
Elephantopus scaber L. Polysaccharides Alleviate Heat Stress-Induced Systemic Inflammation in Mice via Modulation of Characteristic Gut Microbiota and Metabolites

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction of Crude Polysaccharides
2.2. Composition Analysis
2.3. Animal Ethics Statement
2.4. Animals and Study Design
2.5. Temperature-Humidity Index (THI) Measurements
2.6. Physiological Variables and Sample Collection
2.7. Measurement of Inflammatory Markers and Lipopolysaccharide (LPS)
2.8. Haematoxylin and Eosin (H&E) Staining of Intestinal Tissue
2.9. Western Blot (WB) Analysis
2.10. Gut Microbiota Analysis
2.10.1. DNA Extraction
2.10.2. 16S rRNA Amplicon Pyrosequencing and Sequence Analysis
2.10.3. Bioinformatics and Statistical Analysis
2.11. SCFA Analysis
2.12. Neurotransmitter Analysis
2.13. Pro-Inflammatory Experiment of Valeric and Glutamic Acid In Vitro
2.13.1. Cytotoxicity Test of Valeric and Glutamic Acid
2.13.2. Pro-Inflammatory Test of Valeric and Glutamic Acid
2.14. Statistical Analysis
3. Results
3.1. Basic Physicochemical Properties of ESLP
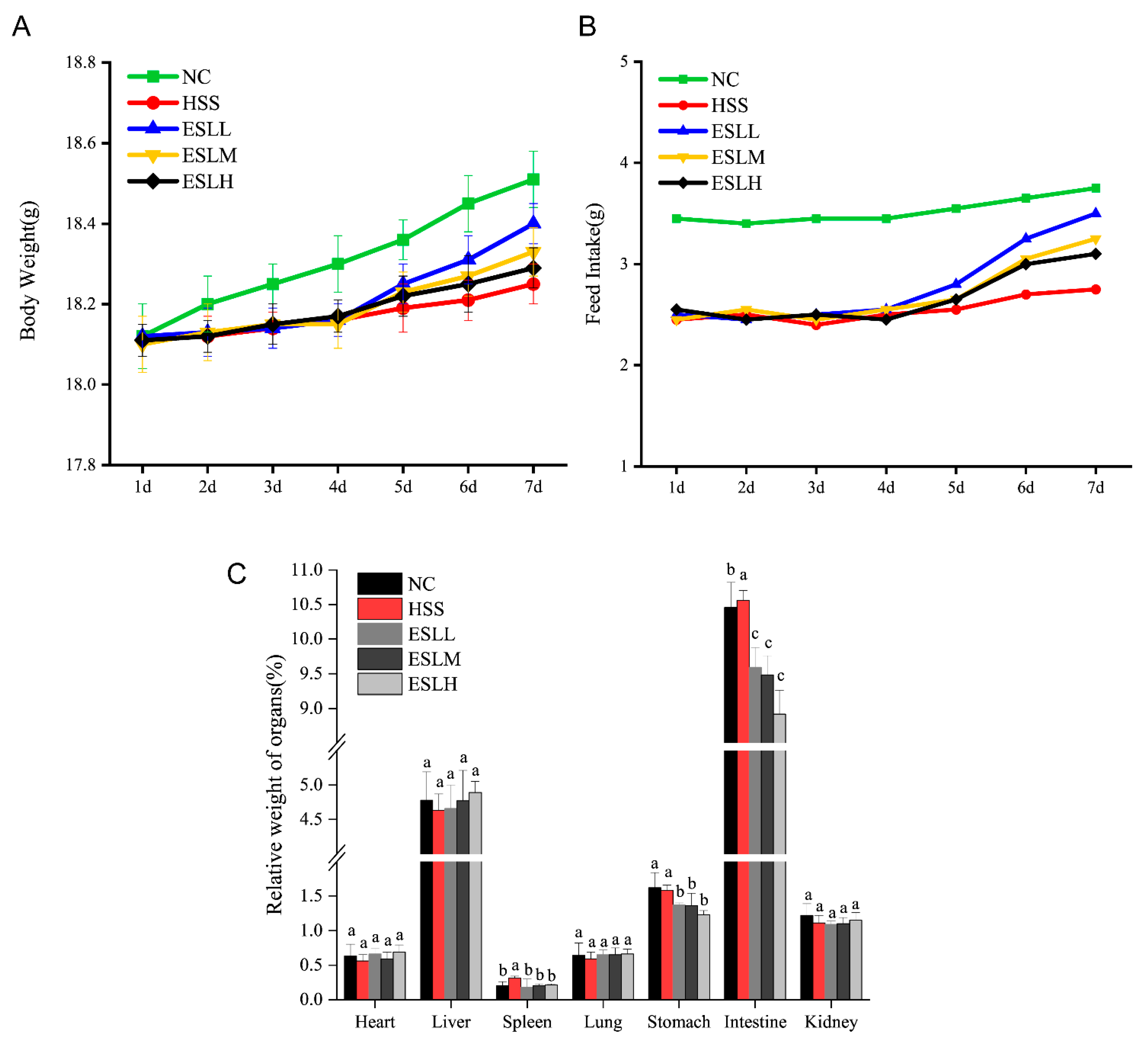
3.2. Effect of ESLP on the Body Weight, Food Intake, Viscera Coefficients, Body Surface Temperature and Physiological Behaviour of Mice
3.3. Effects of ESLP on Inflammatory Markers and LPS on Heat-Stressed Mice
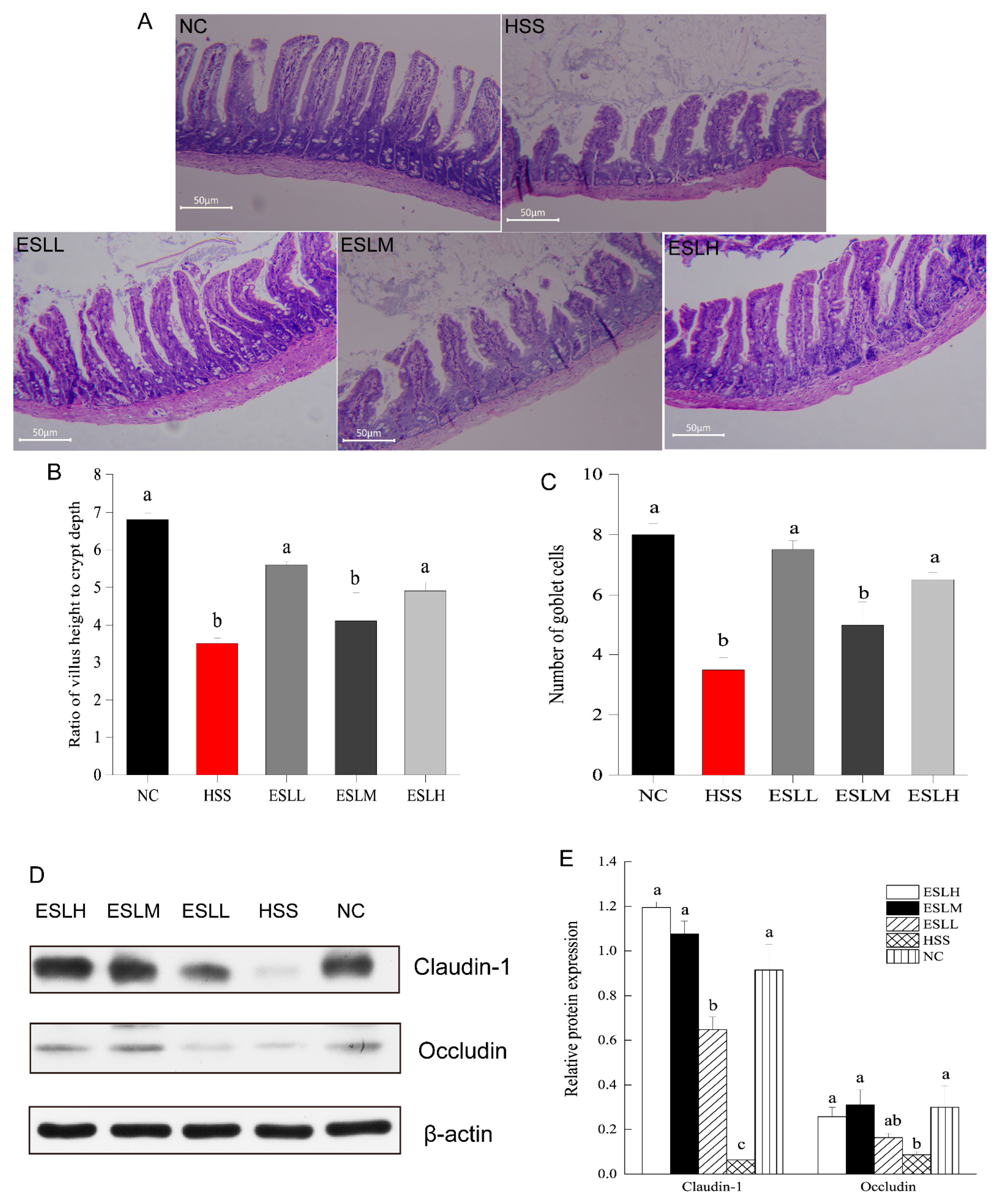
3.4. Effect of ESLP Treatment on Intestinal Tissue
3.5. Effect of ESLP Treatment on the Structure and Function of the Gut Microbiota in Heat-Stressed Mice
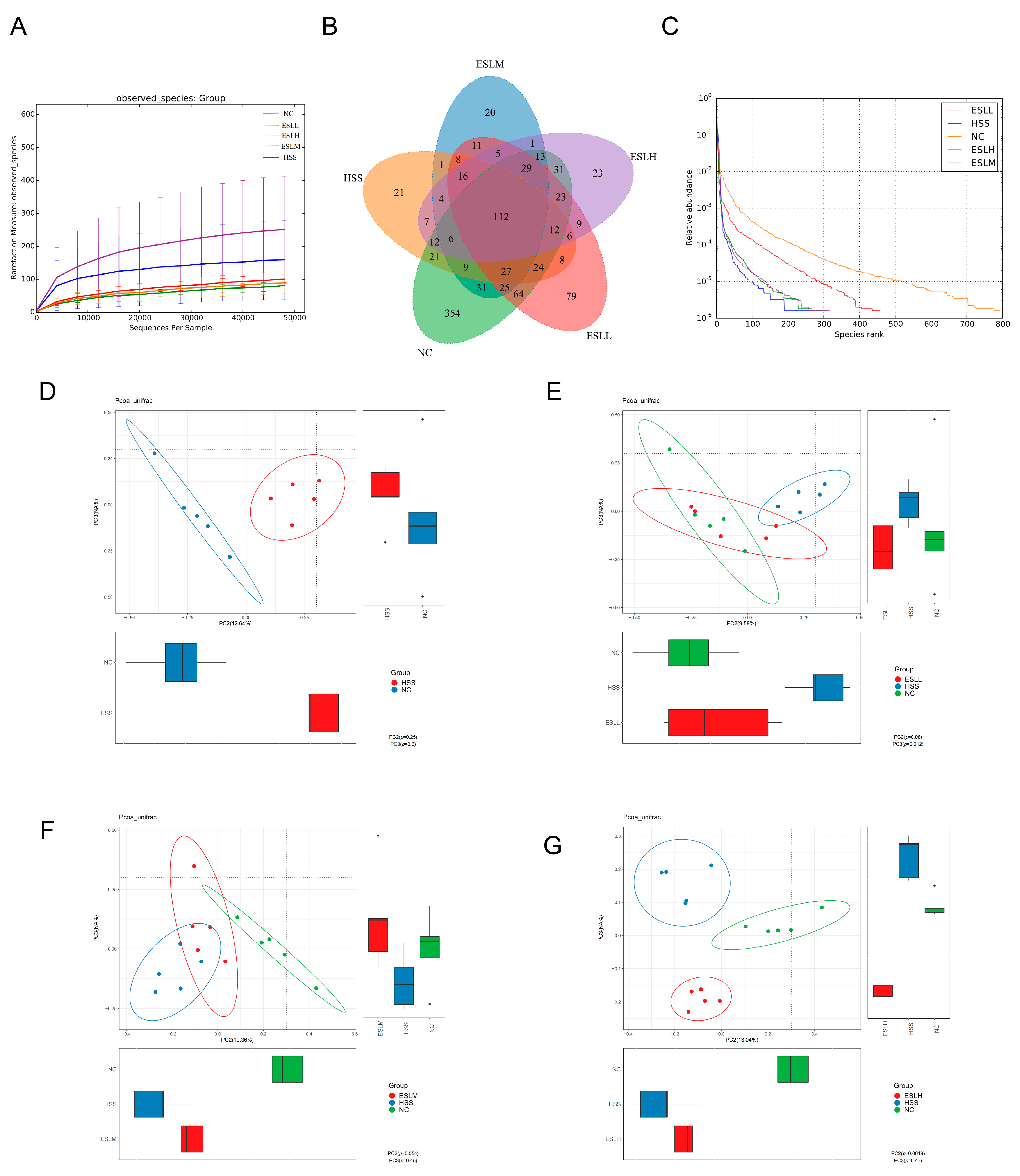
3.5.1. ESLP Maintained the Diversity of the Gut Microbiota in Heat-Stressed Mice
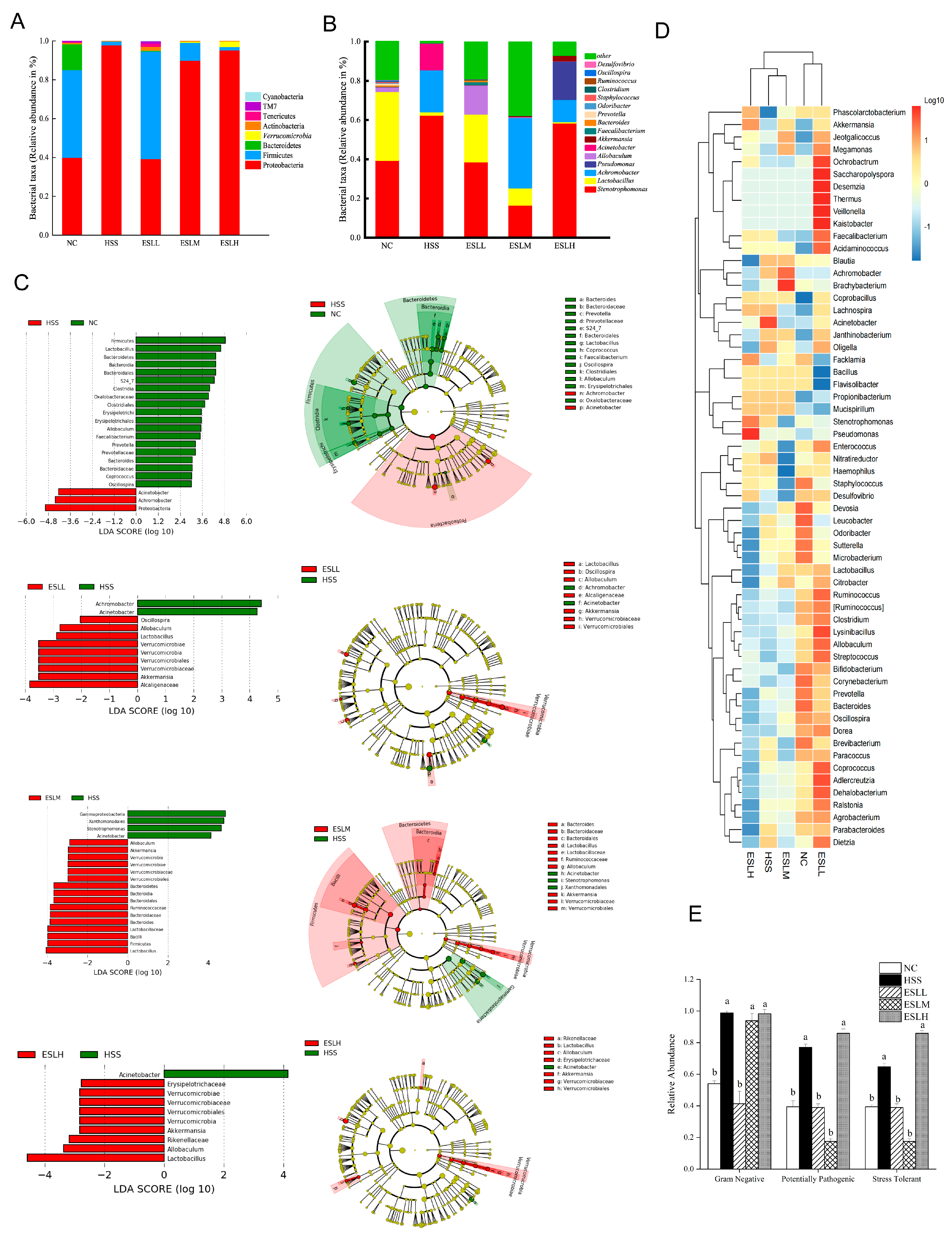
3.5.2. ESLP Maintained the Composition of the Gut Microbiota in Heat-Stressed Mice
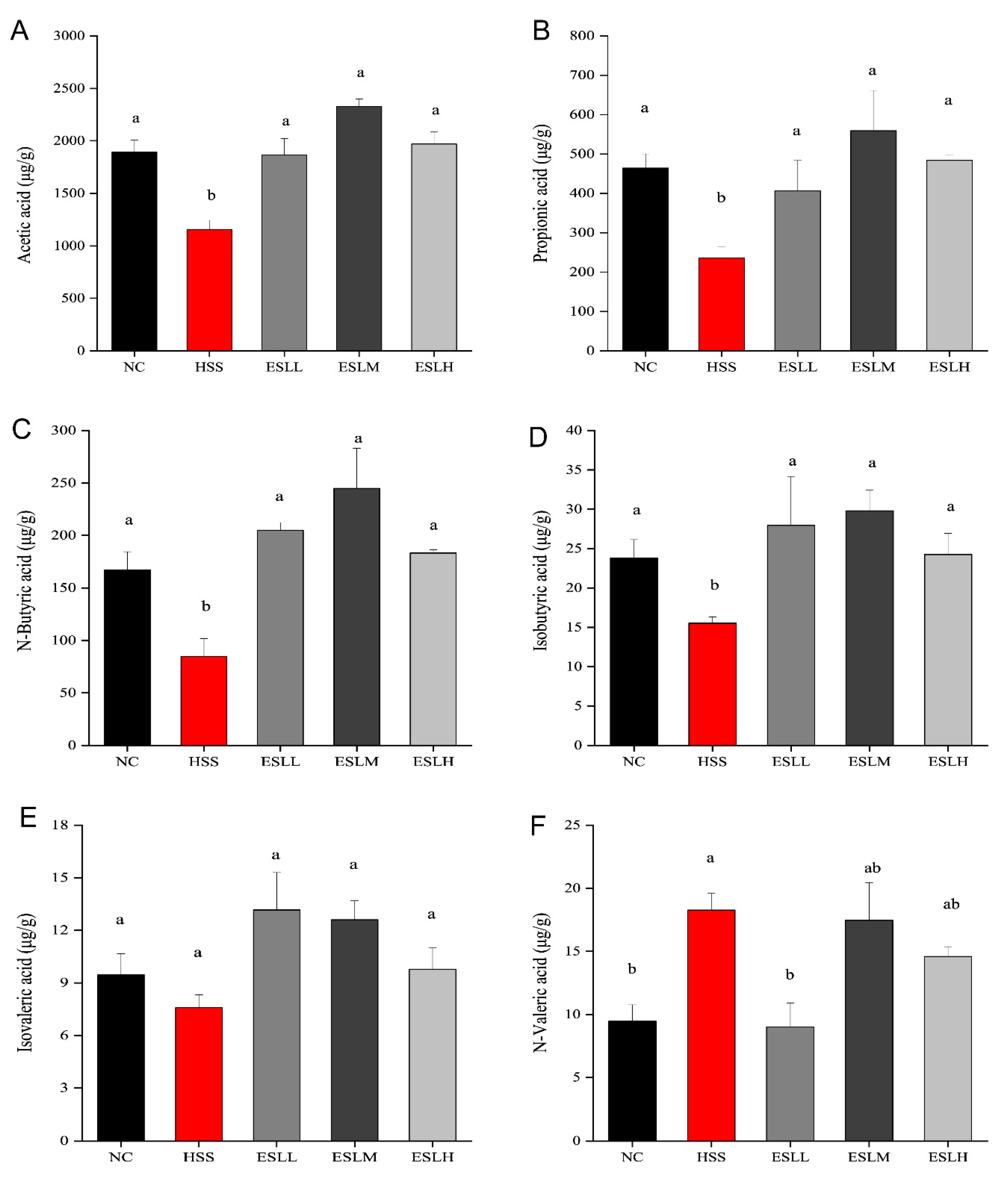
3.6. Effects of ESLP on SCFAs in Heat-Stressed Mice
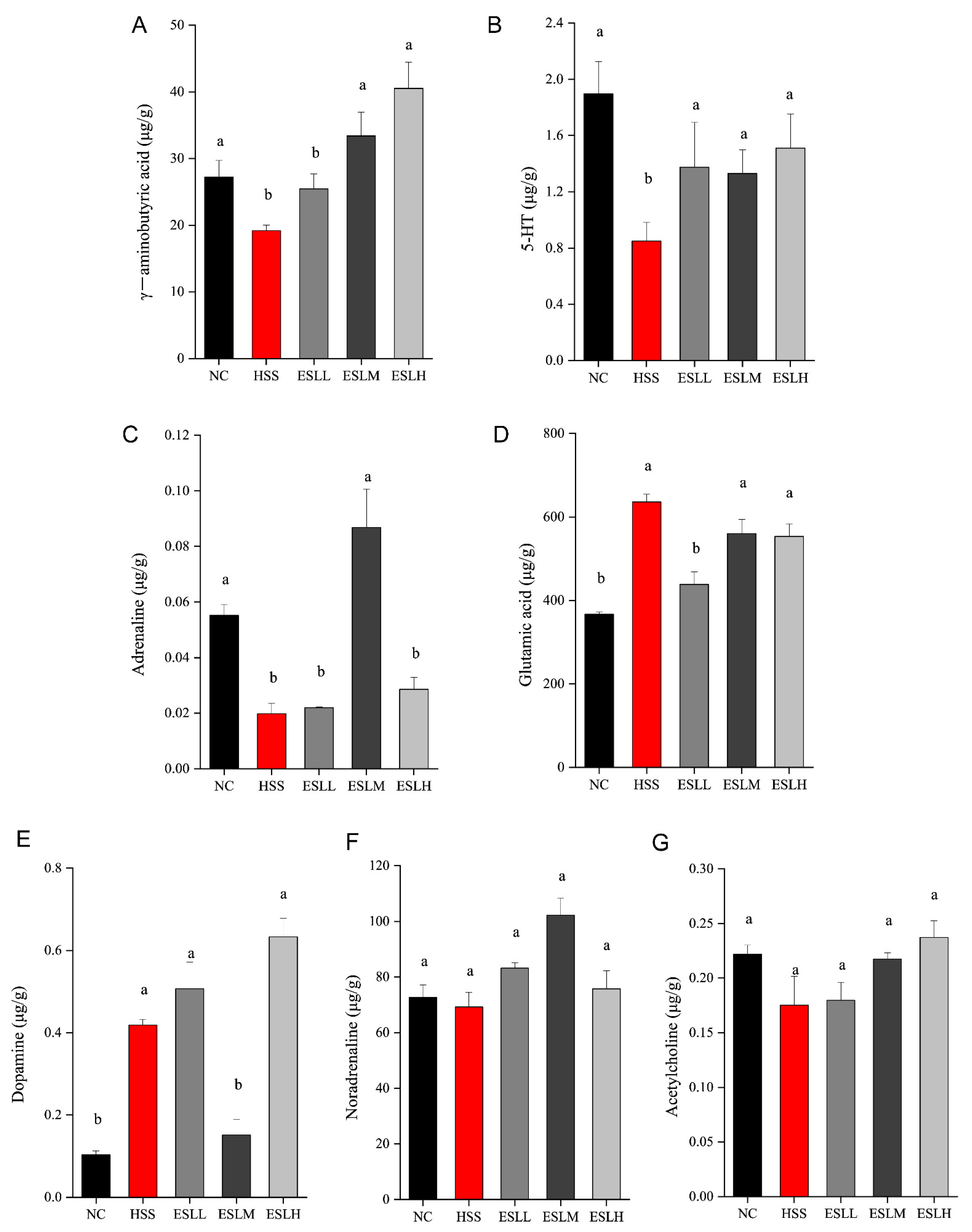
3.7. Effects of ESLP on Neurotransmitters in Heat-Stressed Mice
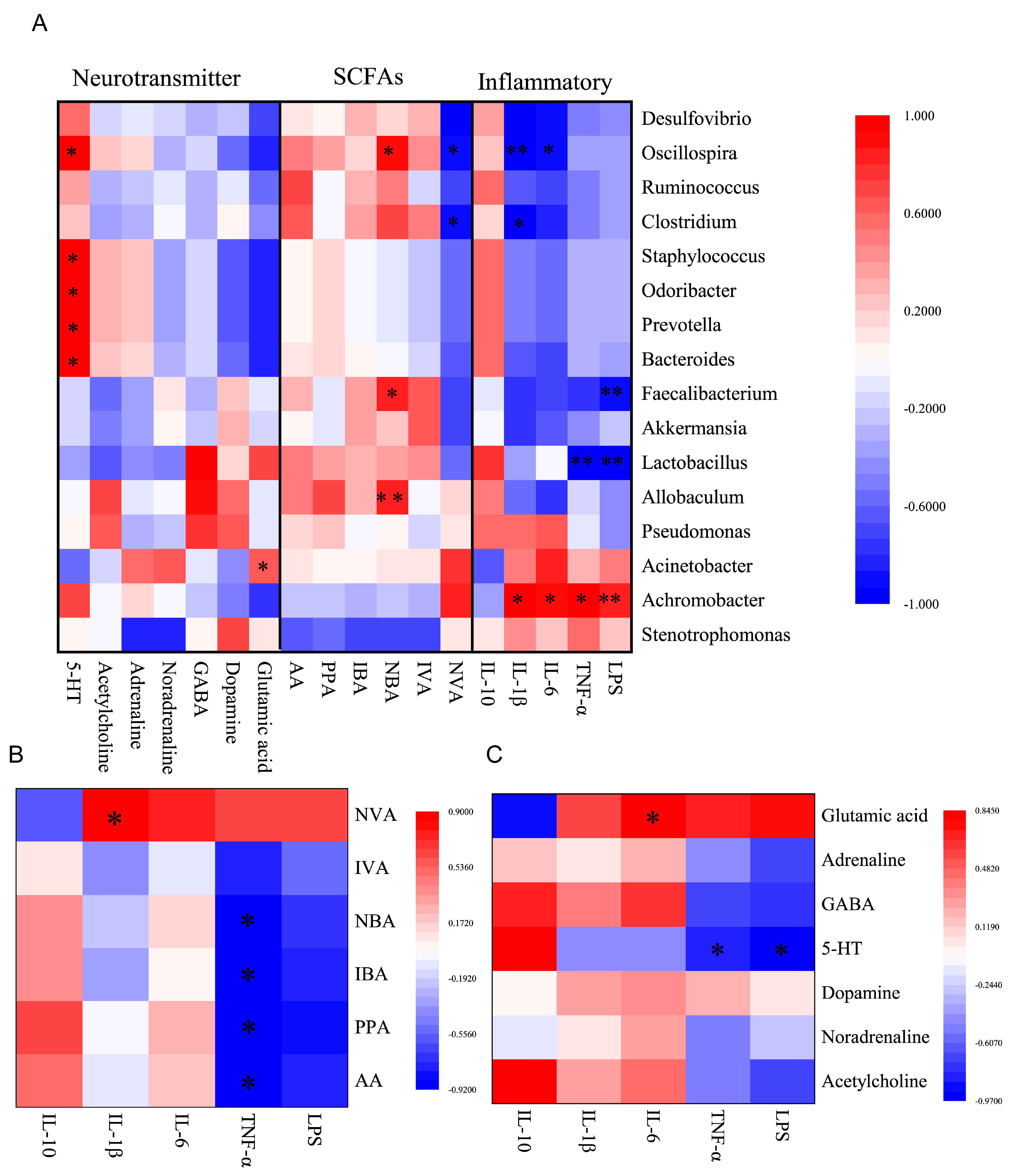
3.8. Correlation Analysis of Inflammation Markers, SCFAs, Neurotransmitters and Gut Microbiota
