

Exploring the Association between Anxiety, Depression, and Gut Microbiota during Pregnancy: Findings from a Pregnancy Cohort Study in Shijiazhuang, Hebei Province, China

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
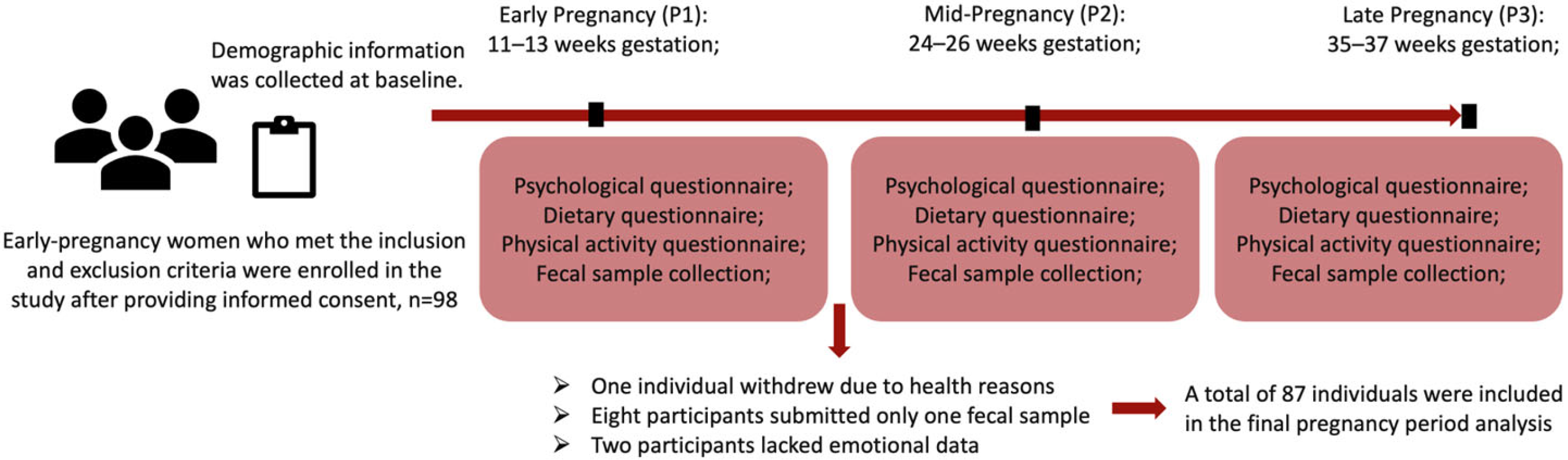
2.2. Study Design
2.3. Population
2.4. Measurements
2.4.1. Basic Information
2.4.2. Measurement of Anxiety
2.4.3. Measurement of Depression
2.4.4. Fecal Sample Collection
2.4.5. Shotgun Metagenomic Profiling
2.4.6. Assessment of Other Covariates
2.4.7. Statistical Analysis
3. Results
3.1. Characteristics of Participants
3.2. Anxiety and Depression
3.3. An Overview of Gut-Microbiota Findings
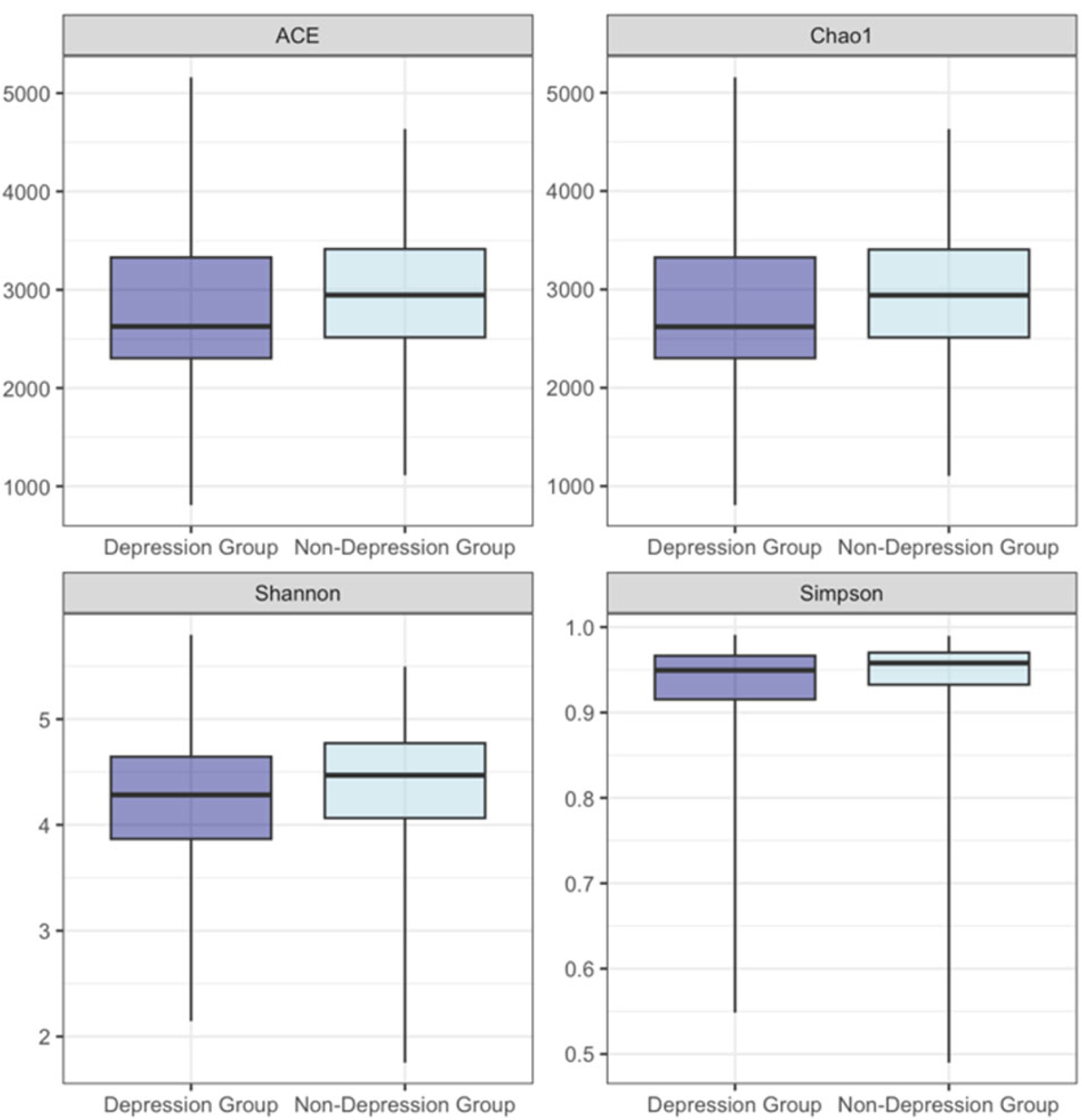
3.4. Alpha Diversity
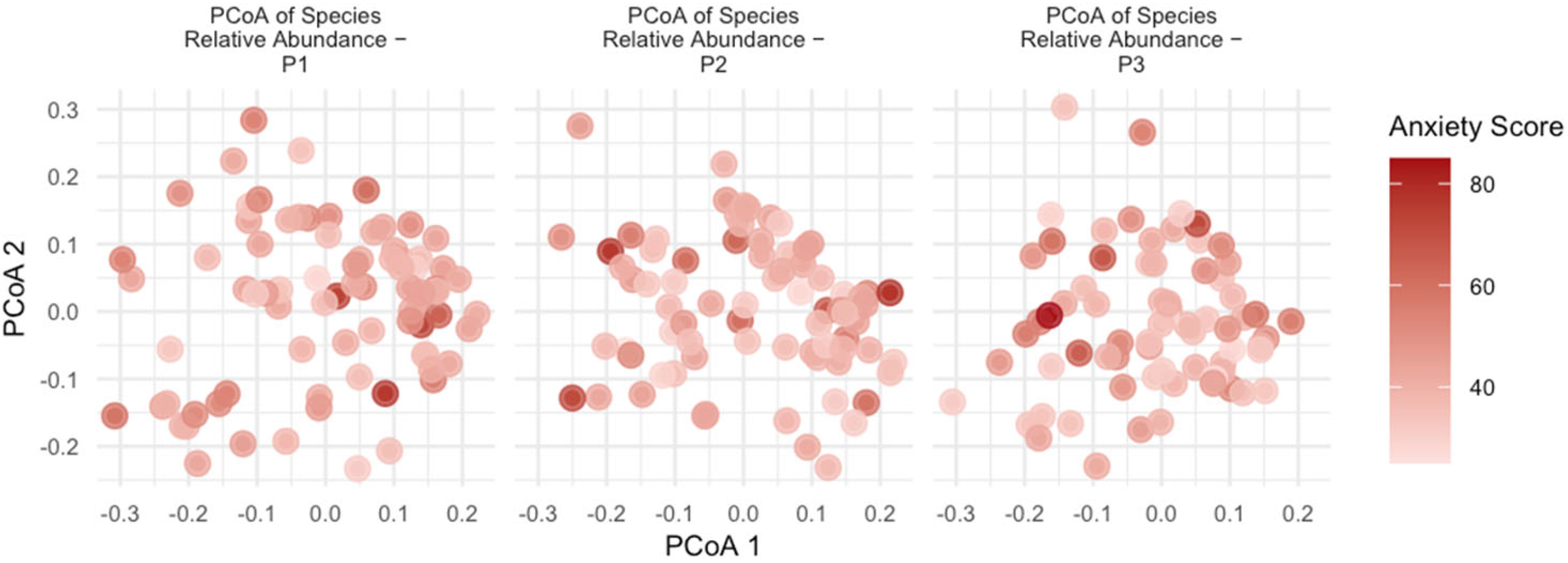
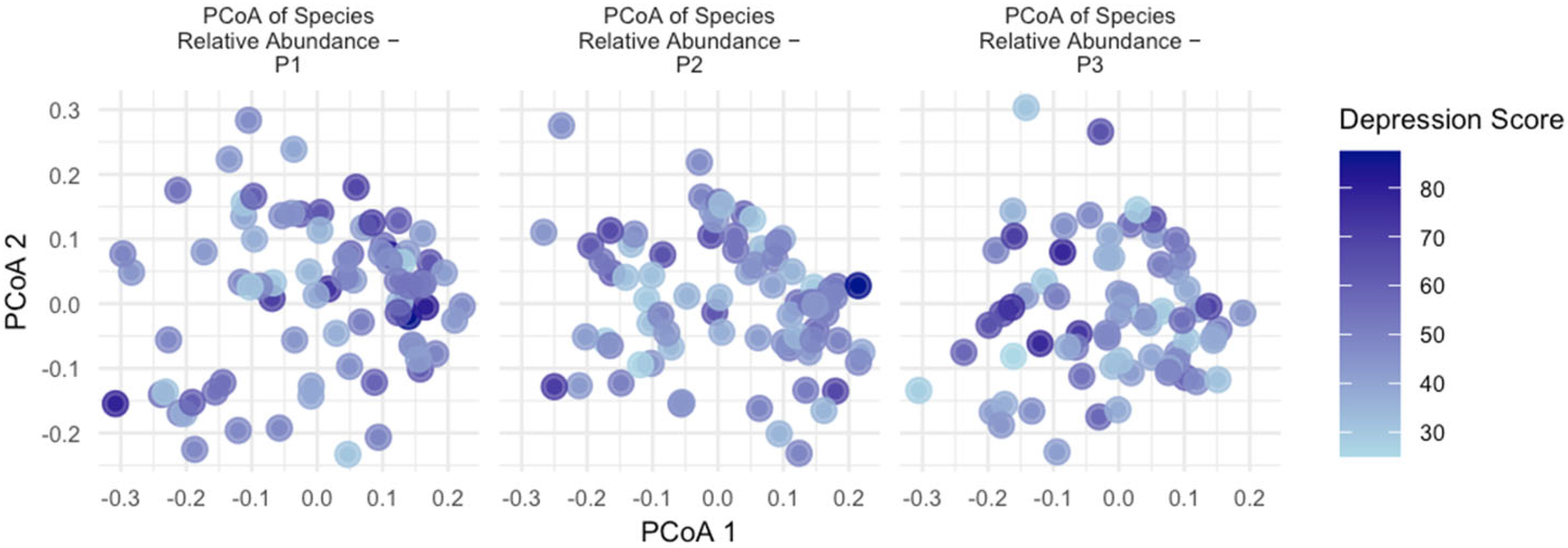
3.5. Beta Diversity
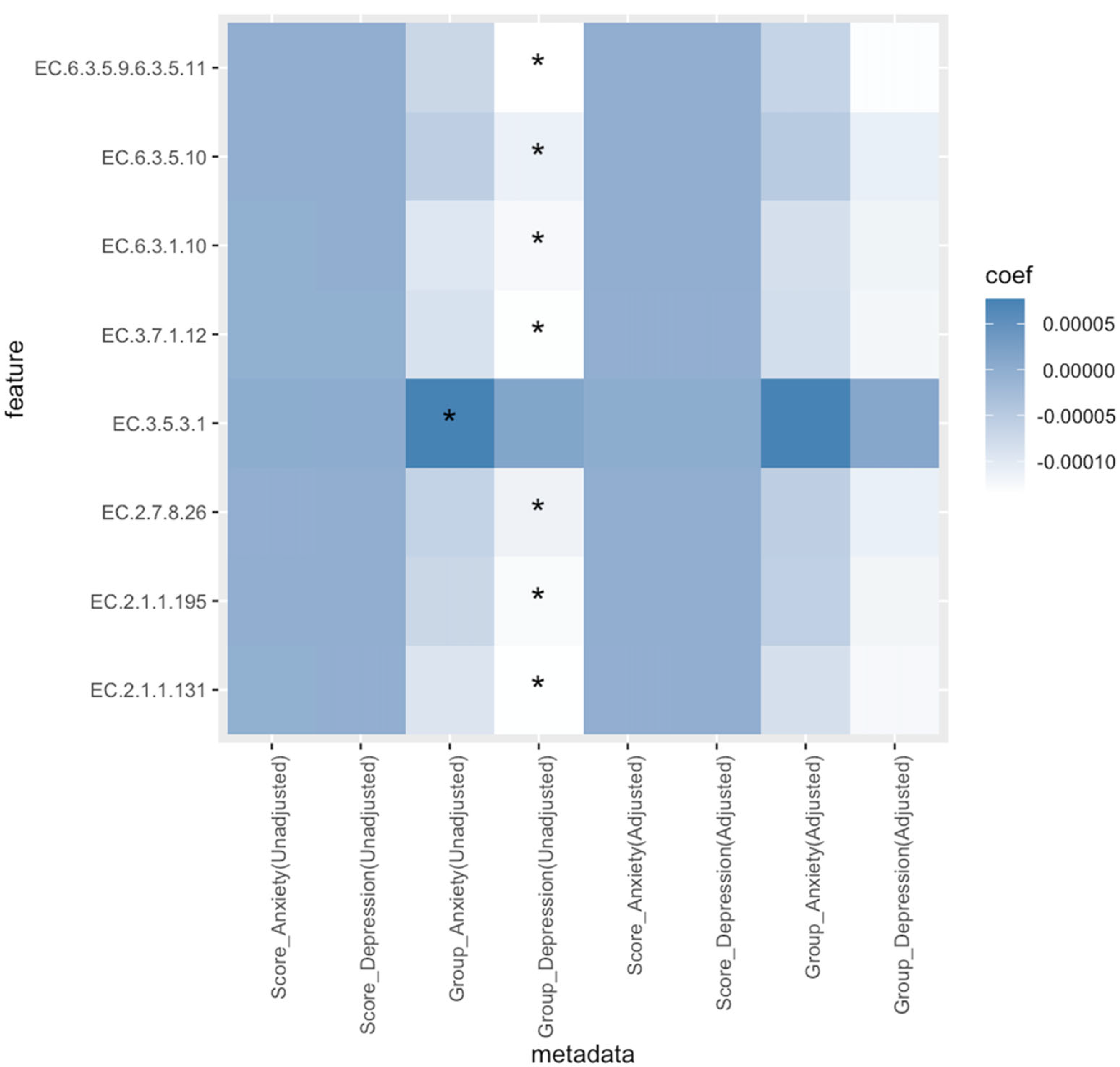
3.6. Taxonomies and Pathways Associated with Anxiety and Depression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dennis, C.L.; Falah-Hassani, K.; Shiri, R. Prevalence of antenatal and postnatal anxiety: Systematic review and meta-analysis. Br. J. Psychiatry 2017, 210, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Woody, C.A.; Ferrari, A.J.; Siskind, D.J.; Whiteford, H.A.; Harris, M.G. A systematic review and meta-regression of the prevalence and incidence of perinatal depression. J. Affect. Disord. 2017, 219, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, K.; Fellmeth, G.; Edwards, S.; Calvert, C.; Bennett, P.; Campbell, O.M.R.; Fuhr, D.C. The global burden of perinatal common mental health disorders and substance use among migrant women: A systematic review and meta-analysis. Lancet Public Health 2023, 8, e203–e216. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, H.C.; Sullivan, E.L.; Nousen, E.K.; Sullivan, C.A.; Huang, E.; Rincon, M.; Nigg, J.T.; Loftis, J.M. Maternal prenatal depression predicts infant negative affect via maternal inflammatory cytokine levels. Brain Behav. Immun. 2018, 73, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Qin, S. Mental health at the forefront of women-centred health-care systems. Lancet 2023, 402, 445. [Google Scholar] [CrossRef] [PubMed]
- Sherer, M.L.; Voegtline, K.M.; Park, H.S.; Miller, K.N.; Shuffrey, L.C.; Klein, S.L.; Osborne, L.M. The immune phenotype of perinatal anxiety. Brain Behav. Immun. 2022, 106, 280–288. [Google Scholar] [CrossRef]
- Brann, E.; Skalkidou, A.; Schwarz, J.; Papadopoulos, F.C.; Sundstrom Poromaa, I.; Fransson, E. Longitudinal assessment of inflammatory markers in the peripartum period by depressive symptom trajectory groups. Brain Behav. Immun. Health 2022, 22, 100468. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wan, J.; Choe, U.; Pham, Q.; Schoene, N.W.; He, Q.; Li, B.; Yu, L.; Wang, T.T.Y. Interactions between Food and Gut Microbiota: Impact on Human Health. Annu. Rev. Food Sci. Technol. 2019, 10, 389–408. [Google Scholar] [CrossRef]
- Lu, X.W.; Shi, Z.; Jiang, L.L.; Zhang, S.Y. Maternal gut microbiota in the health of mothers and offspring: From the perspective of immunology. Front. Immunol. 2024, 15, 1362784. [Google Scholar] [CrossRef]
