
Revisiting PINI Scoring in Light of Recent Biological Advances


Abstract
1. Introduction
2. Biomarkers Pertaining to the PINI Formula
2.1. Albumin (ALB)
2.2. Transthyretin (TTR)
2.3. C-Reactive Protein (CRP)
2.4. Alpha-1 Acid Glycoprotein (AGP)
3. Discussion
3.1. The RBP–Retinol Binary
3.2. The Iron-Deficient Anemia Burden
4. Diagnostic Tools and Dietary Strategies
5. What Lesson Is to Be Learnt from the TTR Story?
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
- ≥40: life-threatening state
- 31 to 40: severe GHS state
- 21 to 30: medium impact GHS state
- 11 to 20: moderate impact
- 1 to 10: low impact
- ≤0: steady state
References
- Ingenbleek, Y.; Carpentier, Y.A. A prognostic inflammatory and nutritional index scoring critically ill patients. Int. J. Vitam. Nutr. Res. 1985, 55, 91–101. [Google Scholar] [PubMed]
- Pressac, M.; Vignoli, L.; Aymard, P.; Ingenbleek, Y. Usefulness of a prognostic inflammatory and nutritional index in pediatric clinical practice. Clin. Chim. Acta 1990, 188, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Kudlácková, M.; Anděl, M.; Hájková, H.; Nováková, J. Acute phase proteins and prognostic inflammatory and nutritional index (PINI) in moderately burned children aged up to 3 Years. Burns 1990, 16, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Gϋnel, E.; Cağlayan, O.; Cağlayan, F.; Sahin, T.K. Acute-phase changes in children recovering from minor surgery. Pediatr. Surg. Int. 1998, 14, 199–201. [Google Scholar] [CrossRef]
- Vehe, K.L.; Brown, R.O.; Kuhl, D.A.; Boucher, B.; Luther, R.W.; Kudsk, K.A. The prognostic inflammatory and nutritional index in traumatized patients receiving enteral nutrition support. J. Am. Coll. Nutr. 1991, 10, 355–363. [Google Scholar] [CrossRef]
- Schlossmacher, P.; Hasselmann, M.; Meyer, N.; Kara, F.; Delabranche, X.; Kummerlen, C.; Ingenbleek, Y. The prognostic value of nutritional and inflammatory indices in critically ill patients with acute respiratory failure. Clin. Chem. Lab. Med. 2002, 40, 1339–1343. [Google Scholar] [CrossRef]
- Dessi, M.; Noce, A.; Agnoli, A.; De Angelis, S.; Fuiano, L.; Tozzo, C.; Taccone-Gallucci, M.; Federici, G. The usefulness of the prognostic and nutritional index in a haemodialysis population. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 811–815. [Google Scholar] [CrossRef]
- Gharsallah, H.; Hajjej, Z.; Naas, I.; Aouni, Z.; Stambouli, I.; Ferjani, M. Assessment of nutritional status and prognosis in surgical intensive care unit: The prognostic and inflammatory index (PINI). Int. J. Nutr. Food Sci. 2014, 5, 477–483. [Google Scholar] [CrossRef]
- Walsh, D.; Mahmoud, F.; Barna, B. Assessment of nutritional status and prognosis in advanced cancer: Interleukin-6, C-reactive protein, and the prognostic and inflammatory nutritional index. Support Care Cancer 2003, 11, 60–62. [Google Scholar] [CrossRef]
- Dupire, S.; Wemeau, M.; Debarri, H.; Pascal, L.; Hivert, B.; Willekens, C.; Boyle, E.; Manier, S.; Thielemens, B.; Onraed, B.; et al. Prognostic value of PINI index in patients with multiple myeloma. Eur. J. Hematol. 2012, 88, 306–313. [Google Scholar] [CrossRef]
- Kirov, K.M.; Xu, H.P.; Crenn, P.; Goater, P.; Tzanis, D.; Bouhadiba, M.T.; Abdelhafidh, K.; Kirova, Y.M.; Bonvalot, S. Role of nutritional status in the early postoperative prognosis of patients operated for retropperitoneal liposarcoma (RLS): A single center experience. Eur. J. Surg. Oncol. 2019, 45, 261–267. [Google Scholar] [CrossRef]
- Bonnefoy, M.; Ayzac, L.; Ingenbleek, Y.; Kostka, T.; Boisson, R.C.; Bienvenu, J. Usefulness of the prognostic inflammatory and nutritional index (PINI) in hospitalized elderly patients. Int. J. Vitam. Nutr. Res. 1998, 68, 189–195. [Google Scholar]
- WHO. Serum Retinol Concentrations for Determining the Prevalence of Vitamin A Deficiency in Populations; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- WHO. Serum Ferritin Concentrations for the Assessment of Iron Status and Iron Deficiency in Populations: Vitamin and Mineral Nutrition Information System; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Ancell, H. Course of lectures on the physiology and pathology of the blood and the other animal fluids. Lancet 1839, 1, 222–231. [Google Scholar]
- Rothschild, M.A.; Oratz, M.; Schreiber, S.S. Albumin synthesis. N. Engl. J. Med. 1972, 286, 748–757. [Google Scholar] [CrossRef]
- Peters, T., Jr. Serum Albumin. In The Plasma Proteins. Structure, Function and Genetic Control; Putnam, F.W., Ed.; Academic Press: New York, NY, USA, 1975; Volume 1, pp. 133–181. [Google Scholar]