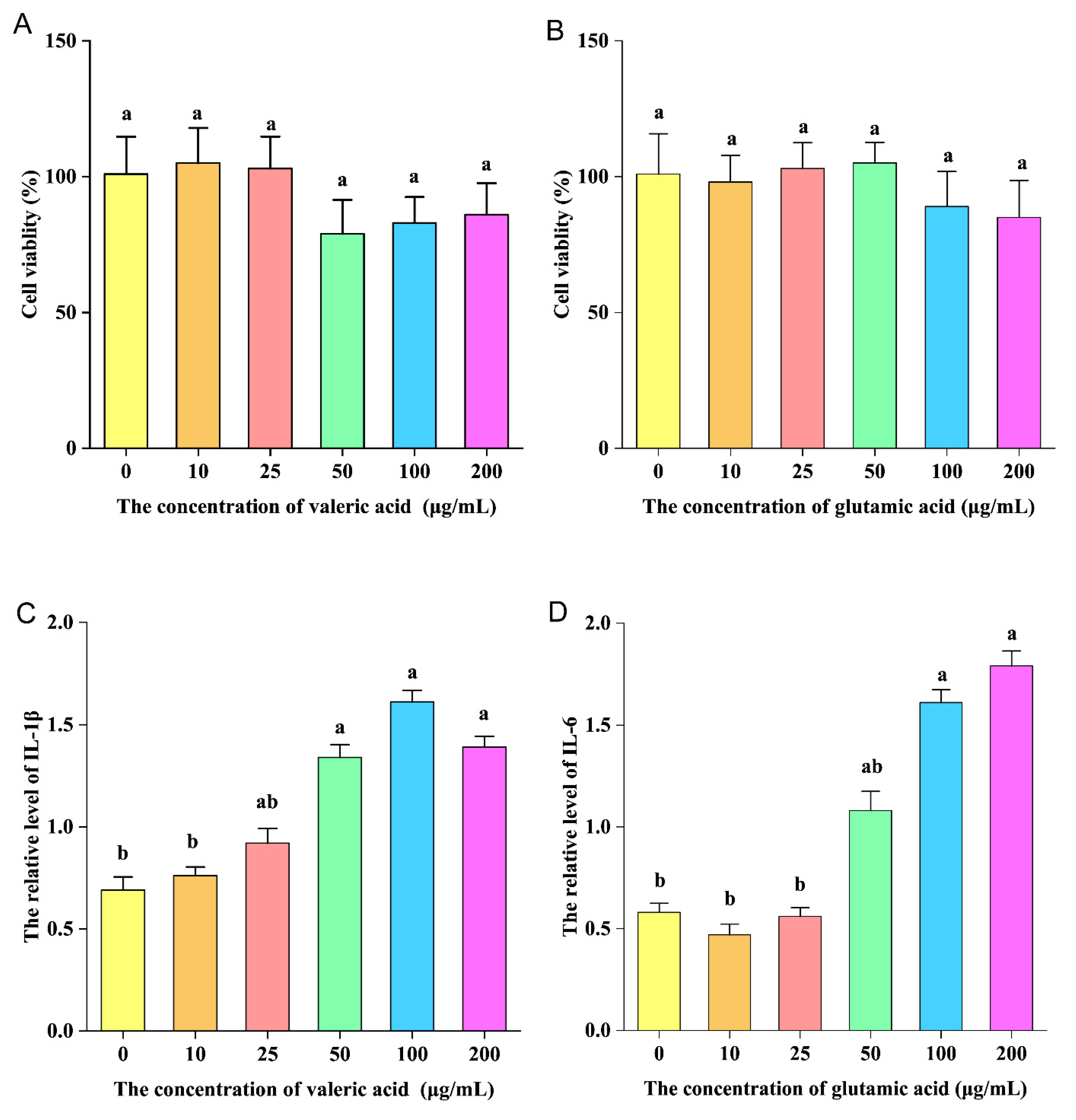
3.9. Pro-Inflammatory Activity of Valeric and Glutamic Acid In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cantet, J.M.; Yu, Z.; Ríus, A.G. Heat Stress-Mediated Activation of Immune-Inflammatory Pathways. Antibiotics 2021, 10, 1285. [Google Scholar] [CrossRef] [PubMed]
- Khajehnasiri, F.; Akhondzadeh, S.; Mortazavi, S.B.; Allameh, A.; Khavanin, A.; Zamanian, Z. Oxidative Stress and Depression among Male Shift Workers in Shahid Tondgouyan Refinery. Iran. J. Psychiatry 2014, 4, 11–20. [Google Scholar]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. Heat stress as a predisposing factor for necrotic enteritis in broiler chicks. Avian Pathol. 2018, 47, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Presbitero, A.; Melnikov, V.R.; Krzhizhanovskaya, V.V.; Sloot, P.M.A. A unifying model to estimate the effect of heat stress in the human innate immunity during physical activities. Sci. Rep. 2021, 11, 16688. [Google Scholar] [CrossRef] [PubMed]
- Pease, S.; Bouadma, L.; Kermarrec, N.; Schortgen, F.; Régnier, B.; Wolff, M. Early organ dysfunction course, cooling time and outcome in classic heatstroke. Intensive Care Med. 2009, 35, 1454–1458. [Google Scholar] [CrossRef] [PubMed]
- Hiradeve, S.M.; Rangari, V.D. Elephantopus scaber Linn.: A review on its ethnomedical, phytochemical and pharmacological profile. J. Appl. Biomed. 2014, 12, 49–61. [Google Scholar] [CrossRef]
- Chan, C.K.; Supriady, H.; Goh, B.H.; Kadir, H.A. Elephantopus scaber induces apoptosis through ROS-dependent mitochondrial signaling pathway in HCT116 human colorectal carcinoma cells. J. Ethnopharmacol. 2015, 168, 291–304. [Google Scholar] [CrossRef]
- Liu, H.W.; Dong, X.F.; Tong, J.M.; Zhang, Q. Alfalfa polysaccharides improve the growth performance and antioxidant status of heat-stressed rabbits. Livest. Sci. 2010, 131, 88–93. [Google Scholar] [CrossRef]
- Sohail, M.U.; Ijaz, A.; Younus, M.; Shabbir, M.Z.; Kamran, Z.; Ahmad, S.; Anwar, H.; Yousaf, M.S.; Ashraf, K.; Shahzad, A. Effect of supplementation of mannan oligosaccharide and probiotic on growth performance, relative weights of viscera, and population of selected intestinal bacteria in cyclic heat-stressed broilers. J. Appl. Poult. Res. 2013, 22, 485–491. [Google Scholar] [CrossRef]
