
Impact of High-Fat Diet and Exercise on Bone and Bile Acid Metabolism in Rats

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diet and Exercise
2.3. Microcomputed Tomography (microCT)
2.4. Biomechanical Analysis
2.5. Extraction of Bile Acids
2.6. Mass Spectrometry Analyses
2.7. Statistics
3. Results
3.1. Exercise Cannot Revert Long-Term HFD-Related Bone Loss
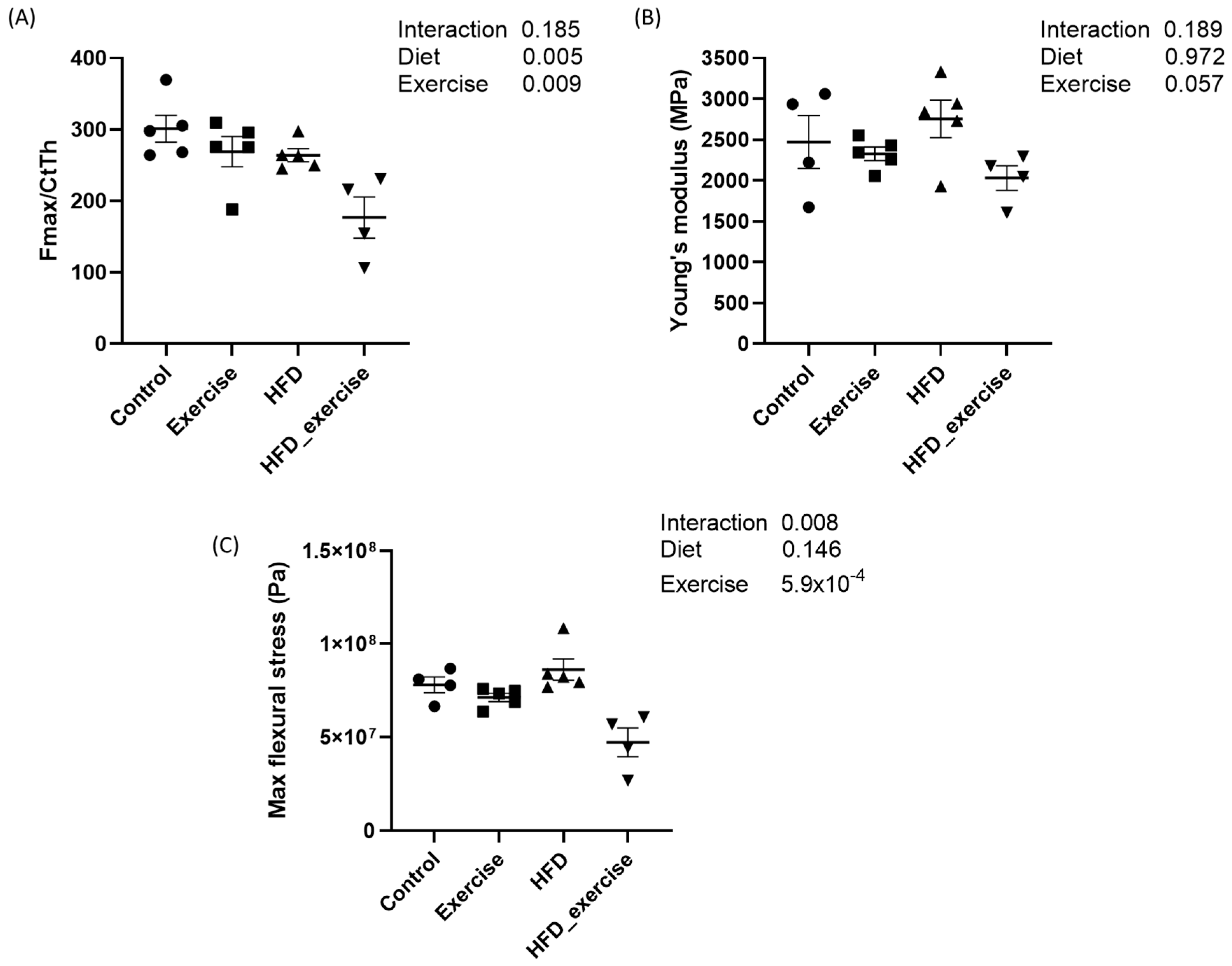
3.2. Long-Term HFD and Exercise Have Effect on Bone Mechanical Properties
3.3. HFD-Related Alterations of Serum Bile Acid Profile Cannot Be Reverted by Exercise
3.4. Long-Term HFD Dysregulates Bile Acid Levels in Stool That Cannot Be Reverted by Exercise
3.5. Long-Term HFD Affects the Bile Acid Ratios That Cannot Be Reverted by Exercise
3.6. Bile Acid Alterations Correlate with Bone Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Health Organisation. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 11 May 2024).