- Straus, D.S.; Marten, N.W.; Hayden, J.M.; Burke, E.J. Protein restriction specifically decreases the abundance of serum albumin and transthyretin nuclear transcripts in rat liver. J. Nutr. 1994, 124, 1041–1051. [Google Scholar] [CrossRef]
- Robbins, J. Structure and function of thyroid transport proteins. In Thyroid Hormone Metabolism; Harland, W.A., Orr, J.S., Eds.; Academic Press: London, UK, 1975; pp. 1–22. [Google Scholar]
- Young, V.R.; Pellett, P.L. Current concepts concerning indispensable amino acid needs in adults and their implications for international nutrition planning. Food Nutr. Bull. 1990, 12, 289–300. [Google Scholar] [CrossRef]
- Soeters, P.B.; Wolfe, R.R.; Shenkin, A. Hypoalbuminemia: Pathogenesis and clinical significance. J. Parenter. Enter. Nutr. 2019, 43, 181–193. [Google Scholar] [CrossRef]
- Fuka, T.; Wada, Y.; Kawakami, S.; Miyaji, K. Serum albumin redox states: More than oxidative stress biomarker. Antioxidants 2021, 10, 503. [Google Scholar] [CrossRef]
- Tojo, A.; Kinugasa, S. Mechanisms of glomerular filtration and tubular reabsorption. Int. J. Nephrol. 2012. [Google Scholar] [CrossRef]
- Nielsen, R.; Christensen, E.I.; Birn, H. Megalin and cubilin in proximal tubule protein reabsorption: From experimental models to human disease. Kidney Int. 2016, 89, 58–67. [Google Scholar] [CrossRef]
- Gekle, M. Renal tubule albumin transport. Annu. Rev. Physiol. 2005, 67, 573–594. [Google Scholar] [CrossRef]
- Haraldsson, B.S.; Johsson, E.K.; Rippe, B. Glomerular permselectivity is dependent on adequate serum concentrations of orosomucoid. Kidney Int. 1992, 41, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Ballerman, B.J.; Nyström, J.; Haraldsson, B. The glomerular endothelium restricts albumin filtration. Front. Med. 2021, 8, 766689. [Google Scholar] [CrossRef] [PubMed]
- Viasus, D.; Garcia-Vidal, C.; Simonetti, A.; Manresa, F.; Dorca, J.; Gudiol, F.; Carratalà, J. Prognostic value of serum albumin levels in hospitalized adults with community-acquired pneumonia. J. Infect. 2013, 66, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Shigematsu, H.; Iwata, E.; Nakajima, H.; Tanaka, M.; Okuda, A.; Kawasaki, S.; Suga, Y.; Masuda, K.; Tanaka, Y. Hypoalbuminemia increased the length of stay in the treatment of postoperative acute surgical site infection in spinal surgery. Spine 2020, 45, 1564–1571. [Google Scholar] [CrossRef]
- Qian, S.Y.; Jin, D.; Chen, Z.B.; Ye, Y.C.; Xiang, W.W.; Ye, L.M.; Pan, J.Y. Hypoalbuminemia, a novel pronostic factor for prediction of long-term outcomes in critially ill patients with septic shock. Int. J. Clin. Exp. Med. 2019, 12, 7401–7409. [Google Scholar]
- Anderson, C.G.; Altmann, A. The electrophoretic serum-protein pattern in malignant malnutrition. Lancet 1951, 257, 203–204. [Google Scholar] [CrossRef]
- Whitehead, R.G.; Coward, W.A.; Lunn, P.G. Serum-albumin concentration and the onset of kwashiorkor. Lancet 1973, 7794, 63–66. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; De Visscher, M.; De Nayer, P. Measurement of prealbumin as index of protein-calorie malnutrition. Lancet 1972, 7768, 106–109. [Google Scholar] [CrossRef]
- Ingenbleek, Y. Protein-Calorie Malnutrition in Infants of Young Age. Repercussions on Thyroid Function and Serum Carrier-Proteins. Ph.D. Thesis, Catholic University of Louvain, Ottignies-Louvain-la-Neuve, Belgium, 1977. [Google Scholar]
- Kanai, M.; Raz, A.; Goodman, D.S. Retinol-binding protein: The transport protein for vitamin A in human plasma. J. Clin. Investig. 1968, 47, 2025–2044. [Google Scholar] [CrossRef]
- Monaco, H.L. The transthyretin-retinol binding protein complex. In Recent Advances in Transthyretin Evolution, Structure and Biological Functions; Richardson, D.S., Cody, V., Eds.; Springer: Berlin, Germany, 2009; pp. 123–142. [Google Scholar]
- Forbes, G.B. Human Body Composition: Growth, Aging, Nutrition, and Activity; Springer: Berlin, Germany, 1987; pp. 1–350. [Google Scholar]
- Cohn, S.H.; Vartsky, D.; Yasumura, S.; Vaswani, A.N.; Ellis, K.J. Indexes of cell body mass: Nitrogen versus potassium. Am. J. Physiol. 1983, 244, E305–E310. [Google Scholar] [CrossRef]
- Bienvenu, J.; Jeppson, J.O.; Ingenbleek, Y. Transthyretin & retinol-binding protein. In Serum Proteins in Clinical Medicine; Ritchie, R.F., Navolotskaia, O., Eds.; Foundation for Blood Research: Scarborough, ME, USA, 1996; pp. 9.011–9.018. [Google Scholar]
- Ingenbleek, Y. Plasma transthyretin reflects the fluctuations of lean body mass in health and disease. In Recent Advances in Transthyretin Evolution, Structure and Biological Functions; Richardson, D.S., Cody, V., Eds.; Springer: Berlin, Germany, 2009; pp. 329–357. [Google Scholar]
