
Network Pharmacology and Molecular Modeling Techniques in Unraveling the Underlying Mechanism of Citri Reticulatae Pericarpium aganist Type 2 Diabetic Osteoporosis

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioactive Compounds of Citri Reticulatae Pericarpium (CRP) and Screening of Effect Targets
2.2. Collection of Targets Related to Type 2 Diabetic Osteoporosis
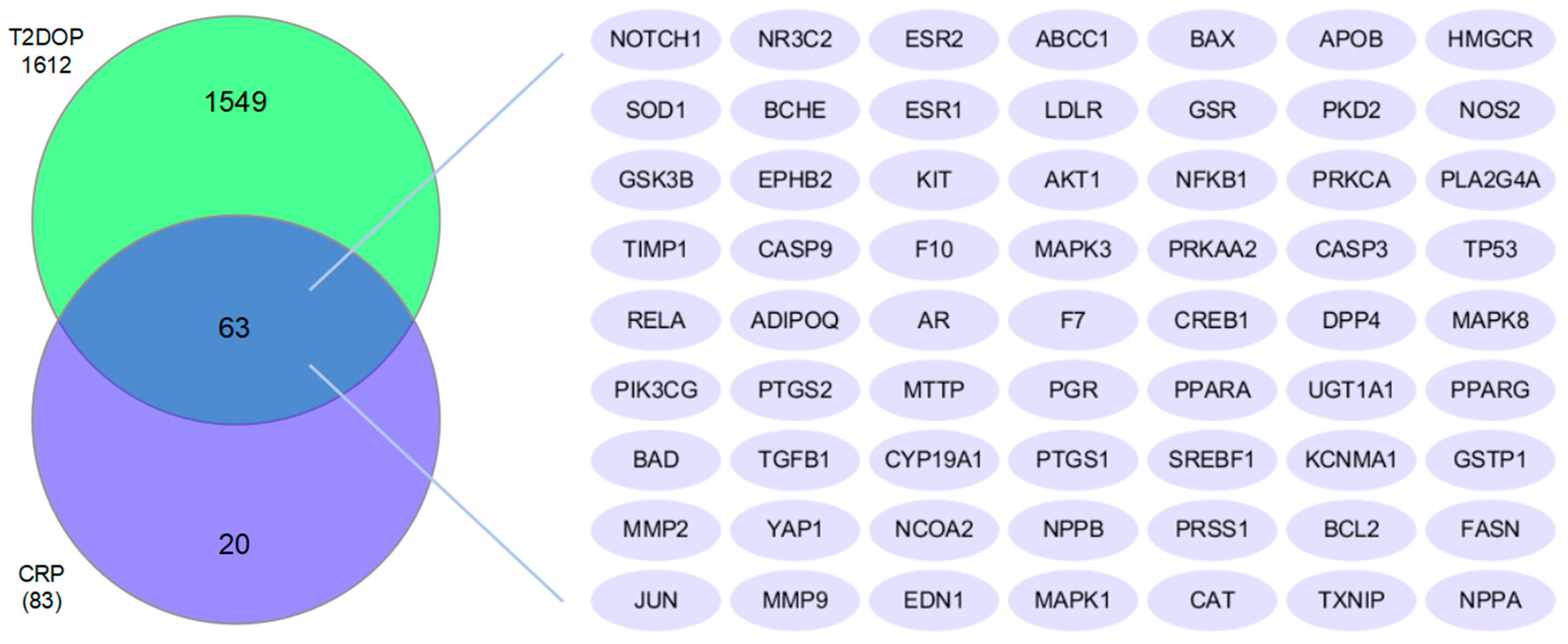
2.3. Determination of the Common Targets of Citri Reticulatae Pericarpium and T2DOP
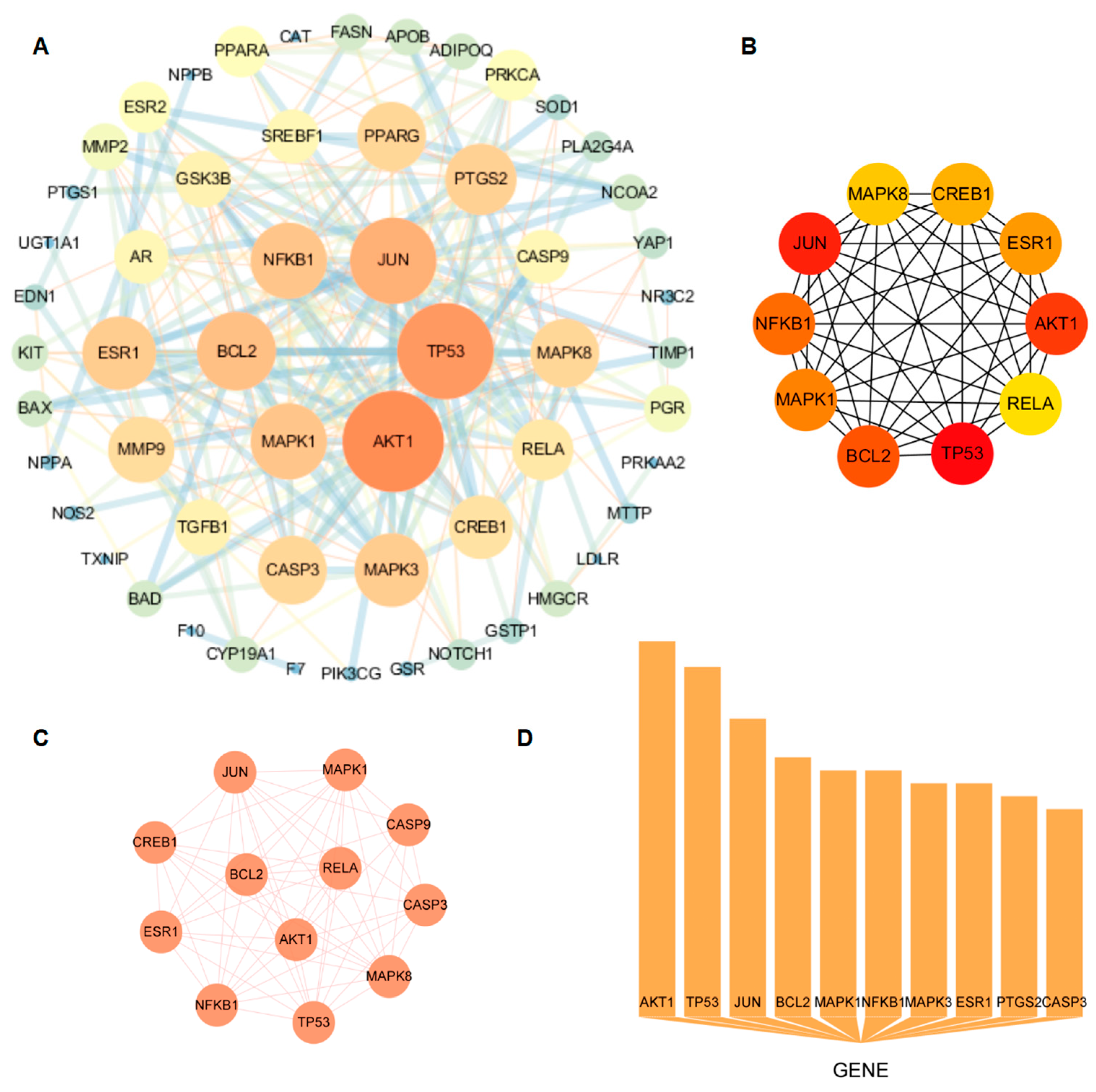
2.4. Protein–Protein Interaction Network Construction and Core Target Acquisition
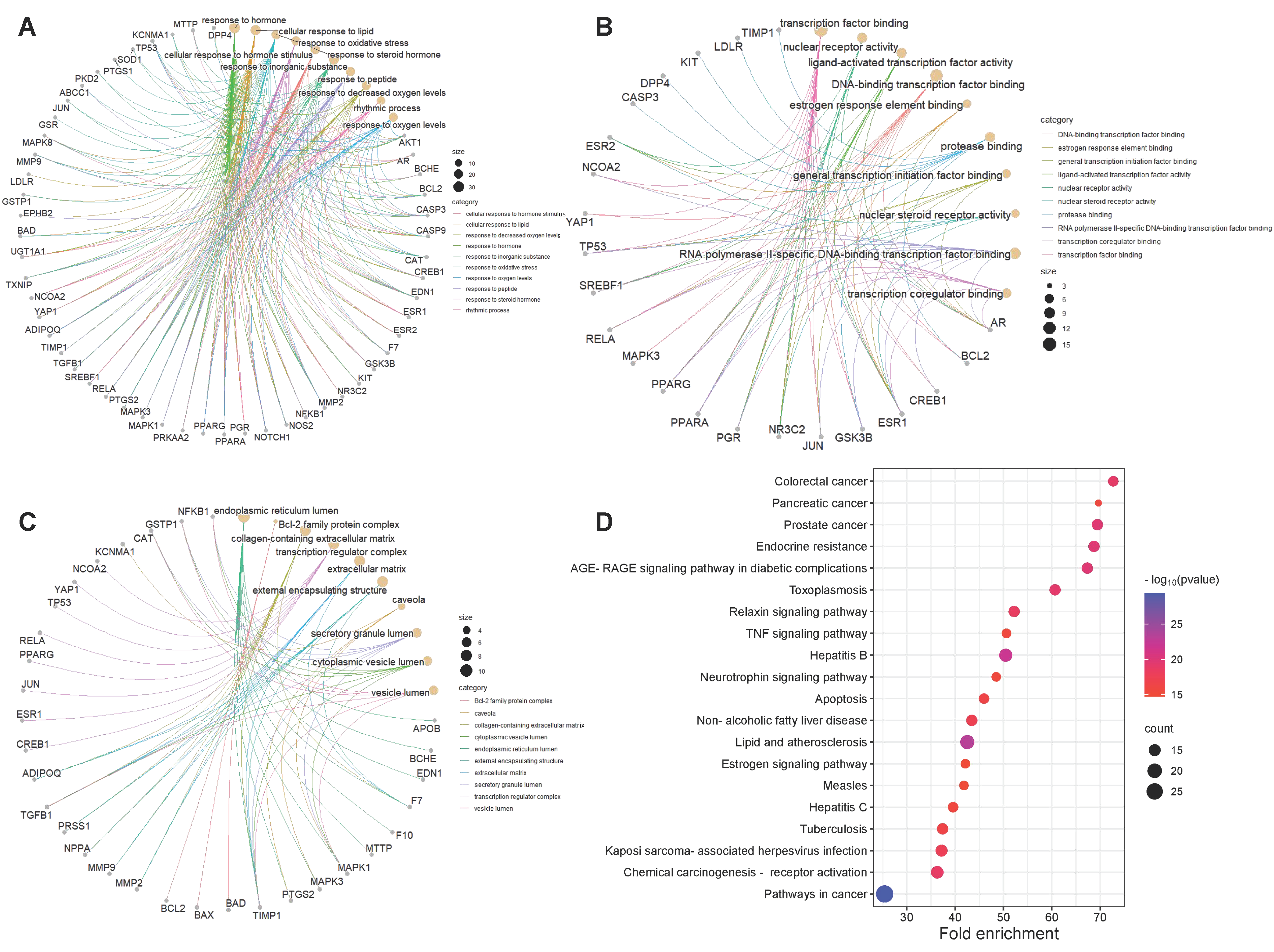
2.5. GO and KEGG Pathway Enrichment Analysis
2.6. Construction of Target–Pathway Interaction Networks
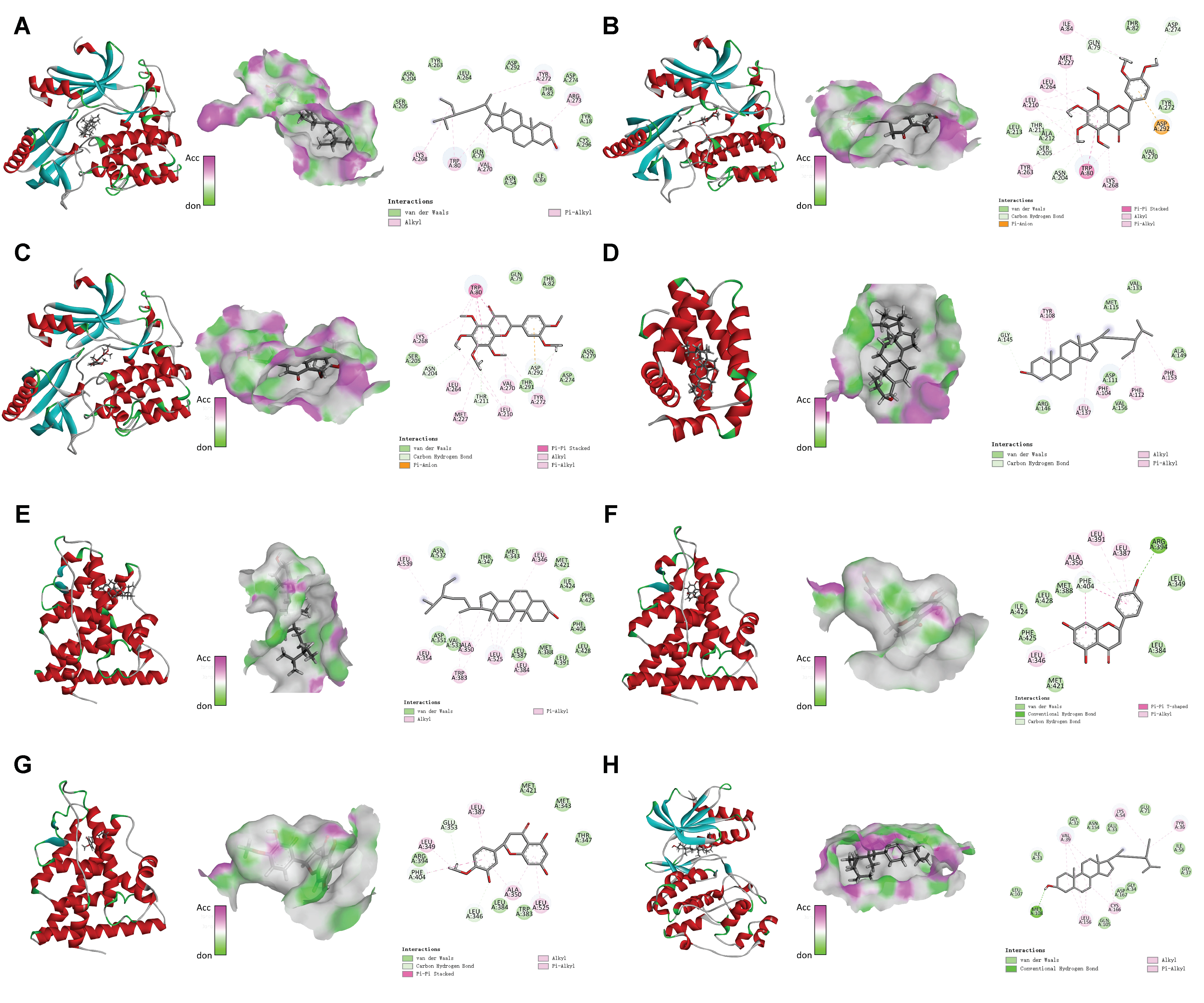
2.7. Molecular Docking
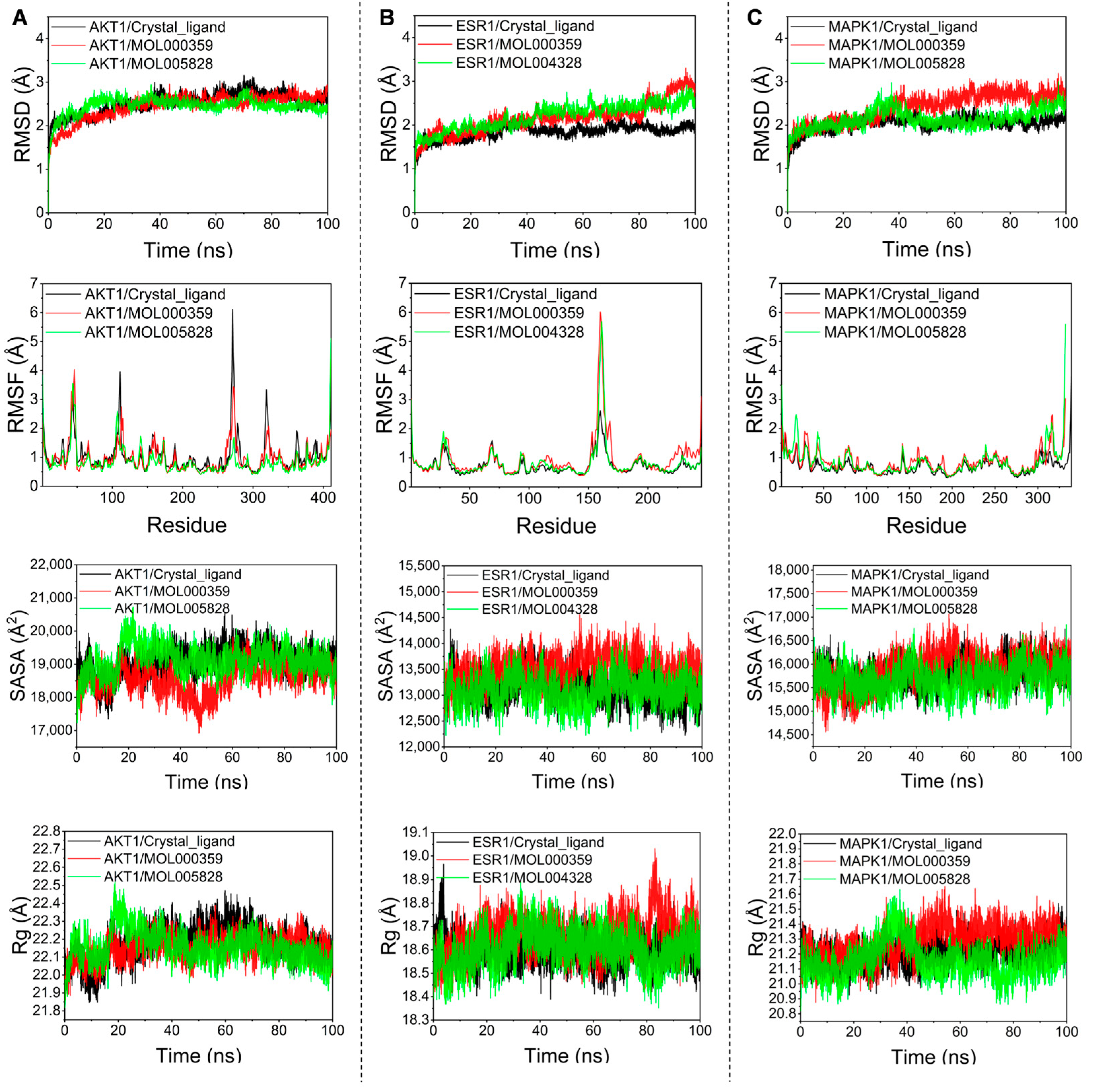
2.8. Molecular Dynamics Simulation
2.9. Binding Free Energy Calculations of MMGBSA
3. Results
3.1. The Bioactive Compounds of Citri Reticulatae Pericarpium (CRP) and Effect Target Results
3.2. Targets Related to Type 2 Diabetic Osteoporosis (T2DOP)
3.3. Common Targets of Citri Reticulatae Pericarpium and Type 2 Diabetic Osteoporosis
3.4. PPI Network and Core Target Analysis
3.5. Common-Target GO Analysis and KEGG Pathway Analysis
3.6. Molecular Docking
3.7. Molecular Dynamics Simulation
3.8. Binding Free Energy Calculations of MMGBSA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, X.; Bie, L.Y.; Pang, D.R.; Li, X.; Yang, L.F.; Chen, D.D.; Wang, Y.R.; Gao, Y. The role of autophagy in the treatment of type II diabetes and its complications: A review. Front. Endocrinol. 2023, 14, 1228045. [Google Scholar] [CrossRef] [PubMed]
- Sheu, A.; Green, J.R.; White, C.P.; Center, J.R. Contributors to impaired bone health in type 2 diabetes. Trends Endocrinol. Metab. 2023, 34, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, I.; Pantazidou, G.; Kokkalis, Z.; Georgopoulos, N.; Jelastopulu, E. Systematic Review: Are the Elderly With Diabetes Mellitus Type 2 Prone to Fragility Fractures? Cureus J. Med. Sci. 2021, 13, e14514. [Google Scholar] [CrossRef] [PubMed]