- Fox, C.; Eichelberger, K. Maternal microbiome and pregnancy outcomes. Fertil. Steril. 2015, 104, 1358–1363. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Robertson, R.C.; Manges, A.R.; Finlay, B.B.; Prendergast, A.J. The Human Microbiome and Child Growth—First 1000 Days and Beyond. Trends Microbiol. 2019, 27, 131–147. [Google Scholar] [CrossRef]
- Fung, T.C. The microbiota-immune axis as a central mediator of gut-brain communication. Neurobiol. Dis. 2020, 136, 104714. [Google Scholar] [CrossRef]
- Galley, J.D.; Mashburn-Warren, L.; Blalock, L.C.; Lauber, C.L.; Carroll, J.E.; Ross, K.M.; Hobel, C.; Coussons-Read, M.; Dunkel Schetter, C.; Gur, T.L. Maternal anxiety, depression and stress affects offspring gut microbiome diversity and bifidobacterial abundances. Brain Behav. Immun. 2023, 107, 253–264. [Google Scholar] [CrossRef]
- Christian, L.M. The gut microbiome and mental health: Taking baby steps. Brain Behav. Immun. 2019, 81, 22–23. [Google Scholar] [CrossRef]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef]
- Fang, Q.; Tu, Y.; Fan, X.; Zang, T.; Wu, N.; Qiu, T.; Li, Y.; Bai, J.; Liu, Y. Inflammatory cytokines and prenatal depression: Is there a mediating role of maternal gut microbiota? J. Psychiatr. Res. 2023, 164, 458–467. [Google Scholar] [CrossRef]
- Rajasekera, T.A.; Galley, J.D.; Mackos, A.R.; Chen, H.J.; Mitchell, J.G.; Kleinman, J.J.; Cappelucci, P.; Mashburn-Warren, L.; Lauber, C.L.; Bailey, M.T.; et al. Stress and depression-associated shifts in gut microbiota: A pilot study of human pregnancy. Brain Behav. Immun. Health 2024, 36, 100730. [Google Scholar] [CrossRef]
- Hechler, C.; Borewicz, K.; Beijers, R.; Saccenti, E.; Riksen-Walraven, M.; Smidt, H.; de Weerth, C. Association between Psychosocial Stress and Fecal Microbiota in Pregnant Women. Sci. Rep. 2019, 9, 4463. [Google Scholar] [CrossRef]
- Zhou, B.F.; Coorperative Meta-Analysis Group of China Obesity Task Force. Predictive values of body mass index and waist circumference for risk factors of certain related diseases in Chinese adults—Study on optimal cut-off points of body mass index and waist circumference in Chinese adults. Biomed. Environ. Sci. 2002, 15, 83–96. [Google Scholar]
- Zung, W.W. A rating instrument for anxiety disorders. Psychosom. J. Consult. Liaison Psychiatry 1971, 12, 371–379. [Google Scholar] [CrossRef]
- Yue, C.Y.; Liu, C.P.; Wang, J.; Zhang, M.; Wu, H.J.; Li, C.R.; Yang, X.L. Association between social support and anxiety among pregnant women in the third trimester during the coronavirus disease 2019 (COVID-19) epidemic in Qingdao, China: The mediating effect of risk perception. Int. J. Soc. Psychiatry 2021, 67, 120–127. [Google Scholar] [CrossRef]
- Liao, M.L.; Fang, F.; Liu, G.X.; Zhang, Y.X.; Deng, C.Q.; Zhang, X.Q. Influencing factors and correlation of anxiety, psychological stress sources, and psychological capital among women pregnant with a second child in Guangdong and Shandong Province. J. Affect. Disord. 2020, 264, 115–122. [Google Scholar] [CrossRef]
- Yue, J.M.; Zang, X.Y.; Le, Y.Y.; An, Y.Y. Anxiety, depression and PTSD among children and their parent during 2019 novel coronavirus disease (COVID-19) outbreak in China. Curr. Psychol. 2022, 41, 5723–5730. [Google Scholar] [CrossRef]
- Zung, W.W. A self-rating depression scale. Arch. Gen. Psychiatry 1965, 12, 63–70. [Google Scholar] [CrossRef]
