
Plant-Derived as Alternatives to Animal-Derived Bioactive Peptides: A Review of the Preparation, Bioactivities, Structure–Activity Relationships, and Applications in Chronic Diseases

Abstract
1. Introduction
2. Methods
3. Preparation of PDBPs
3.1. Enzymatic Hydrolysis
3.1.1. Monoenzymatic Hydrolysis
3.1.2. Complex Enzymatic Hydrolysis
3.1.3. Microbial Fermentation
3.2. Separation and Purification
4. Bioactivities of PDBPs
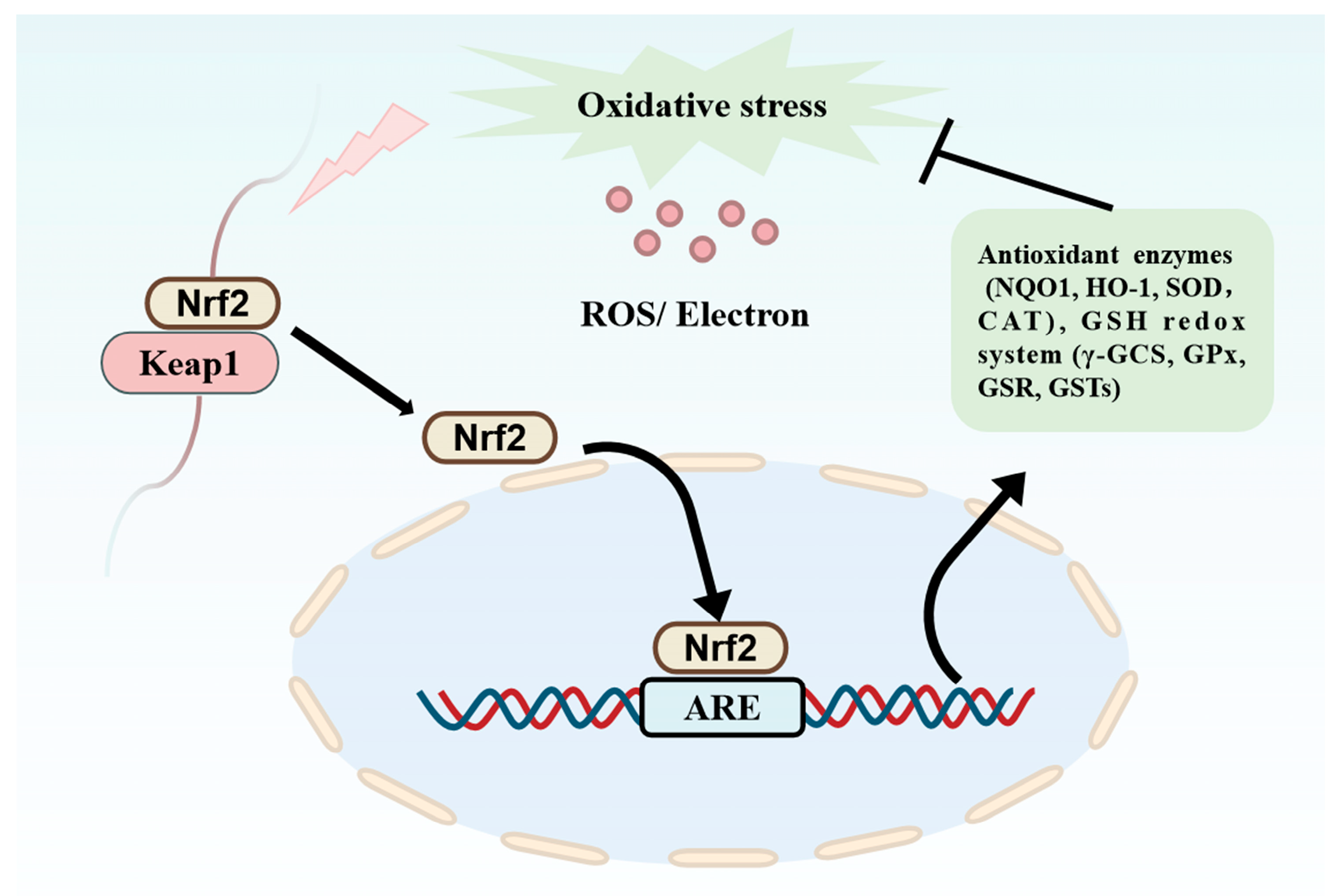
4.1. Antioxidant Activity
4.2. Antibacterial Activity
4.3. Anticancer Activity
4.4. Antihypertensive Activity
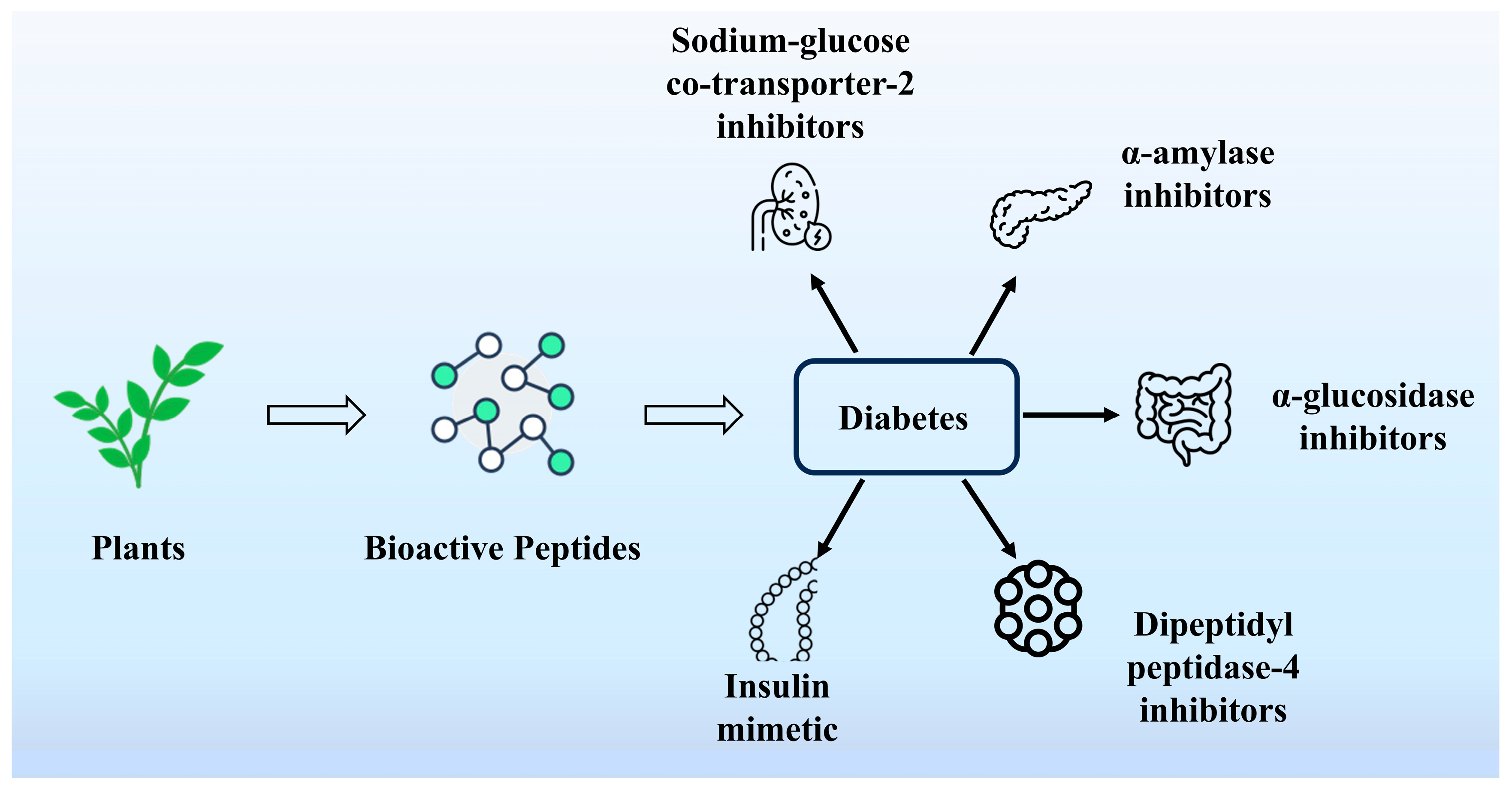
4.5. Hypoglycemic Activity
4.6. Immunoregulatory Activity
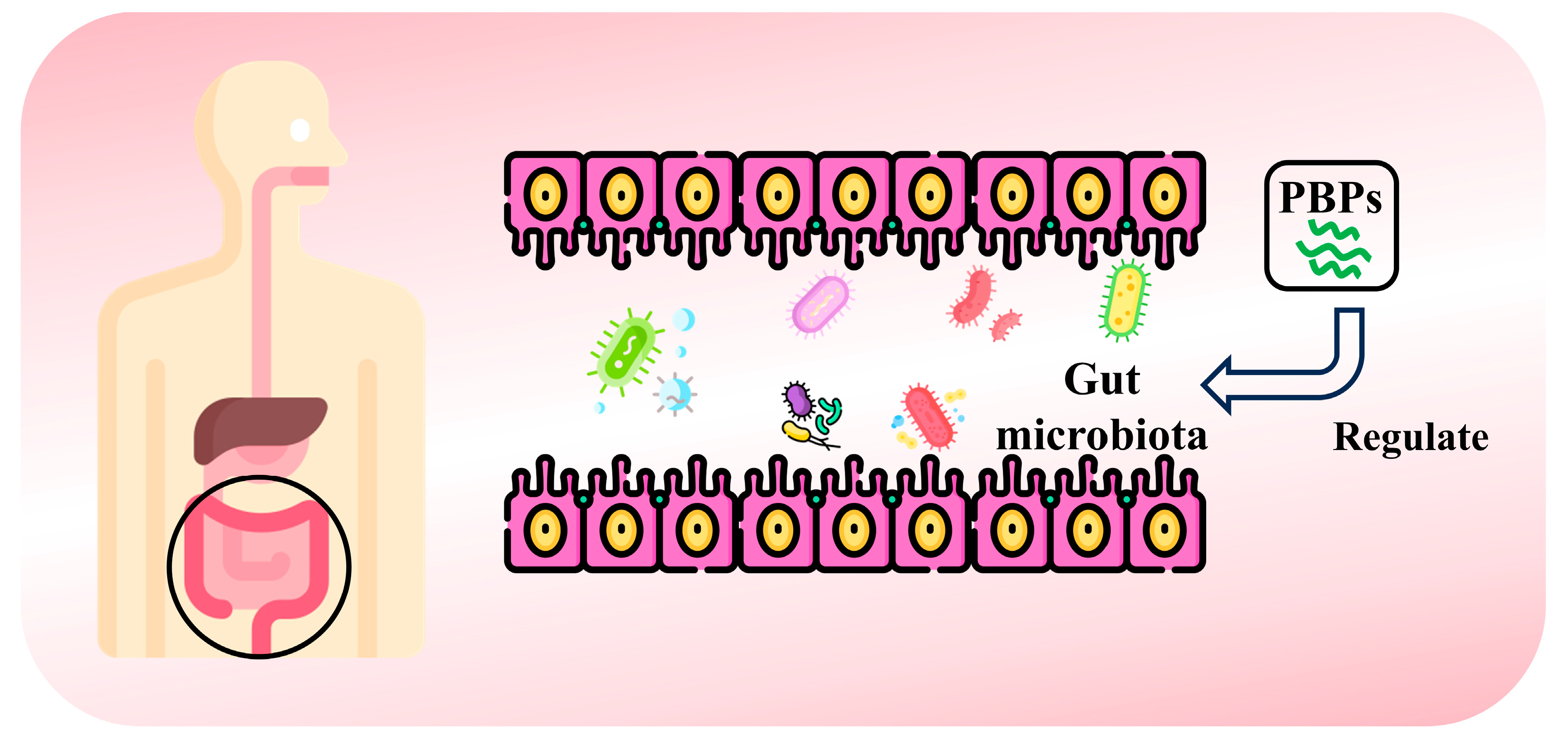
4.7. Regulation of Gut Flora
4.8. Other Biological Activities
5. Structure–Activity Relationship of PDBPs
5.1. Molecular Weight
5.2. Amino Acid Composition and Sequence
5.3. Secondary Structure
6. Application of PDBPs in Chronic Diseases
7. Limitations
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Thakur, S.; Pandey, A.K.; Verma, K.; Shrivastava, A.; Singh, N. Plant-based protein as an alternative to animal proteins: A review of sources, extraction methods and applications. Int. J. Food Sci. Technol. 2024, 59, 488–497. [Google Scholar] [CrossRef]
- Päivärinta, E.; Itkonen, S.T.; Pellinen, T.; Lehtovirta, M.; Erkkola, M.; Pajari, A.-M. Replacing Animal-Based Proteins with Plant-Based Proteins Changes the Composition of a Whole Nordic Diet—A Randomised Clinical Trial in Healthy Finnish Adults. Nutrients 2020, 12, 943. [Google Scholar] [CrossRef] [PubMed]
- Görgüç, A.; Gençdağ, E.; Yılmaz, F.M. Bioactive peptides derived from plant origin by-products: Biological activities and techno-functional utilizations in food developments—A review. Food Res. Int. 2020, 136, 109504. [Google Scholar] [CrossRef] [PubMed]
- Sosalagere, C.; Adesegun Kehinde, B.; Sharma, P. Isolation and functionalities of bioactive peptides from fruits and vegetables: A reviews. Food Chem. 2022, 366, 130494. [Google Scholar] [CrossRef] [PubMed]
- Aderinola, T.A.; Duodu, K.G. Production, health-promoting properties and characterization of bioactive peptides from cereal and legume grains. BioFactors 2022, 48, 972–992. [Google Scholar] [CrossRef]
- Li, X.; Guo, M.; Chi, J.; Ma, J. Bioactive Peptides from Walnut Residue Protein. Molecules 2020, 25, 1285. [Google Scholar] [CrossRef]
- Nourmohammadi, E.; Sadeghi Mahoonak, A. Health Implications of Bioactive Peptides: A Review. Int. J. Vitam. Nutr. Res. 2018, 88, 319–343. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Strappe, P.; Shang, W.T.; Zhou, Z.K. Functional peptides derived from rice bran proteins. Crit. Rev. Food Sci. Nutr. 2019, 59, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, X.; Kim, S.J.; Zhang, W. Antimycin-type depsipeptides: Discovery, biosynthesis, chemical synthesis, and bioactivities. Nat. Prod. Rep. 2016, 33, 1146–1165. [Google Scholar] [CrossRef]
- Cruz-Casas, D.E.; Aguilar, C.N.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Chávez-González, M.L.; Flores-Gallegos, A.C. Enzymatic hydrolysis and microbial fermentation: The most favorable biotechnological methods for the release of bioactive peptides. Food Chem. Mol. Sci. 2021, 3, 100047. [Google Scholar] [CrossRef] [PubMed]
- Karami, Z.; Akbari-adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Nasri, M. Chapter Four—Protein Hydrolysates and Biopeptides: Production, Biological Activities, and Applications in Foods and Health Benefits. A Review. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 81, pp. 109–159. [Google Scholar]
- Sun, X.; Acquah, C.; Aluko, R.E.; Udenigwe, C.C. Considering food matrix and gastrointestinal effects in enhancing bioactive peptide absorption and bioavailability. J. Funct. Foods 2020, 64, 103680. [Google Scholar] [CrossRef]
- Liu, W.; Chen, X.; Li, H.; Zhang, J.; An, J.; Liu, X. Anti-Inflammatory Function of Plant-Derived Bioactive Peptides: A Review. Foods 2022, 11, 2361. [Google Scholar] [CrossRef] [PubMed]
- Kannan, A.; Hettiarachchy, N.S.; Lay, J.O.; Liyanage, R. Human cancer cell proliferation inhibition by a pentapeptide isolated and characterized from rice bran. Peptides 2010, 31, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef]
- Duffuler, P.; Bhullar, K.S.; de Campos Zani, S.C.; Wu, J. Bioactive Peptides: From Basic Research to Clinical Trials and Commercialization. J. Agric. Food Chem. 2022, 70, 3585–3595. [Google Scholar] [CrossRef]
- Lins, M.; Puppin Zandonadi, R.; Raposo, A.; Ginani, V.C. Food Waste on Foodservice: An Overview through the Perspective of Sustainable Dimensions. Foods 2021, 10, 1175. [Google Scholar] [CrossRef]
- Memarpoor-Yazdi, M.; Karbalaei-Heidari, H.R.; Khajeh, K. Production of the renewable extremophile lipase: Valuable biocatalyst with potential usage in food industry. Food Bioprod. Process. 2017, 102, 153–166. [Google Scholar] [CrossRef]
- Courrol, D.d.S.; Silva, C.C.F.d.; Prado, L.G.; Chura-Chambi, R.M.; Morganti, L.; de Souza, G.O.; Heinemann, M.B.; Isaac, L.; Conte, F.P.; Portaro, F.C.V.; et al. Leptolysin, a Leptospira secreted metalloprotease of the pappalysin family with broad-spectrum activity. Front. Cell. Infect. Microbiol. 2022, 12, 966370. [Google Scholar] [CrossRef] [PubMed]
- An, G.; Wei, M.; Wang, Z.; Zhao, X.; Li, X.; He, Z. Effects of enzymolysis method on the preparation of peptides from wheat flour. Food Biosci. 2022, 49, 101956. [Google Scholar] [CrossRef]
- Xiang, H.; Waterhouse, D.-S.; Liu, P.; Waterhouse, G.I.N.; Li, J.; Cui, C. Pancreatic lipase-inhibiting protein hydrolysate and peptides from seabuckthorn seed meal: Preparation optimization and inhibitory mechanism. LWT 2020, 134, 109870. [Google Scholar] [CrossRef]
- Song, W.; Kong, X.; Hua, Y.; Chen, Y.; Zhang, C.; Chen, Y. Identification of antibacterial peptides generated from enzymatic hydrolysis of cottonseed proteins. LWT 2020, 125, 109199. [Google Scholar] [CrossRef]
- Chen, L.; Li, D.; Zhu, C.; Rong, Y.; Zeng, W. Characterisation of antioxidant peptides from enzymatic hydrolysate of golden melon seeds protein. Int. J. Food Sci. Technol. 2021, 56, 5904–5912. [Google Scholar] [CrossRef]
- Tian, S.; Du, K.; Yan, F.; Li, Y. Microwave-assisted enzymatic hydrolysis of wheat germ albumin to prepare polypeptides and influence on physical and chemical properties. Food Chem. 2022, 374, 131707. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qiao, Y.; Shi, B.; Dia, V.P. Alcalase and bromelain hydrolysis affected physicochemical and functional properties and biological activities of legume proteins. Food Struct. 2021, 27, 100178. [Google Scholar] [CrossRef]