- Halade, G.V.; Rahman, M.M.; Williams, P.J.; Fernandes, G. High fat diet-induced animal model of age-associated obesity and osteoporosis. J. Nutr. Biochem. 2010, 21, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.; Gupta, S.R.; Moustafa, A.F.; Chao, A.M. Sex/Gender Differences in Obesity Prevalence, Comorbidities, and Treatment. Curr. Obes. Rep. 2021, 10, 458–466. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Reid, I.R.; Ames, R.; Evans, M.C.; Sharpe, S.; Gamble, G.; France, J.T.; Lim, T.M.; Cundy, T.F. Determinants of total body and regional bone mineral density in normal postmenopausal women--a key role for fat mass. J. Clin. Endocrinol. Metab. 1992, 75, 45–51. [Google Scholar] [PubMed]
- Felson, D.T.; Zhang, Y.; Hannan, M.T.; Anderson, J.J. Effects of weight and body mass index on bone mineral density in men and women: The Framingham study. J. Bone Miner. Res. 1993, 8, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Ravn, P.; Cizza, G.; Bjarnason, N.H.; Thompson, D.; Daley, M.; Wasnich, R.D.; McClung, M.; Hosking, D.; Yates, A.J.; Christiansen, C. Low body mass index is an important risk factor for low bone mass and increased bone loss in early postmenopausal women. Early Postmenopausal Intervention Cohort (EPIC) study group. J. Bone Miner. Res. 1999, 14, 1622–1627. [Google Scholar] [CrossRef] [PubMed]
- Compston, J. Type 2 diabetes mellitus and bone. J. Intern. Med. 2018, 283, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Bonds, D.E.; Larson, J.C.; Schwartz, A.V.; Strotmeyer, E.S.; Robbins, J.; Rodriguez, B.L.; Johnson, K.C.; Margolis, K.L. Risk of fracture in women with type 2 diabetes: The Women’s Health Initiative Observational Study. J. Clin. Endocrinol. Metab. 2006, 91, 3404–3410. [Google Scholar] [CrossRef] [PubMed]
- Starup-Linde, J.; Frost, M.; Vestergaard, P.; Abrahamsen, B. Epidemiology of Fractures in Diabetes. Calcif. Tissue Int. 2017, 100, 109–121. [Google Scholar] [CrossRef]
- Vestergaard, P. Discrepancies in bone mineral density and fracture risk in patients with type 1 and type 2 diabetes—A meta-analysis. Osteoporos. Int. 2007, 18, 427–444. [Google Scholar] [CrossRef]
- Yu, E.W.; Putman, M.S.; Derrico, N.; Abrishamanian-Garcia, G.; Finkelstein, J.S.; Bouxsein, M.L. Defects in cortical microarchitecture among African-American women with type 2 diabetes. Osteoporos. Int. 2015, 26, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Insull, W., Jr. Clinical utility of bile acid sequestrants in the treatment of dyslipidemia: A scientific review. South Med. J. 2006, 99, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [PubMed]
- Houten, S.M.; Watanabe, M.; Auwerx, J. Endocrine functions of bile acids. EMBO J. 2006, 25, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Giannini, C.; Mastromauro, C.; Scapaticci, S.; Gentile, C.; Chiarelli, F. Role of bile acids in overweight and obese children and adolescents. Front. Endocrinol 2022, 13, 1011994. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Song, Y.W.; Zhang, L.; Zheng, F.J.; Wang, X.M.; Zhuang, X.H.; Wu, F.; Liu, J. Association between bile acid metabolism and bone mineral density in postmenopausal women. Clinics 2020, 75, e1486. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Y.; Luo, Q. The Association of Serum Total Bile Acids with Bone Mineral Density in Chinese Adults Aged 20–59: A Retrospective Cross-Sectional Study. Front. Endocrinol. 2022, 13, 817437. [Google Scholar] [CrossRef]
- Deng, D.; Pan, C.; Wu, Z.; Sun, Y.; Liu, C.; Xiang, H.; Yin, P.; Shang, D. An Integrated Metabolomic Study of Osteoporosis: Discovery and Quantification of Hyocholic Acids as Candidate Markers. Front. Pharmacol. 2021, 12, 725341. [Google Scholar] [CrossRef]
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Bile Acid Receptors and Gastrointestinal Functions. Liver Res. 2019, 3, 31–39. [Google Scholar] [CrossRef]
- Cho, S.W.; An, J.H.; Park, H.; Yang, J.Y.; Choi, H.J.; Kim, S.W.; Park, Y.J.; Kim, S.Y.; Yim, M.; Baek, W.Y.; et al. Positive regulation of osteogenesis by bile acid through FXR. J. Bone Miner. Res. 2013, 28, 2109–2121. [Google Scholar] [CrossRef]
- Zheng, T.; Kang, J.H.; Sim, J.S.; Kim, J.W.; Koh, J.T.; Shin, C.S.; Lim, H.; Yim, M. The farnesoid X receptor negatively regulates osteoclastogenesis in bone remodeling and pathological bone loss. Oncotarget 2017, 8, 76558–76573. [Google Scholar] [CrossRef]
