
Health-Promoting Effects, Phytochemical Constituents and Molecular Genetic Profile of the Purple Carrot ‘Purple Sun’ (Daucus carota L.)

 ,
,  ,
,  ,
,  ,
,  ,
,  , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Molecular Genetic Analysis of Purple Carrot Plants
2.3. Preparation of Purple Carrot Extract (PCE)
2.4. LC-ESI/LTQOrbitrap/MS/MS Analysis
2.5. Reactive Oxygen Species (ROS) and Antioxidant Enzymes in Polymorphonuclear Leukocytes after Opsonized Zymosan Stress (OZ-Stressed PMNs)
2.6. Antimicrobial Activity Assays: Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Determination
2.7. Cell Lines, Culture Conditions
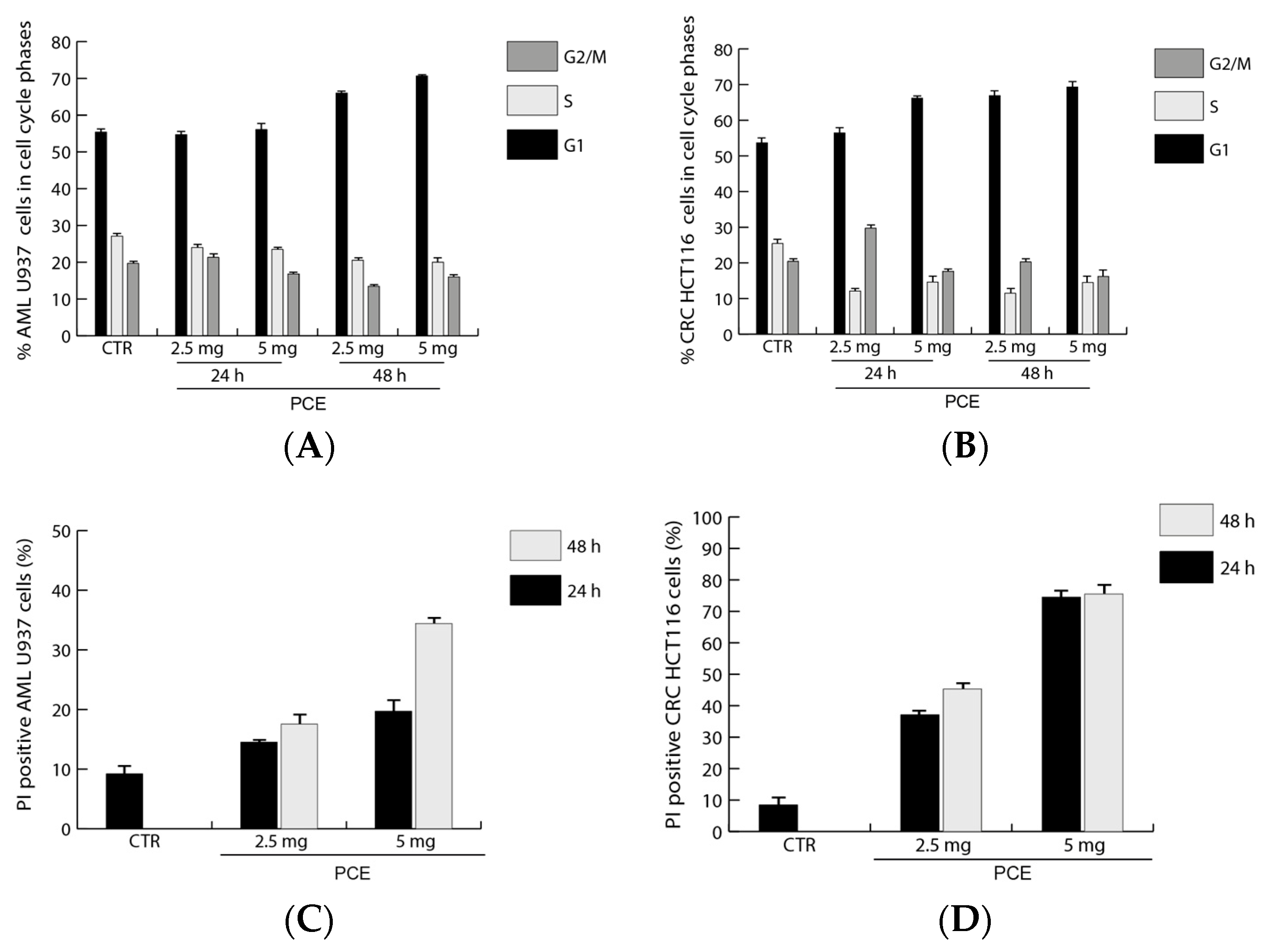
2.8. Cell Cycle Analysis
2.9. Total Protein Extraction and Western Blot Analysis
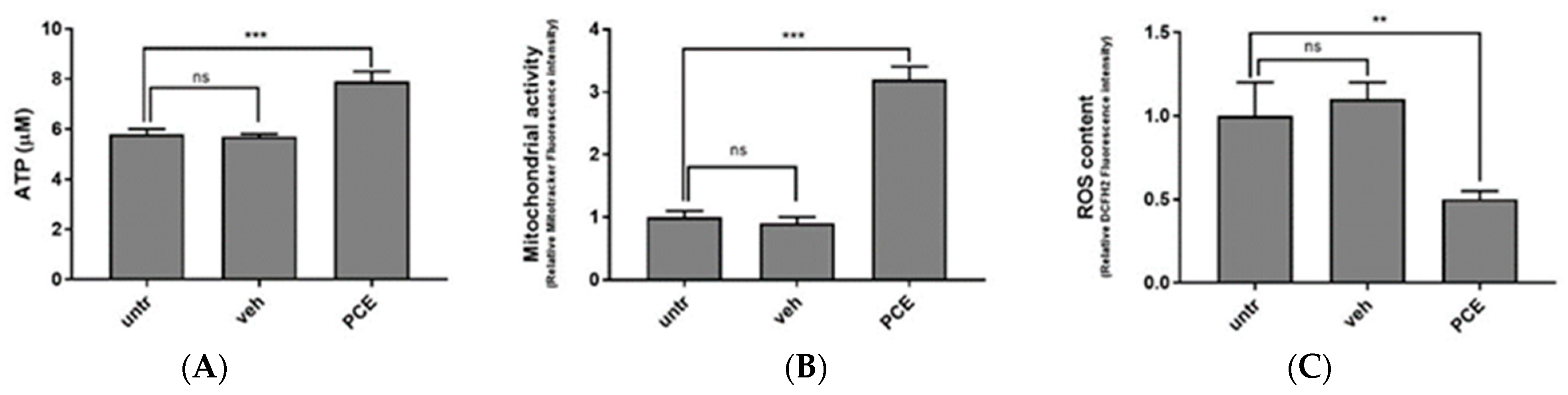
2.10. ATP and Mitochondrial Stimulatory Activity of PCE in Muscle Cells
2.11. Selection of Cyanidin Protein Targets and Molecular Docking
2.12. Statistical Analysis
3. Results and Discussion
3.1. Assessment of Genetic Diversity among Purple Carrot Plants
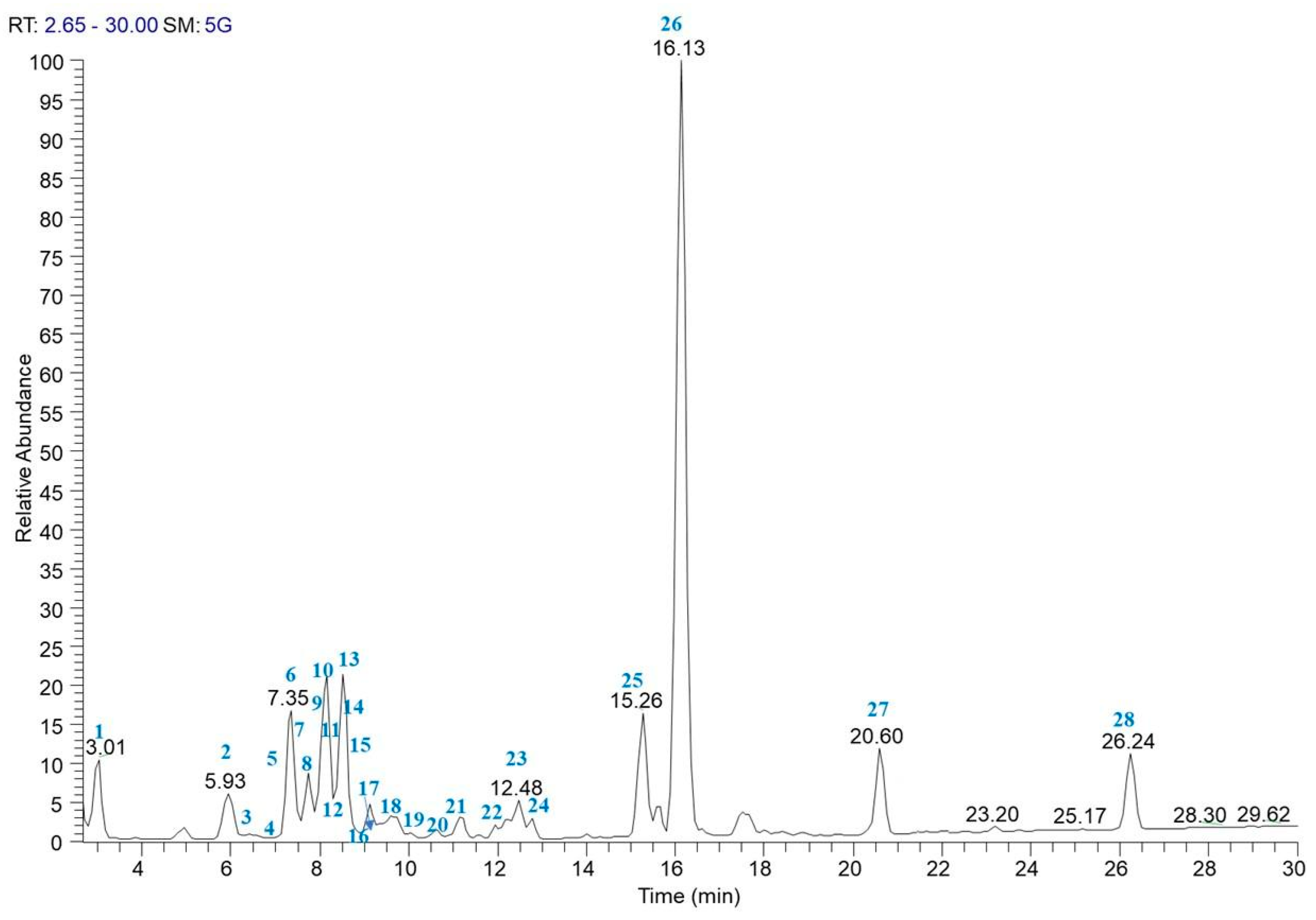
3.2. LC-ESI/LTQOrbitrap/MS/MS Analysis of Purple Carrot Extract
3.3. Biological Activities of Purple Carrot Extract (PCE) and Potential Beneficial Effects on Human Health
3.3.1. Characterization of Antioxidant Activity of PCE
3.3.2. Determination of Antibacterial Activity of PCE
3.3.3. Analysis of Antiproliferative Activity of PCE on Hematological and Epithelial Cancer Cells
3.3.4. Prediction of Cyanidin Protein Targets and Molecular Docking
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alamnia, T.T.; Sargent, G.M.; Kelly, M. Dietary patterns and associations with metabolic risk factors for non-communicable disease. Sci. Rep. 2023, 13, 21028. [Google Scholar] [CrossRef]
- Zhang, Y.; Giovannucci, E.L. Ultra-processed foods and health: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2022, 63, 10836–10848. [Google Scholar] [CrossRef]
- D’Amelia, V.; Aversano, R.; Chiaiese, P.; Carputo, D. The antioxidant properties of plant flavonoids: Their exploitation by molecular plant breeding. Phytochem. Rev. 2018, 17, 611–625. [Google Scholar] [CrossRef]
- Bontempo, P.; De Masi, L.; Rigano, D. Functional Properties of Natural Products and Human Health. Nutrients 2023, 15, 2961. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the Art of Anthocyanins: Antioxidant Activity, Sources, Bioavailability, and Therapeutic Effect in Human Health. Antioxidants 2020, 9, 451. [Google Scholar] [CrossRef]
- Câmara, J.S.; Locatelli, M.; Pereira, J.A.M.; Oliveira, H.; Arlorio, M.; Fernandes, I.; Perestrelo, R.; Freitas, V.; Bordiga, M. Behind the Scenes of Anthocyanins—From the Health Benefits to Potential Applications in Food, Pharmaceutical and Cosmetic Fields. Nutrients 2022, 14, 5133. [Google Scholar] [CrossRef]
- Bontempo, P.; Carafa, V.; Grassi, R.; Basile, A.; Tenore, G.C.; Formisano, C.; Rigano, D.; Altucci, L. Antioxidant, antimicrobial and antiproliferative activities of Solanum tuberosum L. var. Vitelotte. Food Chem. Toxicol. 2013, 55, 304–312. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The Therapeutic Potential of Anthocyanins: Current Approaches Based on Their Molecular Mechanism of Action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef]
