
Maternal Weight Intervention in the Perinatal Period Improves Liver Health in the Offspring of Mothers with Obesity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
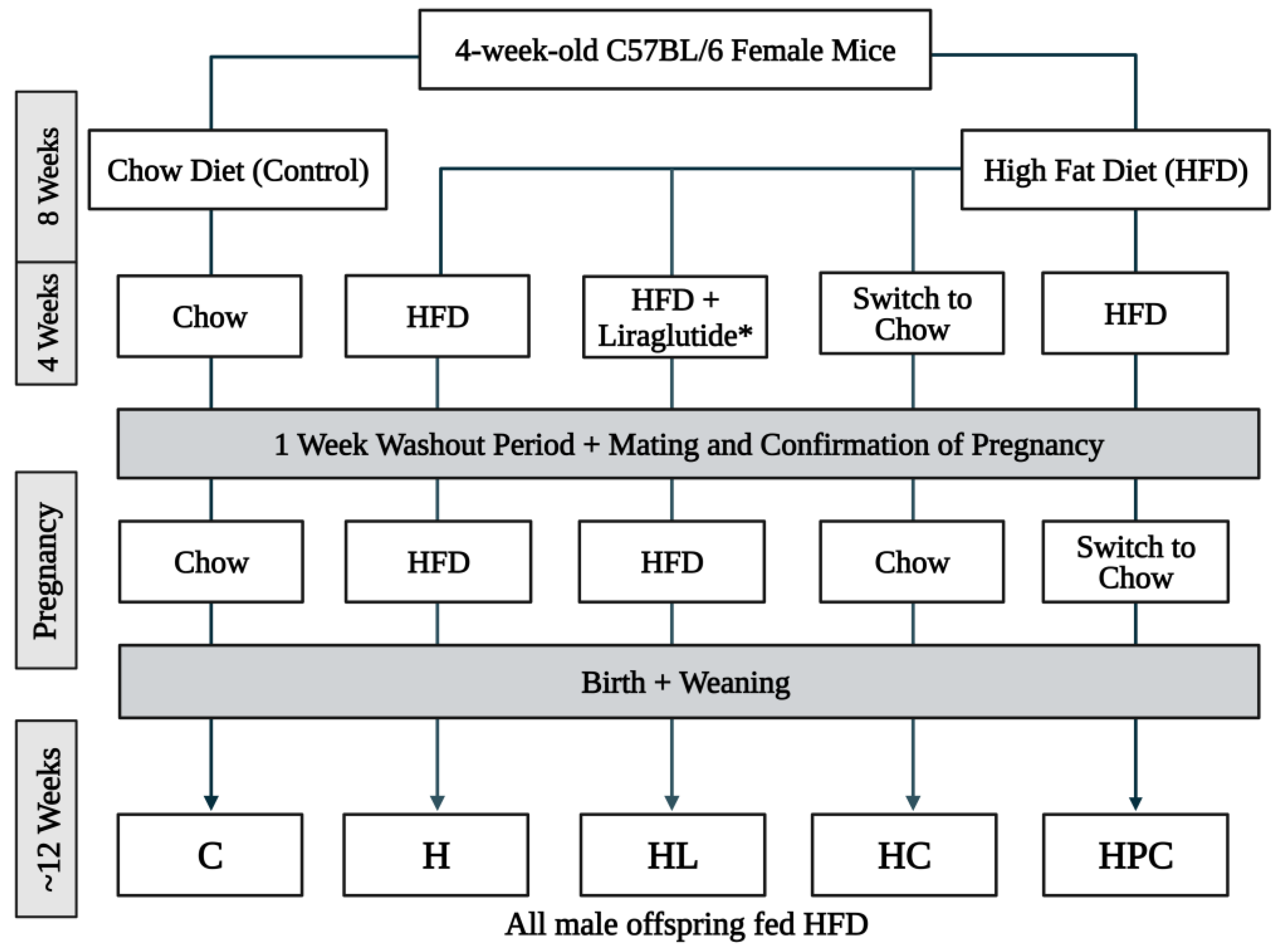
2.1. Animal Experiments
2.2. Bioassays
2.3. Quantitative Real-Time PCR
2.4. Analysis of Structural Changes to the Liver
2.5. Immunohistochemistry
2.6. Statistical Methods
3. Results
3.1. Maternal HFD Causes Metabolic Disturbances in the Offspring at Postnatal Week 12
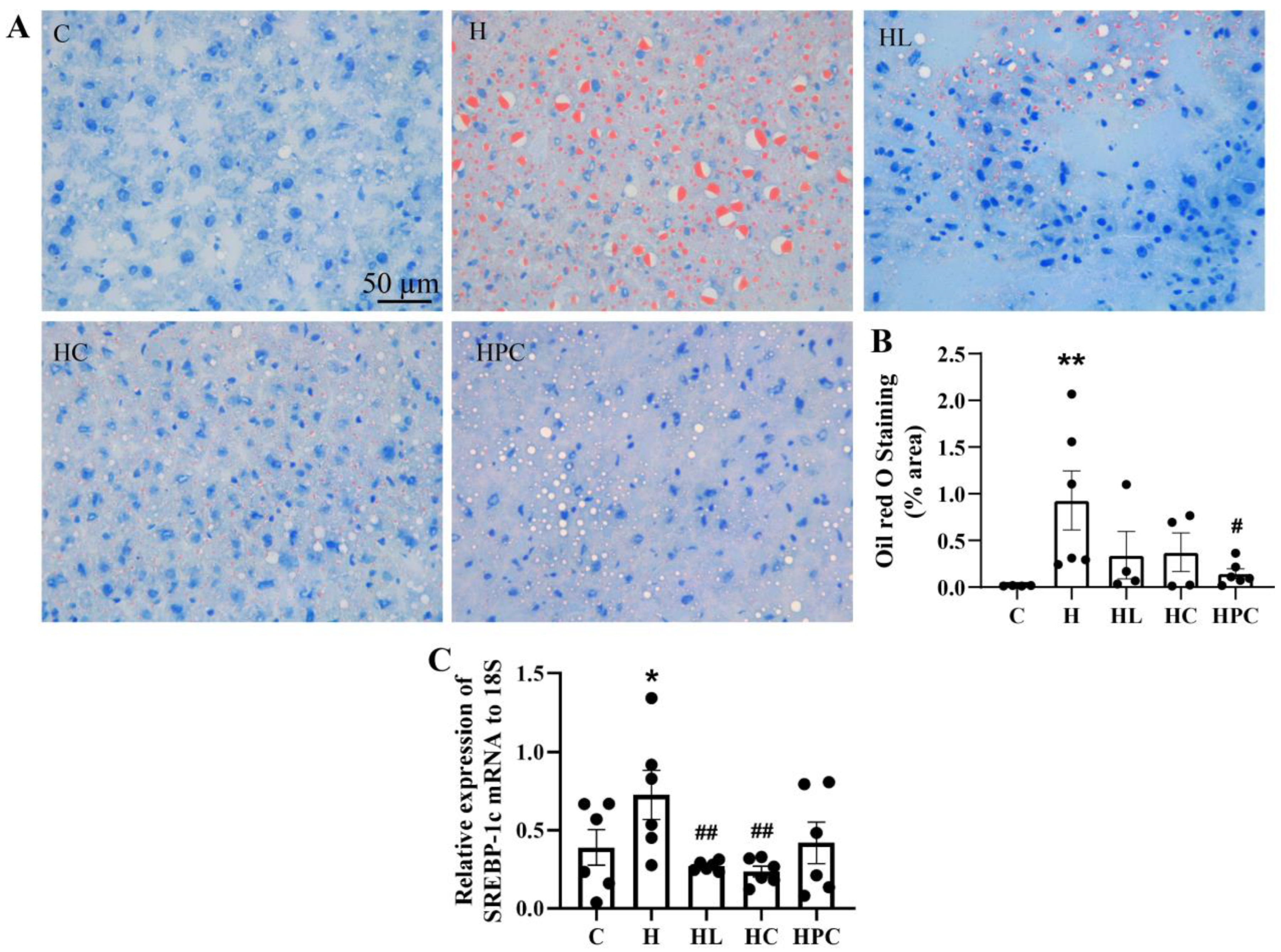
3.2. Maternal Obesity Induces Steatosis in the Liver of Offspring
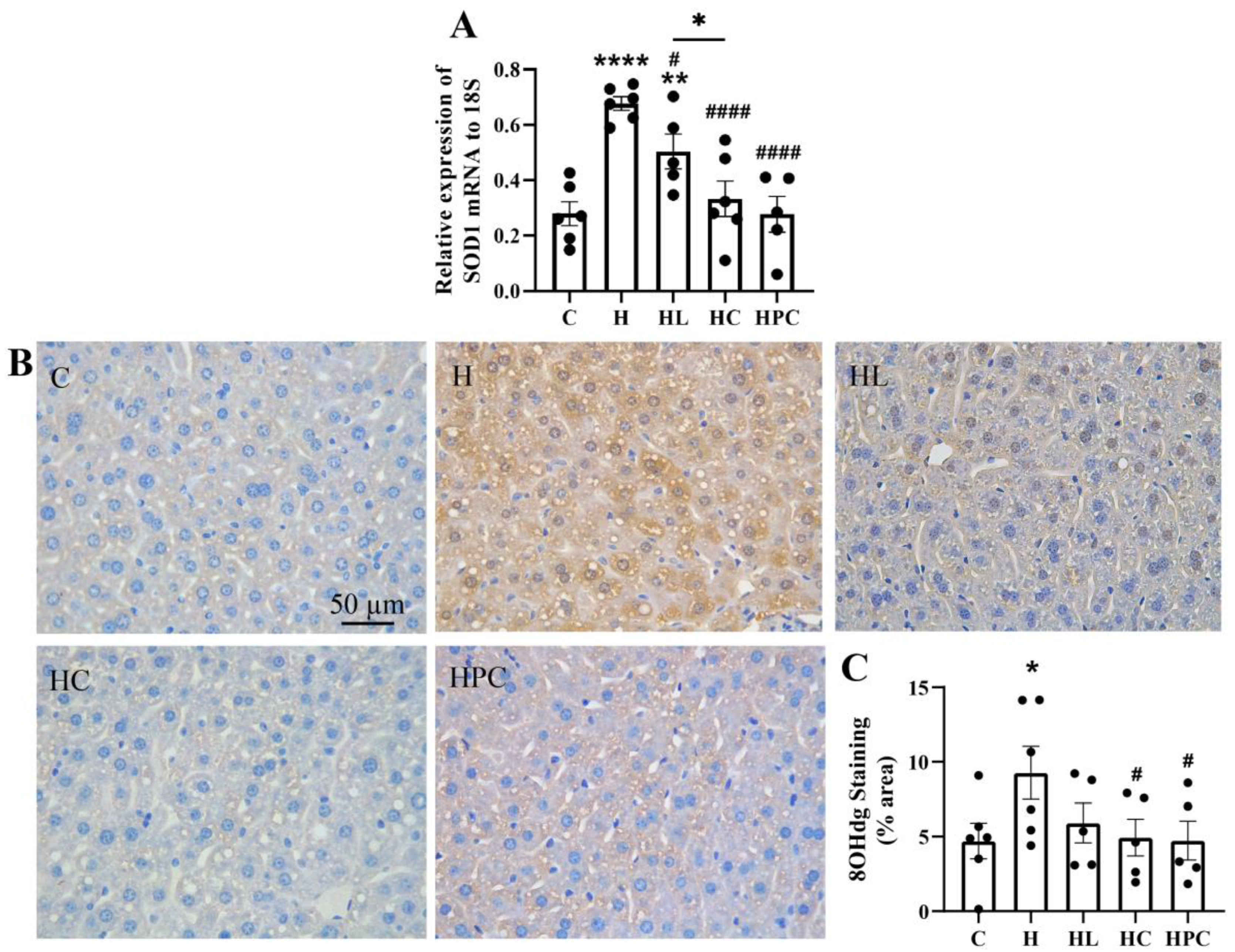
3.3. Maternal Obesity Increases Inflammation and Oxidative Stress in the Liver of Offspring Which Is Attenuated by Weight Loss in the Perinatal Period
3.4. Fibrotic Levels in the Liver of Offspring Are Increased Due to Maternal Obesity and Restored by Weight Loss in the Perinatal Period
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef] [PubMed]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment. Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Bellentani, S.; Saccoccio, G.; Masutti, F.; Crocè, L.S.; Brandi, G.; Sasso, F.; Cristanini, G.; Tiribelli, C. Prevalence of and Risk Factors for Hepatic Steatosis in Northern Italy. Ann. Intern. Med. 2000, 132, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Hoyumpa, A.M., Jr.; Greene, H.L.; Dunn, G.D.; Schenker, S. Fatty liver: Biochemical and clinical considerations. Am. J. Dig. Dis. 1975, 20, 1142–1170. [Google Scholar] [CrossRef]
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of Childhood and Adult Obesity in the United States, 2011–2012. JAMA 2014, 311, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, H.M.; Mercer, B.M.; Catalano, P.M. The influence of obesity and diabetes on the prevalence of macrosomia. Am. J. Obstet. Gynecol. 2004, 191, 964–968. [Google Scholar] [CrossRef]