- Napoli, N.; Chandran, M.; Pierroz, D.D.; Abrahamsen, B.; Schwartz, A.V.; Ferrari, S.L.; Grp, I.O.F.B.D.W. Mechanisms of diabetes mellitus-induced bone fragility. Nat. Rev. Endocrinol. 2017, 13, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.Y.; Xiong, Z.; Liu, K.; Chang, Y.; Shu, L.; Gao, G.; Zhang, C. Identification of kaempferol as an OSX upregulator by network pharmacology-based analysis of qianggu Capsule for osteoporosis. Front. Pharmacol. 2022, 13, 1011561. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, X.; Yu, R. Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation. Nutrients 2023, 15, 4294. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Zhou, C.; Khan, H.; Fu, M.; Cheang, W.S. Citri Reticulatae Pericarpium (Chenpi) Protects against Endothelial Dysfunction and Vascular Inflammation in Diabetic Rats. Nutrients 2022, 14, 5221. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Sun, S.; Guo, Y.; Liu, Y.; Yang, D.; Li, G.; Lu, S. Citri Reticulatae Pericarpium (Chenpi): Botany, ethnopharmacology, phytochemistry, and pharmacology of a frequently used traditional Chinese medicine. J. Ethnopharmacol. 2018, 220, 265–282. [Google Scholar] [CrossRef]
- Zou, J.; Wang, J.; Ye, W.; Lu, J.; Li, C.; Zhang, D.; Ye, W.; Xu, S.; Chen, C.; Liu, P.; et al. Citri Reticulatae Pericarpium (Chenpi): A multi-efficacy pericarp in treating cardiovascular diseases. Biomed. Pharmacother. 2022, 154, 113626. [Google Scholar] [CrossRef]
- Yari, Z.; Movahedian, M.; Imani, H.; Alavian, S.M.; Hedayati, M.; Hekmatdoost, A. The effect of hesperidin supplementation on metabolic profiles in patients with metabolic syndrome: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Nutr. 2020, 59, 2569–2577. [Google Scholar] [CrossRef]
- Martin, B.R.; McCabe, G.P.; McCabe, L.; Jackson, G.S.; Horcajada, M.N.; Offord-Cavin, E.; Peacock, M.; Weaver, C.M. Effect of Hesperidin With and Without a Calcium (Calcilock) Supplement on Bone Health in Postmenopausal Women. J. Clin. Endocrinol. Metab. 2016, 101, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Liu, T.; Kou, Y.; Rong, X.; Meng, L.; Cui, Y.; Gao, R.; Hu, S.; Li, M. The Synergistic Effect of Zuogui Pill and Eldecalcitol on Improving Bone Mass and Osteogenesis in Type 2 Diabetic Osteoporosis. Med.