCGCAGTCAAG |
| Fbxo32 | ACCCAAGAAGAGAGCAGTATG | GACTCCCAGCCATCCAATTA |
| Trim63 | GGACTACTTTACTCTGGACTTAGAAC | CAGCCTCCTCTTCTGTAAACTC |
| β-actin | CCTAAGGCCAACCGTGAAA | TGGTACGACCAGAGGCATA |
| Nutrition | NCD | NCD-McPc 4% | HFD | HFD-McPc 4% |
|---|---|---|---|---|
| Casein | 200 | 189.93 | 200 | 191.36 |
| Corn starch | 397.486 | 368.33 | 72.8 | 47.78 |
| Dextrose | 132 | 132 | - | - |
| Maltodextrin | - | - | 100 | 100 |
| Sucrose | 100 | 100 | 172.8 | 172.8 |
| Cellulose | 50 | 50 | 50 | 50 |
| Soybean Oil | 70 | 69.23 | 25 | 24.34 |
| t-Butylhydroquinone | 0.014 | 0.014 | - | - |
| Lard | - | - | 177.5 | 177.5 |
| Mineral mix | - | - | 10 | 10 |
| Dicalcium phosphate | - | - | 13 | 13 |
| Calcium carbonate | - | - | 5.5 | 5.5 |
| Potassium citrate | - | - | 16.5 | 16.5 |
| Salt Mix | 35 | 35 | - | - |
| Vitamin Mix | 10 | 10 | 10 | 10 |
| L-cystine | 3 | 3 | 3 | 3 |
| Choline Bitartrate | 2.5 | 2.5 | 2 | 2 |
| FD&C red dye | - | - | 0.05 | 0.05 |
| McPc | - | 40 | - | 34.33 |
| Total (g) | 1000 | 1000 | 858.15 | 858.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-S.; Han, J.-H.; Lim, C.-H.; Fang, X.-Q.; Jang, H.-S.; Lee, S.-Y.; Yim, W.-J.; Lim, J.-H. Effects of Fermented Polygonum cuspidatum on the Skeletal Muscle Functions. Nutrients 2024, 16, 305. https://doi.org/10.3390/nu16020305
Kim Y-S, Han J-H, Lim C-H, Fang X-Q, Jang H-S, Lee S-Y, Yim W-J, Lim J-H. Effects of Fermented Polygonum cuspidatum on the Skeletal Muscle Functions. Nutrients. 2024; 16(2):305. https://doi.org/10.3390/nu16020305
Chicago/Turabian StyleKim, Young-Seon, Ji-Hye Han, Chang-Hoon Lim, Xue-Quan Fang, Hyeock-Soon Jang, Sang-Yun Lee, Woo-Jong Yim, and Ji-Hong Lim. 2024. "Effects of Fermented Polygonum cuspidatum on the Skeletal Muscle Functions" Nutrients 16, no. 2: 305. https://doi.org/10.3390/nu16020305
APA StyleKim, Y.-S., Han, J.-H., Lim, C.-H., Fang, X.-Q., Jang, H.-S., Lee, S.-Y., Yim, W.-J., & Lim, J.-H. (2024). Effects of Fermented Polygonum cuspidatum on the Skeletal Muscle Functions. Nutrients, 16(2), 305. https://doi.org/10.3390/nu16020305