- Ren, L.-K.; Fan, J.; Yang, Y.; Liu, X.-F.; Wang, B.; Bian, X.; Wang, D.-F.; Xu, Y.; Liu, B.-X.; Zhu, P.-Y.; et al. Identification, in silico selection, and mechanism study of novel antioxidant peptides derived from the rice bran protein hydrolysates. Food Chem. 2023, 408, 135230. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-F.; Li, Y.-Q.; Wang, C.-Y.; Liang, Y.; Zhao, X.-Z.; He, J.-X.; Mo, H.-Z. Physicochemical, functional and antioxidant properties of mung bean protein enzymatic hydrolysates. Food Chem. 2022, 393, 133397. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vaquero, M.; Mora, L.; Hayes, M. In Vitro and In Silico Approaches to Generating and Identifying Angiotensin-Converting Enzyme I Inhibitory Peptides from Green Macroalga Ulva lactuca. Mar. Drugs 2019, 17, 204. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Peng, M.; Yu, M.; Jiang, B.; Wu, H.; Chen, J. Effect of Enzymatic Hydrolysis on the Zinc Binding Capacity and in vitro Gastrointestinal Stability of Peptides Derived From Pumpkin (Cucurbita pepo L.) Seeds. Front. Nutr. 2021, 8, 647782. [Google Scholar] [CrossRef]
- Shen, Y.; Fang, L.; Liu, C.; Wang, J.; Wu, D.; Zeng, Q.; Leng, Y.; Min, W. Effect of bi-enzyme hydrolysis on the properties and composition of hydrolysates of Manchurian walnut dreg protein. Food Chem. 2024, 447, 138947. [Google Scholar] [CrossRef]
- Ozón, B.; Cotabarren, J.; Valicenti, T.; Graciela Parisi, M.; David Obregón, W. Chia expeller: A promising source of antioxidant, antihypertensive and antithrombotic peptides produced by enzymatic hydrolysis with Alcalase and Flavourzyme. Food Chem. 2022, 380, 132185. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, F.; Chen, H.; Shu, G. Preparation and identification of novel angiotensin-I-converting enzyme inhibitory peptides from Moringa oleifera leaf. LWT 2024, 205, 116472. [Google Scholar] [CrossRef]
- Cao, M.; Li, W.; Li, H.; Zhang, J.; Liu, Y.; Liu, X. Antioxidant and ACE inhibitory activities of peptides prepared from adzuki bean by semi-solid enzymatic hydrolysis. Food Biosci. 2022, 47, 101620. [Google Scholar] [CrossRef]
- Liu, W.; Yu, S.; Han, Y.; Chen, L.; An, J.; Li, H.; Liu, X. Systematic sequence characterization of enzymatic-derived soybean peptides for precision enhancement of anti-inflammatory properties. Food Biosci. 2024, 60, 104292. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Yarmand, M.S.; Ghanbarzadeh, B.; Mirzaee, H. Characterization of bioactive peptides produced from green lentil (Lens culinaris) seed protein concentrate using Alcalase and Flavourzyme in single and sequential hydrolysis. J. Food Process. Preserv. 2021, 45, e15932. [Google Scholar] [CrossRef]
- Goyal, N.; Hajare, S.N.; Gautam, S. Release of an encrypted, highly potent ACE-inhibitory peptide by enzymatic hydrolysis of moth bean (Vigna aconitifolia) protein. Front. Nutr. 2023, 10, 1167259. [Google Scholar] [CrossRef]
- Gao, Y.-L.; Wang, C.-S.; Zhu, Q.-H.; Qian, G.-Y. Optimization of Solid-State Fermentation with Lactobacillus brevis and Aspergillus oryzae for Trypsin Inhibitor Degradation in Soybean Meal. J. Integr. Agric. 2013, 12, 869–876. [Google Scholar] [CrossRef]
- Razavizadeh, S.; Alencikiene, G.; Vaiciulyte-Funk, L.; Ertbjerg, P.; Salaseviciene, A. Utilization of fermented and enzymatically hydrolyzed soy press cake as ingredient for meat analogues. LWT 2022, 165, 113736. [Google Scholar] [CrossRef]
- Babini, E.; Taneyo-Saa, D.L.; Tassoni, A.; Ferri, M.; Kraft, A.; Grän-Heedfeld, J.; Bretz, K.; Roda, A.; Michelini, E.; Calabretta, M.M.; et al. Microbial Fermentation of Industrial Rice-Starch Byproduct as Valuable Source of Peptide Fractions with Health-Related Activity. Microorganisms 2020, 8, 986. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, W.; Xue, J.; Yang, J.; Chen, X.; Shao, Y.; Kwok, L.-y.; Bilige, M.; Mang, L.; Zhang, H. Angiotensin-converting enzyme inhibitory activity of Lactobacillus helveticus strains from traditional fermented dairy foods and antihypertensive effect of fermented milk of strain H9. J. Dairy Sci. 2014, 97, 6680–6692. [Google Scholar] [CrossRef] [PubMed]
- Tonini, S.; Tlais, A.Z.A.; Galli, B.D.; Helal, A.; Tagliazucchi, D.; Filannino, P.; Zannini, E.; Gobbetti, M.; Di Cagno, R. Lentils protein isolate as a fermenting substrate for the production of bioactive peptides by lactic acid bacteria and neglected yeast species. Microb. Biotechnol. 2024, 17, e14387. [Google Scholar] [CrossRef] [PubMed]
- Marulo, S.; De Caro, S.; Nitride, C.; Di Renzo, T.; Di Stasio, L.; Ferranti, P.; Reale, A.; Mamone, G. Bioactive peptides released by lactic acid bacteria fermented pistachio beverages. Food Biosci. 2024, 59, 103988. [Google Scholar] [CrossRef]
- Ma, J.; Su, K.; Chen, M.; Wang, S. Study on the antioxidant activity of peptides from soybean meal by fermentation based on the chemical method and AAPH-induced oxidative stress. Food Sci. Nutr. 2023, 11, 6634–6647. [Google Scholar] [CrossRef]
- Huang, W.; Xu, H.; Pan, J.; Dai, C.; Mintah, B.K.; Dabbour, M.; Zhou, R.; He, R.; Ma, H. Mixed-Strain Fermentation Conditions Screening of Polypeptides from Rapeseed Meal and the Microbial Diversity Analysis by High-Throughput Sequencing. Foods 2022, 11, 3285. [Google Scholar] [CrossRef] [PubMed]
- Chirinos, R.; Pedreschi, R.; Campos, D. Enzyme-assisted hydrolysates from sacha inchi (Plukenetia volubilis) protein with in vitro antioxidant and antihypertensive properties. J. Food Process. Preserv. 2020, 44, e14969. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Q.; Kong, D.; Xu, P. Production and Functionality of Food-derived Bioactive Peptides: A Review. Mini-Rev. Med. Chem. 2018, 18, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Alavi, F.; Ciftci, O.N. Purification and fractionation of bioactive peptides through membrane filtration: A critical and application review. Trends Food Sci. Technol. 2023, 131, 118–128. [Google Scholar] [CrossRef]
- Agyei, D.; Ongkudon, C.M.; Wei, C.Y.; Chan, A.S.; Danquah, M.K. Bioprocess challenges to the isolation and purification of bioactive peptides. Food Bioprod. Process. 2016, 98, 244–256. [Google Scholar] [CrossRef]
- Dou, L.; Zhang, Z.; Yang, W.; Chen, Y.; Luo, K.; Kan, J. Separation and purification of antioxidant peptides from Idesia polycarpa Maxim. cake meal and study of conformational relationship between them. Food Sci. Nutr. 2024, 12, 6206–6225. [Google Scholar] [CrossRef]
- Mijiti, Y.; Wali, A.; Jian, Y.; Rozi, P.; Yili, A.; Aisa, H.A. Isolation Purification and Characterization of Antimicrobial Peptides from Cuminum cyminum L. Seeds. Int. J. Pept. Res. Ther. 2018, 24, 525–533. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.; Li, X. Novel peptides with α-glucosidase inhibitory activity from Changii Radix hydrolysates. Process Biochem. 2021, 111, 200–206. [Google Scholar] [CrossRef]