-Lith. 2023, 59, 1414. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.F.; Shao, L.H.; Wu, X.Y.; Liu, C.; Ni, S.; Dai, T.; Liu, H.W.; Zhao, H.B. Electrospun sandwich mesh structures loaded with naringenin and vitamin K2 polycaprolactone/gelatin nanofibers synergistically promote bone regeneration. Mater. Today Bio 2023, 23, 100794. [Google Scholar] [CrossRef] [PubMed]
- Muhamad, M.L.N.; Ekeuku, S.O.; Wong, S.K.; Chin, K.Y. A Scoping Review of the Skeletal Effects of Naringenin. Nutrients 2022, 14, 4851. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Song, M.; Katsumata, S.-I.; Uehara, M.; Suzuki, K.; Ohigashi, H. Citrus nobiletin suppresses bone loss in ovariectomized ddY mice and collagen-induced arthritis in DBA/1J mice: Possible involvement of receptor activator of NF-kappaB ligand (RANKL)-induced osteoclastogenesis regulation. Biofactors 2007, 30, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.d.C.; Moura Fideles, S.O.; Bertoni Reis, C.H.; Bellini, M.Z.; Bastos Mazuqueli Pereira, E.d.S.; Pilon, J.P.G.; de Marchi, M.A.; Penteado Detregiachi, C.R.; Prync Flato, U.A.; de Moraes Trazzi, B.F.; et al. Therapeutic Effects of Citrus Flavonoids Neohesperidin, Hesperidin and Its Aglycone, Hesperetin on Bone Health. Biomolecules 2022, 12, 626. [Google Scholar] [CrossRef] [PubMed]
- Trzeciakiewicz, A.; Habauzit, V.; Mercier, S.; Barron, D.; Urpi-Sarda, M.; Manach, C.; Offord, E.; Horcajada, M.N. Molecular Mechanism of Hesperetin-7-O-glucuronide, the Main Circulating Metabolite of Hesperidin, Involved in Osteoblast Differentiation. J. Agric. Food Chem. 2010, 58, 668–675. [Google Scholar] [CrossRef]
- Li, S.; Zhang, B. Traditional Chinese medicine network pharmacology: Theory, methodology and application. Chin. J. Nat. Med. 2013, 11, 110–120. [Google Scholar] [CrossRef]
- Batool, S.; Javed, M.R.; Aslam, S.; Noor, F.; Javed, H.M.F.; Seemab, R.; Rehman, A.; Aslam, M.F.; Paray, B.A.; Gulnaz, A. Network Pharmacology and Bioinformatics Approach Reveals the Multi-Target Pharmacological Mechanism of Fumaria indica in the Treatment of Liver Cancer. Pharmaceuticals 2022, 15, 654. [Google Scholar] [CrossRef]
