Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
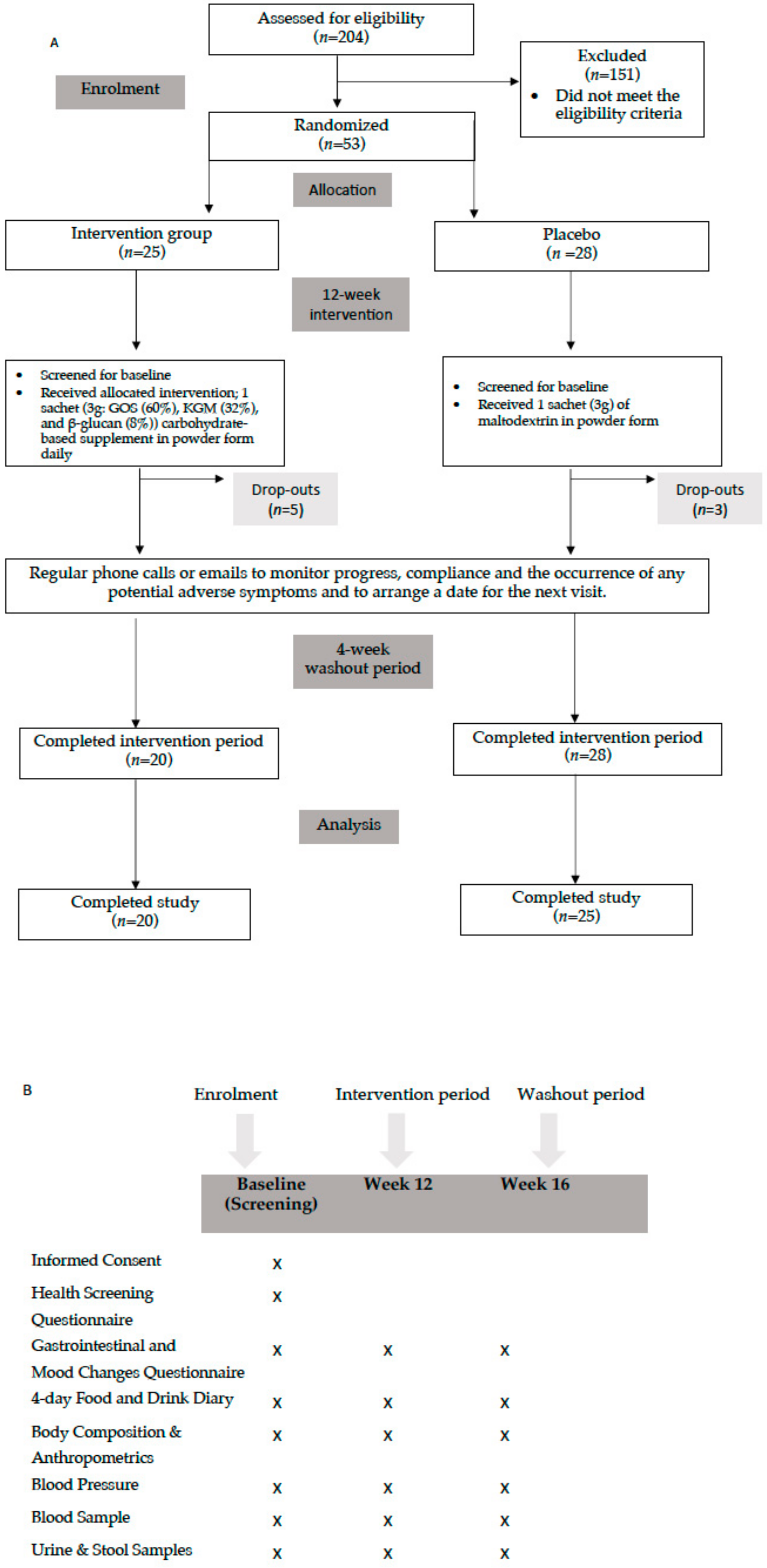
2.1. Study Design and Ethics
2.2. Inclusion and Exclusion Criteria
2.3. Study Participants
2.4. Intervention
2.5. Outcomes
2.5.1. Samples Collection for Blood Lipids, Immune/Inflammatory Markers, and Gut Microbiota Assessments
2.5.2. Glucose Biomarkers
2.5.3. Intestinal Permeability
2.5.4. Gut Microbiota Profiling
2.5.5. Anthropometric and Body Composition Analyses
2.5.6. Blood Pressure (BP)