- Heslehurst, N.; Rankin, J.; Wilkinson, J.R.; Summerbell, C.D. A nationally representative study of maternal obesity in England, UK: Trends in incidence and demographic inequalities in 619 323 births, 1989–2007. Int. J. Obes. 2010, 34, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Nohr, E.A.; Vaeth, M.; Baker, J.L.; Sørensen, T.I.A.; Olsen, J.; Rasmussen, K.M. Combined associations of prepregnancy body mass index and gestational weight gain with the outcome of pregnancy. Am. J. Clin. Nutr. 2008, 87, 1750–1759. [Google Scholar] [CrossRef]
- Hagström, H.; Simon, T.G.; Roelstraete, B.; Stephansson, O.; Söderling, J.; Ludvigsson, J.F. Maternal obesity increases the risk and severity of NAFLD in offspring. J. Hepatol. 2021, 75, 1042–1048. [Google Scholar] [CrossRef]
- McCurdy, C.E.; Bishop, J.M.; Williams, S.M.; Grayson, B.E.; Smith, M.S.; Friedman, J.E.; Grove, K.L. Maternal high-fat diet triggers lipotoxicity in the fetal livers of nonhuman primates. J. Clin. Investig. 2009, 119, 323–335. [Google Scholar] [CrossRef]
- Bruce, K.D.; Cagampang, F.R.; Argenton, M.; Zhang, J.; Ethirajan, P.L.; Burdge, G.C.; Bateman, A.C.; Clough, G.F.; Poston, L.; Hanson, M.A.; et al. Maternal high-fat feeding primes steatohepatitis in adult mice offspring, involving mitochondrial dysfunction and altered lipogenesis gene expression. Hepatology 2009, 50, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Kominiarek, M.A.; Chauhan, S.P. Obesity Before, During, and After Pregnancy: A Review and Comparison of Five National Guidelines. Am. J. Perinatol. 2016, 33, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Purcell, K.; Sumithran, P.; Prendergast, L.A.; Bouniu, C.J.; Delbridge, E.; Proietto, J. The effect of rate of weight loss on long-term weight management: A randomised controlled trial. Lancet Diabetes Endocrinol. 2014, 2, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D. The structure and function of the glucagon-like peptide-1 receptor and its ligands. Br. J. Pharmacol. 2012, 166, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, N.; Chen, H.; Pollock, C.A.; Glastras, S.J. Preconception weight loss improves fertility and maternal outcomes in obese mice. J. Endocrinol. 