- Li, Z.; Huang, J.; Wang, F.; Li, W.; Wu, X.; Zhao, C.; Zhao, J.; Wei, H.; Wu, Z.; Qian, M.; et al. Dual Targeting of Bile Acid Receptor-1 (TGR5) and Farnesoid X Receptor (FXR) Prevents Estrogen-Dependent Bone Loss in Mice. J. Bone Miner. Res. 2019, 34, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Monte, M.J.; Marin, J.J.; Antelo, A.; Vazquez-Tato, J. Bile acids: Chemistry, physiology, and pathophysiology. World J. Gastroenterol. 2009, 15, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Dawson, P.A.; Lan, T.; Rao, A. Bile acid transporters. J. Lipid Res. 2009, 50, 2340–2357. [Google Scholar] [CrossRef]
- Ocvirk, S.; O’Keefe, S.J.D. Dietary fat, bile acid metabolism and colorectal cancer. Semin. Cancer Biol. 2021, 73, 347–355. [Google Scholar] [CrossRef]
- Yoshitsugu, R.; Kikuchi, K.; Iwaya, H.; Fujii, N.; Hori, S.; Lee, D.G.; Ishizuka, S. Alteration of Bile Acid Metabolism by a High-Fat Diet Is Associated with Plasma Transaminase Activities and Glucose Intolerance in Rats. J. Nutr. Sci. Vitaminol. 2019, 65, 45–51. [Google Scholar] [CrossRef]
- Ruiz-Gaspa, S.; Guanabens, N.; Jurado, S.; Combalia, A.; Peris, P.; Monegal, A.; Pares, A. Bilirubin and bile acids in osteocytes and bone tissue. Potential role in the cholestatic-induced osteoporosis. Liver Int. 2020, 40, 2767–2775. [Google Scholar] [CrossRef]
- Ruiz-Gaspa, S.; Dubreuil, M.; Guanabens, N.; Combalia, A.; Peris, P.; Monegal, A.; Pares, A. Ursodeoxycholic acid decreases bilirubin-induced osteoblast apoptosis. Eur. J. Clin. Investig. 2014, 44, 1206–1214. [Google Scholar] [CrossRef]
- Chin, S.H.; Kahathuduwa, C.N.; Binks, M. Physical activity and obesity: What we know and what we need to know. Obes. Rev. 2016, 17, 1226–1244. [Google Scholar] [CrossRef] [PubMed]
- Bonaiuti, D.; Shea, B.; Iovine, R.; Negrini, S.; Robinson, V.; Kemper, H.C.; Wells, G.; Tugwell, P.; Cranney, A. Exercise for preventing and treating osteoporosis in postmenopausal women. Cochrane Database Syst. Rev. 2002, 3, CD000333. [Google Scholar]
- Kohrt, W.M.; Bloomfield, S.A.; Little, K.D.; Nelson, M.E.; Yingling, V.R.; American College of Sports Medicine. American College of Sports Medicine Position Stand: Physical activity and bone health. Med. Sci. Sports Exerc. 2004, 36, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.B.; Oliveira, J.; Bauman, A.; Fairhall, N.; Kwok, W.; Sherrington, C. Evidence on physical activity and osteoporosis prevention for people aged 65+ years: A systematic review to inform the WHO guidelines on physical activity and sedentary behaviour. Int. J. Behav. Nutr. Phys. Act. 2020, 17, 150. [Google Scholar] [CrossRef]
- Danese, E.; Salvagno, G.L.; Tarperi, C.; Negrini, D.; Montagnana, M.; Festa, L.; Sanchis-Gomar, F.; Schena, F.; Lippi, G. Middle-distance running acutely influences the concentration and composition of serum bile acids: Potential implications for cancer risk? Oncotarget 2017, 8, 52775–52782. [Google Scholar] [CrossRef] [PubMed]
- Morville, T.; Sahl, R.E.; Trammell, S.A.; Svenningsen, J.S.; Gillum, M.P.; Helge, J.W.; Clemmensen, C. Divergent effects of resistance and endurance exercise on plasma bile acids, FGF19, and FGF21 in humans. JCI Insight 2018, 3, e122737. [Google Scholar] [CrossRef] [PubMed]
- Maurer, A.; Ward, J.L.; Dean, K.; Billinger, S.A.; Lin, H.; Mercer, K.E.; Adams, S.H.; Thyfault, J.P. Divergence in aerobic capacity impacts bile acid metabolism in young women. J. Appl. Physiol. 2020, 129, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, M.D.; Almer, G.; Kaiser, M.; Zelzer, S.; Meinitzer, A.; Scharnagl, H.; Sedej, S.; Gruber, H.J.; Herrmann, M. The effects of long-term moderate exercise and Western-type diet on oxidative/nitrosative stress, serum lipids and cytokines in female Sprague Dawley rats. Eur. J. Nutr. 2022, 61, 255–268. [Google Scholar] [CrossRef]
- Prodinger, P.M.; Burklein, D.; Foehr, P.; Kreutzer, K.; Pilge, H.; Schmitt, A.; Eisenhart-Rothe, R.V.; Burgkart, R.; Bissinger, O.; Tischer, T. Improving results in rat fracture models: Enhancing the efficacy of biomechanical testing by a modification of the experimental setup. BMC Musculoskelet. Disord. 2018, 19, 243. [Google Scholar] [CrossRef]
- Singh, A.; Scholze, M.; Hammer, N. On the influence of surface coating on tissue biomechanics—Effects on rat bones under routine conditions with implications for image-based deformation detection. BMC Musculoskelet. Disord. 2018, 19, 387. [Google Scholar] [CrossRef]