2.5.7. Dietary Intake and Analyses
2.5.8. Gastrointestinal Changes (GI)
2.5.9. Statistical Analysis
3. Results
3.1. Subjects
3.2. Outcomes
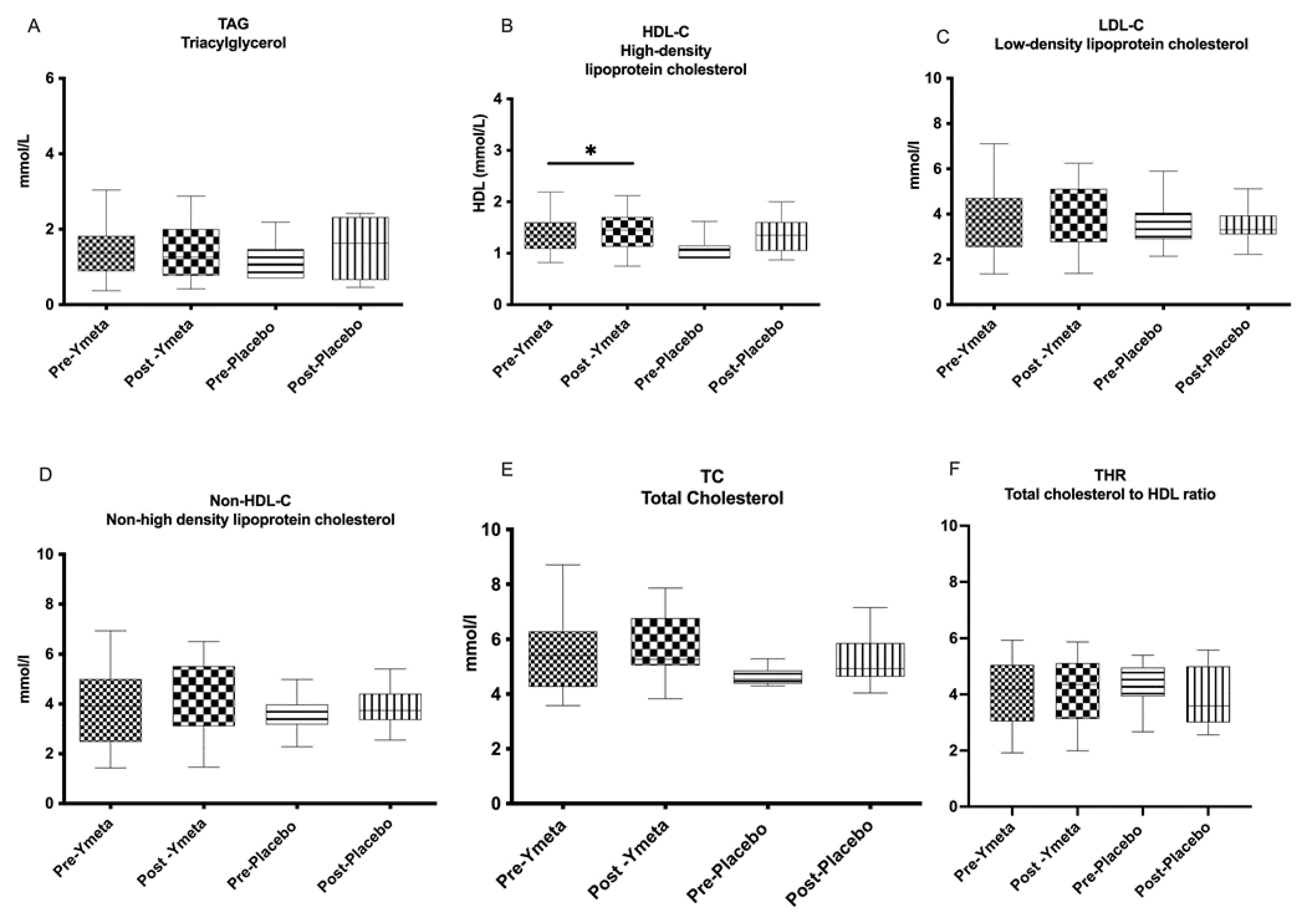
3.2.1. Blood Lipids and Immune/Inflammatory Markers
3.2.2. Glucose Biomarkers
3.2.3. Intestinal Permeability
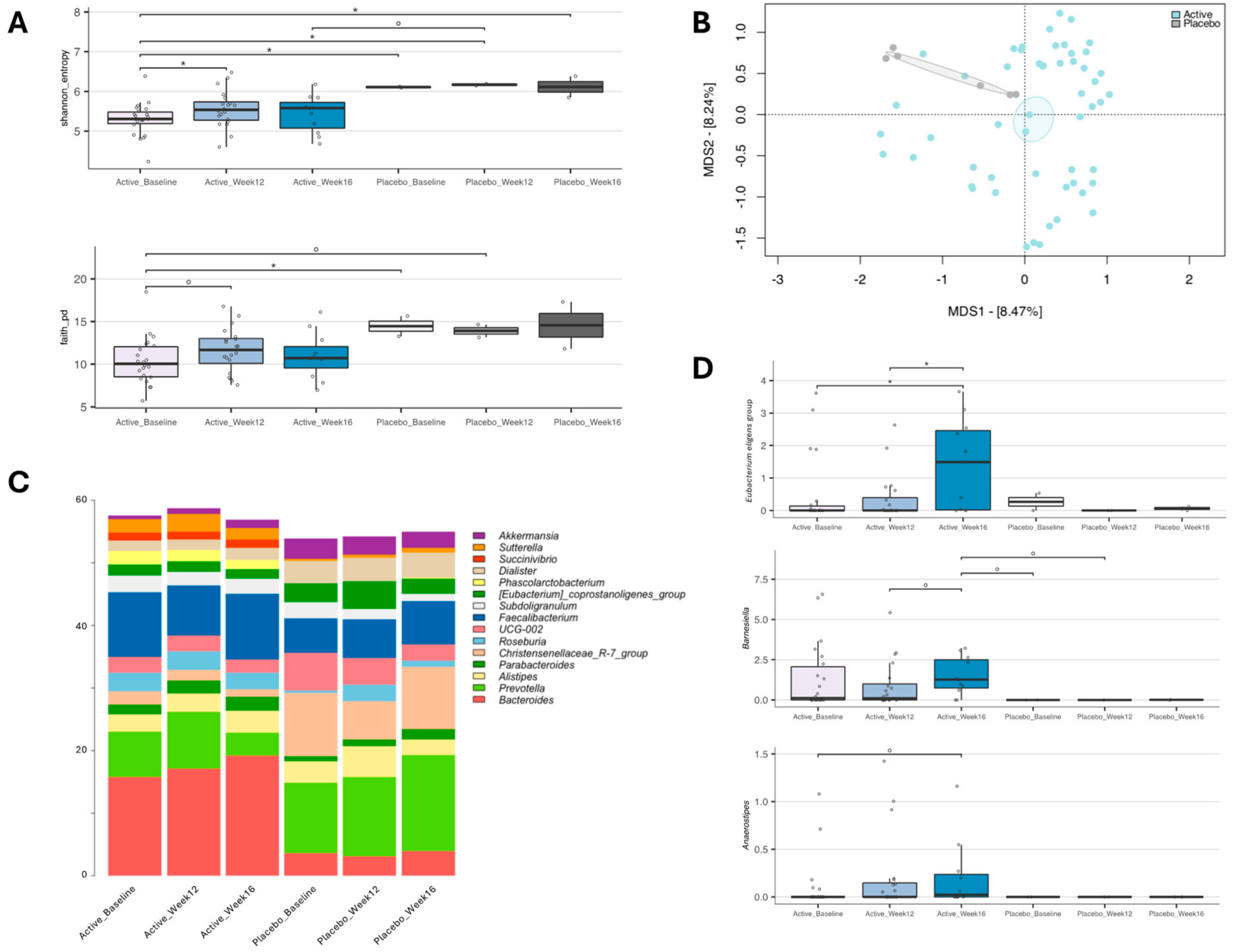
3.2.4. Impact of YMETA Supplementation on the Gut Microbiome
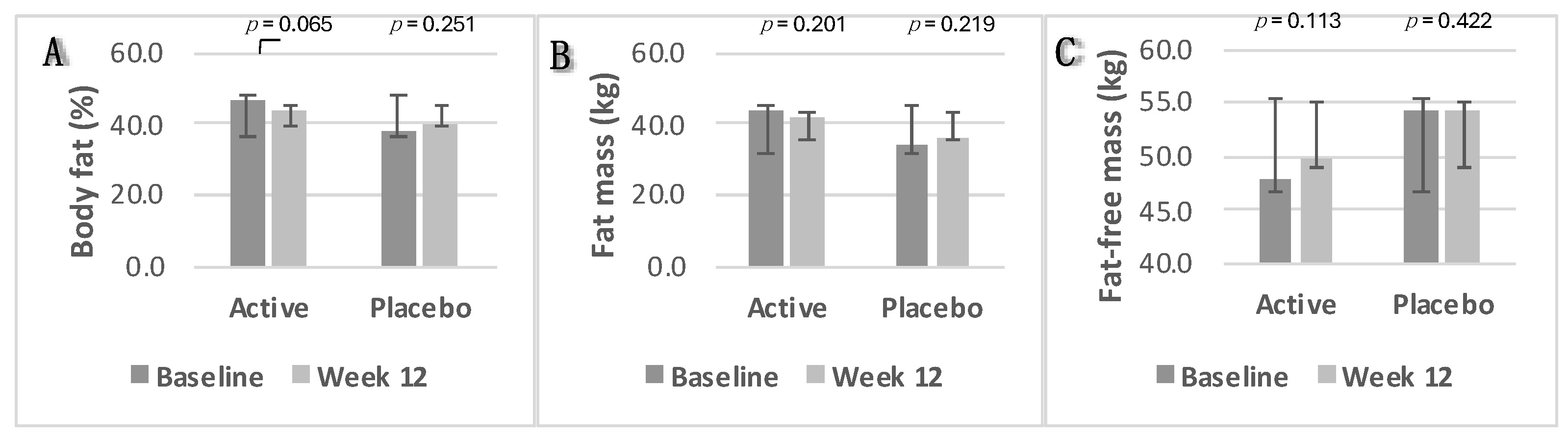
3.2.5. Anthropometrics and Body Composition Analysis