2022, 253, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Tajaddini, A.; Kendig, M.D.; Prates, K.V.; Westbrook, R.F.; Morris, M.J. Male Rat Offspring Are More Impacted by Maternal Obesity Induced by Cafeteria Diet than Females-Additive Effect of Postweaning Diet. Int. J. Mol. Sci. 2022, 23, 1442. [Google Scholar] [CrossRef] [PubMed]
- Meakin, A.S.; Nathanielsz, P.W.; Li, C.; Clifton, V.L.; Wiese, M.D.; Morrison, J.L. Maternal obesity impacts fetal liver androgen signalling in a sex-specific manner. Life Sci. 2023, 122344. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.; Helguero, L.A.; González-Granillo, M.; Melo, T.; Couto, D.; Angelin, B.; Domingues, M.R.; Li, X.; Kutter, C.; Korach-André, M. Molecular programming modulates hepatic lipid metabolism and adult metabolic risk in the offspring of obese mothers in a sex-specific manner. Commun. Biol. 2022, 5, 1057. [Google Scholar] [CrossRef]
- Oben, J.A.; Mouralidarane, A.; Samuelsson, A.-M.; Matthews, P.J.; Morgan, M.L.; McKee, C.; Soeda, J.; Fernandez-Twinn, D.S.; Martin-Gronert, M.S.; Ozanne, S.E.; et al. Maternal obesity during pregnancy and lactation programs the development of offspring non-alcoholic fatty liver disease in mice. J. Hepatol. 2010, 52, 913–920. [Google Scholar] [CrossRef]
- Koo, S.-H.; Dutcher, A.K.; Towle, H.C. Glucose and Insulin Function through Two Distinct Transcription Factors to Stimulate Expression of Lipogenic Enzyme Genes in Liver*. J. Biol. Chem. 2001, 276, 9437–9445. [Google Scholar] [CrossRef]
- Shimano, H.; Horton, J.D.; Hammer, R.E.; Shimomura, I.; Brown, M.S.; Goldstein, J.L. Overproduction of cholesterol and fatty acids causes massive liver enlargement in transgenic mice expressing truncated SREBP-1a. J. Clin. Investig. 1996, 98, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Tajima, K.; Zolzaya, K.; Sato, K.; Inoue, R.; Yoneda, M.; Fujita, K.; Nozaki, Y.; Kubota, K.C.; Haga, H.; et al. Protection from non-alcoholic steatohepatitis and liver tumourigenesis in high fat-fed insulin receptor substrate-1-knockout mice despite insulin resistance. Diabetologia 2012, 55, 3382–3391. [Google Scholar] [CrossRef] [PubMed]