- Amplatz, B.; Zohrer, E.; Haas, C.; Schaffer, M.; Stojakovic, T.; Jahnel, J.; Fauler, G. Bile acid preparation and comprehensive analysis by high performance liquid chromatography-high-resolution mass spectrometry. Clin. Chim. Acta 2017, 464, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Humbert, L.; Maubert, M.A.; Wolf, C.; Duboc, H.; Mahe, M.; Farabos, D.; Seksik, P.; Mallet, J.M.; Trugnan, G.; Masliah, J.; et al. Bile acid profiling in human biological samples: Comparison of extraction procedures and application to normal and cholestatic patients. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 899, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Atkinson, E.J.; Riggs, B.L.; Melton, L.J., 3rd. Relationship between body composition and bone mass in women. J. Bone Miner. Res. 1996, 11, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, G.; Kang, J.; Hou, Y.; Jiang, F.; Yuan, W.; Shi, J. Obesity and risk of hip fracture in adults: A meta-analysis of prospective cohort studies. PLoS ONE 2013, 8, e55077. [Google Scholar] [CrossRef] [PubMed]
- Nielson, C.M.; Srikanth, P.; Orwoll, E.S. Obesity and fracture in men and women: An epidemiologic perspective. J. Bone Miner. Res. 2012, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.E.; Watts, N.B.; Chapurlat, R.; Cooper, C.; Boonen, S.; Greenspan, S.; Pfeilschifter, J.; Silverman, S.; Diez-Perez, A.; Lindsay, R.; et al. Obesity is not protective against fracture in postmenopausal women: GLOW. Am. J. Med. 2011, 124, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Picke, A.K.; Sylow, L.; Moller, L.L.V.; Kjobsted, R.; Schmidt, F.N.; Steejn, M.W.; Salbach-Hirsch, J.; Hofbauer, C.; Bluher, M.; Saalbach, A.; et al. Differential effects of high-fat diet and exercise training on bone and energy metabolism. Bone 2018, 116, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Scheller, E.L.; Khoury, B.; Moller, K.L.; Wee, N.K.; Khandaker, S.; Kozloff, K.M.; Abrishami, S.H.; Zamarron, B.F.; Singer, K. Changes in Skeletal Integrity and Marrow Adiposity during High-Fat Diet and after Weight Loss. Front. Endocrinol. 2016, 7, 102. [Google Scholar] [CrossRef]
- Zernicke, R.F.; Salem, G.J.; Barnard, R.J.; Schramm, E. Long-term, high-fat-sucrose diet alters rat femoral neck and vertebral morphology, bone mineral content, and mechanical properties. Bone 1995, 16, 25–31. [Google Scholar] [CrossRef]
- Cao, J.J.; Gregoire, B.R.; Gao, H. High-fat diet decreases cancellous bone mass but has no effect on cortical bone mass in the tibia in mice. Bone 2009, 44, 1097–1104. [Google Scholar] [CrossRef]
- Silva, M.J.; Eekhoff, J.D.; Patel, T.; Kenney-Hunt, J.P.; Brodt, M.D.; Steger-May, K.; Scheller, E.L.; Cheverud, J.M. Effects of High-Fat Diet and Body Mass on Bone Morphology and Mechanical Properties in 1100 Advanced Intercross Mice. J. Bone Miner. Res. 2019, 34, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, R.M.; Garcia-Irigoyen, O.; Moschetta, A. Bile acids and colon cancer: Is FXR the solution of the conundrum? Mol. Asp. Med. 2017, 56, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, R.A.; Camastra, S.; Nannipieri, M.; Astiarraga, B.; Castro-Perez, J.; Xie, D.; Wang, L.; Chakravarthy, M.; Ferrannini, E. Increased Bile Acid Synthesis and Impaired Bile Acid Transport in Human Obesity. J. Clin. Endocrinol. Metab. 2016, 101, 1935–1944. [Google Scholar] [CrossRef]
- Wan, Y.; Yuan, J.; Li, J.; Li, H.; Zhang, J.; Tang, J.; Ni, Y.; Huang, T.; Wang, F.; Zhao, F.; et al. Unconjugated and secondary bile acid profiles in response to higher-fat, lower-carbohydrate diet and associated with related gut microbiota: A 6-month randomized controlled-feeding trial. Clin. Nutr. 2020, 39, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Eyssen, H.J.; De Pauw, G.; Van Eldere, J. Formation of hyodeoxycholic acid from muricholic acid and hyocholic acid by an unidentified gram-positive rod termed HDCA-1 isolated from rat intestinal microflora. Appl. Environ. Microbiol. 1999, 65, 3158–3163. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, T.; Jiang, R.; Zhao, A.; Wu, Q.; Kuang, J.; Sun, D.; Ren, Z.; Li, M.; Zhao, M.; et al. Hyocholic acid species improve glucose homeostasis through a distinct TGR5 and FXR signaling mechanism. Cell Metab. 2021, 33, 791–803.e7. [Google Scholar] [CrossRef] [PubMed]
- Nakade, Y.; Kitano, R.; Sakamoto, K.; Kimoto, S.; Yamauchi, T.; Inoue, T.; Kobayashi, Y.; Ohashi, T.; Sumida, Y.; Ito, K.; et al. Characteristics of bile acid composition in high fat diet-induced nonalcoholic fatty liver disease in obese diabetic rats. PLoS ONE 2021, 16, e0247303. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Fan, X.; Lu, Y.; Chen, D.; Zhao, Y.; Qi, K. Dietary acetic acid suppress high-fat diet-induced obesity in mice by altering taurine conjugated bile acids metabolism. Curr. Res. Food Sci. 2022, 5, 1976–1984. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef]