- Ingenbleek, Y.; Bernstein, L.H. Plasma transthyretin as a biomarker of lean body mass and catabolic states. Adv. Nutr. 2015, 6, 572–580. [Google Scholar] [CrossRef]
- Ingenbleek, Y. Plasma transthyretin is a nutritional biomarker in human morbidities. Front. Med. 2022, 6, 540–550. [Google Scholar] [CrossRef]
- Dellière, S.D.; Pouga, L.; Neveux, N.; Hernvann, A.; de Bandt, J.P.; Cynober, L. Assessment of transthyretin cut-off values for a better screening of malnutrition: Retrospective determination and prospective validation. Clin. Nutr. 2021, 40, 907–911. [Google Scholar] [CrossRef]
- Dramaix, M.; Brasseur, D.; Donnen, P.; Bawhere, P.; Porignon, D.; Tonglet, R.; Hennart, P. Prognostic indices for mortality of hospitalized children in central Africa. Am. J. Epidemiol. 1996, 143, 1235–1243. [Google Scholar] [CrossRef]
- Tillet, W.S.; Francis, T., Jr. Serological reactions in pneumonia with a non-protein somatic fraction of pneumococcus. J. Exp. Med. 1930, 52, 561–571. [Google Scholar] [CrossRef]
- Floyd-Smith, G.; Whitehead, A.S.; Colten, H.R.; Franke, U. The human C-reactive gene (CRP) and serum amyloid P component gene (APCS)) are located on the proximal long arm of chromosome 1. Immunogenetics 1983, 24, 171–176. [Google Scholar]
- Agrawal, A.; Kilpatrick, J.M.; Volanakis, J.E. Structure and function of human C-reactive protein. In Acute Phase Proteins; Mackiewicz, A., Kushner, I., Bauman, H., Eds.; CRC Press: London, UK, 1993; pp. 79–92. [Google Scholar]
- Whicher, J.T.; Banks, R.E.; Thompson, D.; Evans, S.W. The measurement of acute-phase proteins as disease markers. In Acute Phase Proteins; Mackiewicz, A., Kushner, I., Bauman, H., Eds.; CRC Press: London, UK, 1993; pp. 633–650. [Google Scholar]
- Baumann, H.; Gauldie, J. The acute phase response. Immunol. Today 1994, 15, 74–80. [Google Scholar] [CrossRef]
- Weimer, H.E.; Mehl, J.W.; Winzler, R.J. Studies on the mucoproteins of human plasma. V. Isolation and characterization of a homogeneous mucoprotein. J. Biol. Chem. 1950, 185, 561–568. [Google Scholar] [CrossRef]
- Schmid, K. α1-Acid glycoprotein. In The Plasma Proteins; Putnam, F.W., Ed.; Academic Press: New York, NY, USA, 1975; pp. 183–228. [Google Scholar]
- Dente, L. Human α1-acid glycoprotein genes. Progr. Clin. Biol. Res. 1989, 300, 85–98. [Google Scholar]
- Bienvenu, J.; Sann, L.; Bienvenu, F.; Lahet, C.; Divry, P.; Cotte, J.; Bethenod, M. Laser nephelometry of orosomucoid in serum of newborns: Reference intervals and relation to bacterial infection. Clin. Chem. 1981, 27, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Maraj, M.; Hetwer, P.; Kuśnierz-Cabala, B.; Maziarz, B.; Dumnicka, P.; Kuźniewski, M.; Ceranowicz, P. α1-Acid glycoprotein and dietary intake in end-stage renal disease patients. Nutrients 2021, 13, 3671. [Google Scholar] [CrossRef] [PubMed]
- Prowse, K.P.; Baumann, H. Interleukin-1 and Interleukin-6 stimulate acute phase protein production in primary mouse hepatocytes. J. Leukoc. Biol. 1989, 45, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Ceciliani, F.; Lecchi, C. The immune functions of α1-acid glycoprotein. Curr. Protein Pept. Sci. 2019, 20, 505–524. [Google Scholar] [CrossRef]
- Xiao, K.; Su, S.; Yan, P.; Han, B.; Li, J.; Wang, H.; Jia, Y.; Li, X.; Xie, L. Alpha1-acid glycoprotein as a biomarker for the early diagnosis and monitoring the prognosis of sepsis. J. Crit. Care 2015, 30, 744–751. [Google Scholar] [CrossRef]
- Friedman, M.J. Control of malaria virulence by alpha 1-acid glycoprotein (orosomucoid), an acute-phase (inflammatory) reactant. Proc. Natl. Acad. Sci. USA 1983, 80, 5421–5424. [Google Scholar] [CrossRef]
- Rawat, R.; Stoltzfus, R.J.; Ntozini, R.; Mutasa, K.; Iliff, P.J.; Humphrey, J.H. Influence of inflammation as measured by alpha-1-acid glycoprotein on iron status indicators among HIV-positive postpartum Zimbabwean women. Eur. J. Clin. Nutr. 2009, 63, 787–793. [Google Scholar] [CrossRef]
- Ayoya, M.A.; Spiekermann-Brouwer, G.M.; Stoltzfus, R.J.; Nemeth, E.; Habicht, J.P.; Ganz, T.; Rawat, R.; Traoré, A.K.; Garza, C. Alpha 1-acid glycoprotein, hepcidin, C-reactive protein, and serum ferritin are correlated in anemic schoolchildren with Schistosoma haematobium. Am. J. Clin. Nutr. 2010, 91, 1784–1790. [Google Scholar] [CrossRef]
- Kim, S.U.; Jeon, M.Y.; Lim, T.S. Diagnostic performance of serum asialo-α1-acid glycoprotein for advanced liver fibrosis or cirhosis in patients with chronic hepatitis B or nonalcoholic fatty liver disease. Korean J. Gastroenterol. 2019, 74, 341–348. [Google Scholar] [CrossRef]
- Liang, J.; Zhu, J.; Wang, M.; Singal, A.G.; Odewole, M.; Kagan, S.; Renteria, V.; Liu, S.; Parikh, N.D.; Lubman, D.M. Evaluation of AGP fucosylation as a marker for hepatocellular carcinoma of three different etiologies. Sci. Rep. 2019, 9, 11580. [Google Scholar] [CrossRef]