- Nio, Y.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Funata, M.; Yamaguchi, M.; Ueki, K.; Kadowaki, T. Monocyte chemoattractant protein-1 (MCP-1) deficiency enhances alternatively activated M2 macrophages and ameliorates insulin resistance and fatty liver in lipoatrophic diabetic A-ZIP transgenic mice. Diabetologia 2012, 55, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Malti, N.; Merzouk, H.; Merzouk, S.A.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative stress and maternal obesity: Feto-placental unit interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, S.R.; Mulligan, C.M.; Janssen, R.C.; Baker, P.R., 2nd; Bergman, B.C.; D’Alessandro, A.; Nemkov, T.; Maclean, K.N.; Jiang, H.; Dean, T.A.; et al. Switching obese mothers to a healthy diet improves fetal hypoxemia, hepatic metabolites, and lipotoxicity in non-human primates. Mol. Metab. 2018, 18, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Ishiba, H.; Sumida, Y.; Seko, Y.; Tanaka, S.; Yoneda, M.; Hyogo, H.; Ono, M.; Fujii, H.; Eguchi, Y.; Suzuki, Y.; et al. Type IV Collagen 7S Is the Most Accurate Test For Identifying Advanced Fibrosis in NAFLD With Type 2 Diabetes. Hepatol. Commun. 2021, 5, 559–572. [Google Scholar] [CrossRef]
- Ren, J.; Wang, X.; Yee, C.; Gorrell, M.D.; McLennan, S.V.; Twigg, S.M. Sitagliptin Is More Effective Than Gliclazide in Preventing Pro-Fibrotic and Pro-Inflammatory Changes in a Rodent Model of Diet-Induced Non-Alcoholic Fatty Liver Disease. Molecules 2022, 27, 727. [Google Scholar] [CrossRef]
- Hall, P.; Cash, J. What is the real function of the liver ‘function’ tests? Ulster. Med. J. 2012, 81, 30–36. [Google Scholar]
- Bouanane, S.; Merzouk, H.; Benkalfat, N.B.; Soulimane, N.; Merzouk, S.A.; Gresti, J.; Tessier, C.; Narce, M. Hepatic and very low-density lipoprotein fatty acids in obese offspring of overfed dams. Metabolism 2010, 59, 1701–1709. [Google Scholar] [CrossRef]