- Xu, D.; Li, W.; Huang, Y.; He, J.; Tian, Y. The effect of selenium and polysaccharide of Atractylodes macrocephala Koidz.(PAMK) on immune response in chicken spleen under heat stress. Biol. Trace Elem. Res. 2014, 160, 232–237. [Google Scholar] [CrossRef]
- Xu, D.; Li, B.; Cao, N.; Li, W.; Tian, Y.; Huang, Y. The protective effects of polysaccharide of Atractylodes macrocephala Koidz (PAMK) on the chicken spleen under heat stress via antagonizing apoptosis and restoring the immune function. Oncotarget 2017, 8, 70394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial effect of intestinal fermentation of natural polysaccharides. Nutrients 2018, 10, 1055. [Google Scholar] [CrossRef]
- Ge, Y.; Ahmed, S.; Yao, W.; You, L.; Zheng, J.; Hileuskaya, K. Regulation effects of indigestible dietary polysaccharides on intestinal microflora: An overview. J. Food Biochem. 2021, 45, e13564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, J.; Cheng, H.; Wang, H.; Tan, Y.; Feng, W.; Peng, C. Interactions between polysaccharides and gut microbiota: A metabolomic and microbial review. Food Res. Int. 2022, 160, 111653. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Wang, Y.; Huang, L.; Shen, M.; Yu, Y.; Yu, Q.; Chen, Y.; Xie, J. Review of the relationships among polysaccharides, gut microbiota, and human health. Food Res. Int. 2021, 140, 109858. [Google Scholar] [CrossRef]
- Shang, Q.; Song, G.; Zhang, M.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Xie, F.; He, H.; Johnston, L.J.; Dai, X.; Wu, C.; Ma, X. Dietary fiber-derived short-chain fatty acids: A potential therapeutic target to alleviate obesity-related nonalcoholic fatty liver disease. Obes. Rev. 2021, 22, e13316. [Google Scholar] [CrossRef]