- Sathya, R.; MubarakAli, D.; Mehboob Nousheen, M.G.; Vasimalai, N.; Thajuddin, N.; Kim, J.-W. An Investigation of Pepsin Hydrolysate of Short Antibacterial Peptides Derived from Limnospira Sp. Appl. Biochem. Biotechnol. 2022, 194, 5580–5593. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, S.; Wang, L.; Huang, D.; Chen, S. Identification and characterization of an angiotensin-I converting enzyme inhibitory peptide from enzymatic hydrolysate of rape (Brassica napus L.) bee pollen. LWT 2021, 147, 111502. [Google Scholar] [CrossRef]
- Yuan, H.; Luo, Z.; Ban, Z.; Reiter, R.J.; Ma, Q.; Liang, Z.; Yang, M.; Li, X.; Li, L. Bioactive peptides of plant origin: Distribution, functionality, and evidence of benefits in food and health. Food Funct. 2022, 13, 3133–3158. [Google Scholar] [CrossRef] [PubMed]
- Tirichen, H.; Yaigoub, H.; Xu, W.; Wu, C.; Li, R.; Li, Y. Mitochondrial Reactive Oxygen Species and Their Contribution in Chronic Kidney Disease Progression Through Oxidative Stress. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, X. Purification, Identification and Evaluation of Antioxidant Peptides from Pea Protein Hydrolysates. Molecules 2023, 28, 2952. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, R.; Wang, J.; Tong, Y.; Zhang, J.; Li, Z.; Zhang, H.; Abbas, Z.; Si, D.; Wei, X. Isolation, Characterization, and Functional Properties of Antioxidant Peptides from Mulberry Leaf Enzymatic Hydrolysates. Antioxidants 2024, 13, 854. [Google Scholar] [CrossRef] [PubMed]
- Liboureau, P.; Pampanin, D.M. Effects of vegetative propagation on protein content and bioactivity of the red seaweed Palmaria palmata. Food Chem. 2024, 455, 139929. [Google Scholar] [CrossRef]
- Dada, S.O.; Ehie, G.C.; Osukoya, O.A.; Anadozie, S.O.; Adewale, O.B.; Kuku, A. In vitro antioxidant and anti-inflammatory properties of Artocarpus altilis (Parkinson) Fosberg (seedless breadfruit) fruit pulp protein hydrolysates. Sci. Rep. 2023, 13, 1493. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Cui, C.; Yan, F.; Li, N.; Sun, X.; Wang, F.; Wang, C. In Vitro Protective Effect of Pea-Derived Peptides (PPs) via the Keap1/Nrf2 Signaling Pathway on Alpha-Gliadin-Sensitizing Peptide Induced Cacao-2 Cells. Mol. Nutr. Food Res. 2024, 68, 2400010. [Google Scholar] [CrossRef]
- Lemus-Conejo, A.; Villanueva-Lazo, A.; Martin, M.E.; Millan, F.; Millan-Linares, M.C. Sacha Inchi (Plukenetia volubilis L.) Protein Hydrolysate as a New Ingredient of Functional Foods. Foods 2024, 13, 2045. [Google Scholar] [CrossRef]
- Wang, L.; Qu, L.; He, B. Preparation, identification and molecular docking of two novel anti-aging peptides from perilla seed. Heliyon 2024, 10, e33604. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sun, H.-N.; Zhang, M.; Mu, T.-H.; Khan, N.M. Production, identification and characterization of antioxidant peptides from potato protein by energy-divergent and gathered ultrasound assisted enzymatic hydrolysis. Food Chem. 2023, 405, 134873. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, Z.; Xue, N.; Ma, Y.; Jiao, J.; Wang, C.; Zhang, K.; Lin, Y.; Li, S.; Guo, Z.; et al. Identification of a novel oligopeptide from defatted walnut meal hydrolysate as a potential neuroprotective agent. Food Funct. 2024, 15, 5566–5578. [Google Scholar] [CrossRef]
- Ma, C.; Wu, X. Cyperus peptide SFRWQ inhibits oxidation and inflammation in RAW264.7 cell model. Int. J. Biol. Macromol. 2024, 267, 131272. [Google Scholar] [CrossRef]
- Igbokwe, C.J.; Feng, Y.; Louis, H.; Benjamin, I.; Quaisie, J.; Duan, Y.; Tuly, J.A.; Cai, M.; Zhang, H. Novel antioxidant peptides identified from coix seed by molecular docking, quantum chemical calculations and invitro study in HepG2 cells. Food Chem. 2024, 440, 138234. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ren, J.; Wu, H.; Zhang, X.; Han, L.; Gu, R. Inhibitory effects and action mechanism of five antioxidant peptides derived from wheat gluten on cells oxidative stress injury. Food Biosci. 2023, 56, 103236. [Google Scholar] [CrossRef]
- Tao, L.; Gu, F.; Liu, Y.; Yang, M.; Wu, X.-Z.; Sheng, J.; Tian, Y. Preparation of antioxidant peptides from Moringa oleifera leaves and their protection against oxidative damage in HepG2 cells. Front. Nutr. 2022, 9, 1062671. [Google Scholar] [CrossRef]
- Fan, H.; Bhullar, K.S.; Wang, Z.; Wu, J. Soybean-Derived Tripeptide Leu–Ser–Trp (LSW) Protects Human Vascular Endothelial Cells from TNFα-Induced Oxidative Stress and Inflammation via Modulating TNFα Receptors and SIRT1. Foods 2022, 11, 3372. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Huang, P.; Liang, Y.; Wang, Q.; Miao, J. The antioxidant peptides from walnut protein hydrolysates and their protective activity against alcoholic injury. Food Funct. 2024, 15, 5315–5328. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Bucataru, C.; Ciobanasu, C. Antimicrobial peptides: Opportunities and challenges in overcoming resistance. Microbiol. Res. 2024, 286, 127822. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- Karami, Z.; Muangnapoh, C.; Suthienkul, O.; Suriyarak, S.; Duangmal, K. Exploring novel peptides in adzuki bean and mung bean hydrolysates with potent antibacterial activity. Int. J. Food Sci. Technol. 2024, 59, 4829–4840. [Google Scholar] [CrossRef]
- Kong, X.; Song, W.; Hua, Y.; Li, X.; Chen, Y.; Zhang, C.; Chen, Y. Insights into the antibacterial activity of cottonseed protein-derived peptide against Escherichia coli. Food Funct. 2020, 11, 10047–10057. [Google Scholar] [CrossRef]
- Hu, Y.; Ling, Y.; Qin, Z.; Huang, J.; Jian, L.; Ren, D.F. Isolation, identification, and synergistic mechanism of a novel antimicrobial peptide and phenolic compound from fermented walnut meal and their application in Rosa roxbughii Tratt spoilage fungus. Food Chem. 2024, 433, 137333. [Google Scholar] [CrossRef]
- Garzón, A.G.; Veras, F.F.; Brandelli, A.; Drago, S.R. Purification, identification and in silico studies of antioxidant, antidiabetogenic and antibacterial peptides obtained from sorghum spent grain hydrolysate. LWT 2022, 153, 112414. [Google Scholar] [CrossRef]
- Zhao, Q.; He, L.; Wang, X.; Ding, X.; Li, L.; Tian, Y.; Huang, A. Characterization of a Novel Antimicrobial Peptide Isolated from Moringa oleifera Seed Protein Hydrolysates and Its Membrane Damaging Effects on Staphylococcus aureus. J. Agric. Food Chem. 2022, 70, 6123–6133. [Google Scholar] [CrossRef]
- Sollazzo, F.; Di Nitto, M.; Rosito, L.; Torino, F.; Alvaro, R.; Lacarbonara, F.; Vellone, E.; Durante, A. Caregivers’ contribution to self-care for patients treated with oral anticancer agents: A qualitative descriptive study. Eur. J. Oncol. Nurs. 2023, 64, 102327. [Google Scholar] [CrossRef]