- Bontempo, P.; De Masi, L.; Carafa, V.; Rigano, D.; Scisciola, L.; Iside, C.; Grassi, R.; Molinari, A.M.; Aversano, R.; Nebbioso, A.; et al. Anticancer activities of anthocyanin extract from genotyped Solanum tuberosum L. “Vitelotte”. J. Funct. Foods 2015, 19, 584–593. [Google Scholar] [CrossRef]
- Schmutz, C.; Will, F.; Varga, E.; Jaunecker, C.; Pahlke, G.; Berger, W.; Marko, D. In Vitro Inhibitory Potential of Different Anthocyanin-Rich Berry Extracts in Murine CT26 Colon Cancer Cells. Molecules 2023, 28, 7684. [Google Scholar] [CrossRef]
- Grzebelus, D.; Iorizzo, M.; Senalik, D.; Ellison, S.; Cavagnaro, P.; Macko-Podgorni, A.; Heller-Uszynska, K.; Kilian, A.; Nothnagel, T.; Allender, C.; et al. Diversity, genetic mapping, and signatures of domestication in the carrot (Daucus carota L.) genome, as revealed by Diversity Arrays Technology (DArT) markers. Mol. Bred. 2014, 33, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Tariq, R.M.S.; Azam, M.; Akhtar, S. Phytochemicals in Daucus carota and Their Health Benefits—Review Article. Foods 2019, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Crupi, P.; Faienza, M.F.; Naeem, M.Y.; Corbo, F.; Clodoveo, M.L.; Muraglia, M. Overview of the Potential Beneficial Effects of Carotenoids on Consumer Health and Well-Being. Antioxidants 2023, 12, 1069. [Google Scholar] [CrossRef] [PubMed]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of Many Colors Provide Basic Nutrition and Bioavailable Phytochemicals Acting as a Functional Food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Sirignano, C.; Hammami, S.; El Mokni, R.; Blagborough, A.M.; Luciano, P.; Rigano, D.; Scafati, O.T. Polyoxygenated germacranes from Daucus carota and their antimalarial transmission blocking activity. Phytochemistry 2021, 183, 112632. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.B.; Carvajal, S.; Beretta, V.; Bannoud, F.; Fangio, M.F.; Berli, F.; Fontana, A.; Salomón, M.V.; Gonzalez, R.; Valerga, L.; et al. Characterization of Purple Carrot Germplasm for Antioxidant Capacity and Root Concentration of Anthocyanins, Phenolics, and Carotenoids. Plants 2023, 12, 1796. [Google Scholar] [CrossRef] [PubMed]
- Soleti, R.; Mallegol, P.; Hilairet, G.; Frifra, M.; Perrin, F.; Dubois-Laurent, C.; Huet, S.; Pignon, P.; Basset, L.; Geoffriau, E.; et al. Carrot Genotypes Contrasted by Root Color and Grown under Different Conditions Displayed Differential Pharmacological Profiles in Vascular and Metabolic Cells. Nutrients 2020, 12, 337. [Google Scholar] [CrossRef] [PubMed]
- Nunziata, A.; Ruggieri, V.; Petriccione, M.; De Masi, L. Single Nucleotide Polymorphisms as Practical Molecular Tools to Support European Chestnut Agrobiodiversity Management. Int. J. Mol. Sci. 2020, 21, 4805. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- De Masi, L.; Siviero, P.; Castaldo, D.; Cautela, D.; Esposito, C.; Laratta, B. Agronomic, chemical and genetic profiles of hot peppers (Capsicum annuum ssp.). Mol. Nutr. Food Res. 2007, 51, 1053–1062. [Google Scholar] [CrossRef]