- Biggs, J.T.; Wylie, L.T.; Ziegler, V.E. Validity of the Zung self-rating depression scale. Br. J. Psychiatry 1978, 132, 381–385. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, J.G.; Baker, P.N.; Liao, B.Z.; Yu, X.Y. The prevalence and associated factors of prenatal depression and anxiety in twin pregnancy: A cross-sectional study in Chongqing, China. BMC Pregnancy Childbirth 2022, 22, 877. [Google Scholar] [CrossRef]
- Chen, Q.Q.; Lin, F.M.; Chen, D.H.; Ye, Y.M.; Gong, G.M.; Chen, F.F.; Huang, S.F.; Peng, S.L. Analysis of mental health status and related factors in patients with acute cerebral infarction. World J. Psychiatry 2023, 13, 793–802. [Google Scholar] [CrossRef]
- Wang, Y.N.; Di, Y.; Ye, J.J.; Wei, W.B. Study on the public psychological states and its related factors during the outbreak of coronavirus disease 2019 (COVID-19) in some regions of China. Psychol. Health Med. 2021, 26, 13–22. [Google Scholar] [CrossRef]
- Li, M.X.; Zhang, G.H.; Cui, L.J.; Zhang, L.; Zhou, Q.; Mu, C.X.; Chi, R.X.; Zhang, N.; Ma, G.S. Dynamic changes in gut microbiota during pregnancy among Chinese women and influencing factors: A prospective cohort study. Front. Microbiol. 2023, 14, 1114228. [Google Scholar] [CrossRef]
- Shahar, D.; Shai, I.; Vardi, H.; Brener-Azrad, A.; Fraser, D. Development of a semi-quantitative Food Frequency Questionnaire (FFQ) to assess dietary intake of multiethnic populations. Eur. J. Epidemiol. 2003, 18, 855–861. [Google Scholar] [CrossRef]
- Prevention CCFDCA. Chinese Food Composition Table, 2nd ed.; Peking University Medical Press: Beijing, China, 2009. [Google Scholar]
- Fan, M.; Lyu, J.; He, P. Chinese guidelines for data processing and analysis concerning the International Physical Activity Questionnaire. Zhonghua Liu Xing Bing Xue Za Zhi 2014, 35, 961–964. [Google Scholar]
- Feranchuk, S.; Belkova, N.; Potapova, U.; Kuzmin, D.; Belikov, S. Evaluating the use of diversity indices to distinguish between microbial communities with different traits. Res. Microbiol. 2018, 169, 254–261. [Google Scholar] [CrossRef]
- Li, Y.P.; Wang, D.D.; Satija, A.; Ivey, K.L.; Li, J.; Wilkinson, J.E.; Li, R.F.; Baden, M.; Chan, A.T.; Huttenhower, C.; et al. Plant-Based Diet Index and Metabolic Risk in Men: Exploring the Role of the Gut Microbiome. J. Nutr. 2021, 151, 2780–2789. [Google Scholar] [CrossRef]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable association discovery in population-scale meta-omics studies. PLoS Comput. Biol. 2021, 17, e1009442. [Google Scholar] [CrossRef]
- Wang, K.; Mehta, R.S.; Ma, W.; Nguyen, L.H.; Wang, D.D.; Ghazi, A.R.; Yan, Y.; Al-Shaar, L.; Wang, Y.; Hang, D.; et al. The gut microbiome modifies the associations of short- and long-term physical activity with body weight changes. Microbiome 2023, 11, 121. [Google Scholar] [CrossRef]
- Kennedy, K.M.; Plagemann, A.; Sommer, J.; Hofmann, M.; Henrich, W.; Barrett, J.F.R.; Surette, M.G.; Atkinson, S.; Braun, T.; Sloboda, D.M. Parity modulates impact of BMI and gestational weight gain on gut microbiota in human pregnancy. Gut Microbes 2023, 15, 2259316. [Google Scholar] [CrossRef]
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for enriching next-generation health-promoting gut bacteria through prebiotics and other dietary components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef]