- Gao, Y.; Ji, W.; Lu, M.; Wang, Z.; Jia, X.; Wang, D.; Cao, P.; Hu, C.; Sun, X.; Wang, Z. Systemic pharmacological verification of Guizhi Fuling decoction in treating endometriosis-associated pain. J. Ethnopharmacol. 2022, 297, 115540. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. Bmc Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; Hartline, E.; et al. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the Amber biomolecular simulation package. Wiley Interdiscip. Rev.-Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Wang, J.M.; Wang, W.; Kollman, P.A. Antechamber: An accessory software package for molecular mechanical calculations. Abstr. Pap. Am. Chem. Soc. 2001, 222, U403. [Google Scholar]
- Wang, J.M.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Mark, P.; Nilsson, L. Structure and dynamics of the TIP3P, SPC, and SPC/E water models at 298 K. J. Phys. Chem. A 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Sagui, C.; Darden, T.A. Molecular dynamics simulations of biomolecules: Long-range electrostatic effects. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 155–179. [Google Scholar] [CrossRef]
- Kräutler, V.; Van Gunsteren, W.F.; Hünenberger, P.H. A fast SHAKE: Algorithm to solve distance constraint equations for small molecules in molecular dynamics simulations. J. Comput. Chem. 2001, 22, 501–508. [Google Scholar] [CrossRef]
- Larini, L.; Mannella, R.; Leporini, D. Langevin stabilization of molecular-dynamics simulations of polymers by means of quasisymplectic algorithms. J. Chem. Phys. 2007, 126, 104101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, Y.; Fong, P.; Mao, S.; Wang, Q. The application of the MM/GBSA method in the binding pose prediction of FGFR inhibitors. Phys. Chem. Chem. Phys. 2020, 22, 9656–9663. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Wang, J.; Li, Y.; Wang, W. Assessing the Performance of the MM/PBSA and MM/GBSA Methods. 1. The Accuracy of Binding Free Energy Calculations Based on Molecular Dynamics Simulations. J. Chem. Inf. Model. 2011, 51, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, G.; Del Rio, A.; Degliesposti, G.; Sgobba, M. Fast and Accurate Predictions of Binding Free Energies Using MM-PBSA and MM-GBSA. J. Comput. Chem. 2010, 31, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Roe, D.R.; Simmerling, C. Improved Generalized Born Solvent Model Parameters for Protein Simulations. J. Chem. Theory Comput. 2013, 9, 2020–2034. [Google Scholar] [CrossRef]