- MacQueen, H.A.; Sadler, D.A.; Moore, S.A.; Daya, S.; Brown, J.Y.; Shuker, D.E.G.; Seaman, M.; Wassif, W.S. Deleterious effects of a cafeteria diet on the livers of nonobese rats. Nutr. Res. 2007, 27, 38–47. [Google Scholar] [CrossRef]
- Xu, H.; Fu, Q.; Zhou, Y.; Xue, C.; Olson, P.; Lynch, E.C.; Zhang, K.K.; Wu, C.; Murano, P.; Zhang, L.; et al. A long-term maternal diet intervention is necessary to avoid the obesogenic effect of maternal high-fat diet in the offspring. J. Nutr. Biochem. 2018, 62, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Martínez-Samayoa, P.M.; Rodríguez-González, G.L.; Nathanielsz, P.W. Dietary intervention prior to pregnancy reverses metabolic programming in male offspring of obese rats. J. Physiol. 2010, 588, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Peterson, G.E.; Pollom, R.D. Liraglutide in clinical practice: Dosing, safety and efficacy. Int. J. Clin. Pract. 2010, 64, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, H.; Yang, L.; Zhao, M.; Magnussen, C.G.; Xi, B. Associations Between Gestational Weight Gain and Adverse Birth Outcomes: A Population-Based Retrospective Cohort Study of 9 Million Mother-Infant Pairs. Front. Nutr. 2022, 9, 811217. [Google Scholar] [CrossRef]
- Poston, L.; Bell, R.; Croker, H.; Flynn, A.C.; Godfrey, K.M.; Goff, L.; Hayes, L.; Khazaezadeh, N.; Nelson, S.M.; Oteng-Ntim, E.; et al. Effect of a behavioural intervention in obese pregnant women (the UPBEAT study): A multicentre, randomised controlled trial. Lancet Diabetes Endocrinol. 2015, 3, 767–777. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| 18S | CGGCTACCACATACCAAGGAA | GCTGGAATTAACCGCGGCT |
| COL-I | CCCCGGGACTCCTGGACTT | GCTCCGACACGCCCTCTCTC |
| COL-III | CCTGGAGCCCCTGGACTAATAG | GCCCATTGCACCAGGTTCT |
| COL-IV | ATGGCTTGCCTGGAGAGATAGG | TGGTTGCCGTTTGAGTCCTGGA |
| MCP-1 | AGGTCCCTGTCATGCTTCTG | GCTGCTGGTGATCCTCTTGT |
| SOD1 | GGTGAACCAGTTGTGTTGTCAG | ATGAGGTCCTGCACTGGTACAG |
| SREBP-1c | CATGGATTGCACATTTGAAGAC | GCAGGAGAAGAGAAGCTCTCAG |
| Measure | C | H | HL | HC | HPC |
|---|---|---|---|---|---|