- Norouzi, P.; Mirmohammadi, M.; Houshdar Tehrani, M.H. Anticancer peptides mechanisms, simple and complex. Chem. Biol. Interact. 2022, 368, 110194. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Y.; Xia, S.; Ding, X. Finding and isolation of novel peptides with anti-proliferation ability of hepatocellular carcinoma cells from mung bean protein hydrolysates. J. Funct. Foods 2019, 62, 103557. [Google Scholar] [CrossRef]
- Chen, Z.; Li, W.; Santhanam, R.K.; Wang, C.; Gao, X.; Chen, Y.; Wang, C.; Xu, L.; Chen, H. Bioactive peptide with antioxidant and anticancer activities from black soybean [Glycine max (L.) Merr.] byproduct: Isolation, identification and molecular docking study. Eur. Food Res. Technol. 2019, 245, 677–689. [Google Scholar] [CrossRef]
- Taghizadeh, S.F.; Azizi, M.; Asili, J.; Madarshahi, F.S.; Rakhshandeh, H.; Fujii, Y. Therapeutic peptides of Mucuna pruriens L.: Anti-genotoxic molecules against human hepatocellular carcinoma and hepatitis C virus. Food Sci. Nutr. 2021, 9, 2908–2914. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Bhagyawant, S.S. Bioactive peptide of Cicer arietinum L. induces apoptosis in human endometrial cancer via DNA fragmentation and cell cycle arrest. 3 Biotech 2021, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Visuddho, V.; Halim, P.; Helen, H.; Muhar, A.M.; Iqhrammullah, M.; Mayulu, N.; Surya, R.; Tjandrawinata, R.R.; Ribeiro, R.I.; Tallei, T.E.; et al. Modulation of Apoptotic, Cell Cycle, DNA Repair, and Senescence Pathways by Marine Algae Peptides in Cancer Therapy. Mar. Drugs 2024, 22, 338. [Google Scholar] [CrossRef]
- Wu, B.; Cheng, H.; Li, X.; Yang, Q.; Hao, S.; Wang, C.; Sun, B. Identification and functional analysis of phycocyanin-derived bioactive peptides with non-small cell lung cancer cell inhibition. Algal Res. 2024, 79, 103467. [Google Scholar] [CrossRef]
- Du, X.; Xiao, S.; Luo, Q.; Liu, X.; Liu, J. Laminaria japonica cyclic peptides exert anti-colorectal carcinoma effects through apoptosis induction in vitro and in vivo. J. Pept. Sci. 2022, 28, e3385. [Google Scholar] [CrossRef]
- Rasaratnam, K.; Nantasenamat, C.; Phaonakrop, N.; Roytrakul, S.; Tanyong, D. A novel peptide isolated from garlic shows anticancer effect against leukemic cell lines via interaction with Bcl-2 family proteins. Chem. Biol. Drug Des. 2021, 97, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Wattayagorn, V.; Kongsema, M.; Tadakittisarn, S.; Chumnanpuen, P. Riceberry rice bran protein hydrolyzed fractions induced apoptosis, senescence and G1/S cell cycle arrest in human colon cancer cell lines. Appl. Sci. 2022, 12, 6917. [Google Scholar] [CrossRef]
- Rout, A.; Duhan, S.; Umer, M.; Li, M.; Kalra, D. Atherosclerotic cardiovascular disease risk prediction: Current state-of-the-art. Heart 2024, 110, 1005. [Google Scholar] [CrossRef] [PubMed]
- Bregonzio, C. Angiotensin-converting enzyme inhibitors stimulate cerebral arteriogenesis. Acta Physiol. 2022, 234, e13765. [Google Scholar] [CrossRef] [PubMed]
- Windarto, S.; Lee, M.-C.; Nursyam, H.; Hsu, J.-L. First Report of Screening of Novel Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from the Red Alga Acrochaetium sp. Mar. Biotechnol. 2022, 24, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Dong, Y.; Zhang, M.; Li, Z.; Bu, G.; Chen, F. Identification and molecular interactions of novel ACE inhibitory peptides from rapeseed protein. Food Chem. 2023, 422, 136085. [Google Scholar] [CrossRef]
- Zou, Z.; Wang, M.; Wang, Z.; Aluko, R.E.; He, R. Antihypertensive and antioxidant activities of enzymatic wheat bran protein hydrolysates. J. Food Biochem. 2020, 44, e13090. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, C.; Xiao, S.; Wang, Q.; Zhou, A. Two Novel Angiotensin-Converting Enzyme (ACE) Inhibitory and ACE2 Upregulating Peptides from the Hydrolysate of Pumpkin (Cucurbita moschata) Seed Meal. J. Agric. Food Chem. 2024, 72, 10909–10922. [Google Scholar] [CrossRef]
- Qoms, M.S.; Arulrajah, B.; Ibadullah, W.Z.W.; Ramli, N.S.; Shamsudin, R.; Chau, D.-M.; Saari, N. Antihypertensive, Antidiabetic, and Antioxidant Properties of Novel Azolla pinnata Fern Protein Hydrolysates: Inhibition Mechanism, Stability, Profiling, and Molecular Docking. Food Bioprocess Technol. 2024. [Google Scholar] [CrossRef]
- Bernier, M.-È.; Thibodeau, J.; Bazinet, L. Enzymatic Hydrolysis of Water Lentil (Duckweed): An Emerging Source of Proteins for the Production of Antihypertensive Fractions. Foods 2024, 13, 323. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, D.; Shao, L.; Zheng, Y.; Hao, W.; Kan, Y.; Cao, J.; Yu, H.; Liu, J. Oil palm kernel globulin antihypertensive peptides: Isolation and characterization, ACE inhibition mechanisms, zinc-chelating activity, security and stability. Front. Pharmacol. 2023, 14, 1225256. [Google Scholar] [CrossRef] [PubMed]
- Prangthip, P.; Panbangred, W.; Reamtong, O. Potential antihypertensive activity of novel peptides from green basil leaves. BMC Complement. Med. Ther. 2023, 23, 282. [Google Scholar] [CrossRef]
- Fan, X.; Ma, X.; Maimaitiyiming, R.; Aihaiti, A.; Yang, J.; Li, X.; Wang, X.; Pang, G.; Liu, X.; Qiu, C.; et al. Study on the preparation process of quinoa anti-hypertensive peptide and its stability. Front. Nutr. 2023, 9, 1119042. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Peng, D.; Yao, Y.; Huang, K.; Wang, J.; Ma, Z.; Fu, J.; Xu, Y. Optogenetic therapeutic strategies for diabetes mellitus. J. Diabetes 2024, 16, e13557. [Google Scholar] [CrossRef] [PubMed]
- Chan-Zapata, I.; Sandoval-Castro, C.; Segura-Campos, M.R. Proteins and peptides from vegetable food sources as therapeutic adjuvants for the type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2022, 62, 2673–2682. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wu, S.; Jia, C.; Cui, C.; Sun-Waterhouse, D. Active peptides with hypoglycemic effect obtained from hemp (Cannabis sativa L) protein through identification, molecular docking, and virtual screening. Food Chem. 2023, 429, 136912. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Li, P.; Xiao, Z.-W.; Lou, W.-Y. Preparation and identification of hypoglycemic bioactive peptide from Amygdalus communis L. by multienzyme hydrolysis. Process Biochem. 2024, 136, 292–300. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, F.; He, Z.; Fang, X.; Liu, X. Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods 2023, 12, 393. [Google Scholar] [CrossRef]
- Xu, F.; Mejia, E.G.d.; Chen, H.; Rebecca, K.; Pan, M.; He, R.; Yao, Y.; Wang, L.; Ju, X. Assessment of the DPP-IV inhibitory activity of a novel octapeptide derived from rapeseed using Caco-2 cell monolayers and molecular docking analysis. J. Food Biochem. 2020, 44, e13406. [Google Scholar] [CrossRef]