- Mareckova, K.; Marecek, R.; Jani, M.; Zackova, L.; Andryskova, L.; Brazdil, M.; Nikolova, Y.S. Association of Maternal Depression During Pregnancy and Recent Stress With Brain Age Among Adult Offspring. JAMA Netw. Open 2023, 6, e2254581. [Google Scholar] [CrossRef]
- Borkent, J.; Ioannou, M.; Laman, J.D.; Haarman, B.C.M.; Sommer, I.E.C. Role of the gut microbiome in three major psychiatric disorders. Psychol. Med. 2022, 52, 1222–1242. [Google Scholar] [CrossRef]
- Kumar, A.; Pramanik, J.; Goyal, N.; Chauhan, D.; Sivamaruthi, B.S.; Prajapati, B.G.; Chaiyasut, C. Gut Microbiota in Anxiety and Depression: Unveiling the Relationships and Management Options. Pharmaceuticals 2023, 16, 565. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, Y.; Celniker, S.E.; Xia, Y.; Mao, J.H.; Snijders, A.M.; Chang, H. Gut microbiome partially mediates and coordinates the effects of genetics on anxiety-like behavior in Collaborative Cross mice. Sci. Rep. 2021, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Yay, E.; Yilmaz, M.; Toygar, H.; Balci, N.; Rivas, C.A.; Kilic, B.B.; Zirh, A.; Paster, B.; Kantarci, A. Parkinson′s disease alters the composition of subgingival microbiome. J. Oral Microbiol. 2023, 15, 2250650. [Google Scholar] [CrossRef]
- Kim, S.Y.; Woo, S.Y.; Raza, S.; Ho, D.; Jeon, S.W.; Chang, Y.; Ryu, S.; Kim, H.L.; Kim, H.N. Association between gut microbiota and anxiety symptoms: A large population-based study examining sex differences. J. Affect. Disord. 2023, 333, 21–29. [Google Scholar] [CrossRef]
- Kraaij, R.; Schuurmans, I.K.; Radjabzadeh, D.; Tiemeier, H.; Dinan, T.G.; Uitterlinden, A.G.; Hillegers, M.; Jaddoe, V.W.V.; Duijts, L.; Moll, H.; et al. The gut microbiome and child mental health: A population-based study. Brain Behav. Immun. 2023, 108, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.I.; Bastiaanssen, T.F.S.; Long-Smith, C.; Morkl, S.; Berding, K.; Ritz, N.L.; Strain, C.; Patangia, D.; Patel, S.; Stanton, C.; et al. The gut microbiome in social anxiety disorder: Evidence of altered composition and function. Transl. Psychiatry 2023, 13, 95. [Google Scholar] [CrossRef]
- Ma, K.; Guo, L.; Xu, A.; Cui, S.; Wang, J.H. Molecular Mechanism for Stress-Induced Depression Assessed by Sequencing miRNA and mRNA in Medial Prefrontal Cortex. PLoS ONE 2016, 11, e0159093. [Google Scholar] [CrossRef]
- Reininghaus, E.Z.; Platzer, M.; Kohlhammer-Dohr, A.; Hamm, C.; Morkl, S.; Bengesser, S.A.; Fellendorf, F.T.; Lahousen-Luxenberger, T.; Leitner-Afschar, B.; Schoggl, H.; et al. PROVIT: Supplementary Probiotic Treatment and Vitamin B7 in Depression-A Randomized Controlled Trial. Nutrients 2020, 12, 3422. [Google Scholar] [CrossRef] [PubMed]
- Brushett, S.; Gacesa, R.; Vila, A.V.; Gois, M.F.B.; Andreu-Sanchez, S.; Swarte, J.C.; Klaassen, M.A.Y.; Collij, V.; Sinha, T.; Bolte, L.A.; et al. Gut feelings: The relations between depression, anxiety, psychotropic drugs and the gut microbiome. Gut Microbes 2023, 15, 2281360. [Google Scholar] [CrossRef]