- Haeusler, R.A.; Astiarraga, B.; Camastra, S.; Accili, D.; Ferrannini, E. Human insulin resistance is associated with increased plasma levels of 12alpha-hydroxylated bile acids. Diabetes 2013, 62, 4184–4191. [Google Scholar] [CrossRef]
- Jiao, N.; Baker, S.S.; Chapa-Rodriguez, A.; Liu, W.; Nugent, C.A.; Tsompana, M.; Mastrandrea, L.; Buck, M.J.; Baker, R.D.; Genco, R.J.; et al. Suppressed hepatic bile acid signalling despite elevated production of primary and secondary bile acids in NAFLD. Gut 2018, 67, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Mangat, S.; Sheinfil, A.; Weisburger, J.H.; Wynder, E.L. Effect of type and amount of dietary fat and 1,2-dimethylhydrazine on biliary bile acids, fecal bile acids, and neutral sterols in rats. Cancer Res. 1977, 37, 2132–2137. [Google Scholar] [PubMed]
- Hori, S.; Abe, T.; Lee, D.G.; Fukiya, S.; Yokota, A.; Aso, N.; Shirouchi, B.; Sato, M.; Ishizuka, S. Association between 12alpha-hydroxylated bile acids and hepatic steatosis in rats fed a high-fat diet. J. Nutr. Biochem. 2020, 83, 108412. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, W.; Yoshida, R.; Liu, H.; Hori, S.; Otsubo, Y.; Tanaka, Y.; Sato, M.; Ishizuka, S. The ratio of 12alpha to non-12-hydroxylated bile acids reflects hepatic triacylglycerol accumulation in high-fat diet-fed C57BL/6J mice. Sci. Rep. 2022, 12, 16707. [Google Scholar] [CrossRef] [PubMed]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism of growth inhibition by free bile acids in lactobacilli and bifidobacteria. J. Bacteriol. 2006, 188, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.C.; Lee, Y.S.; Bae, I.H.; Lee, C.H.; Shin, H.I.; Ha, H.J.; Franceschi, R.T.; Choi, H.S.; Koh, J.T. The orphan nuclear receptor SHP is a positive regulator of osteoblastic bone formation. J. Bone Miner. Res. 2010, 25, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Okamoto, A.Y.; Shan, B. FXR, a bile acid receptor and biological sensor. Trends Cardiovasc. Med. 2000, 10, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, H.; Lu, Y.F.; Lei, X.; Cui, J.Y.; Ellis, E.; Strom, S.C.; Klaassen, C.D. Potency of individual bile acids to regulate bile acid synthesis and transport genes in primary human hepatocyte cultures. Toxicol. Sci. 2014, 141, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Modica, S.; Gadaleta, R.M.; Moschetta, A. Deciphering the nuclear bile acid receptor FXR paradigm. Nucl. Recept Signal 2010, 8, e005. [Google Scholar] [CrossRef]
- Nishida, S.; Ishizawa, M.; Kato, S.; Makishima, M. Vitamin D Receptor Deletion Changes Bile Acid Composition in Mice Orally Administered Chenodeoxycholic Acid. J. Nutr. Sci. Vitaminol. 2020, 66, 370–374. [Google Scholar] [CrossRef]
- Almer, G.; Semeraro, M.; Meinitzer, A.; Enko, D.; Rodriguez-Blanco, G.; Gall, B.; Horvath, A.; Moissl-Eichinger, C.; Till, H.; Gruber, H.; et al. Impact of long-term high dietary fat intake and regular exercise on serum TMAO and microbiome composition in female rats. Nutr. Healthy Ageing 2023, 8, 157–170. [Google Scholar] [CrossRef]
- Talebian, R.; Hashem, O.; Gruber, R. Taurocholic acid lowers the inflammatory response of gingival fibroblasts, epithelial cells, and macrophages. J. Oral. Sci. 2020, 62, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.; Jaeschke, H.; Copple, B.L. Bile acids induce inflammatory genes in hepatocytes: A novel mechanism of inflammation during obstructive cholestasis. Am. J. Pathol. 2011, 178, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R.; Kaser, A.; Pines, A.; Dotan, I. Gut, inflammation and osteoporosis: Basic and clinical concepts. Gut 2008, 57, 684–694. [Google Scholar] [CrossRef] [PubMed]
- McCabe, L.R.; Irwin, R.; Tekalur, A.; Evans, C.; Schepper, J.D.; Parameswaran, N.; Ciancio, M. Exercise prevents high fat diet-induced bone loss, marrow adiposity and dysbiosis in male mice. Bone 2019, 118, 20–31. [Google Scholar] [CrossRef]
- Mercer, K.E.; Maurer, A.; Pack, L.M.; Ono-Moore, K.; Spray, B.J.; Campbell, C.; Chandler, C.J.; Burnett, D.; Souza, E.; Casazza, G.; et al. Exercise training and diet-induced weight loss increase markers of hepatic bile acid (BA) synthesis and reduce serum total BA concentrations in obese women. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E864–E873. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bile Acid | Control | Exercise | HFD | HFD and Exercise | Two-Way ANOVA p | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Median [IQR] | n | Median [IQR] | n | Median [IQR] | n | Median [IQR] | HFD | Exercise | Interaction | |