- Kolodziejczyk, J.; Masullo, M.; Olas, B.; Piacente, S.; Wachowicz, B. Effects of garcinol and guttiferone K isolated from Garcinia cambogia on oxidative/nitrative modifications in blood platelets and plasma. Platelets 2009, 20, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy: The Principles and Practice of Numerical Classification, 1st ed.; W. H. Freeman: San Francisco, CA, USA, 1973; ISBN 978-0716706977. [Google Scholar]
- PAST 4. Available online: https://www.nhm.uio.no/english/research/resources/past/ (accessed on 5 June 2023).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Sokal, R.R.; Michener, C.D. A Statistical Method for Evaluating Relationships. Univ. Kans. Sci. Bull. 1958, 38, 1409–1448. [Google Scholar]
- Kjell, T.; Øyvind, M.A. Color stability of anthocyanins in aqueous solutions at various pH values. Food Chem. 2005, 89, 427–440. [Google Scholar]
- Hosseini, S.H.; Masullo, M.; Cerulli, A.; Martucciello, S.; Ayyari, M.; Pizza, C.; Piacente, S. Antiproliferative cardenolides from the aerial parts of Pergularia tomentosa. J. Nat. Prod. 2019, 82, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Masullo, M.; Lauro, G.; Cerulli, A.; Bifulco, G.; Piacente, S. Corylus avellana: A Source of Diarylheptanoids With α-Glucosidase Inhibitory Activity Evaluated by in vitro and in silico Studies. Front. Plant Sci. 2022, 13, 805660. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Russi, S.; Bruno, M.; Maresca, V.; Vaglica, A.; Ilardi, V.; Zanfardino, A.; Di Napoli, M.; Varcamonti, M.; Cianciullo, P.; et al. Dihydrophenanthrenes from a Sicilian accession of Himantoglossum robertianum (Loisel.) P. Delforge showed antioxidant, antimicrobial, and antiproliferative activities. Plants 2021, 10, 2776. [Google Scholar] [CrossRef] [PubMed]
- Manna, A.; Saha, P.; Sarkar, A.; Mukhopadhyay, D.; Bauri, A.K.; Kumar, D.; Das, P.; Chattopadhyay, S.; Chatterjee, M. Malabaricone-A induces a redox imbalance that mediates apoptosis in U937 Cell Line. PLoS ONE 2012, 7, e36938. [Google Scholar] [CrossRef]
- Napolitano, A.; Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Cicio, A.; Bruno, M.; Piacente, S.; Maresca, V.; Cianciullo, P.; Capasso, L.; et al. The chemical composition of the aerial parts of Stachys spreitzenhoferi (Lamiaceae) growing in Kythira Island (Greece), and their antioxidant, antimicrobial, and antiproliferative properties. Phytochemistry 2022, 203, 113373. [Google Scholar] [CrossRef]
- Barbosa, P.O.; Pala, D.; Silva, C.T.; Oliveira de Souza, M.; Ferreira do Amaral, J.; Lima Vieira, R.A.; Andrezza de Freitas Folly, G.; Pinheiro Volp, A.C.; Nascimento de Freitas, R. Açai (Euterpe oleracea Mart.) pulp dietary intake improves cellular antioxidant enzymes and biomarkers of serum in healthy women. Nutrition 2016, 32, 674–680. [Google Scholar] [CrossRef]