- Weiser, J.; Shenkin, P.S.; Still, W.C. Approximate atomic surfaces from linear combinations of pairwise overlaps (LCPO). J. Comput. Chem. 1999, 20, 217–230. [Google Scholar] [CrossRef]
- Hamann, C.; Kirschner, S.; Guenther, K.-P.; Hofbauer, L.C. Bone, sweet bone-osteoporotic fractures in diabetes mellitus. Nat. Rev. Endocrinol. 2012, 8, 297–305. [Google Scholar] [CrossRef]
- Zhang, Z.; Ji, C.; Wang, Y.-N.; Liu, S.; Wang, M.; Xu, X.; Zhang, D. Maresin1 Suppresses High-Glucose-Induced Ferroptosis in Osteoblasts via NRF2 Activation in Type 2 Diabetic Osteoporosis. Cells 2022, 11, 2560. [Google Scholar] [CrossRef]
- Tong, A.H.Y.; Drees, B.; Nardelli, G.; Bader, G.D.; Brannetti, B.; Castagnoli, L.; Evangelista, M.; Ferracuti, S.; Nelson, B.; Paoluzi, S.; et al. A combined experimental and computational strategy to define protein interaction networks for peptide recognition modules. Science 2002, 295, 321–324. [Google Scholar] [CrossRef]
- Hers, I.; Vincent, E.E.; Tavare, J.M. Akt signalling in health and disease. Cell. Signal. 2011, 23, 1515–1527. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Rotwein, P. Selective Signaling by Akt1 Controls Osteoblast Differentiation and Osteoblast-Mediated Osteoclast Development. Mol. Cell. Biol. 2012, 32, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, Y.; Shi, L.; He, K.; Sun, Y.; Ding, Y.; Meng, B.; Zhang, J.; Xiang, L.; Dong, J.; et al. Myeloid-derived growth factor (MYDGF) protects bone mass through inhibiting osteoclastogenesis and promoting osteoblast differentiation. Embo Rep. 2022, 23, e53509. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, W.; He, W.; Wang, H.; Chen, Q.; Song, H. Saikosaponin a inhibits RANKL-induced osteoclastogenesis by suppressing NF-κB and MAPK pathways. Int. Immunopharmacol. 