- Ji, X.; Hou, C.; Zhang, X.; Han, L.; Yin, S.; Peng, Q.; Wang, M. Microbiome-metabolomic analysis of the impact of Zizyphus jujuba cv. Muzao polysaccharides consumption on colorectal cancer mice fecal microbiota and metabolites. Int. J. Biol. Macromol. 2019, 131, 1067–1076. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Tan, Y.; Ao, H.; Wang, J.; Peng, C. Polysaccharides from Atractylodes macrocephala Koidz. Ameliorate ulcerative colitis via extensive modification of gut microbiota and host metabolism. Food Res. Int. 2020, 138, 109777. [Google Scholar] [CrossRef]
- Chen, P.; Hei, M.; Kong, L.; Liu, Y.; Yang, Y.; Mu, H.; Zhang, X.; Zhao, S.; Duan, J. One water-soluble polysaccharide from Ginkgo biloba leaves with antidepressant activities via modulation of the gut microbiome. Food Funct. 2019, 10, 8161–8171. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Huang, G.; Chen, G. Extraction, structural analysis, derivatization and antioxidant activity of polysaccharide from Chinese yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Brewer, C.F. Determination of the concentrations of oligosaccharides, complex type carbohydrates, and glycoproteins using the phenol-sulfuric acid method. Carbohydr. Res. 1994, 254, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wang, C.; Sun, L.; Hu, L.; Fang, Z.; Deng, Q.; Zhao, J.; Gooneratne, R. Preliminary Report on Intestinal Flora Disorder, Faecal Short-Chain Fatty Acid Level Decline and Intestinal Mucosal Tissue Weakening Caused by Litchi Extract to Induce Systemic Inflammation in HFA Mice. Nutrients 2022, 14, 776. [Google Scholar] [CrossRef]
- Iqbal, H.; Kim, S.K.; Cha, K.M.; Jeong, M.S.; Ghosh, P.; Rhee, D.K. Korean Red Ginseng alleviates neuroinflammation and promotes cell survival in the intermittent heat stress-induced rat brain by suppressing oxidative stress via estrogen receptor beta and brain-derived neurotrophic factor upregulation. J. Ginseng. Res. 2020, 44, 593–602. [Google Scholar] [CrossRef]
- Ducray, H.A.G.; Globa, L.; Pustovyy, O.; Morrison, E.; Vodyanoy, V.; Sorokulova, I. Yeast fermentate prebiotic improves intestinal barrier integrity during heat stress by modulation of the gut microbiota in rats. J. Appl. Microbiol. 2019, 127, 1192–1206. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, L.; Zhang, P.; Zhou, Y.; Huang, X.; Yan, Q.; Tan, Z.; Tang, S.; Wan, F. Alterations in nutrient digestibility and performance of heat-stressed dairy cows by dietary L-theanine supplementation. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2022, 11, 350–358. [Google Scholar] [CrossRef]