- Ieven, M.; Vanden Berghe, D.A.; Mertens, F.; Vlietinck, A.; Lammens, E. Screening of higher plants for biological activities. I. Antimicrobial activity. Planta Med. 1979, 36, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Ericcson, H.M.; Sherris, J.C. Antibiotic sensitivity testing: Report of an international collaborative study. Acta Pathol. Microbiol. Scand. 1971, 217, 1–90. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- De Masi, L.; Castaldo, D.; Galano, G.; Minasi, P.; Laratta, B. Genotyping of fig (Ficus carica L) via RAPD markers. J. Sci. Food Agric. 2005, 85, 2235–2242. [Google Scholar] [CrossRef]
- Shim, S.; Jørgensen, R. Genetic structure in cultivated and wild carrots (Daucus carota L.) revealed by AFLP analysis. Theor. Appl. Genet. 2000, 101, 227–233. [Google Scholar] [CrossRef]
- Iorizzo, M.; Senalik, D.A.; Ellison, S.L.; Grzebelus, D.; Cavagnaro, P.F.; Allender, C.; Brunet, J.; Spooner, D.M.; Van Deynze, A.; Simon, P.W. Genetic structure and domestication of carrot (Daucus carota subsp. sativus) (Apiaceae). Am. J. Bot. 2013, 100, 930–938. [Google Scholar] [CrossRef]
- Domblides, A.; Domblides, E. Rapid Genetic Assessment of Carrot Varieties Based on AFLP Analysis. Horticulturae 2023, 9, 298. [Google Scholar] [CrossRef]
- Stelmach, K.; Macko-Podgórni, A.; Allender, C.; Grzebelus, D. Genetic diversity structure of western-type carrots. BMC Plant Biol. 2021, 21, 200. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.S.; Schug, K.A. Structural characterization of cyanidin-3,5-diglucoside and pelargonidin-3,5-diglucoside anthocyanins: Multi-dimensional fragmentation pathways using high performance liquid chromatography-electrospray ionization-ion trap-time of flight mass spectrometry. Int. J. Mass Spectrom. 2011, 308, 71–80. [Google Scholar] [CrossRef]
- Algarra, M.; Fernandes, A.; Mateus, N.; de Freitas, V.; Esteves da Silva, J.C.G.; Casado, J. Anthocyanin profile and antioxidant capacity of black carrots (Daucus carota L. ssp. sativus var. atrorubens Alef.) from Cuevas Bajas, Spain. J. Food Compos. Anal. 2014, 33, 71–76. [Google Scholar] [CrossRef]
- Yusuf, E.; Wojdylo, A.; Oszmianski, J.; Nowicka, P. Nutritional, phytochemical characteristics and in vitro effect on α-amylase, α-glucosidase, lipase, and cholinesterase activities of 12 coloured carrot varieties. Foods 2021, 10, 808. [Google Scholar] [CrossRef]
- Cerulli, A.; Napolitano, A.; Hosek, J.; Masullo, M.; Pizza, C.; Piacente, S. Antioxidant and in vitro preliminary anti-inflammatory activity of Castanea sativa (Italian cultivar “Marrone di Roccadaspide” PGI) burs, leaves and chestnuts extracts and their metabolite profiles by LC-ESI/LTQOrbitrap/MS/MS. Antioxidants 2021, 10, 278. [Google Scholar] [CrossRef]
- Masullo, M.; Cerulli, A.; Pizza, C.; Piacente, S. Pouteria lucuma Pulp and Skin: In Depth Chemical Profile and Evaluation of Antioxidant Activity. Molecules 2021, 26, 5236. [Google Scholar] [CrossRef]
- Cerulli, A.; Lauro, G.; Masullo, M.; Cantone, V.; Olas, B.; Kontek, B.; Nazzaro, F.; Bifulco, G.; Piacente, S. Cyclic diarylheptanoids from Corylus avellana green leafy covers: Determination of their absolute configurations and evaluation of their antioxidant and antimicrobial activities. J. Nat. Prod. 2017, 80, 1703–1713. [Google Scholar] [CrossRef]