- Thomann, A.K.; Wüstenberg, T.; Wirbel, J.; Knoedler, L.L.; Thomann, P.A.; Zeller, G.; Ebert, M.P.; Lis, S.; Reindl, W. Depression and fatigue in active IBD from a microbiome perspective-a Bayesian approach to faecal metagenomics. BMC Med. 2022, 20, 366. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ryu, J.S.; Rim, C.; Shin, J.U.; Kwon, M.S. Possible role of arginase 1 positive microglia on depressive/anxiety-like behaviors in atopic dermatitis mouse model. Arch. Pharm. Res. 2022, 45, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Elgun, S.; Kumbasar, H. Increased serum arginase activity in depressed patients. Prog. Neuropsychopharmacol. Biol. Psychiatry 2000, 24, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Stiernborg, M.; Debelius, J.W.; Yang, L.L.; Skott, E.; Millischer, V.; Giacobini, M.; Melas, P.A.; Boulund, F.; Lavebratt, C. Bacterial gut microbiome differences in adults with ADHD and in children with ADHD on psychostimulant medication. Brain Behav. Immun. 2023, 110, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Fresquet, V.; Williams, L.; Raushel, F.M. Mechanism of cobyrinic acid a,c-diamide synthetase from Salmonella typhimurium LT2. Biochemistry 2004, 43, 10619–10627. [Google Scholar] [CrossRef]
- Maggio-Hall, L.A.; Escalante-Semerena, J.C. In vitro synthesis of the nucleotide loop of cobalamin by Salmonella typhimurium enzymes. Proc. Natl. Acad. Sci. USA 1999, 96, 11798–11803. [Google Scholar] [CrossRef]
- Roper, J.M.; Raux, E.; Brindley, A.A.; Schubert, H.L.; Gharbia, S.E.; Shah, H.N.; Warren, M.J. The enigma of cobalamin (vitamin B) biosynthesis in: Identification and characterization of a functional corrin pathway. J. Biol. Chem. 2000, 275, 40316–40323. [Google Scholar] [CrossRef]
- Scott, A.I.; Roessner, C.A.; Stolowich, N.J.; Spencer, J.B.; Min, C.; Ozaki, S.I. Biosynthesis of vitamin B12. Discovery of the enzymes for oxidative ring contraction and insertion of the fourth methyl group. FEBS Lett. 1993, 331, 105–108. [Google Scholar] [CrossRef]
- Warren, M.J.; Raux, E.; Schubert, H.L.; Escalante-Semerena, J.C. The biosynthesis of adenosylcobalamin (vitamin B12). Nat. Prod. Rep. 2002, 19, 390–412. [Google Scholar] [CrossRef]
- Moore, S.J.; Lawrence, A.D.; Biedendieck, R.; Deery, E.; Frank, S.; Howard, M.J.; Rigby, S.E.; Warren, M.J. Elucidation of the anaerobic pathway for the corrin component of cobalamin (vitamin B12). Proc. Natl. Acad. Sci. USA 2013, 110, 14906–14911. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Xiang, S.S.; Ye, K.; Zheng, Y.Q.; Feng, X.; Zhu, X.; Chen, J.; Chen, Y.W. Cobalamin (Vitamin B12) Induced a Shift in Microbial Composition and Metabolic Activity in an Colon Simulation. Front. Microbiol. 2018, 9, 2780. [Google Scholar] [CrossRef]
- Guetterman, H.M.; Huey, S.L.; Knight, R.; Fox, A.M.; Mehta, S.; Finkelstein, J.L. Vitamin B-12 and the Gastrointestinal Microbiome: A Systematic Review. Adv. Nutr. 2022, 13, 530–558. [Google Scholar] [CrossRef] [PubMed]
- Sangle, P.; Sandhu, O.; Aftab, Z.; Anthony, A.T.; Khan, S. Vitamin B12 Supplementation: Preventing Onset and Improving Prognosis of Depression. Cureus J. Med. Sci. 2020, 12, e11169. [Google Scholar] [CrossRef] [PubMed]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The Gut-Brain Axis and the Microbiome: Mechanisms and Clinical Implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Rieder, R.; Wisniewski, P.J.; Alderman, B.L.; Campbell, S.C. Microbes and mental health: A review. Brain Behav. Immun. 