

Late, but Not Early, Night Sleep Loss Compromises Neuroendocrine Appetite Regulation and the Desire for Food

 ,
, 

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Study Design and Procedure
2.3. Feelings of Hunger and Appetite
2.4. Blood Samples and Assays
2.5. Statistical Analyses
3. Results
3.1. Sleep
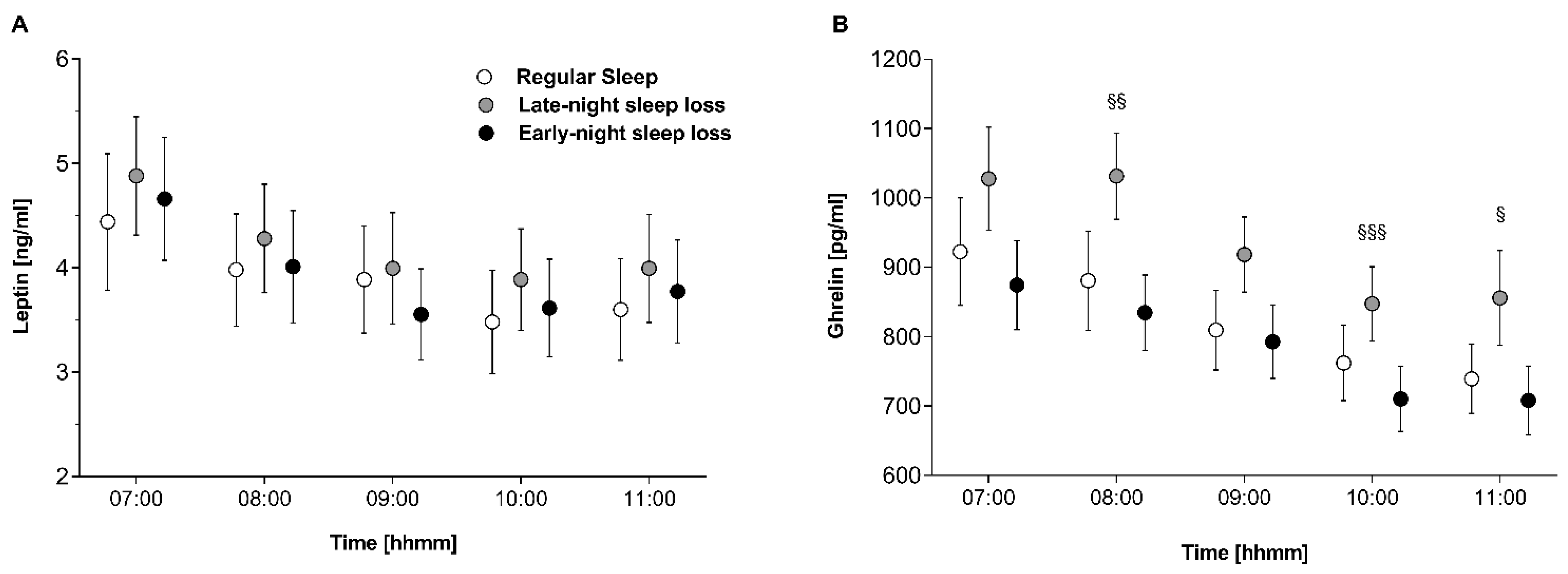
3.2. Orexigenic/Anorexigenic Balance
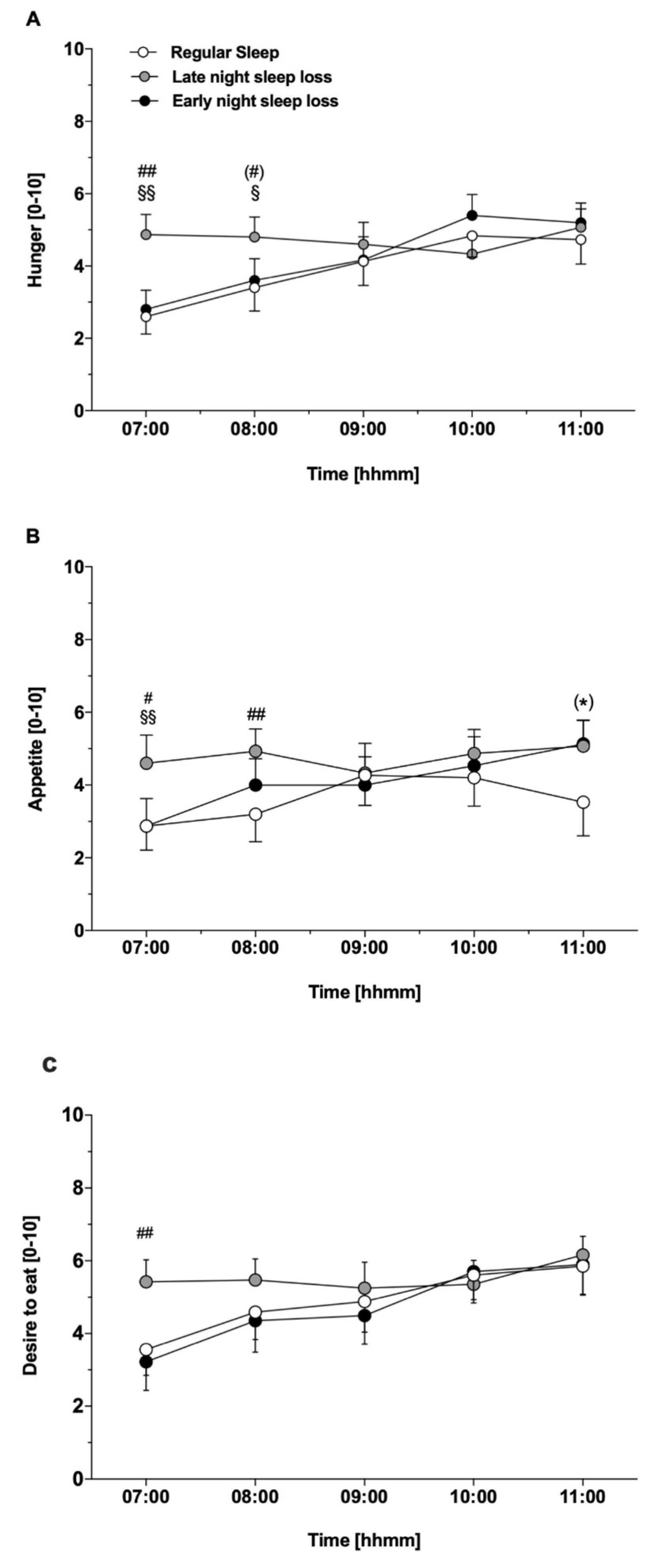
3.3. Feelings of Hunger and Appetite
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ford, E.S.; Cunningham, T.J.; Croft, J.B. Trends in Self-Reported Sleep Duration among US Adults from 1985 to 2012. Sleep 2015, 38, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, R.P.; Patel, S. The epidemiology of sleep and obesity. Sleep Health 2017, 3, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Cappuccio, F.P.; Taggart, F.M.; Kandala, N.-B.; Currie, A.; Peile, E.; Stranges, S.; Miller, M.A. Meta-Analysis of Short Sleep Duration and Obesity in Children and Adults. Sleep 2008, 31, 619–626. [Google Scholar] [CrossRef]
- Xie, J.; Li, Y.; Zhang, Y.; Vgontzas, A.N.; Basta, M.; Chen, B.; Xu, C.; Tang, X. Sleep duration and metabolic syndrome: An updated systematic review and meta-analysis. Sleep Med. Rev. 2021, 59, 101451. [Google Scholar] [CrossRef]
- Che, T.; Yan, C.; Tian, D.; Zhang, X.; Liu, X.; Wu, Z. The Association between Sleep and Metabolic Syndrome: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2021, 12, 773646. [Google Scholar] [CrossRef]
- Schmid, S.M.; Hallschmid, M.; Schultes, B. The metabolic burden of sleep loss. Lancet Diabetes Endocrinol. 2015, 3, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.N.; Oster, H. How sleep and wakefulness influence circadian rhythmicity: Effects of insufficient and mistimed sleep on the animal and human transcriptome. J. Sleep Res. 2015, 24, 476–493. [Google Scholar] [CrossRef]
- Koopman, A.D.; Rauh, S.P.; Riet, E.V.T.; Groeneveld, L.; Van Der Heijden, A.A.; Elders, P.J.; Dekker, J.M.; Nijpels, G.; Beulens, J.W.; Rutters, F. The Association between Social Jetlag, the Metabolic Syndrome, and Type 2 Diabetes Mellitus in the General Population: The New Hoorn Study. J. Biol. Rhythm. 2017, 32, 359–368. [Google Scholar] [CrossRef]
- Hulsegge, G.; Loef, B.; van Kerkhof, L.W.; Roenneberg, T.; van der Beek, A.J.; Proper, K.I. Shift work, sleep disturbances and social jetlag in healthcare workers. J. Sleep Res. 2019, 28, e12802. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Dinich, J.; Merrow, M.; Roenneberg, T. Social Jetlag: Misalignment of Biological and Social Time. Chronobiol. Int. 2006, 23, 497–509. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, L.; Zhang, Y.; Zhang, B.; He, Y.; Xie, S.; Li, M.; Miao, X.; Chan, E.Y.Y.; Tang, J.L.; et al. Meta-analysis on night shift work and risk of metabolic syndrome. Obes. Rev. 2014, 15, 709–720. [Google Scholar] [CrossRef]