- Tian, Y.; Kriisa, M.; Foste, M.; Kutt, M.-L.; Zhou, Y.; Laaksonen, O.; Yang, B. Impact of enzymatic pre-treatment on composition of nutrients and phytochemicals of canola (Brassica napus) oil press residues. Food Chem. 2022, 387, 132911. [Google Scholar] [CrossRef]
- Mizgier, P.; Kucharska, A.Z.; Sokół-Łętowska, A.; Kolniak-Ostek, J.; Kidoń, M.; Fecka, I. Characterization of phenolic compounds and antioxidant and anti-inflammatory properties of red cabbage and purple carrot extracts. J. Funct. Foods 2016, 21, 133–146. [Google Scholar] [CrossRef]
- Fayek, N.M.; Farag, M.A.; Monem, A.R.A.; Moussa, M.Y.; Abd-Elwahab, S.M.; El-Tanbouly, N.D. Comparative metabolite profiling of four Citrus peel cultivars via ultra-performance liquid chromatography coupled with quadrupole- time-of-flight-mass spectrometry and multivariate data analyses. J. Chromatogr. Sci. 2019, 57, 349–360. [Google Scholar] [CrossRef]
- Cho, J.-G.; Cha, B.-J.; Seo, W.-D.; Jeong, R.-H.; Shrestha, S.; Kim, J.-Y.; Kang, H.-C.; Baek, N.-I. Feruloyl sucrose esters from Oryza sativa roots and their tyrosinase inhibition activity. Chem. Nat. Compd. 2015, 51, 1094–1098. [Google Scholar] [CrossRef]
- Szczepańska, J.; Barba, F.J.; Skąpska, S.; Marszałek, K. High pressure processing of carrot juice: Effect of static and multi-pulsed pressure on the polyphenolic profile, oxidoreductases activity and colour. Food Chem. 2020, 307, 125549. [Google Scholar] [CrossRef]
- Średnicka-Tober, D.; Kopczyńska, K.; Góralska-Walczak, R.; Hallmann, E.; Barański, M.; Marszałek, K.; Kazimierczak, R. Are Organic Certified Carrots Richer in Health-Promoting Phenolics and Carotenoids than the Conventionally Grown Ones? Molecules 2022, 27, 4184. [Google Scholar] [CrossRef]
- Grebenstein, C.; Choi, Y.H.; Rong, J.; de Jong, T.J.; Tamis, W.L.M. Metabolic fingerprinting reveals differences between shoots of wild and cultivated carrot (Daucus carota L.) and suggests maternal inheritance or wild trait dominance in hybrids. Phytochemistry 2011, 72, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Zhang, L.; Fu, H.-W.; Yang, S.-L.; Tian, J.-K. Two New Guaiane Sesquiterpenes from the Fruits of Daucus carota. Helv. Chim. Acta 2009, 92, 2769–2773. [Google Scholar] [CrossRef]
- Montilla, E.C.; Arzaba, M.R.; Hillebrand, S.; Winterhalter, P. Anthocyanin Composition of Black Carrot (Daucus carota ssp. sativus var. atrorubens Alef.) Cultivars Antonina, Beta Sweet, Deep Purple, and Purple Haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, M.; Maresca, V.; Basile, A.; Pacifico, S.; Fiorentino, A.; Bruno, M.; Badalamenti, N.; Kapelusz, M.; Marino, P.; Capasso, L.; et al. Phytochemical Characterization, Antioxidant, and Anti-Proliferative Activities of Wild and Cultivated Nigella damascene Species Collected in Sicily (Italy). Antioxidants 2024, 13, 402. [Google Scholar] [CrossRef]