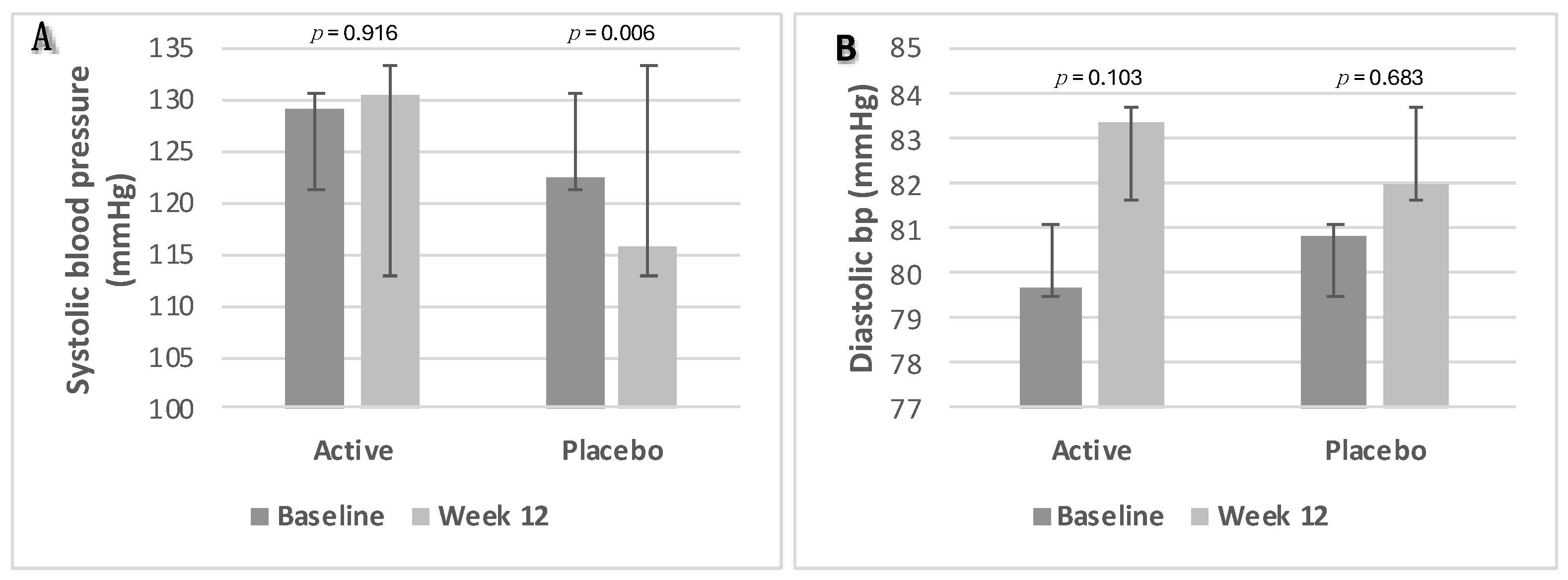
3.2.6. Blood Pressure
3.2.7. Dietary Intake and Analyses
3.2.8. Gastrointestinal (GI) Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rooney, M.R.; Wang, D.; Ogurtsova, K.; Ozkan, B.; Echouffo, J.B.; Boyko, E.J.; Magliano, D.J.; Selvin, E. Global Prevalence of Prediabetes. Diabetes Care 2023, 46, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Diabetes Prevention Program Research Group. Long-term safety, tolerability, and weight loss associated with metformin in the Diabetes Prevention Program Outcomes Study. Diabetes Care 2012, 35, 731–737. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Echouffo-Tcheugui, J.B.; Selvin, E. Prediabetes and What It Means: The Epidemiological Evidence. Annu. Rev. Public Health 2021, 42, 59–77. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The Top 10 Causes of Death. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 28 May 2024).