2015, 25, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Li, J.; Zhang, J.; Xue, Q.; Qin, R.; Wang, R.; Goltzman, D.; Miao, D. 17β-estradiol plays the anti-osteoporosis role via a novel ESR1-Keap1-Nrf2 axis-mediated stress response activation and Tmem119 upregulation. Free. Radic. Biol. Med. 2023, 195, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Pelusi, L.; Mandatori, D.; Di Pietrantonio, N.; Del Pizzo, F.; Di Tomo, P.; Di Pietro, N.; Buda, R.; Genovese, S.; Epifano, F.; Pandolfi, A.; et al. Estrogen Receptor 1 (ESR1) and the Wnt/β-Catenin Pathway Mediate the Effect of the Coumarin Derivative Umbelliferon on Bone Mineralization. Nutrients 2022, 14, 3209. [Google Scholar] [CrossRef] [PubMed]
- Velentza, L.; Wickstrom, M.; Kogner, P.; Ohlsson, C.; Zaman, F.; Savendahl, L. Pharmacological inhibition of BCL-2 with the FDA-approved drug venetoclax impairs longitudinal bone growth. Sci. Rep. 2023, 13, 8054. [Google Scholar] [CrossRef]
- Zhang, F.; Peng, W.; Wang, T.; Zhang, J.; Dong, W.; Wang, C.; Xie, Z.; Luo, H.; Liu, G. Lnc Tmem235 promotes repair of early steroid-induced osteonecrosis of the femoral head by inhibiting hypoxia-induced apoptosis of BMSCs. Exp. Mol. Med. 2022, 54, 1991–2006. [Google Scholar] [CrossRef]
- Yu, T.; You, X.M.; Zhou, H.C.; Kang, A.; He, W.B.; Li, Z.H.; Li, B.; Xia, J.; Zhu, H.; Zhao, Y.G.; et al. p53 plays a central role in the development of osteoporosis. Aging 2020, 12, 10473–10487. [Google Scholar] [CrossRef]
- Novack, D.V. Role of NF-κB in the skeleton. Cell Res. 2011, 21, 169–182. [Google Scholar] [CrossRef]
- Feng, X.; Zhu, S.; Qiao, J.; Ji, Z.; Zhou, B.; Xu, W. CX3CL1 promotes M1 macrophage polarization and osteoclast differentiation through NF-?B signaling pathway in ankylosing spondylitis in vitro. J. Transl. Med. 2023, 21, 573. [Google Scholar] [CrossRef]
- Linares, G.R.; Xing, W.; Govoni, K.E.; Chen, S.-T.; Mohan, S. Glutaredoxin 5 regulates osteoblast apoptosis by protecting against oxidative stress. Bone 2009, 44, 795–804. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wauquier, F.; Leotoing, L.; Coxam, V.; Guicheux, J.; Wittrant, Y. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar] [CrossRef]