- Damiano, S.; Muscariello, E.; La Rosa, G.; Di Maro, M.; Mondola, P.; Santillo, M. Dual Role of Reactive Oxygen Species in Muscle Function: Can Antioxidant Dietary Supplements Counteract Age-Related Sarcopenia? Int. J. Mol. Sci. 2019, 20, 3815. [Google Scholar] [CrossRef]
- Lian, D.; Chen, M.-M.; Wu, H.; Deng, S.; Hu, X. The Role of Oxidative Stress in Skeletal Muscle Myogenesis and Muscle Disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Islam, P.; Subhan, N.; Rahman, M.M.; Khan, F.; Burrows, G.E.; Nahar, L.; Sarker, S.D. Potential health benefits of anthocyanins in oxidative stress related disorders. Phytochem. Rev. 2021, 20, 705–749. [Google Scholar] [CrossRef]
- Rasheed, H.; Shehzad, M.; Rabail, R.; Kowalczewski, P.Ł.; Kidoń, M.; Jeżowski, P.; Ranjha, M.M.A.N.; Rakha, A.; Din, A.; Aadil, R.M. Delving into the Nutraceutical Benefits of Purple Carrot against Metabolic Syndrome and Cancer: A Review. Appl. Sci. 2022, 12, 3170. [Google Scholar] [CrossRef]
- De Masi, L.; Bontempo, P.; Rigano, D.; Stiuso, P.; Carafa, V.; Nebbioso, A.; Piacente, S.; Montoro, P.; Aversano, R.; D’Amelia, V.; et al. Comparative Phytochemical Characterization, Genetic Profile, and Antiproliferative Activity of Polyphenol-Rich Extracts from Pigmented Tubers of Different Solanum tuberosum Varieties. Molecules 2020, 25, 233. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.I.; Hartl, M. Strategies to target the cancer driver MYC in tumor cells. Front. Oncol. 2023, 13, 1142111. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Bates, S.; Peters, G. Evidence for different modes of action of cyclin-dependent kinase inhibitors: p15 and p16 bind to kinases, p21 and p27 bind to cyclins. Oncogene 1995, 11, 1581–1588. [Google Scholar]
- You, W.; Zheng, W.; Weiss, S.; Chua, K.F.; Steegborn, C. Structural basis for the activation and inhibition of Sirtuin 6 by quercetin and its derivatives. Sci. Rep. 2019, 9, 19176. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.L.; Fivush, A.M.; Watson, M.A.; Galardi, C.M.; Lewis, M.C.; Moore, L.B.; Parks, D.J.; Wilson, J.G.; Tippin, T.K.; Binz, J.G.; et al. Identification of a nonsteroidal liver X receptor agonist through parallel array synthesis of tertiary amines. J. Med. Chem. 2002, 45, 1963–1966. [Google Scholar] [CrossRef] [PubMed]
- Fradera, X.; Vu, D.; Nimz, O.; Skene, R.; Hosfield, D.; Wynands, R.; Cooke, A.J.; Haunsø, A.; King, A.; Bennett, D.J.; et al. X-ray structures of the LXRalpha LBD in its homodimeric form and implications for heterodimer signaling. J. Mol. Biol. 2010, 399, 120–132. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Wang, X.; Liu, Y.; Xia, M. Cyanidin-3-O-β-glucoside induces oxysterol efflux from endothelial cells: Role of liver X receptor alpha. Atherosclerosis 2012, 223, 299–305. [Google Scholar] [CrossRef]
- Senger, S.; Chan, C.; Convery, M.A.; Hubbard, J.A.; Shah, G.P.; Watson, N.S.; Young, R.J. Sulfonamide-related conformational effects and their importance in structure-based design. Bioorganic Med. Chem. Lett. 2007, 17, 2931–2934. [Google Scholar] [CrossRef] [PubMed]