- Andrade, E.F.; Lobato, R.V.; de Araújo, T.V.; Zangerônimo, M.G.; de Sousa, R.V.; Pereira, L.J. Effect of beta-glucans in the control of blood glucose levels of diabetic patients: A systematic review. Nutr. Hosp. 2015, 31, 170–177. [Google Scholar]
- Wang, G.; Wang, H.; Jin, Y.; Xiao, Z.; Umar Yaqoob, M.; Lin, Y.; Chen, H.; Wang, M. Galactooligosaccharides as a protective agent for intestinal barrier and its regulatory functions for intestinal microbiota. Food Res. Int. 2020, 155, 111003. [Google Scholar] [CrossRef]
- Megur, A.; Daliri, E.B.-M.; Baltriukienė, D.; Burokas, A. Prebiotics as a Tool for the Prevention and Treatment of Obesity and Diabetes: Classification and Ability to Modulate the Gut Microbiota. Int. J. Mol. Sci. 2022, 23, 6097. [Google Scholar] [CrossRef] [PubMed]
- Kassaian, N.; Feizi, A.; Rostami, S.; Aminorroaya, A.; Yaran, M.; Amini, M. The effects of 6 mo of supplementation with probiotics and synbiotics on gut microbiota in the adults with prediabetes: A double blind randomized clinical trial. Nutrition 2020, 79–80, 110854. [Google Scholar]
- Wang, X.; Yang, J.; Qiu, X.; Wen, Q.; Liu, M.; Zhou, D.; Chen, Q. Probiotics, Pre-biotics and Synbiotics in the Treatment of Pre-diabetes: A Systematic Review of Randomized Controlled Trials. Front Public Health. 2021, 26, 645035. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Costabile, A.; Buttarazzi, I.; Kolida, S.; Quercia, S.; Baldini, J.; Swann, J.R.; Brigidi, P.; Gibson, G.R. An in vivo assessment of the cholesterol-lowering efficacy of Lactobacillus plantarum ECGC 13110402 in normal to mildly hypercholesterolaemic adults. PLoS ONE 2017, 12, e0187964. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Corona, G.; Sarnsamak, K.; Atar-Zwillenberg, D.; Yit, C.; King, A.J.; Vauzour, D.; Barone, M.; Turroni, S.; Brigidi, P.; et al. Wholegrain fermentation affects gut microbiota composition, phenolic acid metabolism and pancreatic beta cell function in a rodent model of type 2 diabetes. Front. Microbiol. 2022, 13, 1004679. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- IBM. SPSS Statistics, version 28, Computer Program; IBM: Chicago, IL, USA, 2021. [Google Scholar]
- Culhane, A.C.; Thioulouse, J.; Perrière, G.; Higgins, D.G. MADE4: An R package for multivariate analysis of gene expression data. Bioinformatics 2021, 21, 2789–2790. [Google Scholar] [CrossRef] [PubMed]
- Public Health England. Government Dietary Recommendations Government Recommendations for Energy and Nutrients for Males and Females Aged 1–18 Years and 19+ Years; GOV.UK Public Health England: London, UK, 2016. [Google Scholar]
- Mao, T.; Huang, F.; Zhu, X.; Wei, D.; Chen, L. Effects of dietary fiber on glycemic control and insulin sensitivity in patients with type 2 diabetes: A systematic review and meta-analysis. J. Funct. Foods 2021, 82, 104500. [Google Scholar] [CrossRef]
- Ali, M.K.; Bullard, K.M.; Saydah, S.; Imperatore, G.; Gregg, E.W. Cardiovascular and renal burdens of prediabetes in the USA: Analysis of data from serial cross-sectional surveys, 1988–2014. Lancet Diabetes Endocrinol. 2018, 6, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Cai, X.; Mai, W.; Li, M.; Hu, Y. Association between prediabetes and risk of cardiovascular disease and all cause mortality: Systematic review and meta-analysis. BMJ 2021, 355, i5953. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Kondal, D.; Rubinstein, A.; Irazola, V.; Gutierrez, L.; Miranda, J.J.; Bernabé-Ortiz, A.; Lazo-Porras, M.; Levitt, N.; Steyn, K.; et al. A Multiethnic Study of Pre-Diabetes and Diabetes in LMIC. Glob. Heart 2016, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on the substantiation of health claims related to konjac mannan (glucomannan) and reduction of body weight (ID 854, 1556, 3725), reduction of post-prandial glycaemic responses (ID 1559), maintenance of normal blood glucose concentration. EFSA J. 2010, 8, 1798. [Google Scholar] [CrossRef]
- Monteiro, C.R.M.; Ávila, P.F.; Pereira, M.A.F.; Pereira, G.N.; Bordignon, S.E.; Zanella, E.; Stambuk, B.U.; de Oliveira, D.; Goldbeck, R.; Poletto, P. Hydrothermal treatment on depolymerization of hemicellulose of mango seed shell for the production of xylooligosaccharides. Carbohydr. Polym. 2021, 253, 117274. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Yuan, J.; Li, D. Biological activity of galacto-oligosaccharides: A review. Front. Microbiol. 2023, 13, 993052. [Google Scholar] [CrossRef] [PubMed]