- Wang, L.; An, J.; Song, S.; Mei, M.; Li, W.; Ding, F.; Liu, S. Electroacupuncture preserves intestinal barrier integrity through modulating the gut microbiota in DSS-induced chronic colitis. Life Sci. 2020, 261, 118473. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Song, J.; Li, Y.; Zhou, L.; Xu, H.; Wu, K.; Gao, J.; Zhao, M.; Zheng, Y. Bergamot polysaccharides relieve DSS-induced ulcerative colitis via regulating the gut microbiota and metabolites. Int. J. Biol. Macromol. 2023, 253, 127335. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.; Li, X.; Zhang, X.; Huang, Y.; Fei, Q.; Han, Y.; Cai, R.; Gao, Y.; Qi, Y. Ethanol extract of Elephantopus scaber Linn. Attenuates inflammatory response via the inhibition of NF-κB signaling by dampening p65-DNA binding activity in lipopolysaccharide-activated macrophages. J. Ethnopharmacol. 2020, 250, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Doan, H.V.; Hoseinifar, S.H.; Sringarm, K.; Jaturasitha, S.; Khamlor, T.; Dawood, M.A.O.; Esteban, M.Á.; Soltani, M.; Musthafa, M.S. Effects of elephant’s foot (Elephantopus scaber) extract on growth performance, immune response, and disease resistance of nile tilapia (Oreochromis niloticus) fingerlings. Fish Shellfish Immunol. 2019, 93, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Cui, H.; Cheng, B.; Fang, S.; Xu, J.; Gu, Q. Chemical constituents from the roots of Elephantopus scaber L. Biochem. Syst. Ecol. 2014, 54, 65–67. [Google Scholar] [CrossRef]
- Ho Do, M.; Seo, Y.S.; Park, H.Y. Polysaccharides: Bowel health and gut microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 1212–1224. [Google Scholar] [CrossRef]
- Gan, L.; Wang, J.; Guo, Y. Polysaccharides influence human health via microbiota-dependent and -independent pathways. Front. Nutr. 2022, 9, 1030063. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, Y.; Wen, C.; Zhou, Y. Protective effects of dietary mannan oligosaccharide on heat stress-induced hepatic damage in broilers. Environ. Sci. Pollut. Res. Int. 2020, 27, 29000–29008. [Google Scholar] [CrossRef]