| Total | 21 | 39.7 [19.4–72.8] | 23 | 41.3 [18.4–79.0] | 16 | 14.6 [9.5–20.6] | 12 | 12.9 [4.7–19.0] | 1.0 × 10−6 | 0.60 | 0.66 |
| Conjugated | 21 | 8.2 [5.1–12.3] | 23 | 5.9 [3.4–8.5] | 16 | 3.7 [1.8–7.3] | 12 | 3.1 [1.4–6.2] | 0.002 | 0.24 | 0.54 |
| Free | 21 | 33.6 [14.3–58.9] | 23 | 36.5 [13.5–51.7] | 16 | 8.4 [3.5–14.7] | 12 | 5.0 [0.7–12.1] | 2.4 × 10−7 | 0.60 | 0.45 |
| Primary | 21 | 35.5 [18.8–61.4] | 23 | 35.1 [17.5–66.4] | 16 | 11.4 [7.4–16.9] | 12 | 9.0 [3.2–15.7] | 6.4 × 10−8 | 0.44 | 0.63 |
| Secondary | 21 | 3.1 [0.8–8.7] | 23 | 3.7 [0.9–8.9] | 16 | 2.8 [1.5–3.8] | 12 | 2.5 [1.5–3.7] | 0.278 | 0.74 | 0.57 |
| 12-α-OH | 21 | 16.2 [9.4–27.0] | 23 | 12.2 [8.3–20.6] | 16 | 7.0 [5.2–12.0] | 12 | 5.0 [1.9–11.4] | 9.6 × 10−5 | 0.19 | 0.88 |
| CA | 21 | 11.4 [6.4–21.2] | 23 | 10.1 [5.6–15.0] | 15 | 5.3 [2.7–7.4] | 9 | 4.1 [0.6–7.1] | 2.0 × 10−5 | 0.44 | 0.86 |
| GCA | 21 | 1.0 [0.4–2.1] | 23 | 1.0 [0.7–1.5] | 16 | 0.1 [0.1–0.2] | 11 | 0.09 [0.07–0.2] | 4.3 × 10−12 | 0.80 | 0.67 |
| CDCA | 21 | 12.7 [2.3–18.2] | 23 | 10.7 [3.7–22.8] | 16 | 0.9 [0.3–2.5] | 11 | 0.7 [0.02–1.9] | 2.2 × 10−9 | 0.96 | 0.41 |
| TCDCA | 21 | 1.3 [0.9–2.1] | 23 | 1.0 [0.5–1.6] | 16 | 0.4 [0.2–0.8] | 11 | 0.3 [0.2–0.7] | 1.0 × 10−6 | 0.14 | 0.60 |
| UDCA | 21 | 0.1 [0.1–0.4] | 23 | 0.2 [0.1–0.6] | 16 | 0.1 [0.01–0.05] | 11 | 0.05 [0.03–0.1] | 3.9 × 10−5 | 0.17 | 0.29 |
| GUDCA | 15 | 0.005 [0.002–0.011] | 18 | 0.005 [0.003–0.0125] | 7 | 0.002 [0.001–0.004] | 10 | 0.002 [0.001–0.002] | 2.4 × 10−4 | 0.44 | 0.44 |
| LCA | 19 | 0.07 [0.04–0.1] | 23 | 0.09 [0.04–0.19] | 13 | 0.01 [0.01–0.03] | 8 | 0.008 [0.002–0.03] | 2.8 × 10−7 | 0.91 | 0.43 |
| αMUA | 20 | 3.7 [1.9–8.2] | 22 | 4.7 [2.0–8.0] | 13 | 0.8 [0.3–1.2] | 7 | 0.9 [0.5–1.2] | 3.0 × 10−6 | 0.52 | 0.98 |
| TαMUA | 21 | 1.8 [1.1–2.1] | 23 | 1.1 [0.8–1.9] | 16 | 0.7 [0.4–1.1] | 11 | 0.7 [0.3–0.9] | 1.0 × 10−5 | 0.13 | 0.84 |
| βMUA | 18 | 0.8 [0.4–1.6] | 22 | 0.59 [0.2–1.8] | 13 | 0.1 [0.04–0.2] | 7 | 0.1 [0.1–0.2] | 2.9 × 10−7 | 0.86 | 0.15 |
| THDCA | 18 | 0.04 [0.02–0.13] | 18 | 0.1 [0.1–0.2] | 16 | 0.2 [0.1–0.4] | 11 | 0.3 [0.2–0.8] | 2.0 × 10−6 | 0.08 | 0.93 |
| Bile Acid | Control | Exercise | HFD | HFD and Exercise | Two-Way ANOVA p | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | HFD | Exercise | Interaction | |
| Total | 15 | 33.9 [26.1–44.4] | 16 | 37.5 [30.6–42.8] | 13 | 70.6 [59.6–133.3] | 10 | 101.4 [58.5–122.8] | 7.3 × 10−9 | 0.30 | 0.83 |
| Conjugated | 15 | 0.5 [0.4–0.5] | 16 | 0.4 [0.4–0.6] | 13 | 1.1 [0.9–1.2] | 10 | 1.2 [1.0–1.7] | 1.2 × 10−14 | 0.92 | 0.49 |
| Free | 15 | 33.4 [25.6–43.8] | 16 | 37.0 [30.3–42.4] | 13 | 69.8 [58.5–131.9] | 10 | 100.4 [57.5–121.0] | 1.1 × 10−8 | 0.31 | 0.82 |
| Primary | 15 | 4.8 [3.7–7.5] | 16 | 5.7 [4.0–7.9] | 13 | 8.9 [5.3–18.5] | 10 | 8.5 [3.4–18.5] | 0.06 | 0.87 | 0.49 |
| Secondary | 15 | 30.3 [21.7–39.1] | 16 | 32.1 [25.8–37.8] | 13 | 67.1 [52.6–114.6] | 10 | 83.1 [41.5–107.3] | 2.3 × 10−9 | 0.33 | 0.89 |
| 12-α-OH | 15 | 3.3 [2.2–5.7] | 16 | 3.4 [2.7–4.5] | 13 | 25.4 [21.1–54.4] | 10 | 37.2 [22.2–47.7] | 1.6 × 10−15 | 0.52 | 0.84 |
| Non-12-α-OH | 15 | 31.1 [22.8–38.6] | 16 | 34.1 [27.7–38.7] | 13 | 47.2 [37.3–87.5] | 10 | 63.3 [36.5–75.1] | 5.8 × 10−5 | 0.44 | 0.57 |
| TCA | 15 | 0.1 ± 0.1 | 16 | 0.12 ± 0.04 | 13 | 0.5 ± 0.1 | 10 | 0.5 ± 0.1 | 1.4 × 10−27 | 0.64 | 0.54 |
| GCA | 15 | 0.05 ± 0.03 | 16 | 0.04 ± 0.03 | 13 | 0.1 ± 0.1 | 10 | 0.1 ± 0.1 | 1.3 × 10−4 | 0.57 | 0.78 |
| DCA | 15 | 2.8 [1.7–5.0] | 16 | 3.0 [2.2–3.8] | 13 | 23.8 [19.6–52.8] | 10 | 31.0 [16.8–44.8] | 4.1 × 10−15 | 0.51 | 0.99 |
| TDCA | 15 | 0.005 ± 0.002 | 16 | 0.006 ± 0.005 | 13 | 0.03 ± 0.02 | 10 | 0.04 ± 0.03 | 2.0 × 10−7 | 0.14 | 0.23 |
| GDCA | 14 | 0.006 ± 0.003 | 16 | 0.006 ± 0.005 | 13 | 0.02 ± 0.01 | 10 | 0.02 ± 0.01 | 9.3 × 10−5 | 0.77 | 0.77 |
| HDCA | 15 | 7.9 ± 5.3 | 16 | 12.5 ± 7.9 | 13 | 30.2 ± 17.3 | 10 | 34.1 ± 15.6 | 2.1 × 10−8 | 0.20 | 0.91 |
| THDCA | 15 | 0.05 ± 0.04 | 16 | 0.05 ± 0.03 | 13 | 0.1 ± 0.02 | 10 | 0.1 ± 0.03 | 1.3 × 10−5 | 0.54 | 0.82 |
| Ratio | Control | Exercise | HFD | HFD and Exercise | Two-Way ANOVA p | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | n | Median [IQR]/Mean ± SD | HFD | Exercise | Interaction | |