- Fidor, A.; Konkel, R.; Mazur-Marzec, H. Bioactive Peptides Produced by Cyanobacteria of the Genus Nostoc: A Review. Mar. Drugs 2019, 17, 561. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, Y.; Ma, C.-m.; Bian, X.; Liu, X.-f.; Wang, Y.; Chen, F.-l.; Wang, B.; Zhang, G.; Zhang, N. Characterization of the structure, antioxidant activity and hypoglycemic activity of soy (Glycine max L.) protein hydrolysates. Food Res. Int. 2023, 173, 113473. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha Vijayan, P.; Xavier, J.; Valappil, M.P. A review of immune modulators and immunotherapy in infectious diseases. Mol. Cell. Biochem. 2024, 479, 1937–1955. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Jiang, Y.; Zhou, X.; Bi, H.; Yang, B. Structure identification of soybean peptides and their immunomodulatory activity. Food Chem. 2021, 359, 129970. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Lu, X.; Zheng, Y.; Song, H.; Zheng, B. Immunomodulatory effects of lotus seed (Nelumbo nucifera Gaertn.) peptides on macrophages in mice. Food Biosci. 2024, 57, 103494. [Google Scholar] [CrossRef]
- Xu, Z.; Mao, T.-M.; Huang, L.; Yu, Z.-C.; Yin, B.; Chen, M.-L.; Cheng, Y.-H. Purification and identification immunomodulatory peptide from rice protein hydrolysates. Food Agric. Immunol. 2019, 30, 150–162. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Villanueva-Lazo, A.; Millan, F.; Martin-Santiago, V.; Rivero-Pino, F.; Millan-Linares, M.C. Production and identification of immunomodulatory peptides in intestine cells obtained from hemp industrial by-products. Food Res. Int. 2023, 174, 113616. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Pan, L.; Wu, H.; Zhang, L.; Zhang, Y.; Zhang, Y.; Yang, J.; Lin, Z.; Zhang, M. Isolation, Identification, and Immunomodulatory Mechanism of Peptides from Lepidium meyenii (Maca) Protein Hydrolysate. J. Agric. Food Chem. 2022, 70, 4328–4341. [Google Scholar] [CrossRef]
- Liu, P.; Liao, W.; Qi, X.; Yu, W.; Wu, J. Identification of immunomodulatory peptides from zein hydrolysates. Eur. Food Res. Technol. 2020, 246, 931–937. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Liu, Y.; Bi, H.; Zhou, X.; Wen, L.; Yang, B. Characterisation of functional pea protein hydrolysates and their immunomodulatory activity. Int. J. Food Sci. Technol. 2024, 59, 3317–3330. [Google Scholar] [CrossRef]
- Segrist, E.; Cherry, S. Using Diverse Model Systems to Define Intestinal Epithelial Defenses to Enteric Viral Infections. Cell Host Microbe 2020, 27, 329–344. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, D.; Cheng, H.; Wu, J.; Liu, J.; Feng, W.; Peng, C. Repairing gut barrier by traditional Chinese medicine: Roles of gut microbiota. Front. Cell. Infect. Microbiol. 2024, 14, 1389925. [Google Scholar] [CrossRef]
- Wijesekara, T.; Abeyrathne, E.D.; Ahn, D.U. Effect of Bioactive Peptides on Gut Microbiota and Their Relations to Human Health. Foods 2024, 13, 1853. [Google Scholar] [CrossRef] [PubMed]
- Zhi, T.; Hong, D.; Zhang, Z.; Li, S.; Xia, J.; Wang, C.; Wu, Y.; Jia, Y.; Ma, A. Anti-inflammatory and gut microbiota regulatory effects of walnut protein derived peptide LPF in vivo. Food Res. Int. 2022, 152, 110875. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, S.; Zhang, T.; Lv, B.; Jin, Y.; Wang, Y.; Chen, X.; Li, N.; Han, N.; Wu, Y.; et al. Walnut peptide alleviates obesity, inflammation and dyslipidemia in mice fed a high-fat diet by modulating the intestinal flora and metabolites. Front. Immunol. 2023, 14, 1305656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jia, L.; He, H.; Yin, H.; Ming, J.; Hou, T.; Xiang, J. Modulation of oxidative stress and gut microbiota by selenium-containing peptides from Cardamine enshiensis and structural-based characterization. Food Chem. 2022, 395, 133547. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, J.; Liang, F.; Shi, Y.; Zhang, R. Changes in biological activity and gut microbiota of digestion of rice glutelin during storage. J. Cereal Sci. 2022, 104, 103421. [Google Scholar] [CrossRef]
- Han, C.; Kong, X.; Xia, X.; Huang, X.; Mao, Z.; Han, J.; Shi, F.; Liang, Y.; Wang, A.; Zhang, F. Effects of ginseng peptides on the hypoglycemic activity and gut microbiota of a type 2 diabetes mellitus mice model. J. Funct. Foods 2023, 111, 105897. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Zhang, Y.; Zhang, C.; Zhang, J.; Liu, X. Differences in the gut microbiota composition of rats fed with soybean protein and their derived peptides. J. Food Sci. 2021, 86, 5452–5465. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Wang, P.; Liu, C.; Wang, J.; Liu, X.; Liu, J.; Min, W. Hazelnut protein-derived peptide LDAPGHR shows anti-inflammatory activity on LPS-induced RAW264.7 macrophage. J. Funct. Foods 2018, 46, 449–455. [Google Scholar] [CrossRef]
- Feng, T.; Huang, Y.; Tang, Z.; Wei, D.; Mo, J. Anti-fatigue effects of pea (Pisum sativum L.) peptides prepared by compound protease. J. Food Sci. Technol. 2021, 58, 2265–2272. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, L.; Zhao, T.; Zhang, Q.; Liu, Y.; Sun, B.; Su, G.; Zhao, M. Inhibitory Effects of Walnut (Juglans regia) Peptides on Neuroinflammation and Oxidative Stress in Lipopolysaccharide-Induced Cognitive Impairment Mice. J. Agric. Food Chem. 2020, 68, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yang, G.; Li, Y.; Tang, Z.; Du, J.; Song, H.; Xiong, L.; Wang, L.; Weng, Z.; Shen, X. A peptide from wheat germ abolishes the senile osteoporosis by regulating OPG/RANKL/RANK/TRAF6 signaling pathway. Phytomedicine 2022, 104, 154304. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Bao, X.; Liu, X.; Li, X. Flaxseed-derived peptides ameliorate hepatic cholesterol metabolism in Sprague–Dawley rats fed a high-cholesterol and high-fat diet. J. Sci. Food Agric. 2022, 102, 5348–5357. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fang, L.; Wang, X.; Wu, D.; Liu, C.; Liu, X.; Wang, J.; Gao, Y.; Min, W. Structure-Activity Relationship of Pine Nut-Derived Peptides and Their Protective Effect on Nerve-Cell Mitochondria. Foods 2022, 11, 1428. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Study on the structure–activity relationship of watermelon seed antioxidant peptides by using molecular simulations. Food Chem. 2021, 364, 130432. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Li, J.; Xu, J.; Fang, L.; Liu, C.; Wu, D.; Min, W. Structure-activity relationship of walnut peptide in gastrointestinal digestion, absorption and antioxidant activity. LWT 2023, 189, 115521. [Google Scholar] [CrossRef]
- Zhou, Y.; She, X.; Chen, Z.; Wei, Y.; Xiao, Y.; Zhou, X. Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn) protein-derived antioxidant peptides: Mechanisms of action and structure-activity relationship in Caco-2 cell models. Food Sci. Hum. Wellness 2022, 11, 1580–1590. [Google Scholar] [CrossRef]