| Body weight (g) | 27.53 ± 0.66 | 34.45 ± 0.55 **** | 34.52 ± 0.83 ****,†††† | 30.62 ± 0.29 **,### | 32.93 ± 0.50 **** |
| Liver mass (% total body weight) | 4.11 ± 0.19 | 4.04 ± 0.18 | 3.84 ± 0.20 | 4.03 ± 0.17 | 3.89 ± 0.15 |
| Insulin (pmol/L) | 1.03 ± 0.22 | 1.92 ± 0.31 ** | 1.05 ± 0.12 ## | 0.85 ± 0.11 ### | 0.96 ± 0.12 ## |
| HOMA-IR score | 0.072 ± 0.01 | 0.148 ± 0.02 *** | 0.080 ± 0.01 ## | 0.053 ± 0.01 #### | 0.065 ± 0.01 ### |
| Glucose (mmol/L) | 10.77 ± 0.60 | 11.75 ± 0.41 | 12.37 ± 0.40 | 11.02 ± 0.68 | 11.27 ± 0.94 |
| NEFA (mmol/L) | 0.42 ± 0.03 | 0.47 ± 0.03 | 0.48 ± 0.05 | 0.42 ± 0.04 | 0.37 ± 0.02 |
| AST (U/L) | 96.46 ± 11.95 | 107.40 ± 17.20 | 117.80 ± 17.02 | 121.10 ± 14.26 | 106.9 ± 18.68 |
| ALT (U/L) | 15.62 ± 2.48 | 16.92 ± 2.61 | 19.36 ± 2.06 | 22.10 ± 5.27 | 20.98 ± 3.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purcell, A.R.; Rodrigo, N.; Cao, Q.; Joseph, O.; Gill, A.J.; Saad, S.; Pollock, C.A.; Glastras, S.J. Maternal Weight Intervention in the Perinatal Period Improves Liver Health in the Offspring of Mothers with Obesity. Nutrients 2024, 16, 109. https://doi.org/10.3390/nu16010109
Purcell AR, Rodrigo N, Cao Q, Joseph O, Gill AJ, Saad S, Pollock CA, Glastras SJ. Maternal Weight Intervention in the Perinatal Period Improves Liver Health in the Offspring of Mothers with Obesity. Nutrients. 2024; 16(1):109. https://doi.org/10.3390/nu16010109
Chicago/Turabian StylePurcell, Amanda R., Natassia Rodrigo, Qinghua Cao, Olivia Joseph, Anthony J. Gill, Sonia Saad, Carol A. Pollock, and Sarah J. Glastras. 2024. "Maternal Weight Intervention in the Perinatal Period Improves Liver Health in the Offspring of Mothers with Obesity" Nutrients 16, no. 1: 109. https://doi.org/10.3390/nu16010109
APA StylePurcell, A. R., Rodrigo, N., Cao, Q., Joseph, O., Gill, A. J., Saad, S., Pollock, C. A., & Glastras, S. J. (2024). Maternal Weight Intervention in the Perinatal Period Improves Liver Health in the Offspring of Mothers with Obesity. Nutrients, 16(1), 109. https://doi.org/10.3390/nu16010109