- Yu, W.; Zeng, D.; Xiong, Y.; Shan, S.; Yang, X.; Zhao, H.; Lu, W. Health benefits of functional plant polysaccharides in metabolic syndrome: An overview. J. Funct. Foods 2022, 95, 105154. [Google Scholar] [CrossRef]
- Snavely, E.A.; Precit, M. It’s Bordetella, It’s Alcaligenes… No, It’s Achromobacter! Identification, Antimicrobial Resistance, and Clinical Significance of an Understudied Gram-Negative Rod. Clin. Microbiol. Newsl. 2022, 44, 169–177. [Google Scholar] [CrossRef]
- Xu, J.; Molin, G.; Davidson, S.; Roth, B.; Sjöberg, K. CRP in Outpatients with Inflammatory Bowel Disease Is Linked to the Blood Microbiota. Int. J. Mol. Sci. 2023, 24, 10899. [Google Scholar] [CrossRef]
- Vicente, A.R.; Ayala-Rodriguez, C.; Colón-Núñez, C.; De Jesus, G.G.; Pabon, M.A.M. From the Gut to the Heart: Purulent Pericarditis Due to Achromobacter Spp. Bacteremia in a Patient with Systemic Lupus Erythematosus (Sle) and Strongyloides Stercoralis Infection. J. Am. Coll. Cardiol. 2023, 81, 3015. [Google Scholar] [CrossRef]
- Neidhöfer, C.; Berens, C.; Parčina, M. An 18-Year Dataset on the Clinical Incidence and MICs to Antibiotics of Achromobacter spp. (Labeled Biochemically or by MAL-DI-TOF MS as A. xylosoxidans), Largely in Patient Groups Other than Those with CF. Antibiotics 2022, 11, 311. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Liu, J.; Ao, H.; Yue, S.; Peng, C. Targeting gut microbiota for precision medicine: Focusing on the efficacy and toxicity of drugs. Theranostics 2020, 10, 11278–11301. [Google Scholar] [CrossRef] [PubMed]
- Rudiansyah, M.; Abdalkareem Jasim, S.; Azizov, B.S.; Samusenkov, V.; Kamal Abdelbasset, W.; Yasin, G.; Mohammad, H.J.; Jawad, M.A.; Mahmudiono, T.; Hosseini-Fard, S.R. The emerging microbiome-based approaches to IBD therapy: From SCFAs to urolithin A. J. Dig. Dis. 2022, 23, 412–434. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Tian, H.Z. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 11–21. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; Mf, N.M.; Ojha, S.K.; Adem, A. Valeric Acid Protects Dopaminergic Neurons by Suppressing Oxidative Stress, Neuroinflammation and Modulating Autophagy Pathways. Int. J. Mol. Sci. 2020, 21, 7670. [Google Scholar] [CrossRef]