- Kuddus, M.; Aguilar, C.N. Value-Addition in Food Products and Processing through Enzyme Technology; Academic Press: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Peláez, C.; et al. Safety of the extension of use of galacto-oligosaccharides (GOS) as a novel food in food for special medical purposes pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20, e07203. [Google Scholar] [PubMed]
- Ambrogi, V.; Bottacini, F.; Cao, L.; Kuipers, B.; Schoterman, M.; van Sinderen, D. Galacto-oligosaccharides as infant prebiotics: Production, application, bioactive activities and future perspectives. Crit. Rev. Food Sci. Nutr. 2023, 63, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.; Ćorović, M.; Milivojević, A.; Simović, M.; Banjanac, K.; Pjanović, R.; Bezbradica, D. Prebiotic effect of galacto-oligosaccharides on the skin microbiota and determination of their diffusion properties. Int. J. Cosmet. Sci. 2022, 44, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.E.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of Diet With Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology 2017, 153, 87–97. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Tzortzis, G.; Gibson, G.R. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 2013, 143, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Tester, R.; Al-Ghazzewi, F. Glucomannans and nutrition. Food Hydrocoll. 2017, 68, 246–254. [Google Scholar] [CrossRef]
- Fang, Y.; Ma, J.; Lei, P.; Wang, L.; Qu, J.; Zhao, J.; Liu, F.; Yan, X.; Wu, W.; Jin, L.; et al. Konjac Glucomannan: An Emerging Specialty Medical Food to Aid in the Treatment of Type 2 Diabetes Mellitus. Foods 2023, 12, 363. [Google Scholar] [CrossRef] [PubMed]
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition, and Allergies). Scientific Opinion on Dietary Reference Values for carbohydrates and dietary fibre. EFSA J. 2010, 8, 1462. [Google Scholar] [CrossRef]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and their molecular communication with the immune system. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef]
- Hughes, K.R.; Harnisch, L.C.; Alcon-Giner, C.; Mitra, S.; Wright, C.J.; Ketskemety, J.; van Sinderen, D.; Watson, A.J.M.; Hall, L.J. Bifidobacterium breve reduces apoptotic epi- thelial cell shedding in an exopolysaccharide and MyD88-dependent manner. Open Biol. 2017, 7, 160155. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; Gustafsson, J.K.; McDonald, K.G.; Kulkarni, D.H.; Coughlin, P.E.; McCrate, S.; Kim, D.; Hsieh, C.-S.; Hogan, S.P.; Elson, C.O.; et al. Microbial antigen encounter during a preweaning interval is critical for tolerance to gut bacteria. Sci. Immunol. 2017, 2, eaao1314. [Google Scholar] [CrossRef] [PubMed]
- Motei, D.M.; Beteri, B.; Hepsomali, P.; Tzortzis, G.; Vulevic, J.; Costabile, A. Supplementation with postbiotic from Bifidobacterium Breve BB091109 improves inflammatory status and endocrine function in healthy females: A randomized, double-blind, placebo-controlled, parallel-groups study. Front. Microbiol. 2023, 14, 1273861. [Google Scholar] [CrossRef] [PubMed]
- Roney, J.R.; Simmons, Z.L. Elevated psychological stress predicts reduced estradiol concentrations in young women. Adapt. Hum. Behav. Physiol. 2015, 1, 30–40. [Google Scholar] [CrossRef]
- Samaan, R.A. Dietary Fiber for the Prevention of Cardiovascular Disease: Fiber’s Interaction between Gut Microflora, Sugar Metabolism, Weight Control and Cardiovascular Health; Elsevier: London, UK; Academic Press: London, UK, 2017. [Google Scholar]