| Serum | |||||||||||
| TCA/GCA | 22 | 2.0 [0.7–7.3] | 22 | 1.0 [0.6–2.7] | 16 | 14.5 [8.8–21.7] | 11 | 14.6 [5.7–70.9] | 7.0 × 10−12 | 0.33 | 0.14 |
| CA/CDCA | 22 | 1.3 [0.8–2.4] | 22 | 0.8 [0.6–1.4] | 15 | 3.5 [2.1–5.6] | 9 | 4.1 [1.7–6.3] | 5.0 × 10−8 | 0.30 | 0.25 |
| TCDCA/CDCA | 20 | 0.1 [0.1–0.4] | 19 | 0.1 [0.03–0.31] | 4 | 0.5 [0.2–0.8] | 2 | 0.5 [0.1–17.1] | 2.1 × 10−4 | 0.82 | 0.52 |
| TLCA/LCA | 20 | 0.2 [0.1–0.3] | 22 | 0.1 [0.1–0.3] | 12 | 0.6 [0.3–0.8] | 9 | 1.2 [0.3–2.8] | 5.4 × 10−10 | 0.91 | 0.27 |
| THDCA/HDCA | 19 | 0.04 ± 0.06 | 17 | 0.05 ± 0.06 | 16 | 0.4 ± 0.6 | 11 | 0.7 ± 0.1 | 4.2 × 10−4 | 0.22 | 0.25 |
| Stool | |||||||||||
| DCA/CA | 16 | 8.0 [6.7–11.3] | 15 | 11.2 [6.4–14.3] | 13 | 48.9 [13.0–73.4] | 9 | 61.2 [25.2–86.6] | 9.1 × 19−5 | 0.55 | 0.78 |
| GDCA/DCA | 16 | 0.002 [0.001–0.003] | 15 | 0.002 [0.002–0.002] | 13 | 0.0004 [0.0003–0.0005] | 9 | 0.00054 [0.0004–0.0007] | 3.5 × 10−9 | 0.51 | 0.69 |
| TωMUA/ωMUA | 14 | 0.0002 [0.0001–0.0008] | 12 | 0.0004 [0.0002–0.0007] | 10 | 0.0008 [0.0005–0.0035] | 8 | 0.0018 [0.0006–0.006] | 4.7 × 10−4 | 0.22 | 0.82 |
| GHDCA/HDCA | 16 | 0.001 [0.001–0.002] | 15 | 0.001 [0.0006–0.002] | 13 | 0.0006 [0.0004–0.0008] | 9 | 0.0006 [0.0004–0.0008] | 8.0 × 10−6 | 0.17 | 0.44 |
| 12-α-OH/Non 12-α-OH | 16 | 0.1 ± 0.05 | 15 | 0.11 ± 0.02 | 13 | 0.5 ± 0.1 | 9 | 0.6 ± 0.5 | 1.7 × 10−23 | 0.74 | 0.24 |
| Stool vs. serum | |||||||||||
| CA | 16 | 0.02 [0.01–0.05] | 15 | 0.04 [0.02–0.06] | 13 | 0.3 [0.1–2.4] | 8 | 0.3 [0.1–1.6] | 2.7 × 10−7 | 0.64 | 0.72 |
| TCA | 16 | 0.05 [0.04–0.1] | 15 | 0.1 [0.1–0.12] | 13 | 0.3 [0.1–0.5] | 9 | 0.4 [0.2–1.3] | 7.1 × 10−7 | 0.007 | 0.70 |
| GCA | 16 | 0.03 [0.03–0.06] | 15 | 0.05 [0.02–0.08] | 13 | 0.7 [0.3–1.2] | 9 | 0.9 [0.5–1.2] | 3.1 × 10−15 | 0.28 | 0.59 |
| CDCA | 16 | 0.03 [0.01–0.13] | 15 | 0.03 [0.01–0.08] | 13 | 0.9 [0.2–3.8] | 9 | 1.0 [0.1–5.1] | 3.3 × 10−8 | 0.91 | 0.89 |
| TCDCA | 16 | 0.09 [0.06–0.11] | 15 | 0.1 [0.1–0.2] | 13 | 0.5 [0.1–0.7] | 9 | 0.4 [0.3–1.3] | 8.7 × 10−8 | 0.06 | 0.70 |
| GCDCA | 15 | 0.1 [0.1–0.2] | 12 | 0.1 [0.1–0.2] | 4 | 1.8 [0.3–4.3] | 2 | 3.7 [0.7–3.7] | 5.1 × 10−5 | 0.90 | 0.66 |
| DCA | 15 | 25.8 [12.3–53.9] | 15 | 18.4 [11.0–47.1] | 13 | 231.1 [207.8–509.0] | 9 | 340.9 [232.0–611.6] | 2.9 × 10−15 | 0.67 | 0.67 |
| TDCA | 16 | 0.1 [0.05–0.13] | 15 | 0.1 [0.1–0.3] | 13 | 0.7 [0.3–1.3] | 9 | 0.8 [0.3–2.4] | 6.9 × 10−9 | 0.048 | 0.67 |
| LCA | 15 | 100.8 [62.7–224.8] | 15 | 48.3 [26.0–201.8] | 11 | 502.0 [443.6–1394.9] | 7 | 650.6 [515.0–2786.1] | 8.2 × 10−8 | 0.54 | 0.49 |
| UDCA | 16 | 0.8 [0.2–1.8] | 15 | 0.6 [0.4–2.9] | 13 | 7.1 [3.9–17.5] | 9 | 2.8 [2.3–4.6] | 2.3 × 10−7 | 0.10 | 0.051 |
| Bile Acid\uCT Parameter | n | BV/TV | BS/TV | TbTh | TbN | CtAr | CtAr/Tt.Ar | CtTh | PsPm | EcPm | Imax | Imin | J |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Serum | |||||||||||||
| Total BAs | 39 | 0.500 (0.001) | 0.484 (0.002) | 0.513 (0.001) | 0.525 (0.001) | 0.556 (2.3 × 10−4) | 0.602 (5.1 × 10−5) | 0.586 (8.9 × 10−5) | 0.531 (0.001) | 0.399 (0.012) | 0.384 (0.016) | 0.488 (0.002) | 0.422 (0.007) |
| Free BAs | 39 | 0.443 (0.005) | 0.426 (0.007) | 0.523 (0.001) | 0.430 (0.006) | 0.582 (1.0 × 10−4) | 0.564 (1.9 × 10−4) | 0.609 (3.8 × 10−5) | 0.522 (0.001) | 0.391 (0.014) | 0.342 (0.033) | 0.545 (3.3 × 10−4) | 0.430 (0.006) |
| Primary BAs | 0.543 (3.5 × 10−4) | 0.529 (5.4 × 10−4) | 0.541 (3.7 × 10−4) | 0.563 (1.9 × 10−4) | 0.572 (1.4 × 10−4) | 0.633 (1.5 × 10−5) | 0.609 (3.9 × 10−5) | 0.545 (3.4 × 10−4) | 0.392 (0.014) | 0.388 (0.0159 | 0.488 (0.002) | 0.420 (0.008) | |
| Non-12-α-OH BAs | 39 | 0.456 (0.004) | 0.450 (0.004) | 0.499 (0.001) | 0.469 (0.003) | 0.580 (1.1 × 10−4) | 0.593 (7.1 × 10−5) | 0.584 (9.5 × 10−5) | 0.553 (2.6 × 10−4) | 0.444 (0.005) | 0.419 (0.008) | 0.527 (0.001) | 0.463 (0.003) |
| GCA | 38 | 0.539 (4.8 × 10−4) | 0.550 (3.5 × 10−4) | 0.586 (1.1 × 10−4) | 0.553 (3.2 × 10−4) | 0.619 (3.2 × 10−5) | 0.624 (2.8 × 10−5) | 0.649 (1.0 × 10−5) | 0.641 (1.5 × 10−5) | 0.496 (0.001) | 0.424 (0.008) | 0.599 (7.1 × 10−5) | 0.511 (0.001) |
| CDCA | 38 | 0.480 (0.002) | 0.467 (0.003) | 0.545 (4.0 × 10−4) | 0.466 (0.003) | 0.631 (2.1 × 10−5) | 0.610 (4.9 × 10−5) | 0.651 (9.5 × 10−6) | 0.575 (1.6 × 10−4) | 0.437 (0.003) | 0.381 (0.02) | 0.590 (9.7 × 10−5) | 0.478 (0.002) |
| αMUA | 32 | 0.562 (8.2 × 10−4) | 0.543 (0.001) | 0.602 (2.6 × 10−4) | 0.561 (8.3 × 10−4) | 0.677 (2.1 × 10−5) | 0.708 (5.6 × 10−6) | 0.691 (1.2 × 10−5) | 0.672 (2.5 × 10−5) | 0.516 (0.003) | 0.434 (0.01) | 0.622 (1.4 × 10−4) | 0.513 (0.003) |