- Zhu, F.; He, S.; Ni, C.; Wu, Y.; Wu, H.; Wen, L. Study on the structure–activity relationship of rice immunopeptides based on molecular docking. Food Chem. X 2024, 21, 101158. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.-S.; Lu, M.-S.; Chiang, W.-D. Identification and characterization of immunomodulatory peptides from pepsin–soy protein hydrolysates. Bioresour. Bioprocess. 2022, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Velliquette, R.A.; Fast, D.J.; Maly, E.R.; Alashi, A.M.; Aluko, R.E. Enzymatically derived sunflower protein hydrolysate and peptides inhibit NFκB and promote monocyte differentiation to a dendritic cell phenotype. Food Chem. 2020, 319, 126563. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-A.; Kim, Y.-J.; Jin, S.-K.; Choi, H.-J. Antioxidant, Collagenase Inhibitory, and Antibacterial Effects of Bioactive Peptides Derived from Enzymatic Hydrolysate of Ulva australis. Mar. Drugs 2023, 21, 469. [Google Scholar] [CrossRef]
- León Madrazo, A.; Fuentes Ortíz, A.B.; Morales Mendoza, L.F.; Segura Campos, M.R. Antibacterial peptide fractions from chia seeds (Salvia hispanica L.) and their stability to food processing conditions. J. Food Sci. Technol. 2022, 59, 4332–4340. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Hong, Z.; Dai, J.; Li, T.; Bai, Y.; Zhang, L.; Hu, X.; Chen, J.; Sheng, J.; Tian, Y. Isolation and identification of anti-colorectal cancer peptides from walnut proteins and associated in silico analysis. J. Funct. Foods 2024, 112, 105952. [Google Scholar] [CrossRef]
- Taniya, M.; Reshma, M.; Shanimol, P.; Krishnan, G.; Priya, S. Bioactive peptides from amaranth seed protein hydrolysates induced apoptosis and antimigratory effects in breast cancer cells. Food Biosci. 2020, 35, 100588. [Google Scholar] [CrossRef]
- Hu, R.; Xu, J.; Qi, G.; Wang, W.; Sun, X.S.; Li, Y. Antioxidative hydrolysates from corn gluten meal may effectively reduce lipid oxidation and inhibit HepG2 cancer cell growth. J. Agric. Food Res. 2022, 7, 100252. [Google Scholar] [CrossRef]
- Phyo, S.H.; Ghamry, M.; Bao, G.; Zeng, A.; Zhao, W. Potential inhibitory effect of highland barley protein hydrolysates on the formation of advanced glycation end-products (AGEs): A mechanism study. Int. J. Biol. Macromol. 2024, 268, 131632. [Google Scholar] [CrossRef]
- Cai, L.; Wu, S.; Jia, C.; Cui, C. Hydrolysates of hemp (Cannabis sativa L.) seed meal: Characterization and their inhibitory effect on α-glucosidase activity and glucose transport in Caco-2 cells. Ind. Crops Prod. 2023, 205, 117559. [Google Scholar] [CrossRef]
- Chen, H.-J.; Dai, F.-J.; Chen, C.-Y.; Fan, S.-L.; Zheng, J.-H.; Chau, C.-F.; Lin, Y.-S.; Chen, C.-S. Effects of molecular weight fraction on antioxidation capacity of rice protein hydrolysates. Sci. Rep. 2023, 13, 3464. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, Y.; Qi, Z.; Chen, Q.; Cao, Y.; Kong, Q. Preparation and identification of a novel peptide with high antioxidant activity from corn gluten meal. Food Chem. 2023, 424, 136389. [Google Scholar] [CrossRef]
- López-Huertas, E.; Rubí-Villegas, J.; Sánchez-Moreno, L.; Nieto, R. Olive Pomace Extract Contains Low Molecular Weight Peptides and Possesses ACE Inhibitory Activity. Int. J. Mol. Sci. 2024, 25, 3962. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Lu, J.; Zhang, Y.; Wang, J.; Wang, S.; Fan, H.; Zhang, J.; Dai, W.; Gao, J.; Yu, H. Structural properties, antioxidant and immune activities of low molecular weight peptides from soybean dregs (Okara). Food Chem. X 2021, 12, 100175. [Google Scholar] [CrossRef]
- Dhaouafi, J.; Romdhani, M.; Deracinois, B.; Flahaut, C.; Nedjar, N.; Balti, R. Fractionation and identification of bioactive peptides from red macroalgae protein hydrolysates: In silico analysis and in vitro bioactivities. Biocatal. Agric. Biotechnol. 2024, 58, 103211. [Google Scholar] [CrossRef]
- Wang, H.; Chi, X.; Zhang, D. Potential Regulatory Gene Network Associated with the Ameliorative Effect of Oat Antibacterial Peptides on Rat Colitis. Foods 2024, 13, 236. [Google Scholar] [CrossRef]
- César, A.P.C.; Lopes, F.E.S.; Azevedo, F.F.N.; Pinto, Y.O.; Andrade, C.R.; Mesquita, F.P.; Silva, G.O.; Freitas, C.D.T.; Souza, P.F.N. Antioxidant peptides from plants: A review. Phytochem. Rev. 2024, 23, 95–104. [Google Scholar] [CrossRef]
- Abdelhameed, S.A.M.; Vandebroek, L.; de Azambuja, F.; Parac-Vogt, T.N. Redox Activity of Ce(IV)-Substituted Polyoxometalates toward Amino Acids and Peptides. Inorg. Chem. 2020, 59, 10569–10577. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Zhong, Y.; Liu, H. Characterization and relationship analysis of antioxidant and anti-inflammatory peptides in pomelo fruitlet albumin. Food Chem. 2024, 446, 138798. [Google Scholar] [CrossRef]
- Wang, X.; Fu, J.; Bhullar, K.S.; Chen, B.; Liu, H.; Zhang, Y.; Wang, C.; Liu, C.; Su, D.; Ma, X.; et al. Identification, in silico selection, and mechanistic investigation of antioxidant peptides from corn gluten meal hydrolysate. Food Chem. 2024, 446, 138777. [Google Scholar] [CrossRef]
- Wang, L.; Ma, M.; Yu, Z.; Du, S.-k. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef]
- Guo, H.; Hao, Y.; Richel, A.; Everaert, N.; Chen, Y.; Liu, M.; Yang, X.; Ren, G. Antihypertensive effect of quinoa protein under simulated gastrointestinal digestion and peptide characterization. J. Sci. Food Agric. 2020, 100, 5569–5576. [Google Scholar] [CrossRef]
- Roy, S.; Agarwal, T.; Das, A.; Halder, T.; Upadhyaya, G.; Chaubey, B.; Ray, S. The C-terminal stretch of glycine-rich proline-rich protein (SbGPRP1) from Sorghum bicolor serves as an antimicrobial peptide by targeting the bacterial outer membrane protein. Plant Mol. Biol. 2023, 111, 131–151. [Google Scholar] [CrossRef] [PubMed]
- LiWang, A.; Porter, L.L.; Wang, L.-P. Fold-switching proteins. Biopolymers 2021, 112, e23478. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Z.-m.; Feng, Y.; Yu, S.; Li, Z.; Zhang, D.; Wang, C. DPP-IV Inhibitory Peptides from Coix Seed Prolamins: Release, Identification, and Analysis of the Interaction between Key Residues and Enzyme Domains. J. Agric. Food Chem. 2023, 71, 14575–14592. [Google Scholar] [CrossRef] [PubMed]
- Habinshuti, I.; Mu, T.-H.; Zhang, M. Ultrasound microwave-assisted enzymatic production and characterisation of antioxidant peptides from sweet potato protein. Ultrason. Sonochem. 2020, 69, 105262. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Meng, F.; Du, K. Physicochemical properties and structure characteristics of different molecular weight peptides from ultrasonic assisted papain hydrolysate of wheat germ albumin. Ind. Crops Prod. 2024, 211, 118254. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhou, J.; Duan, Y.; Zhang, H.; Ma, H. Effects of slit divergent ultrasound and enzymatic treatment on the structure and antioxidant activity of arrowhead protein. Ultrason. Sonochem. 2018, 49, 294–302. [Google Scholar] [CrossRef]