- Arena, M.P.; Spano, G.; Fiocco, D. β-Glucans and Probiotics. Amer. J. Immunol. 2017, 13, 34–44. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Veronesi, M.; Strocchi, E.; Grandi, E.; Rizzoli, E.; Poli, A.; Marangoni, F.; Borghi, C. A randomized Placebo-Controlled Clinical Trial to Evaluate the Medium-Term Effects of Oat Fibers on Human Health: The Beta-Glucan Effects on Lipid Profile, Glycemia and inTestinal Health (BELT) Study. Nutrients 2020, 12, 686. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Cheung, P.C. Fermentation of β-glucans derived from different sources by bifidobacteria: Evaluation of their bifidogenic effect. J. Agric. Food Chem. 2011, 59, 5986–5992. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Gordon, S. Immune recognition of fungal beta-glucans. Cell Microbiol. 2005, 7, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by beta-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Sun, M.; Wang, T.; Zhang, S.; Song, X.; Liu, X.; Wei, J.; Chen, Q.; Zhong, T.; Qin, J. Gut microbiota and type 1 diabetes: A two-sample bidirectional Mendelian randomization study. Front. Cell. Infect. Microbiol. 2023, 13, 1163898. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Khatib, H.A.; et al. Microbiome connections with host metabolism and habitual diet from 1098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microb. 2020, 12, 1802866. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhang, Y.; Ma, D.; Zhang, W.; Zhang, H. Lactobacillus casei Zhang exerts anti-obesity effect to obese glut1 and gut-specific-glut1 knockout mice via gut microbiota modulation mediated different metagenomic pathways. Eur. J. Nutr. 2022, 61, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al. Longitudinal multi-omics of host-microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, Y.E.; Esquivel-Hernández, D.A.; Sánchez-Castañeda, J.P.; Neri-Rosario, D.; Guardado-Mendoza, R.; Resendis-Antonio, O. Type 2 diabetes, gut microbiome, and systems biology: A novel perspective for a new era. Gut Microb. 2022, 14, 2111952. [Google Scholar] [CrossRef] [PubMed]
- Fongang, B.; Satizabal, C.; Kautz, T.F.; Wadop, Y.N.; Muhammad, J.A.S.; Vasquez, E.; Mathews, J.; Gireud-Goss, M.; Saklad, A.R.; Himali, J.; et al. Cerebral small vessel disease burden is associated with decreased abundance of gut Barnesiella intestinihominis bacterium in the Framingham Heart Study. Sci. Rep. 2023, 13, 13622. [Google Scholar] [CrossRef] [PubMed]
- Doumatey, A.P.; Adeyemo, A.; Zhou, J.; Lei, L.; Adebamowo, S.N.; Adebamowo, C.; Rotimi, C.N. Gut Microbiome Profiles Are Associated With Type 2 Diabetes in Urban Africans. Front. Cell. Infect. Microbiol. 2020, 10, 63. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Godneva, A.; Bar, N.; Kurilshikov, A.; Lotan-Pompan, M.; Weinberger, A.; Fu, J.; Wijmenga, C.; Zhernakova, A.; et al. Structural variation in the gut microbiome associates with host health. Nature 2019, 568, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.H.; Meyer, K.; Lulla, A.; Lewis, C.E.; Carnethon, M.R.; Schreiner, P.J.; Sidney, S.; Shikany, J.M.; Meirelles, O.; Launer, L.J. Gut microbiome and stages of diabetes in middle-aged adults: CARDIA microbiome study. Nutr. Metab. 2023, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kang, W.; Shin, N.R.; Hyun, D.W.; Kim, P.S.; Kim, H.S.; Lee, J.Y.; Tak, E.J.; Sung, H.; Bae, J.W. Anaerostipes hominis sp. nov., a novel butyrate-producing bacteria isolated from faeces of a patient with Crohn’s disease. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, R.; Tanno, H.; Maeno, S.; Endo, A. Spore-forming properties and enhanced oxygen tolerance of butyrate-producing Anaerostipes spp. Anaerobe 2023, 82, 102752. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptives | Intervention (n = 25) | Placebo (n = 25) | p-Value |
|---|---|---|---|
| Age (years) | 53.5 ± 8.8 | 51.0 ± 10.9 | 0.369 |
| Sex (M/F), n/n | 7/18 | 16/9 | |
| Anthropometric Measurements | |||
| Height (cm) | 167.6 ± 8.8 | 172.1 ± 10.5 | 0.095 |
| Weight (kg) | 91.9 ± 20.8 | 87.5 ± 17.5 | 0.414 |
| Body Mass Index (kg/m2) | 32.4 ± 6.3 | 29.0 ± 4.0 | 0.019 |
| Waist Circumference (cm) | 100.2 ± 13.7 | 106.5 ± 30.2 | 0.342 |
| Hip Circumference (cm) | 114.1 ± 13.4 | 101.7 ± 21.4 | 0.015 |
| Waist/Hip Ratio (waist/hip circumference) | 0.88 ± 0.07 | 1.24 ± 1.03 | 0.085 |
| Body Composition | |||
| Body Fat (%) | 46.4 ± 9.4 | 38.3 ± 7.1 | <0.001 |
| Fat Mass (kg) | 43.8 ± 17.8 | 33.7 ± 9.9 | 0.013 |
| Fat Free Mass (kg) | 47.8 ± 8.4 | 53.9 ± 12.2 | 0.043 |
| Blood Pressure | |||
| Systolic Blood Pressure (mmHg) | 130.5 ± 12.7 | 122.7 ± 7.9 | 0.009 |