- Gao, G.; Zhou, J.; Wang, H.; Ding, Y.; Zhou, J.; Chong, P.H.; Zhu, L.; Ke, L. Effects of valerate on intestinal barrier function in cultured Caco-2 epithelial cell monolayers. Mol. Biol. Rep. 2022, 49, 1817–1825. [Google Scholar] [CrossRef]
- Wu, M.; Tian, T.; Mao, Q.; Zou, T.; Chen, J.J. Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl. Psychiatry 2020, 10, 112–132. [Google Scholar] [CrossRef]
- Ahern, G.P. 5-HT and the immune system. Curr. Opin. Pharmacol. 2011, 11, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Ding, L.; Wang, D.; Han, J.; Gao, P. Serotonin: A Potent Immune Cell Modulator in Autoimmune Diseases. Front. Immunol. 2020, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Koopman, N.; Katsavelis, D. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef] [PubMed]
- de Bie, T.; Balvers, M.; Jongsma, M.; Witkamp, R. Dietary Neurotransmitters: The Relative Oral Bioavailability of GABA and Glutamic Acid From Tomato in Healthy Human Volunteers. Curr. Dev. Nutr. 2021, 5, 311. [Google Scholar] [CrossRef]
- Motaghi, S.; Sirchi, M.M.; Hosseininasab, N.s.; Abbasnejad, M.; Esmaili-Mahani, S.; Sepehri, G. Age-Related Changes in Glutamic Acid Decarboxylase 1 Gene Expression in the Medial Prefrontal Cortex and Ventral Hippocampus of Fear-Potentiated Rats Subjected to Isolation Stress. Behav. Brain Res. 2023, 453, 114630. [Google Scholar] [CrossRef]
- Coëffier, M.; Marion, R.; Leplingard, A.; Lerebours, E.; Ducrotté, P.; Déchelotte, P. Glutamine decreases interleukin-8 and interleukin-6 but not nitric oxide and prostaglandins e(2) production by human gut in-vitro. Cytokine 2002, 18, 92–97. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | NC | HSS | ESLL | ESLM | ESLH |
|---|---|---|---|---|---|
| ear base/°C | 36.07 ± 0.29 a | 36.20 ± 0.08 a | 36.12 ± 0.25 a | 36.02 ± 0.20 a | 35.96 ± 0.23 a |