- Bosy-Westphal, A.; Hinrichs, S.; Jauch-Chara, K.; Hitze, B.; Later, W.; Wilms, B.; Settler, U.; Peters, A.; Kiosz, D.; Müller, M.J. Influence of Partial Sleep Deprivation on Energy Balance and Insulin Sensitivity in Healthy Women. Obes. Facts 2008, 1, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Brondel, L.; Romer, M.A.; Nougues, P.M.; Touyarou, P.; Davenne, D. Acute partial sleep deprivation increases food intake in healthy men. Am. J. Clin. Nutr. 2010, 91, 1550–1559. [Google Scholar] [CrossRef]
- Schmid, S.M.; Hallschmid, M.; Jauch-Chara, K.; Wilms, B.; Benedict, C.; Lehnert, H.; Born, J.; Schultes, B. Short-term sleep loss decreases physical activity under free-living conditions but does not increase food intake under time-deprived laboratory conditions in healthy men. Am. J. Clin. Nutr. 2009, 90, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Calvin, A.D.; Carter, R.E.; Adachi, T.; Macedo, P.G.; Albuquerque, F.N.; van der Walt, C.; Bukartyk, J.; Davison, D.E.; Levine, J.A.; Somers, V.K. Effects of Experimental Sleep Restriction on Caloric Intake and Activity Energy Expenditure. Chest 2013, 144, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Broussard, J.L.; Kilkus, J.M.; Delebecque, F.; Abraham, V.; Day, A.; Whitmore, H.R.; Tasali, E. Elevated ghrelin predicts food intake during experimental sleep restriction. Obesity 2016, 24, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Radcliffe, P.N.; Whitney, C.C.; Fagnant, H.S.; Wilson, M.A.; Finlayson, G.; Smith, T.J.; Karl, J.P. Severe sleep restriction suppresses appetite independent of effects on appetite regulating hormones in healthy young men without obesity. Physiol. Behav. 2021, 237, 113438. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Shi, C.; Park, C.G.; Zhao, X.; Reutrakul, S. Effects of sleep restriction on metabolism-related parameters in healthy adults: A comprehensive review and meta-analysis of randomized controlled trials. Sleep Med. Rev. 2019, 45, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, K.; Tasali, E.; Penev, P.; Van Cauter, E. Brief Communication: Sleep Curtailment in Healthy Young Men Is Associated with Decreased Leptin Levels, Elevated Ghrelin Levels, and Increased Hunger and Appetite. Ann. Intern. Med. 2004, 141, 846–850. [Google Scholar] [CrossRef]
- Spiegel, K.; Leproult, R.; L’hermite-Balériaux, M.; Copinschi, G.; Penev, P.D.; Van Cauter, E. Leptin Levels Are Dependent on Sleep Duration: Relationships with Sympathovagal Balance, Carbohydrate Regulation, Cortisol, and Thyrotropin. J. Clin. Endocrinol. Metab. 2004, 89, 5762–5771. [Google Scholar] [CrossRef]