- Bijak, M.; Ziewiecki, R.; Saluk, J.; Ponczek, M.; Pawlaczyk, I.; Krotkiewski, H.; Wachowicz, B.; Nowak, P. Thrombin inhibitory activity of some polyphenolic compounds. Med. Chem. Res. 2014, 23, 2324–2337. [Google Scholar] [CrossRef] [PubMed]
- Bijak, M.; Ponczek, M.B.; Nowak, P. Polyphenol compounds belonging to flavonoids inhibit activity of coagulation factor X. Int. J. Biol. Macromol. 2014, 65, 129–135. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | CAR1 | CAR2 | CAR3 | CAR4 | CAR5 | CAR6 | CAR7 | CAR8 | CAR9 | CAR10 | Polymorphic Loci/Total Loci | Private Loci |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AK10 | 6 | 5 | 6 | 7 | 5 | 4 | 6 | 8 | 5 | 5 | 7/10 (70%) | 1 |
| E10 | 8 | 9 | 8 | 8 | 9 | 7 | 7 | 8 | 8 | 8 | 3/10 (30%) | 1 |
| E11 | 8 | 9 | 9 | 9 | 9 | 9 | 10 | 9 | 8 | 8 | 5/12 (42%) | 2 |
| E14 | 9 | 8 | 11 | 11 | 9 | 9 | 8 | 8 | 5 | 9 | 10/12 (83%) | 0 |
| U1 | 7 | 7 | 7 | 8 | 8 | 7 | 10 | 11 | 7 | 8 | 9/13 (69%) | 1 |
| U3 | 7 | 8 | 8 | 10 | 8 | 9 | 7 | 7 | 7 | 9 | 4/10 (40%) | 0 |
| U4 | 7 | 10 | 7 | 6 | 6 | 9 | 6 | 8 | 8 | 7 | 9/11 (82%) | 0 |
| U19 | 7 | 9 | 8 | 8 | 8 | 9 | 9 | 10 | 8 | 8 | 6/12 (50%) | 0 |
| V01 | 6 | 5 | 7 | 6 | 5 | 5 | 6 | 8 | 5 | 6 | 12/12 (100%) | 0 |
| V06 | 6 | 4 | 5 | 5 | 5 | 6 | 6 | 6 | 6 | 6 | 6/9 (67%) | 0 |
| Private bands | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 5/111 (4.5%) | |
| Total bands | 71 | 74 | 76 | 78 | 72 | 74 | 75 | 83 | 67 | 74 | 71/111 (64%) |
| # | Compound | Rt (min) | Molecular Formula | [M − H]− | [M − 2H]− | Error (ppm) | Characteristic Product Ions |
|---|---|---|---|---|---|---|---|
| 1 | isocitric acid ** | 3.01 | C6H8O7 | 191.1200 | 4.5 | 147, 111 | |
| 2 | protocatechuic acid 4-O-glucoside ** | 5.93 | C13H16O9 | 315.0722 | 3.53 | 153 | |
| 3 | cyanidin 3-O-xylosyl-sinapoylglucosyl-galactoside * | 7.05 | C43H49O24+ | 947.2468 | 1.68 | 785, 285 | |
| 4 | cyanidin 3-O-xylosyl-feruloylglucosyl-galactoside * | 7.35 | C42H47O23+ | 917.2361 | 1.67 | 285 | |
| 5 | cyanidin 3-caffeoylsophoroside 5-glucoside * | 11.60 | C42H47O24+ | 933.2008 | 1.33 | 285 | |
| 6 | 4-hydroxybenzoic acid 4-O-glucoside ** | 7.50 | C13H16O8 | 299.0772 | 3.66 | 137 | |
| 7 | cyanidin 3-O-xylosyl-p-coumaroylglucosyl-galactoside * | 7.50 | C41H45O22+ | 887.2259 | 2.14 | 527, 285 | |
| 8 | pelargonidin 3-O-xylosyl-feruloylglucosyl-galactoside * | 8.86 | C42H47O22+ | 901.2408 | 1.20 | 269 | |
| 9 | sinapoyl-O-hexoside | 8.63 | C17H22O10 | 385.1193 | 2.13 | 205 | |
| 10 | 5-caffeoylquinic acid * | 8.02 | C16H18O9 | 353.0881 | 3.94 | 191, 135 | |
| 11 | peonidin 3-O-xylosyl-galactoside * | 8.09 | C27H31O15+ | 593.1665 | 1.27 | 301 | |
| 12 | feruloyl-O-di-hexosyl-pentoside | 8.63 | C27H38O18 | 649.1987 | 1.29 | 499, 397, 355, 193 | |
| 13 | feruloyl-O-di-hexoside * | 8.55 | C22H30O14 | 517.1563 | 2.16 | 337, 193 | |
| 14 | feruloyl-O-hexoside * | 8.73 | C16H20O9 | 355.1032 | 2.48 | 175 | |
| 15 | dihydromyricetin | 8.88 | C15H12O8 | 319.0457 | 2.75 | 285, 193 | |
| 16 | feruloyl-O-rutinoside | 9.12 | C22H30O13 | 501.1609 | 2.22 | 397, 193 | |
| 17 | ferulic acid * | 9.12 | C10H10O4 | 193.0504 | 4.38 | 178, 149 | |