- Garcia-Rojas, M.D.; Palma-Cordero, G.; Martinez-Ramirez, C.O.; Ponce de Leon-Suarez, V.; Valdes-Flores, M.; Castro-Hernandez, C.; Rubio-Lightbourn, J.; Hernandez-Zamora, E.; Reyes-Maldonado, E.; Velazquez-Cruz, R.; et al. Association of Polymorphisms in Estrogen Receptor Genes (ESR1 and ESR2) with Osteoporosis and Fracture-Involvement of Comorbidities and Epistasis. DNA Cell Biol. 2022, 41, 437–446. [Google Scholar] [CrossRef]
- Asadipooya, K.; Uy, E.M. Advanced Glycation End Products (AGEs), Receptor for AGEs, Diabetes, and Bone: Review of the Literature. J. Endocr. Soc. 2019, 3, 1799–1818. [Google Scholar] [CrossRef] [PubMed]
- Roy, B. Biomolecular basis of the role of diabetes mellitus in osteoporosis and bone fractures. World J. Diabetes 2013, 4, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Taminiau, A.; Draime, A.; Tys, J.; Lambert, B.; Vandeputte, J.; Nathan, N.; Renard, P.; Geerts, D.; Rezsohazy, R. HOXA1 binds RBCK1/HOIL-1 and TRAF2 and modulates the TNF/NF-κB pathway in a transcription-independent manner. Nucleic Acids Res. 2016, 44, 7331–7349. [Google Scholar] [CrossRef][Green Version]
- Bora, K.; Sarma, M.; Kanaujia, S.P.; Dubey, V.K. Dual-target drugs against Leishmania donovani for potential novel therapeutics. Sci. Rep. 2023, 13, 18363. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mol ID | Molecule Name | OB (%) | DL |
|---|---|---|---|
| MOL000359 | sitosterol | 36.91 | 0.75 |
| MOL004328 | naringenin | 59.29 | 0.21 |
| MOL005100 | 5,7-dihydroxy-2-(3-hydroxy-4-methoxyphenyl)chroman-4-one | 47.74 | 0.27 |
| MOL005815 | Citromitin | 86.9 | 0.51 |
| MOL005828 | nobiletin | 61.67 | 0.52 |
| PDB ID | Core Gene | MOL000359 | MOL004328 | MOL005100 | MOL005815 | MOL005828 |
|---|---|---|---|---|---|---|
| 5T01 | JUN | −6.7 | −5.7 | −5.1 | −5.4 | −5.4 |
| 6S9W | AKT1 | −11.2 | −8 | −7.3 | −8.6 | −8.6 |
| 7lhb | BCL2 | −8.2 | −7.3 | −6.7 | −6.5 | −6.7 |
| 2YJA | ESR1 | −8.4 | −8.4 | −8.2 | −7.4 | −7.2 |
| 6SLG | MAPK1 | −8.7 | −7.4 | −6.7 | −7.3 | −7.6 |
| 3GUT | NFKB1 | −6.7 | −6.2 | −6 | −6 | −6.1 |
| 7DHZ | TP53 | −6.6 | −7 | −6.9 | −5.9 | −5.9 |
| System Name | AKT1/Crystal_Ligand | AKT1/MOL000359 | AKT1/MOL005828 |
|---|---|---|---|
| ΔEvdw | −68.07 ± 1.56 | −63.71 ± 1.82 | −52.51 ± 2.05 |
| ΔEelec | −27.54 ± 5.60 | −2.20 ± 2.46 | −13.59 ± 3.53 |