- Zhang, X.; Du, H.; Xu, Z.; Wang, Y.; Guo, X.; Xiao, H.; Li, Y. A novel alcalase-hydrolyzed soybean meal hydrolysates prepared using by-product material: Structure, function property, sensory property, and biological activity. Food Biosci. 2023, 56, 103324. [Google Scholar] [CrossRef]
- Sun, C.; Shan, Y.; Tang, X.; Han, D.; Wu, X.; Wu, H.; Hosseininezhad, M. Effects of enzymatic hydrolysis on physicochemical property and antioxidant activity of mulberry (Morus atropurpurea Roxb.) leaf protein. Food Sci. Nutr. 2021, 9, 5379–5390. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, D.; Guo, C.; Wang, J.; Xue, S. Construction and Reliability and Validity Test of Home Care Assessment Scale for Elderly Patients with Chronic Diseases Based on Intelligent Medical Care. Mob. Inf. Syst. 2022, 2022, 7697036. [Google Scholar] [CrossRef]
- Formanowicz, D. Pathomechanisms of Disturbances Underlying Chronic Disorders. Biomedicines 2024, 12, 131. [Google Scholar] [CrossRef]
- Khan, S.S.; Greenland, P. Comprehensive Cardiovascular Health Promotion for Successful Prevention of Cardiovascular Disease. JAMA 2020, 324, 2036–2037. [Google Scholar] [CrossRef]
- Pappachan, J.M.; Fernandez, C.J.; Chacko, E.C. Diabesity and antidiabetic drugs. Mol. Asp. Med. 2019, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yang, L.; Chen, G.; Xu, F.; Yang, F.; Yu, H.; Li, L.; Dong, X.; Han, J.; Cao, C.; et al. A Review on Drug Delivery System for Tumor Therapy. Front. Pharmacol. 2021, 12, 735446. [Google Scholar] [CrossRef] [PubMed]
- Luvián-Morales, J.; Varela-Castillo, F.O.; Flores-Cisneros, L.; Cetina-Pérez, L.; Castro-Eguiluz, D. Functional foods modulating inflammation and metabolism in chronic diseases: A systematic review. Crit. Rev. Food Sci. Nutr. 2022, 62, 4371–4392. [Google Scholar] [CrossRef] [PubMed]
- Amato, A. Natural Compounds and Healthy Foods: Useful Tools against Onset and Progression of Chronic Diseases. Nutrients 2023, 15, 2898. [Google Scholar] [CrossRef]
- Mahgoub, S.; Alagawany, M.; Nader, M.; Omar, S.M.; Abd El-Hack, M.E.; Swelum, A.; Elnesr, S.S.; Khafaga, A.F.; Taha, A.E.; Farag, M.R.; et al. Recent Development in Bioactive Peptides from Plant and Animal Products and Their Impact on the Human Health. Food Rev. Int. 2023, 39, 511–536. [Google Scholar] [CrossRef]
- Chai, T.-T.; Ee, K.-Y.; Kumar Thirumal, D.; Manan Abd, F.; Wong, F.-C. Plant Bioactive Peptides: Current Status and Prospects Towards Use on Human Health. Protein Pept. Lett. 2021, 28, 623–642. [Google Scholar] [CrossRef]
- Stuermer, E.K.; Besser, M.; Terberger, N.; Bachmann, H.S.; Severing, A.-L. Side Effects of Frequently Used Antihypertensive Drugs on Wound Healing in vitro. Ski. Pharmacol. Physiol. 2019, 32, 162–172. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Ruan, S.; Lu, F.; Tian, W.; Ma, H. Antihypertensive effect of rapeseed peptides and their potential in improving the effectiveness of captopril. J. Sci. Food Agric. 2021, 101, 3049–3055. [Google Scholar] [CrossRef]
- Xingfei, L.; Shunshun, P.; Wenji, Z.; Lingli, S.; Qiuhua, L.; Ruohong, C.; Shili, S. Properties of ACE inhibitory peptide prepared from protein in green tea residue and evaluation of its anti-hypertensive activity. Process Biochem. 2020, 92, 277–287. [Google Scholar] [CrossRef]
- Shu, Y.; Cao, X.-Y.; Chen, J. Preparation and antagonistic effect of ACE inhibitory peptide from cashew. J. Sci. Food Agric. 2019, 99, 6822–6832. [Google Scholar] [CrossRef] [PubMed]
- Blahova, J.; Martiniakova, M.; Babikova, M.; Kovacova, V.; Mondockova, V.; Omelka, R. Pharmaceutical Drugs and Natural Therapeutic Products for the Treatment of Type 2 Diabetes Mellitus. Pharmaceuticals 2021, 14, 806. [Google Scholar] [CrossRef]
- Deng, F.; Liang, Y.; Lei, Y.; Xiong, S.; Rong, J.; Hu, Y. Development and Identification of Novel α-Glucosidase Inhibitory Peptides from Mulberry Leaves. Foods 2023, 12, 3917. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, H.; Wen, Y.; Liu, Y.; Wang, J.; Sun, B. Molecular Mechanism for the α-Glucosidase Inhibitory Effect of Wheat Germ Peptides. J. Agric. Food Chem. 2021, 69, 15231–15239. [Google Scholar] [CrossRef]
- Wang, X.; Fan, Y.; Xu, F.; Xie, J.; Gao, X.; Li, L.; Tian, Y.; Sheng, J. Characterization of the structure, stability, and activity of hypoglycemic peptides from Moringa oleifera seed protein hydrolysates. Food Funct. 2022, 13, 3481–3494. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Rios, E.; Del-Toro-Sanchez Lizette, C. Antioxidant Peptides from Terrestrial and Aquatic Plants Against Cancer. Curr. Protein Pept. Sci. 2018, 19, 368–379. [Google Scholar] [CrossRef]
- Chiangjong, W.; Chutipongtanate, S.; Hongeng, S. Anticancer peptide: Physicochemical property, functional aspect and trend in clinical application (Review). Int. J. Oncol. 2020, 57, 678–696. [Google Scholar] [CrossRef]
- Ma, K.; Wang, Z.; Ju, X.; Huang, J.; He, R. Rapeseed peptide inhibits HepG2 cell proliferation by regulating the mitochondrial and P53 signaling pathways. J. Sci. Food Agric. 2023, 103, 1474–1483. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.-T.; Jung, S.-H.; Han, J.W.; Jo, S.; Kim, I.-G.; Kim, R.-K.; Kahm, Y.-J.; Choi, T.-I.; Kim, C.-H.; et al. A Novel Anticancer Peptide Derived from Bryopsis plumosa Regulates Proliferation and Invasion in Non-Small Cell Lung Cancer Cells. Mar. Drugs 2023, 21, 607. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vital, D.; González de Mejía, E. Peptides from legumes with antigastrointestinal cancer potential: Current evidence for their molecular mechanisms. Curr. Opin. Food Sci. 2018, 20, 13–18. [Google Scholar] [CrossRef]
- Taghizadeh, M.S.; Niazi, A.; Moghadam, A.; Afsharifar, A.R. Novel bioactive peptides of Achillea eriophora show anticancer and antioxidant activities. Bioorg. Chem. 2021, 110, 104777. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation Method | Advantages | Disadvantages |
|---|---|---|
| Enzymatic hydrolysis | No toxic substances are produced Non-destructive amino acid High catalytic efficiency High specificity | High cost |
| Microbial fermentation | High efficiency Low cost | Difficult to control Susceptible to microbial contamination Difficult separation and purification |
| Plant Origins | Sequence/Name | Major Findings | Reference |
|---|---|---|---|
| Pea Protein | YLVN EEHLCFR TFY3 | Through computer simulation of molecular docking, it was found that the anti-oxidation mechanism may be highly related to the activation of the Keap1-Nrf2 pathway by occupying the Keap1-Nrf2 binding site. | [58] |
| Mulberry leaves | EGDAGAQGPPGPAGPAGER RPGPSPGVGAPGK | ↓ ROS ↑ SOD and CAT Both peptides are found to exert protective effects against H2O2-induced chromatin damage and cell apoptosis. | [59] |