2017, 66, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Brain-Gut-Microbiota Axis and Mental Health. Psychosom. Med. 2017, 79, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M., Jr.; Schatzberg, A.F. HPA axis in major depression: Cortisol, clinical symptomatology and genetic variation predict cognition. Mol. Psychiatry 2017, 22, 527–536. [Google Scholar] [CrossRef]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.G.; Sharpley, C.F. Gut microbiome and depression: What we know and what we need to know. Rev. Neurosci. 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Doran, C.M.; Kinchin, I. A review of the economic impact of mental illness. Aust. Health Rev. 2019, 43, 43–48. [Google Scholar] [CrossRef]
- Rhee, S.J.; Kim, H.; Lee, Y.; Lee, H.J.; Park, C.H.K.; Yang, J.; Kim, Y.K.; Ahn, Y.M. The association between serum microbial DNA composition and symptoms of depression and anxiety in mood disorders. Sci. Rep. 2021, 11, 13987. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, A.; Furukawa, T.A.; Salanti, G.; Chaimani, A.; Atkinson, L.Z.; Ogawa, Y.; Leucht, S.; Ruhe, H.G.; Turner, E.H.; Higgins, J.P.T.; et al. Comparative Efficacy and Acceptability of 21 Antidepressant Drugs for the Acute Treatment of Adults With Major Depressive Disorder: A Systematic Review and Network Meta-Analysis. Focus 2018, 16, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Deschasaux, M.; Bouter, K.E.; Prodan, A.; Levin, E.; Groen, A.K.; Herrema, H.; Tremaroli, V.; Bakker, G.J.; Attaye, I.; Pinto-Sietsma, S.J.; et al. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nat. Med. 2018, 24, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zuo, T.; Cheung, C.P.; Gu, W.X.; Wan, Y.T.; Zhang, F.; Chen, N.; Zhan, H.; Yeoh, Y.K.; Niu, J.K.; et al. Population-Level Configurations of Gut Mycobiome Across 6 Ethnicities in Urban and Rural China. Gastroenterology 2021, 160, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, K.; Akyirem, S.; Sartori, L.; Abdul-Latif, A.M.; Akaateba, D.; Bayrampour, H.; Daly, A.; Hadfield, K.; Abiiro, G.A. Measurement of pregnancy-related anxiety worldwide: A systematic review. BMC Pregnancy Childbirth 2022, 22, 331. [Google Scholar] [CrossRef] [PubMed]
- Brunton, R.J.; Dryer, R.; Saliba, A.; Kohlhoff, J. Pregnancy anxiety: A systematic review of current scales. J. Affect. Disord. 2015, 176, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Adler, C.; du Plessis, M.R.; Landau, E.R.; Dashper, S.G.; Reynolds, E.C.; Schwartz, O.S.; Simmons, J.G. Oral microbiome composition, but not diversity, is associated with adolescent anxiety and depression symptoms. Physiol. Behav. 2020, 226, 113126. [Google Scholar] [CrossRef]
- Ritchie, G.; Strodl, E.; Parham, S.; Bambling, M.; Cramb, S.; Vitetta, L. An exploratory study of the gut microbiota in major depression with anxious distress. J. Affect. Disord. 2023, 320, 595–604. [Google Scholar] [CrossRef]
- Xiong, R.G.; Li, J.H.; Cheng, J.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Gan, R.Y.; Li, H.B. The Role of Gut Microbiota in Anxiety, Depression, and Other Mental Disorders as Well as the Protective Effects of Dietary Components. Nutrients 2023, 15, 3258. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Level | P1 | P2 | P3 | Overall | ||||
|---|---|---|---|---|---|---|---|---|---|
| N | Percentage | N | Percentage | N | Percentage | N | Percentage | ||