- Omisade, A.; Buxton, O.M.; Rusak, B. Impact of acute sleep restriction on cortisol and leptin levels in young women. Physiol. Behav. 2010, 99, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.C.; Dorrian, J.; Liu, P.Y.; Van Dongen, H.P.A.; Wittert, G.A.; Harmer, L.J.; Banks, S. Impact of Five Nights of Sleep Restriction on Glucose Metabolism, Leptin and Testosterone in Young Adult Men. PLoS ONE 2012, 7, e41218. [Google Scholar] [CrossRef] [PubMed]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc. Natl. Acad. Sci. USA 2013, 110, 5695–5700. [Google Scholar] [CrossRef] [PubMed]
- Hibi, M.; Kubota, C.; Mizuno, T.; Aritake, S.; Mitsui, Y.; Katashima, M.; Uchida, S. Effect of shortened sleep on energy expenditure, core body temperature, and appetite: A human randomised crossover trial. Sci. Rep. 2017, 7, 39640. [Google Scholar] [CrossRef]
- Nedeltcheva, A.V.; Kilkus, J.M.; Imperial, J.; Kasza, K.; Schoeller, D.A.; Penev, P.D. Sleep curtailment is accompanied by increased intake of calories from snacks. Am. J. Clin. Nutr. 2009, 89, 126–133. [Google Scholar] [CrossRef]
- St-Onge, M.-P.; Roberts, A.L.; Chen, J.; Kelleman, M.; O’keeffe, M.; RoyChoudhury, A.; Jones, P.J. Short sleep duration increases energy intakes but does not change energy expenditure in normal-weight individuals. Am. J. Clin. Nutr. 2011, 94, 410–416. [Google Scholar] [CrossRef]
- Guilleminault, C.; Powell, N.B.; Martinez, S.; Kushida, C.; Raffray, T.; Palombini, L.; Philip, P. Preliminary observations on the effects of sleep time in a sleep restriction paradigm. Sleep Med. 2003, 4, 177–184. [Google Scholar] [CrossRef]
- Shechter, A.; O’Keeffe, M.; Roberts, A.L.; Zammit, G.K.; RoyChoudhury, A.; St-Onge, M.-P. Alterations in sleep architecture in response to experimental sleep curtailment are associated with signs of positive energy balance. Am. J. Physiol. Integr. Comp. Physiol. 2012, 303, R883–R889. [Google Scholar] [CrossRef]
- McNeil, J.; Forest, G.; Hintze, L.J.; Brunet, J.-F.; Finlayson, G.; Blundell, J.E.; Doucet, É. The Effects of Partial Sleep Restriction and Altered Sleep Timing on Appetite and Food Reward. Appetite 2017, 109, 48–56. [Google Scholar] [CrossRef]
- McNeil, J.; Doucet, É.; Brunet, J.-F.; Hintze, L.J.; Chaumont, I.; Langlois, É.; Maitland, R.; Riopel, A.; Forest, G. The Effects of Sleep Restriction and Altered Sleep Timing on Energy Intake and Energy Expenditure. Physiol. Behav. 2016, 164, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Wilms, B.; Chamorro, R.; Hallschmid, M.; Trost, D.; Forck, N.; Schultes, B.; Mölle, M.; Sayk, F.; Lehnert, H.; Schmid, S.M. Timing Modulates the Effect of Sleep Loss on Glucose Homeostasis. J. Clin. Endocrinol. Metab. 2019, 104, 2801–2808. [Google Scholar] [CrossRef] [PubMed]