- Gannon, B.M.; Glesby, M.J.; Finkelstein, J.L.; Raj, T.; Erickson, D.; Mehta, S. A point-of-care assay for alpha-1-acid glycoprotein as a diagnostic tool for rapid, mobile-based determination of inflammation. Curr. Res. Biotechnol. 2019, 1, 41–48. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, N.M.; Kim, S.Y.; Yi, D.Y.; Yun, S.W.; Chae, S.A.; Lim, I.N. Effectiveness of prealbumin as an indicator of growth in neonates. Medicine 2021, 100, e27603. [Google Scholar] [CrossRef]
- Elhasid, R.; Laor, A.; Lischinsky, S.; Postovsky, S.; Weyl Ben Arush, M. Nutritional status of children with solid tumors. Cancer 1999, 86, 119–125. [Google Scholar] [CrossRef]
- Sato, S.; Shiozawa, M.; Nukada, S.; Iguchi, K.; Kazama, K.; Atsumi, Y.; Numata, M.; Tamagawa, H.; Tanaka, K.; Oshima, T.; et al. Preoperative pre-albumin concentration as a predictor of short-term outcomes in elderly patients with colorectal cancer. Anticancer Res. 2021, 41, 5195–5202. [Google Scholar] [CrossRef]
- Devakonda, A.; George, L.; Raoof, S.; Esan, A.; Saleh, A.; Bernstein, L.H. Transthyretin as a marker to predict outcome in critically ill patients. Clin. Biochem. 2008, 41, 1126–1130. [Google Scholar] [CrossRef]
- Yang, H.T.; Yim, H.; Cho, Y.S.; Kim, D.; Hur, J.; Kim, J.H.; Lee, B.C.; Seo, D.K.; Kim, H.S.; Chun, W. Prediction of clinical outcomes for massively-burned patients via transthyretin levels in the early postburn period. J. Trauma Acute Care Surg. 2012, 72, 999–1005. [Google Scholar] [CrossRef]
- Fan, Y.; Sun, Y.; Man, C.; Lang, Y. Preoperative serum prealbumin level and adverse prognosis in patients with hepatocellular carcinoma after hepatectomy: A meta-analysis. Front. Oncol. 2021, 11, 775425. [Google Scholar] [CrossRef]
- Chertow, G.M.; Goldstein-Fuchs, D.J.; Lazarus, J.M.; Kaysen, G.A. Prealbumin, mortality, and cause-specific hospitalization in hemodialysis patients. Kidney Int. 2005, 68, 2794–2800. [Google Scholar] [CrossRef]
- Akashi, M.; Minami, Y.; Haruki, S.; Jujo, K.; Hagiwara, N. Prognostic implications of prealbumin level on admission in patients with acute heart failure referred to a cardiac intensive care unit. J. Cardiol. 2019, 73, 114–119. [Google Scholar] [CrossRef]
- Sugumar, D.; Arockiaraj, J.; Amritanand, R.; David, K.S.; Krishnan, V. Role of biochemical nutritional parameters as predictors of postoperative morbidity in major spine surgeries. Asian Spine J. 2021, 15, 504–511. [Google Scholar] [CrossRef]
- Zinellu, A.; Mangoni, A.A. Serum prealbumin concentrations, COVID-19 severity, and mortality: A systematic review and meta-analysis. Front. Med. 2021, 8, 638529. [Google Scholar] [CrossRef] [PubMed]
- Isono, N.; Imamura, Y.; Ohmura, K.; Ueda, N.; Kawabata, S.; Furuse, M.; Kuroiwa, T. Transthyretin concentrations in acute stroke patients predict convalescent rehabilitation. J. Stroke Cerebrovasc. Dis. 2017, 26, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Ingenbleek, Y.; Bernstein, L.H. Downsizing of lean body mass is a key determinant of Alzheimer’s disease. J. Alzheimers Dis. 2015, 44, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, L.H.; Leukhardt-Fairfield, C.J.; Pleban, W.; Rudolph, R. Usefulness of data on albumin and prealbumin concentrations in determining effectiveness of nutritional support. Clin. Chem. 1989, 35, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Chertow, G.M.; Ackert, K.; Lew, N.L.; Lazarus, J.M.; Lowrie, E.G. Prealbumin is as important as albumin in the nutritional assessment of hemodialysis patients. Kidney Int. 2000, 58, 2512–2517. [Google Scholar] [CrossRef]
- Dalrymple, L.; Johansen, K.L.; Chertow, G.M.; Grimes, B.; Anand, S.; McCullogh, C.E.; Kaysen, G.A. Longitudinal measures of serum albumin and prealbumin concentrations in incident dialysis patients: The comprehensive dialysis study. J. Ren. Nutr. 2013, 23, 91–97. [Google Scholar] [CrossRef]
- Sergi, G.; Coin, A.; Enzi, G.; Volpato, S.; Inelmen, E.M.; Buttarello, M.; Peloso, M.; Mulone, S.; Marin, S.; Bonometto, P. Role of visceral proteins in detecting malnutrition in the elderly. Eur. J. Clin. Nutr. 2006, 60, 203–209. [Google Scholar] [CrossRef]
- Beetham, R.; Dawnay, A.; Ghany, C.; Dubrey, S.; Miles, J. A radioimmunoassay for human urinary prealbumin. Ann. Clin. Biochem. 1993, 30, 377–382. [Google Scholar] [CrossRef]
- Rask, L.; Vahlquist, A.; Peterson, P.A. Studies on two physiological forms of the human retinol binding protein differing in vitamin A and arginine content. J. Biol. Chem. 1971, 246, 6638–6648. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schultze, K.; Raiten, D.J. Biomarkers of nutrition for development (BOND)-Vitamin A review. J. Nutr. 2016, 146, S1816–S1848. [Google Scholar] [CrossRef]