| Age * | Median (IQR) | 29 (5.75) | 29 (6) | 29 (6) | 29 (5.5) | ||||
| Pre-Pregnancy BMI | Underweight | 5 | 6.4% | 5 | 6.5% | 4 | 5.7% | 5 | 5.0% |
| Normal | 47 | 60.3% | 45 | 58.4% | 10 | 14.3% | 52 | 52.0% | |
| Overweight | 15 | 19.2% | 16 | 20.8% | 42 | 60.0% | 17 | 17.0% | |
| Obesity | 11 | 14.1% | 11 | 14.3% | 14 | 20.0% | 13 | 13.0% | |
| Parity | 1 | 47 | 60.3% | 50 | 64.9% | 44 | 62.9% | 53 | 53.0% |
| 2 | 31 | 39.7% | 27 | 35.1% | 26 | 37.1% | 34 | 34.0% | |
| Smoking | Yes | 2 | 2.6% | 2 | 2.6% | 1 | 1.4% | 2 | 2.0% |
| No | 76 | 97.4% | 75 | 97.4% | 69 | 98.6% | 85 | 85.0% | |
| Alcohol | Yes | 15 | 19.2% | 15 | 19.5% | 13 | 18.6% | 17 | 17.0% |
| No | 63 | 80.8% | 62 | 80.5% | 57 | 81.4% | 70 | 70.0% | |
| Energy Intake * | Median (IQR) | 1279 (992) | 1417 (961) | 1730 (1257) | 1434 (1019) | ||||
| Physical Activity Level | High | 18 | 23.1% | 17 | 22.1% | 5 | 7.1% | 40 | 17.8% |
| Median | 36 | 46.2% | 34 | 44.2% | 32 | 47.1% | 103 | 45.8% | |
| Low | 24 | 30.8% | 26 | 33.8% | 32 | 45.7% | 82 | 36.4% | |
| Probiotics | Yes | 3 | 3.8% | 4 | 5.2% | 2 | 2.9% | 9 | 4.0% |
| No | 75 | 96.2% | 73 | 94.8% | 68 | 97.1% | 216 | 96.0% | |
| Prebiotics | Yes | 1 | 1.3% | 2 | 2.6% | 1 | 1.4% | 4 | 1.8% |
| No | 77 | 98.7% | 75 | 97.4% | 69 | 98.6% | 221 | 98.2% | |
| Total | 78 | 77 | 70 | 87 | |||||
| P1 | P2 | P3 | χ2 * | p | |
|---|---|---|---|---|---|
| Anxiety Score | 41.3 (11.0) | 38.8 (13.8) | 40.0 (10.0) | 4.607 | 0.100 |
| Depression Score | 47.5 (15.6) | 46.3 (15.0) | 43.8 (14.7) | 3.704 | 0.157 |
| Group | P1 | P2 | P3 | Overall | χ2 * | p | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Percentage | N | Percentage | N | Percentage | N | Percentage | |||
| Anxiety Group | 16 | 20.5 | 12 | 15.6 | 15 | 21.4 | 43 | 19.1 | 0.960 | 0.600 |
| Non-Anxiety Group | 62 | 79.5 | 65 | 84.4 | 55 | 78.6 | 182 | 80.9 | ||
| Depression Group | 25 | 32.1 | 17 | 22.1 | 16 | 22.9 | 58 | 25.8 | 2.500 | 0.300 |
| Non-Depression Group | 53 | 67.9 | 60 | 77.9 | 54 | 77.1 | 167 | 74.2 | ||
| Variable | Model | Metrics for Alpha Diversity | Estimate | Std. Error | df | t | p |
|---|---|---|---|---|---|---|---|
| Anxiety Score | Unadjusted | ACE | −5.454 | 4.516 | 219.260 | −1.208 | 0.229 |
| Anxiety Score | Adjusted | ACE | −7.137 | 4.660 | 207.118 | −1.532 | 0.127 |
| Anxiety Score | Unadjusted | Chao1 | −5.460 | 4.514 | 219.187 | −1.210 | 0.228 |
| Anxiety Score | Adjusted | Chao1 | −7.149 | 4.657 | 206.977 | −1.535 | 0.126 |
| Anxiety Score | Unadjusted | Shannon Entropy | −0.009 | 0.005 | 218.078 | −2.025 | 0.044 |
| Anxiety Score | Adjusted | Shannon Entropy | −0.011 | 0.005 | 207.035 | −2.234 | 0.027 |
| Anxiety Score | Unadjusted | Simpson Index | −0.001 | 0.001 | 215.991 | −1.787 | 0.075 |
| Anxiety Score | Adjusted | Simpson Index | −0.001 | 0.001 | 205.413 | −1.984 | 0.049 |
| Depression Score | Unadjusted | ACE | 1.149 | 3.737 | 220.385 | 0.308 | 0.759 |
| Depression Score | Adjusted | ACE | −0.838 | 3.833 | 210.214 | −0.219 | 0.827 |
| Depression Score | Unadjusted | Chao1 | 1.137 | 3.735 | 220.343 | 0.304 | 0.761 |
| Depression Score | Adjusted | Chao1 | −0.855 | 3.830 | 210.132 | −0.223 | 0.824 |
| Depression Score | Unadjusted | Shannon Entropy | −0.002 | 0.004 | 219.456 | −0.519 | 0.604 |
| Depression Score | Adjusted | Shannon Entropy | −0.004 | 0.004 | 209.868 | −0.910 | 0.364 |