- Wilms, B.; Kuhr, M.; Chamorro, R.; Klinsmann, N.; Spyra, D.; Mölle, M.; Kalscheuer, H.; Schultes, B.; Lehnert, H.; Schmid, S.M. Chronobiological Aspects of Sleep Restriction Modulate Subsequent Spontaneous Physical Activity. Physiol. Behav. 2020, 215, 112795. [Google Scholar] [CrossRef]
- Flint, A.; Raben, A.; Blundell, J.E.; Astrup, A. Reproducibility, Power and Validity of Visual Analogue Scales in Assessment of Appetite Sensations in Single Test Meal Studies. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 38–48. [Google Scholar] [CrossRef]
- Schmid, S.M.; Hallschmid, M.; Jauch-Chara, K.; Born, J.; Schultes, B. A Single Night of Sleep Deprivation Increases Ghrelin Levels and Feelings of Hunger in Normal-Weight Healthy Men. J. Sleep Res. 2008, 17, 331–334. [Google Scholar] [CrossRef]
- Lowe, M.R.; Butryn, M.L. Hedonic Hunger: A New Dimension of Appetite? Physiol. Behav. 2007, 91, 432–439. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting Components of Reward: “Liking”, “Wanting”, and Learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Soussignan, R.; Schaal, B.; Royet, J.-P. Reward for Food Odors: An FMRI Study of Liking and Wanting as a Function of Metabolic State and BMI. Soc. Cogn. Affect. Neurosci. 2015, 10, 561–568. [Google Scholar] [CrossRef]
- Chamorro, R.; Kannenberg, S.; Wilms, B.; Kleinerüschkamp, C.; Meyhöfer, S.; Park, S.Q.; Lehnert, H.; Oster, H.; Meyhöfer, S.M. Meal Timing and Macronutrient Composition Modulate Human Metabolism and Reward-Related Drive to Eat. Nutrients 2022, 14, 562. [Google Scholar] [CrossRef]
- Wong, P.M.; Hasler, B.P.; Kamarck, T.W.; Muldoon, M.F.; Manuck, S.B. Social Jetlag, Chronotype, and Cardiometabolic Risk. J. Clin. Endocrinol. Metab. 2015, 100, 4612–4620. [Google Scholar] [CrossRef]
- Rusu, A.; Ciobanu, D.; Vonica, C.L.; Bala, C.; Mocan, A.; Sima, D.; Inceu, G.; Craciun, A.; Pop, R.M.; Craciun, C.; et al. Chronic Disruption of Circadian Rhythm with Mistimed Sleep and Appetite—An Exploratory Research. Chronobiol. Int. 2021, 38, 807–816. [Google Scholar] [CrossRef]
- Nechifor, R.E.; Ciobanu, D.; Vonica, C.L.; Popita, C.; Roman, G.; Bala, C.; Mocan, A.; Inceu, G.; Craciun, A.; Rusu, A. Social Jetlag and Sleep Deprivation Are Associated with Altered Activity in the Reward-Related Brain Areas: An Exploratory Resting-State FMRI Study. Sleep Med. 2020, 72, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Barclay, J.L.; Husse, J.; Bode, B.; Naujokat, N.; Meyer-Kovac, J.; Schmid, S.M.; Lehnert, H.; Oster, H. Circadian Desynchrony Promotes Metabolic Disruption in a Mouse Model of Shiftwork. PLoS ONE 2012, 7, e37150. [Google Scholar] [CrossRef]