- Gannon, B.M.; Tanumihardjo, S.A. Comparisons among equation used for retinol isotope dilution in the assessment of total body stores and total liver reserves. J. Nutr. 2015, 145, 847–854. [Google Scholar] [CrossRef]
- Kasper, H.; Brodersen, M.; Schedel, R. Concentrations of vitamin A, retinol-binding protein and prealbumin in response to stress. Acta Hepato-Gastroenterol. 1975, 22, 403–408. [Google Scholar]
- Hoover, H.C., Jr.; Ryan, J.A.; Anderson, E.J.; Fischer, J.E. Nutritional benefits of immediate postoperative jejunal feeding of an elemental diet. Am. J. Surg. 1980, 139, 153–159. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; Van den Schrieck, H.G.; De Nayer, P.; De Visscher, M. The role of retinol-binding protein in protein-calorie malnutrition. Metabolism 1975, 24, 633–641. [Google Scholar] [CrossRef]
- Murakami, T.; Ohnishi, S.; Nishiguchi, S.; Maeda, S.; Araki, S.; Shimada, K. Acute-phase response of mRNAs for serum amyloid P component, C-reactive protein and prealbumin (transthyretin) in mouse liver. Biochem. Biophys. Res. Commun. 1988, 155, 554–560. [Google Scholar] [CrossRef]
- Banks, R.E.; Forbes, M.A.; Storr, M.; Higginson, J.; Thompson, D.; Raynes, J.; Illingworth, J.M.; Perren, T.J.; Selby, P.J.; Whicher, J.T. The acute phase protein response in patients receiving subcutaneous Il-6. Clin. Exp. Immunol. 1995, 102, 217–223. [Google Scholar] [CrossRef]
- de Jong, F.A.; Schreiber, G. Messenger RNA levels of plasma proteins in rat liver during protein depletion and refeeding. J. Nutr. 1987, 117, 1795–1800. [Google Scholar] [CrossRef]
- Peterson, P.A. Studies on the interaction between prealbumin, retinol-binding protein, and vitamin A. J. Biol. Chem. 1971, 246, 44–49. [Google Scholar] [CrossRef]
- Jaconi, S.; Rose, K.; Hughes, G.J.; Saurat, J.H.; Siegenthaler, G. Characterization of two post-translationally processed forms of human serum retinol-binding protein: Altered ratios in chronic renal failure. J. Lipid. Res. 1995, 36, 1247–1253. [Google Scholar] [CrossRef]
- Bernard, A.M.; Vyskocil, A.A.; Mahieu, P.; Lauwerijs, R.R. Assessment of urinary retinol-biding protein as an index of proximal tubular injury. Clin. Chem. 1987, 33, 775–779. [Google Scholar] [CrossRef]
- Xia, Y.; Peng, C.P.; Qu, S.; Liu, F.; Peng, Y. Correlation studies between urinary retinol binding protein and renal tubular damage. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2011, 36, 1008–1011. [Google Scholar] [PubMed]
- Raila, J.; Willnow, T.E.; Schweigert, F.J. Megalin-mediated reuptake of retinol in the kidneys of mice is essential for vitamin A homeostasis. J. Nutr. 2005, 135, 2512–2516. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, D.B.; Princé, H.P.; Burr, W.A.; Bradwell, A.R.; Black, E.G.; Evans, A.E.; Hoffenberg, R. The inter-relationship of thyroid hormones, vitamin A and their binding proteins following acute stress. Clin. Endocrinol. 1978, 8, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K.; Alvarez, J.O.; Stephensen, C.B. Increased urinary retinol loss in children with severe infections. Lancet 1998, 351, 1033–1034. [Google Scholar] [CrossRef]
- Gudas, L.J. Retinoid metabolism: New insights. J. Mol. Endocrinol. 2022, 69, T37–T49. [Google Scholar] [CrossRef]
- Steinhoff, J.S.; Lass, A.; Schupp, M. Retinoid homeostasis and beyond: How retinol-binding protein 4 contributes to health and disease. Nutrients 2022, 14, 1236. [Google Scholar] [CrossRef]
- Willumsen, J.F.; Simmank, K.; Filteau, S.M.; Wagstaff, L.A.; Tomkins, A.M. Toxic damage to the respiratory epithelium induces acute changes in vitamin A metabolism without depleting retinol stores of South African children. J. Nutr. 1997, 127, 1339–1343. [Google Scholar] [CrossRef]
- Donnen, P.; Dramaix, M.; Brasseur, D.; Bitwe, R.; Bisimwa, G.; Hennart, P. The molar ratio of serum retinol-binding protein (RBP) to transthyretin (TTR) is not useful to assess vitamin A status during infection in hospitalized children. Eur. J. Clin. Nut. 2001, 55, 1043–1047. [Google Scholar] [CrossRef]
- Bataille, S.; Landrier, J.F.; Astier, J.; Cado, S.; Sallette, J.; Serveaux, M.; Burtey, S.; Cohen, J.; Tournier, C.; Tournaire, F.; et al. Plasma retinol concentration is mainly driven by transthyretin in hemodialysis patients. J. Ren. Nutr. 2017, 27, 395–401. [Google Scholar] [CrossRef]
- Vaz-Rodrigues, R.; Mazuecos, L.; Villar, M.; Urra, J.M.; Gortázar, C.; de la Fuente, J. Serum biomarkers for nutritional status as predictors in COVID-19 patients before and after vaccination. J. Funct. Foods 2023, 101, 105412. [Google Scholar] [CrossRef]