| Diastolic Blood Pressure (mmHg) | 80.2 ± 8.4 | 80.8 ± 10.2 | 0.814 |
| Variable LBP, μg/mL | Baseline | Post-Intervention | Treatment | Time | Interaction |
|---|---|---|---|---|---|
| Intervention | 14.8 ± 0.3 | 12.4 ± 0.9 | 0.732 | 0.002 * | 0.207 |
| Control | 18.7 ± 0.6 | 15.4 ± 1.1 | 0.6 |
| Descriptives | Intervention (n = 20) | Placebo (n = 25) |
|---|---|---|
| Total Daily Energy Intake (kcals) | 1769.9 ± 525.8 | 2566.7 ± 964.3 |
| Protein (g) (% energy) | 78.3 ± 29.9 (18.1 ± 5.9) | 93.5 ± 33.2 (15.0 ± 1.5) |
| Carbohydrate (g) (% energy) | 194.4 ± 54.2 (43.5 ± 8.5) | 303.4 ± 103.3 (43.1 ± 2.7) |
| Dietary Fibre (g) | 20.5 ± 7.4 | 27.1 ± 9.0 |
| Soluble Fibre (g) | ||
| Starch (g) | 109.4 ± 54.2 | 207.3 ± 77.9 |
| Sugars (g) | 81.5 ± 27.5 | 94.8 ± 37.3 |
| Insoluble Fibre (g) | ||
| Non-Starch Polysaccharides (NSPs) (g) | 15.7 ± 3.9 | 19.5 ± 6.8 |
| Fat (g) (% energy) | 72.8 ± 25.8 (35.6 ± 5.4) | 128.9 ± 63.7 (39.7 ± 19.6) |
| Alcohol (% energy) | 7.2 ± 10.1 (2.7 ± 4.0) | 8.2 ± 9.3 (2.8 ± 4.9) |
| Variable | Baseline | Post-Intervention | Treatment | Time | Interaction |
|---|---|---|---|---|---|
| Abdominal discomfort | |||||
| Intervention | 1.18 ± 0.36 | 1.09 ± 0.34 | 0.938 | 0.0091 | 0.439 |
| Control | 1.67 ± 0.53 | 1.78 ± 0.56 | |||
| Heartburn | |||||
| Intervention | 1.27 ± 0.38 | 1.00 ± 0.31 | 0.067 | 0.583 | 0.884 |
| Control | 1.33 ± 0.42 | 1.11 ± 0.35 | |||
| Acid reflux | |||||
| Intervention | 1.09 ± 0.33 | 1.18 ± 0.374 | 0.810 | 0.224 | 0.380 |
| Control | 1.00 ± 0.31 | 1.01 ± 0.34 | |||
| Hunger pains | |||||
| Intervention | 1.2 ± 0.3 | 1.1 ± 0.2 | 0.817 | 0.068 | 0.842 |
| Control | 1.2 ± 0.2 | 1.3 ± 0.2 | |||
| Nausea | |||||
| Intervention | 1.00 ± 0.30 | 1.55 ± 0.49 | 0.988 | 0.180 | 0.475 |
| Control | 2.33 ± 0.73 | 2.11 ± 0.66 | |||
| Rumbling | |||||
| Intervention | 1.64 ± 0.49 | 1.55 ± 0.49 | 0.558 | 0.057 | 0.805 |
| Control | 2.33 ± 0.74 | 2.11 ± 0.67 | |||
| Bloating | |||||
| Intervention | 2.45 ± 0.74 | 2.00 ± 0.63 | 0.152 | 0.693 | 0.614 |
| Control | 2.11 ± 0.66 | 1.90 ± 0.59 | |||
| Burping | |||||
| Intervention | 1.18 ± 0.36 | 1.45 ± 0.46 | 0.152 | 0.614 | 0.263 |
| Control | 1.78 ± 0.56 | 1.33 ± 0.42 | |||
| Flatus | |||||
| Intervention | 1.27 ± 0.38 | 1.55 ± 0.49 | 0.815 | 0.334 | 0.988 |
| Control | 2.33 ± 0.73 | 1.89 ± 0.59 | |||
| Constipation | |||||
| Intervention | 1.28 ± 0.42 | 1.27 ± 0.40 | 0.161 | 0.161 | 0.879 |
| Control | 1.56 ± 0.49 | 1.11 ± 0.35 | |||
| Diarrheal | |||||
| Intervention | 1.18 ± 0.36 | 1.45 ± 0.46 | 0.747 | 0.187 | 0.555 |
| Control | 1.78 ± 0.56 | 1.33 ± 0.42 | |||
| Loose stools | |||||
| Intervention | 1.36 ± 0.41 | 1.82 ± 0.57 | 0.569 | 0.569 | 0.336 |
| Control | 1.67 ± 0.52 | 1.72 ± 0.64 | |||
| Hard stools | |||||
| Intervention | 1.45 ± 0.44 | 1.09 ± 0.34 | 0.448 | 0.162 | 0.129 |
| Control | 1.00 ± 0.31 | 1.13 ± 0.35 | |||
| Bowel movement | |||||
| Intervention | 1.73 ± 0.52 | 2.00 ± 0.63 | 0.387 | 0.929 | 0.750 |
| Control | 2.00 ± 0.63 | 2.38 ± 0.75 | |||
| Not complete bowel emptiness sensation | |||||
| Intervention | 2.09 ± 0.63 | 1.91 ± 0.60 | 0.103 | 0.481 | 0.518 |
| Control | 1.78 ± 0.56 | 1.38 ± 0.43 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beteri, B.; Barone, M.; Turroni, S.; Brigidi, P.; Tzortzis, G.; Vulevic, J.; Sekulic, K.; Motei, D.-E.; Costabile, A. Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients 2024, 16, 2205. https://doi.org/10.3390/nu16142205
Beteri B, Barone M, Turroni S, Brigidi P, Tzortzis G, Vulevic J, Sekulic K, Motei D-E, Costabile A. Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients. 2024; 16(14):2205. https://doi.org/10.3390/nu16142205
Chicago/Turabian StyleBeteri, Beyda, Monica Barone, Silvia Turroni, Patrizia Brigidi, George Tzortzis, Jelena Vulevic, Karol Sekulic, Diana-Elena Motei, and Adele Costabile. 2024. "Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study" Nutrients 16, no. 14: 2205. https://doi.org/10.3390/nu16142205
APA StyleBeteri, B., Barone, M., Turroni, S., Brigidi, P., Tzortzis, G., Vulevic, J., Sekulic, K., Motei, D.-E., & Costabile, A. (2024). Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients, 16(14), 2205. https://doi.org/10.3390/nu16142205