- McHill, A.W.; Hull, J.T.; McMullan, C.J.; Klerman, E.B. Chronic Insufficient Sleep Has a Limited Impact on Circadian Rhythmicity of Subjective Hunger and Awakening Fasted Metabolic Hormones. Front. Endocrinol. 2018, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.J.L.; Morris, C.J.; Shea, S.A. The Internal Circadian Clock Increases Hunger and Appetite in the Evening Independent of Food Intake and Other Behaviors: Body Clock Controls Hunger. Obesity 2013, 21, 421–423. [Google Scholar] [CrossRef]
- St-Onge, M.-P.; McReynolds, A.; Trivedi, Z.B.; Roberts, A.L.; Sy, M.; Hirsch, J. Sleep restriction leads to increased activation of brain regions sensitive to food stimuli. Am. J. Clin. Nutr. 2012, 95, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Simpson, N.S.; Banks, S.; Dinges, D.F. Sleep Restriction Is Associated with Increased Morning Plasma Leptin Concentrations, Especially in Women. Biol. Res. Nurs. 2010, 12, 47–53. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, M.-P.; O’Keeffe, M.; Roberts, A.L.; Roychoudhury, A.; Laferrere, B. Short Sleep Duration, Glucose Dysregulation and Hormonal Regulation of Appetite in Men and Women. Sleep 2012, 35, 1503–1510. [Google Scholar] [CrossRef]
- Spaeth, A.M.; Dinges, D.F.; Goel, N. Sex and race differences in caloric intake during sleep restriction in healthy adults. Am. J. Clin. Nutr. 2014, 100, 559–566. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyhöfer, S.; Chamorro, R.; Hallschmid, M.; Spyra, D.; Klinsmann, N.; Schultes, B.; Lehnert, H.; Meyhöfer, S.M.; Wilms, B. Late, but Not Early, Night Sleep Loss Compromises Neuroendocrine Appetite Regulation and the Desire for Food. Nutrients 2023, 15, 2035. https://doi.org/10.3390/nu15092035
Meyhöfer S, Chamorro R, Hallschmid M, Spyra D, Klinsmann N, Schultes B, Lehnert H, Meyhöfer SM, Wilms B. Late, but Not Early, Night Sleep Loss Compromises Neuroendocrine Appetite Regulation and the Desire for Food. Nutrients. 2023; 15(9):2035. https://doi.org/10.3390/nu15092035
Chicago/Turabian StyleMeyhöfer, Svenja, Rodrigo Chamorro, Manfred Hallschmid, Denisa Spyra, Nelli Klinsmann, Bernd Schultes, Hendrik Lehnert, Sebastian M. Meyhöfer, and Britta Wilms. 2023. "Late, but Not Early, Night Sleep Loss Compromises Neuroendocrine Appetite Regulation and the Desire for Food" Nutrients 15, no. 9: 2035. https://doi.org/10.3390/nu15092035
APA StyleMeyhöfer, S., Chamorro, R., Hallschmid, M., Spyra, D., Klinsmann, N., Schultes, B., Lehnert, H., Meyhöfer, S. M., & Wilms, B. (2023). Late, but Not Early, Night Sleep Loss Compromises Neuroendocrine Appetite Regulation and the Desire for Food. Nutrients, 15(9), 2035. https://doi.org/10.3390/nu15092035