| ΔGGB | 67.76 ± 4.99 | 26.24 ± 1.79 | 41.47 ± 4.22 |
| ΔGSA | −8.88 ± 0.32 | −7.50 ± 0.06 | −6.42 ± 0.15 |
| ΔGbind | −36.74 ± 1.68 | −47.18 ± 2.18 | −31.05 ± 1.84 |
| System Name | ESR1/Crystal_Ligand | ESR1/MOL000359 | ESR1/MOL004328 |
|---|---|---|---|
| ΔEvdw | −43.91 ± 2.20 | −65.50 ± 1.08 | −35.34 ± 1.51 |
| ΔEelec | −8.07 ± 1.22 | −1.66 ± 0.90 | −24.76 ± 2.63 |
| ΔGGB | 18.13 ± 1.23 | 13.72 ± 0.88 | 32.16 ± 1.78 |
| ΔGSA | −5.34 ± 0.10 | −7.72 ± 0.05 | −5.37 ± 0.10 |
| ΔGbind | −39.19 ± 1.54 | −61.16 ± 0.95 | −33.31 ± 0.87 |
| System Name | MAPK1/Crystal_Ligand | MAPK1/MOL000359 | MAPK1/MOL005828 |
|---|---|---|---|
| ΔEvdw | −57.31± 2.84 | −57.09 ± 2.56 | −50.19 ± 2.73 |
| ΔEelec | −48.87 ± 10.21 | 1.65 ± 1.30 | −11.26 ± 3.86 |
| ΔGGB | 70.44± 9.18 | 27.54 ± 2.73 | 39.15 ± 2.44 |
| ΔGSA | −7.25± 0.23 | −6.73 ± 0.21 | −6.65 ± 0.26 |
| ΔGbind | −43.00± 2.11 | −34.62 ± 1.37 | −28.95 ± 2.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, Y.; Ullah, A.; Zhang, R.; Sun, Y.; Li, J.; Kou, G. Network Pharmacology and Molecular Modeling Techniques in Unraveling the Underlying Mechanism of Citri Reticulatae Pericarpium aganist Type 2 Diabetic Osteoporosis. Nutrients 2024, 16, 220. https://doi.org/10.3390/nu16020220
Li J, Wang Y, Ullah A, Zhang R, Sun Y, Li J, Kou G. Network Pharmacology and Molecular Modeling Techniques in Unraveling the Underlying Mechanism of Citri Reticulatae Pericarpium aganist Type 2 Diabetic Osteoporosis. Nutrients. 2024; 16(2):220. https://doi.org/10.3390/nu16020220
Chicago/Turabian StyleLi, Jiangtao, Ying Wang, Amin Ullah, Ruiyang Zhang, Yuge Sun, Jinjie Li, and Guangning Kou. 2024. "Network Pharmacology and Molecular Modeling Techniques in Unraveling the Underlying Mechanism of Citri Reticulatae Pericarpium aganist Type 2 Diabetic Osteoporosis" Nutrients 16, no. 2: 220. https://doi.org/10.3390/nu16020220
APA StyleLi, J., Wang, Y., Ullah, A., Zhang, R., Sun, Y., Li, J., & Kou, G. (2024). Network Pharmacology and Molecular Modeling Techniques in Unraveling the Underlying Mechanism of Citri Reticulatae Pericarpium aganist Type 2 Diabetic Osteoporosis. Nutrients, 16(2), 220. https://doi.org/10.3390/nu16020220