| neck/°C | 36.40 ± 0.11 a | 36.46 ± 0.19 a | 36.44 ± 0.17 a | 36.46 ± 0.11 a | 36.42 ± 0.19 a |
| rectal/°C | 35.62 ± 0.10 b | 35.94 ± 0.13 a | 35.57 ± 0.06 b | 35.87 ± 0.05 a | 35.70 ± 0.25 a |
| Groups | Shannon | Simpson | Chao1 | ACE | Goods Coverage |
|---|---|---|---|---|---|
| NC | 1.960 ± 0.911 a | 0.662 ± 0.219 a | 263.414 ± 86.015 a | 265.013 ± 81.011 a | 1.000 ± 0.000 a |
| HSS | 1.457 ± 0.411 ab | 0.465 ± 0.045 a | 172.756 ± 25.611 ab | 163.025 ± 30.418 b | 1.000 ± 0.000 a |
| ESLL | 1.847 ± 1.220 a | 0.647 ± 0.111 a | 212.085 ± 100.941 ab | 203.030 ± 89.117 a | 1.000 ± 0.000 a |
| ESLM | 0.557 ± 0.139 b | 0.126 ± 0.037 b | 161.470 ± 42.782 b | 168.618 ± 37.352 b | 1.000 ± 0.000 a |
| ESLH | 1.526 ± 0.323 ab | 0.565 ± 0.028 a | 202.863 ± 25.556 ab | 191.318 ± 24.231 ab | 1.000 ± 0.000 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Sun, D.; Deng, Q.; Sun, L.; Hu, L.; Fang, Z.; Zhao, J.; Gooneratne, R. Elephantopus scaber L. Polysaccharides Alleviate Heat Stress-Induced Systemic Inflammation in Mice via Modulation of Characteristic Gut Microbiota and Metabolites. Nutrients 2024, 16, 262. https://doi.org/10.3390/nu16020262
Wang C, Sun D, Deng Q, Sun L, Hu L, Fang Z, Zhao J, Gooneratne R. Elephantopus scaber L. Polysaccharides Alleviate Heat Stress-Induced Systemic Inflammation in Mice via Modulation of Characteristic Gut Microbiota and Metabolites. Nutrients. 2024; 16(2):262. https://doi.org/10.3390/nu16020262
Chicago/Turabian StyleWang, Chen, Dongfang Sun, Qi Deng, Lijun Sun, Lianhua Hu, Zhijia Fang, Jian Zhao, and Ravi Gooneratne. 2024. "Elephantopus scaber L. Polysaccharides Alleviate Heat Stress-Induced Systemic Inflammation in Mice via Modulation of Characteristic Gut Microbiota and Metabolites" Nutrients 16, no. 2: 262. https://doi.org/10.3390/nu16020262
APA StyleWang, C., Sun, D., Deng, Q., Sun, L., Hu, L., Fang, Z., Zhao, J., & Gooneratne, R. (2024). Elephantopus scaber L. Polysaccharides Alleviate Heat Stress-Induced Systemic Inflammation in Mice via Modulation of Characteristic Gut Microbiota and Metabolites. Nutrients, 16(2), 262. https://doi.org/10.3390/nu16020262