| Red seaweed (Palmaria palmata) | SLLYSDITRPGGNMYTTR (SR18) | The SR18 peptide was found to have extremely high ROS scavenging activity and high ferric reducing ability and may be partially driven by the amino acids Tyr, Asn, and Met. | [60] |
| Artocarpus altilis (Parkinson) Fosberg | AAPPH | The different enzymatic hydrolysis products of the hydrolysis had a higher Fe2+-chelating capacity, DPPH scavenging capacity, and hydrogen peroxide scavenging capacity. | [61] |
| Rice bran | AFDEGPWPK | AFDEGPWPK has the ability to scavenge ORAC and DPPH free radicals, and it can enter the binding pocket of the Kelch structural domain and activate the Keap1/Nrf2/HO-1 pathway. | [27] |
| Pea | Pea-derived peptides (PPs) | ↓ SOD, GR, GSH, GSSG, and ROS | [62] |
| Sacha inchi (Plukenetia volubilis L.) | SIH20B AAGALKKFL LGVKFKGGL | Antioxidant capacity (DPPH free radical chelating capacity, ferric ion reducing antioxidant capacity, and β-carotene-linoleic acid assay) | [63] |
| Perilla seed meal | NFF and PMR perilla seed peptides (PSP) | ↓ ROS | [64] |
| Potato | IFGPM, IDGGGI, HGPHIF, VDDDKDFIPF, LVTVDDDKD, VVTGGKVGNEND, potato protein hydrolysates (PPH) | Fe2+-chelating activity (55.33 µg EDTA/mL), ·OH scavenging activity (230.05 µg Vc/mL), and oxygen radical absorbance capacity (82.24 µg TE/mL) | [65] |
| Defatted walnut meal | AASCDQ | AQ exhibited strong antioxidant activity, which demonstrated significant scavenging ability against DPPH (79.40%), superoxide anion (81.00%), and ABTS (67.09%) radicals. | [66] |
| Cyperus (Cyperus esculentus L.) | SFRWQ | ↑ SOD, CAT, ↓ ROS, TNF-α, IL-6 | [67] |
| Coix seed (Coix lacryma-jobi L.) | FFDR | ↑ GSH, CAT, SOD, GSH-Px ↓ GSSG, MDA | [68] |
| Wheat gluten | LY, PY, YQ, APSY, RGGY | ↑ SOD, CAT, GSH-Px, GSH ↓ ROS, MDA | [69] |
| Moringa oleifera leaves | LALPVYN | ↑ CAT, GSH-Px, SOD ↓ MDA, ROS | [70] |
| Soybean | Leu-Ser-Trp (LSW) | Inhibited oxidative stress and reduced superoxide and malondialdehyde levels | [71] |
| Walnut | HGEPGQQQR VAPFPEVFGK HNVADPQR | Excellent cellular antioxidant activity | [72] |
| Functional Activity | Plant Origins | Molecular Characteristics | Structure–Activity Relationship | Reference |
|---|---|---|---|---|
| Antioxidant peptides | Pine nut (Pinus koraiensis Sieb. et Zucc.) | WYSGK | The presence of Ser increases its beta-pleated sheet content, and the active hydrogen atoms produce a chemical shift. | [134] |
| Antioxidant peptides | Watermelon seed | RDPEER (P1) | The active site of P1 is located at C6H14 on Arg. P1 can bind to DPPH/ABTS through hydrogen bonding and hydrophobic interaction. | [135] |
| Antioxidant peptides | Walnut (Juglans mandshurica Maxim.) | Walnut protein hydrolysate (<3 kDa) and peptides KGHLFPN | The tendency of the layer secondary structure to be randomly curled during digestion and the increase in active hydrogen content are favorable conditions for improving its antioxidant capacity. | [136] |
| Antioxidant peptides | Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn) | CTGFVAVR | CR-8 can enhance the antioxidant capacity of damaged cells by interfering with multiple metabolic pathways. This is associated with hydrophobic amino acids, N-terminal cysteine (Cys), and others. | [137] |
| Immunomodulatory peptides | Rice (Oryza sativa L.) | 11–20 amino acids | S, R, D, E, and T amino acids readily form hydrogen bonds with MHC-II molecules, thereby enhancing innate and adaptive immunity. | [138] |
| Immunomodulatory peptides | Soybean (Glycine max L.) | EKPQQQSSRRGS | EKPQQQQSSRRGS increases phagocytic activity of mouse spleen macrophages and also induces macrophage M1 polarization. | [139] |
| Immunomodulatory peptides | Sunflower seed (Helianthus annuus L.) | MVWGP | MVWGP is the most potent immunomodulatory peptide in all cellular assays, which is attributed to the presence of Met residues. | [140] |
| Antimicrobial peptides | Genus Ulva (Ulvophyceae, Chlorophyta) | HAVY RDRF | HAVY has a large number of hydrogen bonds as an antimicrobial agent and has shown higher antimicrobial efficacy than RDRF in in vitro validation. | [141] |
| Antimicrobial peptides | Moringa oleifera | HVLDTPLL | HVLDTPLL inhibited S. aureus growth by interacting with dihydrofolate reductase and DNA gyrase through hydrogen bonding and hydrophobic interactions. | [80] |
| Antimicrobial peptides | Chia seeds (Salvia hispanica L.) | <1 kDa | Significant inhibition effect was reported against Listeria monocytogenes for components with molecular weight < 1 kDa. | [142] |
| Anticancer peptides | Walnuts (Juglans regia L.) | <1 kDa | The <1 kDa molecular weight fraction (WPH-M1) exhibited more significant inhibition of HCT116 cell proliferation and induction of apoptosis than other fractions. | [143] |
| Anticancer peptides | Amaranth seed (Amaranthus caudutus L.) | Heat denaturation | Bioactive peptides from amaranth seed protein hydrolysates induced apoptosis and antimigratory effects in breast cancer cells. | [144] |
| Anticancer peptides | Corn gluten meal | 3–5 kDa | The fraction 3–5 kDa effectively inhibited the growth of HepG2 cancer cells. | [145] |
| Antidiabetic peptides | Highland barley | F-3 (DH-23.86%) | F-3 exhibited strong antiglycation activity, effectively suppressed the non-fluorescent AGE (CML) and the fructosamine level. | [146] |
| Antidiabetic peptides | Hemp (Cannabis sativa L.) seed meal | 99% of hydrolysate peptides have molecular weights < 5 kDa, 53.95% are 0.5–1 kDa | The peptides inhibited both intracellular disaccharidase and the transport of glucose at different concentrations. | [147] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Liu, J.; He, Z.; Du, R. Plant-Derived as Alternatives to Animal-Derived Bioactive Peptides: A Review of the Preparation, Bioactivities, Structure–Activity Relationships, and Applications in Chronic Diseases. Nutrients 2024, 16, 3277. https://doi.org/10.3390/nu16193277
Sun L, Liu J, He Z, Du R. Plant-Derived as Alternatives to Animal-Derived Bioactive Peptides: A Review of the Preparation, Bioactivities, Structure–Activity Relationships, and Applications in Chronic Diseases. Nutrients. 2024; 16(19):3277. https://doi.org/10.3390/nu16193277
Chicago/Turabian StyleSun, Li, Jinze Liu, Zhongmei He, and Rui Du. 2024. "Plant-Derived as Alternatives to Animal-Derived Bioactive Peptides: A Review of the Preparation, Bioactivities, Structure–Activity Relationships, and Applications in Chronic Diseases" Nutrients 16, no. 19: 3277. https://doi.org/10.3390/nu16193277
APA StyleSun, L., Liu, J., He, Z., & Du, R. (2024). Plant-Derived as Alternatives to Animal-Derived Bioactive Peptides: A Review of the Preparation, Bioactivities, Structure–Activity Relationships, and Applications in Chronic Diseases. Nutrients, 16(19), 3277. https://doi.org/10.3390/nu16193277