| Stool | |||||||||||||
| Conjugated BAs | 32 | −0.606 (2.3 × 10−4) | −0.630 (1.1 × 10−4) | −0.680 (1.8 × 10−5) | −0.583 (4.6 × 10−4) | −0.593 (3.5 × 10−4) | −0.708 (5.8 × 10−6) | −0.689 (1.3 × 10−5) | −0.595 (3.3 × 10−4) | −0.409 (0.020) | −0.291 (0.107) | −0.425 (0.015) | −0.345 (0.053) |
| 12-α-OH BAs | 32 | −0.579 (0.001) | −0.593 (3.5 × 10−4) | −0.618 (1.7 × 10−4) | −0.565 (7.6 × 10−4) | −0.601 (2.7 × 10−4) | −0.662 (3.6 × 10−5) | −0.651 (5.5 × 10−5) | −0.583 (4.6 × 10−4) | −0.474 (0.006) | −0.384 (0.030) | −0.504 (0.003) | −0.427 (0.015) |
| TCA | 32 | −0.618 (1.7 × 10−4) | −0.656 (4.6 × 10−5) | −0.750 (7.6 × 10−7) | −0.595 (3.3 × 10−4) | −0.734 (1.7 × 10−6) | −0.778 (1.6 × 10−7) | −0.805 (2.7 × 10−8) | −0.706 (6.2 × 10−6) | −0.541 (0.003) | −0.418 (0.02) | −0.631 (1.1 × 10−4) | −0.516 (0.002) |
| Bile acid ratios | |||||||||||||
| TCA/GCA (serum) | 38 | −0.491 (0.002) | −0.500 (0.001) | −0.509 (0.001) | −0.474 (0.003) | −0.606 (5.5 × 10−5) | −0.534 (5.5 × 10−4) | −0.594 (8.5 × 10−5) | −0.580 (1.4 × 10−4) | −0.489 (0.002) | −0.439 (0.006) | −0.570 (1.8 × 10−4) | −0.500 (0.001) |
| CA/CDCA (serum) | 36 | −0.452 (0.005) | −0.463 (0.004) | −0.515 (0.001) | −0.457 (0.005) | −0.686 (3.9 × 10−6) | −0.633 (3.4 × 10−5) | −0.641 (2.6 × 10−5) | −0.665 (9.8 × 10−6) | −0.637 (3.0 × 10−5) | −0.543 (6.2 × 10−4) | −0.672 (7.3 × 10−6) | −0.606 (9.0 × 10−5) |
| TLCA/LCA (serum) | 34 | −0.528 (0.001) | −0.520 (0.002) | −0.544 (0.001) | −0.457 (0.007) | −0.628 (7.0 × 10−5) | −0.589 (2.5 × 10−4) | −0.647 (3.6 × 10−5) | −0.541 80.001) | −0.382 (0.026) | −0.416 (0.014) | −0.544 (0.001) | −0.474 (0.005) |
| TωMUA/ωMUA (stool) | 26 | −0.683 (1.2 × 10−4) | −0.699 (7.1 × 10−5) | −0.448 (0.02) | −0.679 (1.4 × 10−4) | −0.411 (0.037) | −0.521 (0.006) | −0.469 (0.016) | −0.449 (0.021) | −0.287 (0.155) | −0.164 (0.424) | −0.289 (0.152) | −0.198 (0.332) |
| CA (stool/serum) | 31 | −0.416 (0.020) | −0.473 80.007) | −0.514 (0.003) | −0.446 (0.012) | −0.521 (0.003) | −0.639 (1.1 × 10−4) | −0.617 (2.2 × 10−4) | −0.517 (0.003) | −0.394 (0.028) | −0.170 (0.359) | −0.430 (0.016) | −0.298 (0.104) |
| GCA (stool/serum) | 31 | −0.604 (3.2 × 10−4) | −0.649 (7.8 × 10−5) | −0.615 (2.3 × 10−4) | −0.653 (6.8 × 10−5) | −0.615 (2.3 × 10−4) | −0.702 (1.1 × 10−5) | −0.690 (1.7 × 10−5) | −0.634 (1.3 × 10−4) | −0443 (0.013) | −0.360 (0.047) | −0.487 (0.005) | −0.407 (0.023) |
| CDCA (stool/serum) | 31 | −0.469 (0.008) | −0.547 (0.001) | −0.554 (0.001) | −0.511 (0.003) | −0.644 (9.1 × 10−5) | −0.699 (1.2 × 10−5) | −0.695 (1.4 × 10−5) | −0.640 (1.1 × 10−4) | −0.511 (0.003) | −0.312 (0.088) | −0.537 (0.002) | −0.427 (0.017) |
| DCA (stool/serum) | 31 | −0.679 (2.7 × 10−5) | −0.683 (2.3 × 10−5) | −0.673 (3.3 × 10−5) | −0.667 (4.2 × 10−5) | −0.726 (3.7 × 10−6) | −0.771 (3.9 × 10−7) | −0.777 (2.8 × 10−7) | −0.685 (2.1 × 10−5) | −0.511 (0.003) | −0.471 (0.007) | −0.607 (2.9 × 10−4) | −0.523 (0.003) |
| LCA (stool/serum) | 29 | −0.595 (0.001) | −0.584 (0.001) | −0.544 (0.002) | −0.587 (0.001) | −0.674 (6.1 × 10−5) | −0.624 (3.0 × 10−4) | −0.618 (3.6 × 10−4) | −0.644 (1.6 × 10−4) | −0.569 (0.001) | −0.565 (0.001) | −0.658 (1.0 × 10−4) | −0.618 (3.5 × 10−4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, N.; Almer, G.; Semeraro, M.D.; Rodriguez-Blanco, G.; Fauler, G.; Anders, I.; Ritter, G.; vom Scheidt, A.; Hammer, N.; Gruber, H.-J.; et al. Impact of High-Fat Diet and Exercise on Bone and Bile Acid Metabolism in Rats. Nutrients 2024, 16, 1744. https://doi.org/10.3390/nu16111744
Alonso N, Almer G, Semeraro MD, Rodriguez-Blanco G, Fauler G, Anders I, Ritter G, vom Scheidt A, Hammer N, Gruber H-J, et al. Impact of High-Fat Diet and Exercise on Bone and Bile Acid Metabolism in Rats. Nutrients. 2024; 16(11):1744. https://doi.org/10.3390/nu16111744
Chicago/Turabian StyleAlonso, Nerea, Gunter Almer, Maria Donatella Semeraro, Giovanny Rodriguez-Blanco, Günter Fauler, Ines Anders, Gerald Ritter, Annika vom Scheidt, Niels Hammer, Hans-Jürgen Gruber, and et al. 2024. "Impact of High-Fat Diet and Exercise on Bone and Bile Acid Metabolism in Rats" Nutrients 16, no. 11: 1744. https://doi.org/10.3390/nu16111744
APA StyleAlonso, N., Almer, G., Semeraro, M. D., Rodriguez-Blanco, G., Fauler, G., Anders, I., Ritter, G., vom Scheidt, A., Hammer, N., Gruber, H.-J., & Herrmann, M. (2024). Impact of High-Fat Diet and Exercise on Bone and Bile Acid Metabolism in Rats. Nutrients, 16(11), 1744. https://doi.org/10.3390/nu16111744