- WHO. Worldwide Prevalence of Anaemia 1995–2005. WHO Global Database on Anaemia; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Suchdev, P.S.; Williams, A.M.; Mei, Z.; Flores-Ayala, R.; Pasricha, S.R.; Rogers, L.M.; Namaste, S. Assessment of iron status in settings of inflammation: Challenges and potential approaches. Am. J. Clin. Nutr. 2017, 106, 1626S–1633S. [Google Scholar] [CrossRef] [PubMed]
- Dagg, J.H.; Smith, J.A.; Goldberg, A. Urinary excretion of iron. Clin. Sci. 1966, 106, 495–503. [Google Scholar]
- Howard, R.L.; Buddington, B.; Alfrey, A.C. Urinary albumin, transferrin and iron excretion in diabetic patients. Kidney Int. 1991, 40, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Ouled Amor, A.A.; Goemaere-Vanneste, J.; Antoine, J.L.; Lauwerijs, R.R.; Lambert, A.; Vandeleene, B. Microtransferrinuria is a more sensitive indicator of early glomerular damage in diabetes than microalbuminuria. Clin. Chem. 1988, 34, 1920–1921. [Google Scholar] [CrossRef] [PubMed]
- MacGillivray, R.T.; Mendez, E.; Sinha, S.K.; Sutton, M.R.; Lineback-Zins, J.; Brew, K. The complete amino acid sequence of human serum transferrin. Proc. Natl. Acad. Sci. USA 1982, 79, 2504–2508. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; McVey Ward, D.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef]
- Weiss, G.; Goodnough, L.T. Anemia of chronic disease. N. Engl. J. Med. 2005, 352, 1011–1023. [Google Scholar] [CrossRef]
- Papanikolaou, G.; Tzilianos, M.; Christakis, J.I.; Bogdanos, D.; Tsimirika, K.; MacFarlane, J.; Goldberg, Y.P.; Sakellaropoulos, N.; Ganz, T.; Nemeth, E. Hepcidin in iron overload disorders. Blood 2005, 105, 4103–4105. [Google Scholar] [CrossRef]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past, present and future. Biochim. Biophys. Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef]
- Thurnham, D.I. Interactions between nutrition and immune function: Using inflammation biomarkers to interpret micronutrient status. Proc. Nutr. Soc. 2014, 73, 1–8. [Google Scholar] [CrossRef]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic effects of inflammation on vitamin A and carotenoids in humans and animal models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- Wagh, V.D.; Deore, B.R. Ready to use therapeutic food (RUTF): An overview. Adv. Life Sci. Health 2015, 2, 1–15. [Google Scholar]
- Thurnham, D.I.; Northrop-Clewes, C.A.; Knowles, J. The use of adjustment factors to address the impact of inflammation on vitamin A and iron status in humans. J. Nutr. 2015, 145, 1137S–1143S. [Google Scholar] [CrossRef]
- Larson, L.M.; Namaste, S.M.; Williams, A.M.; Engle-Stone, R.; Addo, O.Y.; Suchdev, P.S.; Wirth, J.P.; Temple, V.; Serdula, M.; Northrop-Clewes, C.A. Adjusting retinol-binding protein concentrations for inflammation: Biomarkers reflecting inflammation and nutritional determinants of anemia (BRINDA) project. Am. J. Clin. Nutr. 2017, 106, 390S–401S. [Google Scholar]
- Rohner, F.; Namaste, S.M.; Larson, L.M.; Addo, O.Y.; Mei, Z.; Suchdev, P.S.; Williams, A.M.; Sakr Ashour, F.A.; Rawat, R.; Raiten, D.J.; et al. Adjusting soluble transferrin receptor concentrations for inflammation: Biomarkers reflecting inflammation and nutritional determinants of anemia(BRINDA)project. Am. J. Clin. Nutr. 2017, 106, 372S–382S. [Google Scholar] [CrossRef]
- Namaste, S.M.; Rohner, F.; Huang, J.; Bhushan, N.L.; Flores Ayala, R.; Kupka, R.; Mei, Z.; Rawat, R.; Williams, A.M.; Raiten, D.J.; et al. Adjusting ferritin concentrations for inflammation: Biomarkers reflecting inflammation and nutritional determinants of anemia (BRINDA) project. Am. J. Clin. Nutr. 2017, 106, 359S–371S. [Google Scholar] [CrossRef]
- Lynch, S.R.; Dassenko, S.A.; Cook, J.; Juillerat, M.A.; Hurrell, R.F. Inhibitory effect of a soybean-protein-related moiety on iron absorption in humans. Am. J. Clin. Nutr. 1994, 60, 567–572. [Google Scholar] [CrossRef]
- Schmidt, J.A.; Rinaldi, S.; Scalbert, A.; Ferrari, P.; Achaintre, D.; Gunter, M.J.; Appleby, P.N.; Key, T.; Travis, R.C. Plasma concentrations and intakes of amino acids in male meat-eaters, vegetarians and vegans: A cross-sectional analysis in the EPIC-Oxford cohort. Eur. J. Clin. Nutr. 2016, 70, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Forbes, G.B. Stature and lean body mass. Am. J. Clin. Nutr. 1974, 27, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Dewey, K.G.; Beaton, G.; Field, C.; Lonnerdal, B.; Reeds, P. Protein requirements of infants and children. Eur. J. Clin. Nutr. 1996, S119–S150. [Google Scholar]