| 18 | cyanidin-3-O-xylosyl-galactoside * | 9.35 | C26H29O15+ | 579.1358 | 3.73 | 417, 285 | |
| 19 | 5-O-feruloylquinic acid * | 9.63 | C17H20O9 | 367.1035 | 2.97 | 193 | |
| 20 | Rutin * | 10.05 | C27H30O16 | 609.1456 | 1.02 | 463, 301 | |
| 21 | methoxy-dihydromyricetin | 11.94 | C16H14O8 | 333.0615 | 3.10 | 283, 185 | |
| 22 | diferuloyl-O-sucrose | 12.17 | C32H38O17 | 693.2035 | 1.39 | 517, 499, 337 | |
| 23 | diferuloyl-O-hexoside | 12.48 | C26H28O12 | 531.1512 | 2.67 | 351, 193 | |
| 24 | isorhamnetin-O-hexuronoside | 12.79 | C23H24O12 | 491.1193 | 1.75 | 459, 315, 151 | |
| 25 | trihydroxyoctadecadienoic acid (TriHODE) | 15.26 | C18H32O5 | 327.2178 | 3.66 | 291, 229, 211, 171 | |
| 26 | trihydroxyoctanoic acid (TriHOME) | 16.13 | C18H34O5 | 329.2335 | 3.85 | 293, 229, 211, 171 | |
| 27 | 11-acetyloxytorilolone ** | 20.60 | C17H26O4 | 293.1759 | 4.07 | 236, 221 | |
| 28 | hydroxyoctadecadienoic acid(HODE) | 26.24 | C18H32O3 | 295.2278 | 3.62 | 277, 195, 171 |
| MIC | MBC | |||||
|---|---|---|---|---|---|---|
| ATCC | PCE | CTAX | PENG | TET | PCE | |
| Staphylococcus aureus | 13709 | 3.9 | 2 | 0.03 | 2 | 62.5 |
| Enterococcus faecalis | 14428 | 3.9 | R | 8 | 2 | R |
| Proteus vulgaris | 12454 | 15.6 | 2 | 4 | R | R |
| Proteus mirabilis | 7002 | 15.6 | 0.03 | 4 | 32 | R |
| Salmonella typhi | 19430 | 31.3 | 0.5 | 4 | 1 | R |
| Enterobacter cloacae | 10699 | 15.6 | R | 4 | R | R |
| Enterobacter aerogenes | 13048 | 31.3 | R | 4 | R | R |
| Pseudomonas aeruginosa | 27853 | 62.5 | 16 | R | 32 | R |
| Klebsiella pneumoniae | 27736 | 62.5 | 0.1 | R | 16 | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maresca, V.; Capasso, L.; Rigano, D.; Stornaiuolo, M.; Sirignano, C.; Piacente, S.; Cerulli, A.; Marallo, N.; Basile, A.; Nebbioso, A.; et al. Health-Promoting Effects, Phytochemical Constituents and Molecular Genetic Profile of the Purple Carrot ‘Purple Sun’ (Daucus carota L.). Nutrients 2024, 16, 2505. https://doi.org/10.3390/nu16152505
Maresca V, Capasso L, Rigano D, Stornaiuolo M, Sirignano C, Piacente S, Cerulli A, Marallo N, Basile A, Nebbioso A, et al. Health-Promoting Effects, Phytochemical Constituents and Molecular Genetic Profile of the Purple Carrot ‘Purple Sun’ (Daucus carota L.). Nutrients. 2024; 16(15):2505. https://doi.org/10.3390/nu16152505
Chicago/Turabian StyleMaresca, Viviana, Lucia Capasso, Daniela Rigano, Mariano Stornaiuolo, Carmina Sirignano, Sonia Piacente, Antonietta Cerulli, Nadia Marallo, Adriana Basile, Angela Nebbioso, and et al. 2024. "Health-Promoting Effects, Phytochemical Constituents and Molecular Genetic Profile of the Purple Carrot ‘Purple Sun’ (Daucus carota L.)" Nutrients 16, no. 15: 2505. https://doi.org/10.3390/nu16152505
APA StyleMaresca, V., Capasso, L., Rigano, D., Stornaiuolo, M., Sirignano, C., Piacente, S., Cerulli, A., Marallo, N., Basile, A., Nebbioso, A., Giordano, D., Facchiano, A., De Masi, L., & Bontempo, P. (2024). Health-Promoting Effects, Phytochemical Constituents and Molecular Genetic Profile of the Purple Carrot ‘Purple Sun’ (Daucus carota L.). Nutrients, 16(15), 2505. https://doi.org/10.3390/nu16152505