| Depression Score | Unadjusted | Simpson Index | <0.001 | <0.001 | 217.873 | −0.884 | 0.377 |
| Depression Score | Adjusted | Simpson Index | −0.001 | 0.001 | 208.589 | −1.097 | 0.274 |
| Variable | Model | Metrics for Alpha Diversity | Estimate | Std. Error | df | t | p |
|---|---|---|---|---|---|---|---|
| Group_Anxiety | Unadjusted | ACE | −219.618 | 110.415 | 219.382 | −1.989 | 0.048 |
| Group_Anxiety | Adjusted | ACE | −247.762 | 112.065 | 211.760 | −2.211 | 0.028 |
| Group_Anxiety | Unadjusted | Chao1 | −219.358 | 110.368 | 219.445 | −1.988 | 0.048 |
| Group_Anxiety | Adjusted | Chao1 | −247.512 | 112.004 | 211.789 | −2.210 | 0.028 |
| Group_Anxiety | Unadjusted | Shannon Entropy | −0.149 | 0.116 | 220.490 | −1.288 | 0.199 |
| Group_Anxiety | Adjusted | Shannon Entropy | −0.168 | 0.119 | 211.931 | −1.407 | 0.161 |
| Group_Anxiety | Unadjusted | Simpson Index | −0.015 | 0.015 | 220.991 | −1.032 | 0.303 |
| Group_Anxiety | Adjusted | Simpson Index | −0.017 | 0.015 | 211.964 | −1.147 | 0.253 |
| Group_Depression | Unadjusted | ACE | −189.147 | 102.327 | 220.990 | −1.848 | 0.066 |
| Group_Depression | Adjusted | ACE | −218.775 | 103.806 | 211.260 | −2.108 | 0.036 |
| Group_Depression | Unadjusted | Chao1 | −189.116 | 102.274 | 220.985 | −1.849 | 0.066 |
| Group_Depression | Adjusted | Chao1 | −218.818 | 103.737 | 211.205 | −2.109 | 0.036 |
| Group_Depression | Unadjusted | Shannon Entropy | −0.233 | 0.106 | 220.889 | −2.193 | 0.029 |
| Group_Depression | Adjusted | Shannon Entropy | −0.270 | 0.110 | 211.444 | −2.467 | 0.014 |
| Group_Depression | Unadjusted | Simpson Index | −0.027 | 0.013 | 220.300 | −2.049 | 0.042 |
| Group_Depression | Adjusted | Simpson Index | −0.031 | 0.014 | 210.718 | −2.229 | 0.027 |
| Variable | R2 | F | p |
|---|---|---|---|
| Anxiety Score | 0.723% | 3.115 | 0.001 |
| Depression Score | 0.731% | 3.150 | 0.001 |
| Group_Anxiety | 0.651% | 2.808 | 0.001 |
| Group_Depression | 0.810% | 3.497 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chi, R.; Li, M.; Zhang, M.; Zhang, N.; Zhang, G.; Cui, L.; Ma, G. Exploring the Association between Anxiety, Depression, and Gut Microbiota during Pregnancy: Findings from a Pregnancy Cohort Study in Shijiazhuang, Hebei Province, China. Nutrients 2024, 16, 1460. https://doi.org/10.3390/nu16101460
Chi R, Li M, Zhang M, Zhang N, Zhang G, Cui L, Ma G. Exploring the Association between Anxiety, Depression, and Gut Microbiota during Pregnancy: Findings from a Pregnancy Cohort Study in Shijiazhuang, Hebei Province, China. Nutrients. 2024; 16(10):1460. https://doi.org/10.3390/nu16101460
Chicago/Turabian StyleChi, Ruixin, Muxia Li, Man Zhang, Na Zhang, Guohua Zhang, Lijun Cui, and Guansheng Ma. 2024. "Exploring the Association between Anxiety, Depression, and Gut Microbiota during Pregnancy: Findings from a Pregnancy Cohort Study in Shijiazhuang, Hebei Province, China" Nutrients 16, no. 10: 1460. https://doi.org/10.3390/nu16101460
APA StyleChi, R., Li, M., Zhang, M., Zhang, N., Zhang, G., Cui, L., & Ma, G. (2024). Exploring the Association between Anxiety, Depression, and Gut Microbiota during Pregnancy: Findings from a Pregnancy Cohort Study in Shijiazhuang, Hebei Province, China. Nutrients, 16(10), 1460. https://doi.org/10.3390/nu16101460