- Young, V.R.; Borgonha, S. Nitrogen and amino acid requirements: The Massachusetts Institute of Technology amino acid requirement pattern. J. Nutr. 2000, 130, 1814S–1849S. [Google Scholar] [CrossRef]
- Rose, W.C. Human Protein Requirements and their fulfillment in Practice, Proceedings of a Conference in Princetown, USA, 1955; Sponsored jointly by F.A.O., W.H.O. and Josiah Macy Jr. Foundation; Waterlow, J.C., Stephen, J.M.L., Eds.; John Wright & Sons: Bristol, UK, 1957. [Google Scholar]
- Mak, T.N.; Angeles-Agdeppa, I.; Tassy, M.; Capanzana, M.V.; Offord, E.A. The nutritional impact of milk beverages in reducing nutrient inadequacy among children aged one to five years of age in the Philippines: A dietary modelling study. Nutrients 2020, 12, 3330. [Google Scholar] [CrossRef]
- Parikh, P.; Semba, R.; Manary, M.; Swaminathan, S.; Udomkesmalee, E.; Bos, R.; Poh, B.K.; Rojroongwasinkul, N.; Geurts, J.; Sekartini, R.; et al. Animal source foods, rich in essential amino acids, are important for linear growth and development of young children in low- and middle-income countries. Matern. Child Nutr. 2022, 18, e13264. [Google Scholar] [CrossRef]
- Tadesse, S.E.; Zerga, A.A.; Mekonnen, T.C.; Tadesse, A.W.; Hussien, F.M.; Feleke, Y.W.; Anagaw, M.; Ayele, F.Y. Burden and determinants of anemia among under-five children in Africa: Systematic review and meta-analysis. Anemia 2022. [Google Scholar] [CrossRef]
- Kateera, F.; Ingabire, C.M.; Hakizimana, E.; Kalinda, P.; Mens, P.F.; Grobusch, M.P.; Mutesa, L.; van Vugt, M. Malaria, anaemia and under-nutrition: Three frequently co-existing conditions among preschool children in rural Rwanda. Malar. J. 2015, 14, 440–511. [Google Scholar] [CrossRef] [PubMed]
- Diouf, S.; Sylla, A.; Diop, F.; Diallo, A.; Sarr, M. Anemia among apparently healthy Senegalese children. Arch. Pediatr. 2013, 20, 312–313. [Google Scholar] [CrossRef]
- Gedfie, S.; Getawa, S.; Melku, M. Prevalence and associated factors of iron deficiency and iron deficiency anemia among under-5 children: A systematic review and meta-analysis. Glob. Pediatr. Health 2022, 9, 1–13. [Google Scholar] [CrossRef]
- Bahizire, E.; Bahwere, P.; Donnen, P.; Tugirimana, P.L.; Balol’ebwami, S.; Dramaix, M.; Nfundiko, C.; Chirimwami, R.; Mubamgwa, K. High prevalence of anemia but low level of iron deficiency in preschool children during a low transmission period of malaria in rural Kivu, Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 2017, 97, 489–496. [Google Scholar] [CrossRef]
- Kangas, S.T.; Salpéteur, C.; Nikièma, V.; Talley, L.; Briend, A.; Ritz, C.; Friis, H.; Kaestel, P. Vitamin A and iron status of children before and after treament of uncomplicated severe acute malnutrition. Clin. Nutr. 2020, 39, 3512–3519. [Google Scholar] [CrossRef]
- González-Fernández, D.; Nemeth, E.; Pons, E.d.C.; Sinisterra, O.T.; Rueda, D.; Starr, L.; Sangkhae, V.; Murillo, E.; Scott, M.E.; Koski, K.G. Multiple indicators of undernutrition, infection, and inflammation in lactating women are associated with maternal iron status and infant anthropometry in Panama: The MINDI cohort. Nutrients 2022, 14, 3497. [Google Scholar] [CrossRef]
- Keller, U. Nutritional laboratory markers in malnutrition. J. Clin. Med. 2019, 8, 775. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; Van den Schriek, H.G.; De Nayer, P.; De Visscher, M. Albumin, transferrin, and the thyroxine-binding prealbumin/retinol-binding protein complex in assessment of malnutrition. Clin. Chim. Acta 1975, 63, 61–67. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; Young, V.R. Transthyretin (prealbumin) in health and disease: Nutritional implications. Annu. Rev. Nutr. 1994, 14, 495–533. [Google Scholar] [CrossRef]
- McLaren, D.S. The Great Protein Fiasco. Lancet 1974, 7872, 93–96. [Google Scholar] [CrossRef]
- Semba, R.D. The rise and fall of protein malnutrition in global health. Ann. Nutr. Metab. 2016, 69, 79–88. [Google Scholar] [CrossRef]
- Williams, C.D. A nutritional disease of childhood associated with a maize diet. Arch. Dis. Child. 1933, 8, 423–433. [Google Scholar] [CrossRef]
- Brock, J.F.; Autret, M. Kwashiorkor in Africa; Bulletin of the World Health Organization: Geneva, Switzerland, 1952; pp. 1–71. [Google Scholar]
- Evans, D.C.; Corkins, M.R.; Malone, A.; Miller, S.; Mogensen, K.M.; Guenter, P.; Jensen, G.L.; ASPEN Malnutrition Committee. The use of visceral proteins as nutrition markers: An ASPEN position paper. Nutr. Clin. Pract. 2021, 36, 22–28. [Google Scholar] [CrossRef]
- Lacy, M.; Roesch, J.; Langsjoen, J. Things we do for no reason: Prealbumin testing to diagnose malnutrition in the hospitalized patient. J. Hosp. Med. 2019, 14, 239–241. [Google Scholar]
- Zuo, P.; Tong, S.; Yan, Q.; Cheng, L.; Li, Y.; Song, K.; Chen, Y.; Dai, Y.; Gao, H.; Zhang, C. Decreased prealbumin level is associated with increased risk for mortality in elderly hospitalized patients with COVID-19. Nutrition 2020, 78, 110930. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.J.; Lee, H.J.; Han, D.S.; Suh, Y.S.; Lee, Y.H.; Lee, H.S.; Cho, J.J.; Kong, S.H.; Yang, H.K. Prealbumin levels as a useful marker for predicting infectious complicationhs after gastric surgery. J. Gastrointest. Surg. 2011, 15, 2136–2144. [Google Scholar] [CrossRef] [PubMed]
- Ando, Y. Transthyretin: It’s miracle function and pathogenesis. Rinsho Byon 2009, 57, 228–235. [Google Scholar]
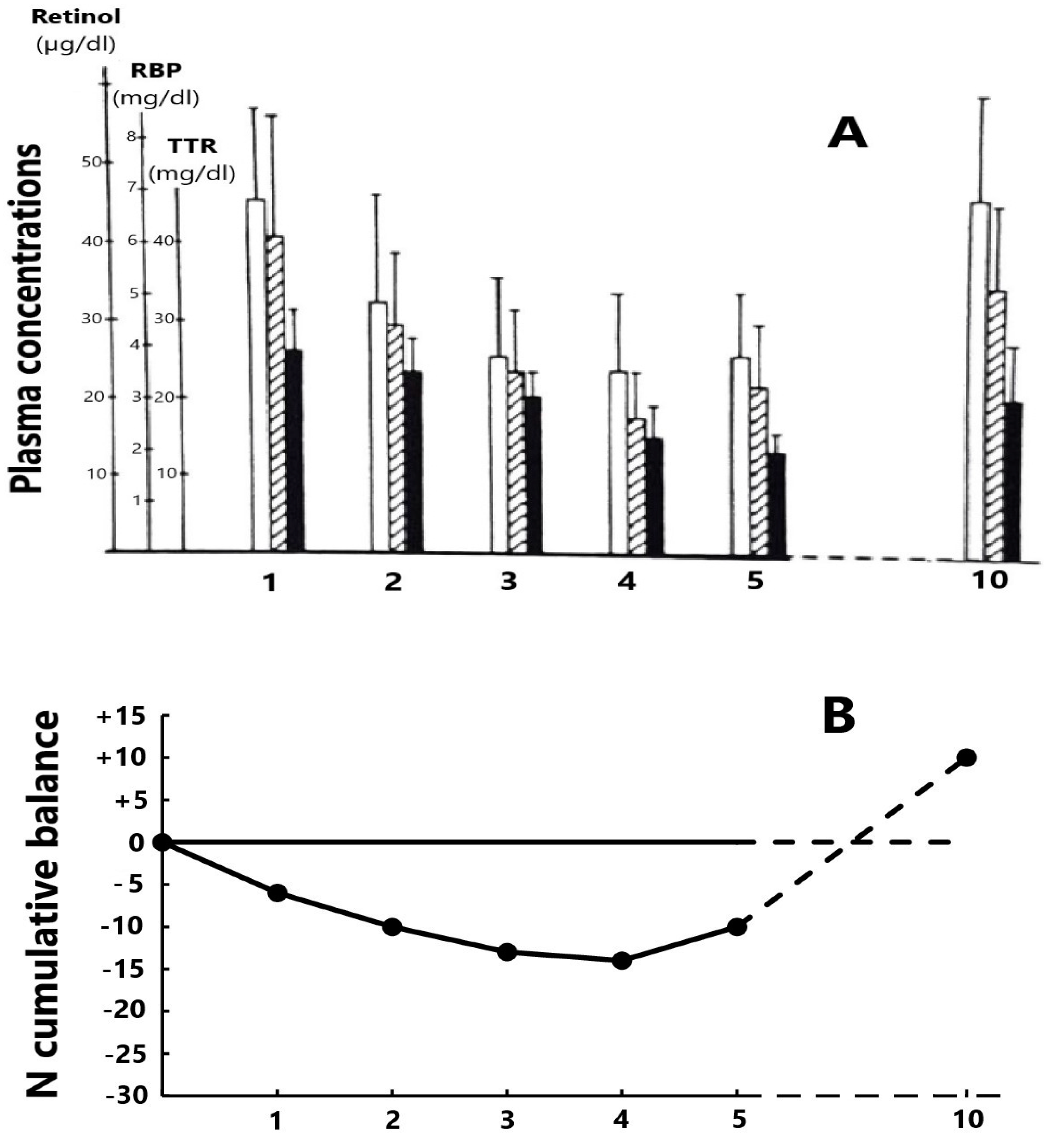
 RBP and ■ TTR). The nadir level is reached on days 4–5 and, in the absence of complications, healing is recorded on day 10 with full restoration of basal RCC values. (B) The cumulative mean N balance in 28 healthy adult subjects undergoing abdominal surgery and who were enterally refed [85]. Urinary excretion of nitrogenous compounds is mainly due to the cytokine-induced breakdown of tissue proteins stored in lean body mass (LBM) whose depletion may be quantified by TTR values showing the same declining profile as that of N.
RBP and ■ TTR). The nadir level is reached on days 4–5 and, in the absence of complications, healing is recorded on day 10 with full restoration of basal RCC values. (B) The cumulative mean N balance in 28 healthy adult subjects undergoing abdominal surgery and who were enterally refed [85]. Urinary excretion of nitrogenous compounds is mainly due to the cytokine-induced breakdown of tissue proteins stored in lean body mass (LBM) whose depletion may be quantified by TTR values showing the same declining profile as that of N.
RBP and ■ TTR). The nadir level is reached on days 4–5 and, in the absence of complications, healing is recorded on day 10 with full restoration of basal RCC values. (B) The cumulative mean N balance in 28 healthy adult subjects undergoing abdominal surgery and who were enterally refed [85]. Urinary excretion of nitrogenous compounds is mainly due to the cytokine-induced breakdown of tissue proteins stored in lean body mass (LBM) whose depletion may be quantified by TTR values showing the same declining profile as that of N.
RBP and ■ TTR). The nadir level is reached on days 4–5 and, in the absence of complications, healing is recorded on day 10 with full restoration of basal RCC values. (B) The cumulative mean N balance in 28 healthy adult subjects undergoing abdominal surgery and who were enterally refed [85]. Urinary excretion of nitrogenous compounds is mainly due to the cytokine-induced breakdown of tissue proteins stored in lean body mass (LBM) whose depletion may be quantified by TTR values showing the same declining profile as that of N.

{kind=link}
| Normal Adult Values | Molecular Conformation | Molecular Mass (MM) | Biological Half-Life (T1/2) | |
|---|---|---|---|---|
| ALB | 35–45 g/L | unglycosylated monopeptide (580 Aas) | 66.5 kDa | 19 days |
| TTR | 280–350 mg/L | unglycosylated tetramer (4 × 127 Aas) | 55 kDa | 2 days |
| CRP | ≤1 mg/L | cyclic pentaxin (5 × 206 Aas) | 118 kDa | 19 h |
| AGP | 0.4–1.4 g/L | monopeptide (183 Aas) 42% glycosylated | 41 kDa | 60–120 h |
| Day 1 (n = 39) | Day 8 (n = 39) | Day 15 (n = 39) | Day 22 (n = 39) | Day 45 (n= 15) | Day 60 (n= 15) | |
|---|---|---|---|---|---|---|
| Retinol | 11.62 ± 4.31 | 25.93 ± 9.99 | 37.86 ± 9.60 | 42.35 ± 9.88 | 39.97 ± 7.79 | 40.05 ± 7.21 |
| (μg/100 mL) | (2.1–19.3) | (9.9–44.7) | (15.2–54.3) | (32.7–56.6) | (29.9–51.3) | (30.4–50.3) |
| RBP | 1.62 ± 0.71 | 3.57 ± 131 | 4.96 ± 1.59 | 5.43 ± 0.88 | 5.21 ± 1.01 | 5.17 ± 1.02 |
| (mg/100 mL) | (0.3–3.5) | (1.5–6.1) | (2.0–8.5) | (4.3–7.6) | (4.4–8.2) | (4.1–7.9) |
| TTR | 6.45 ± 1.86 | 13.03 ± 4.05 | 19.78 ± 5.25 | 22.23 ± 3.24 | 21.72 ± 3.44 | 21.63 ± 3.56 |
| (mg/100 mL) | (3.4–11.5) | (7.2–22.6) | (9.6–32.0) | (17.1–29.8) | (18.2–29.9) | (17.6–28.1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingenbleek, Y. Revisiting PINI Scoring in Light of Recent Biological Advances. Nutrients 2023, 15, 1846. https://doi.org/10.3390/nu15081846
Ingenbleek Y. Revisiting PINI Scoring in Light of Recent Biological Advances. Nutrients. 2023; 15(8):1846. https://doi.org/10.3390/nu15081846
Chicago/Turabian StyleIngenbleek, Yves. 2023. "Revisiting PINI Scoring in Light of Recent Biological Advances" Nutrients 15, no. 8: 1846. https://doi.org/10.3390/nu15081846
APA StyleIngenbleek, Y. (2023). Revisiting PINI Scoring in Light of Recent Biological Advances. Nutrients, 15(8), 1846. https://doi.org/10.3390/nu15081846