
Modulation of Beta-Amyloid-Activated Primary Human Neutrophils by Dietary Phenols from Virgin Olive Oil

 ,
,  , , , , and
, , , , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical Characterization of PF
2.3. Blood Collection and Neutrophil Isolation
2.4. Cytotoxicity Assay
2.5. Cell Activation and Exposure to PF
2.6. Flow Cytometry
2.7. RNA Isolation and qRT-PCR Analysis
2.8. Quantification of Cytokine Levels
2.9. Statistical Analysis
3. Results and Discussion
3.1. Characterization of the Phenolic Fraction
3.2. Effect of PF and HTyr on Neutrophil Viability
3.3. Regulation of Myeloperoxidase, Neutrophil Elastase, and Cyclooxygenase-2 Gene Expression
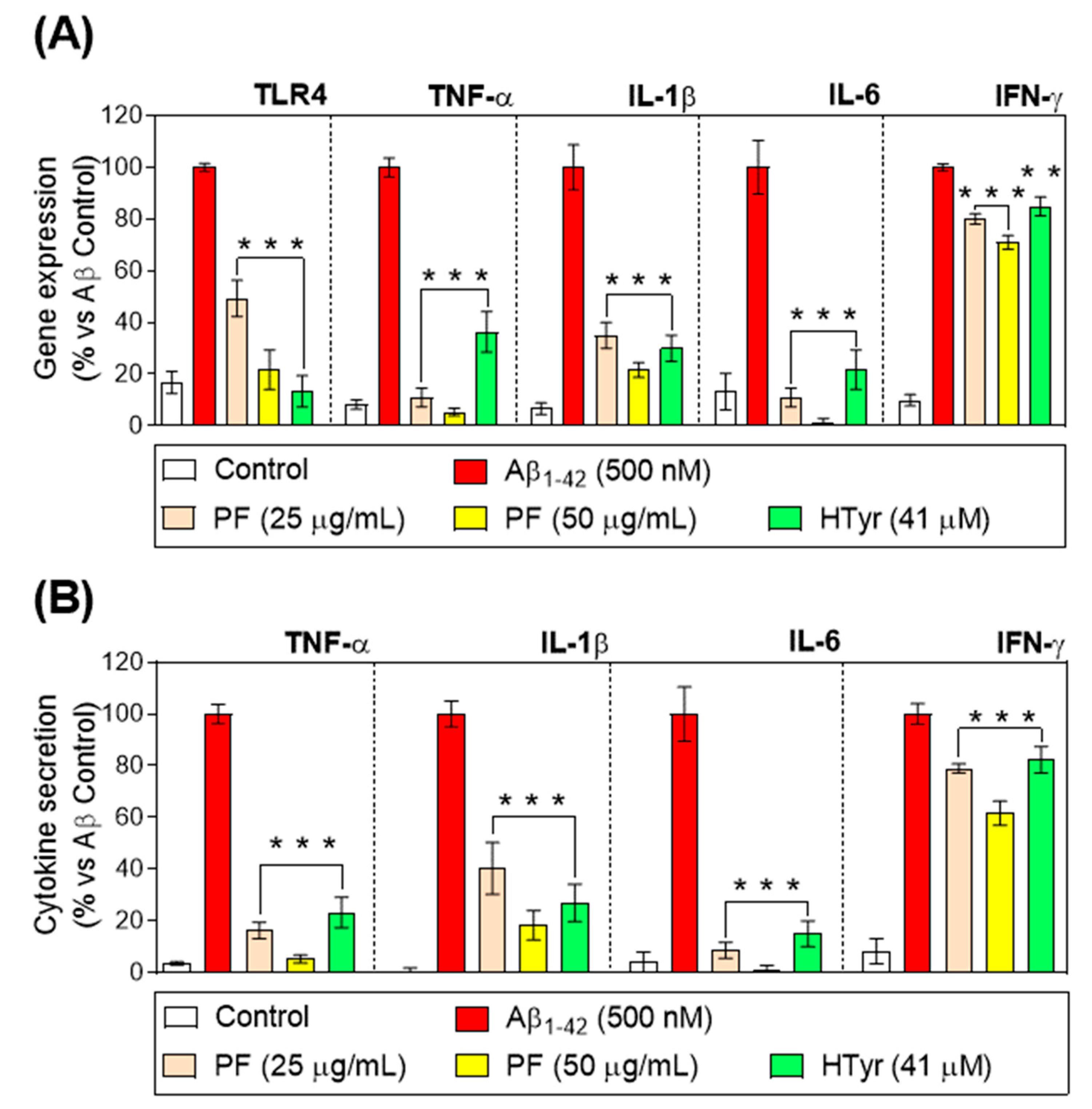
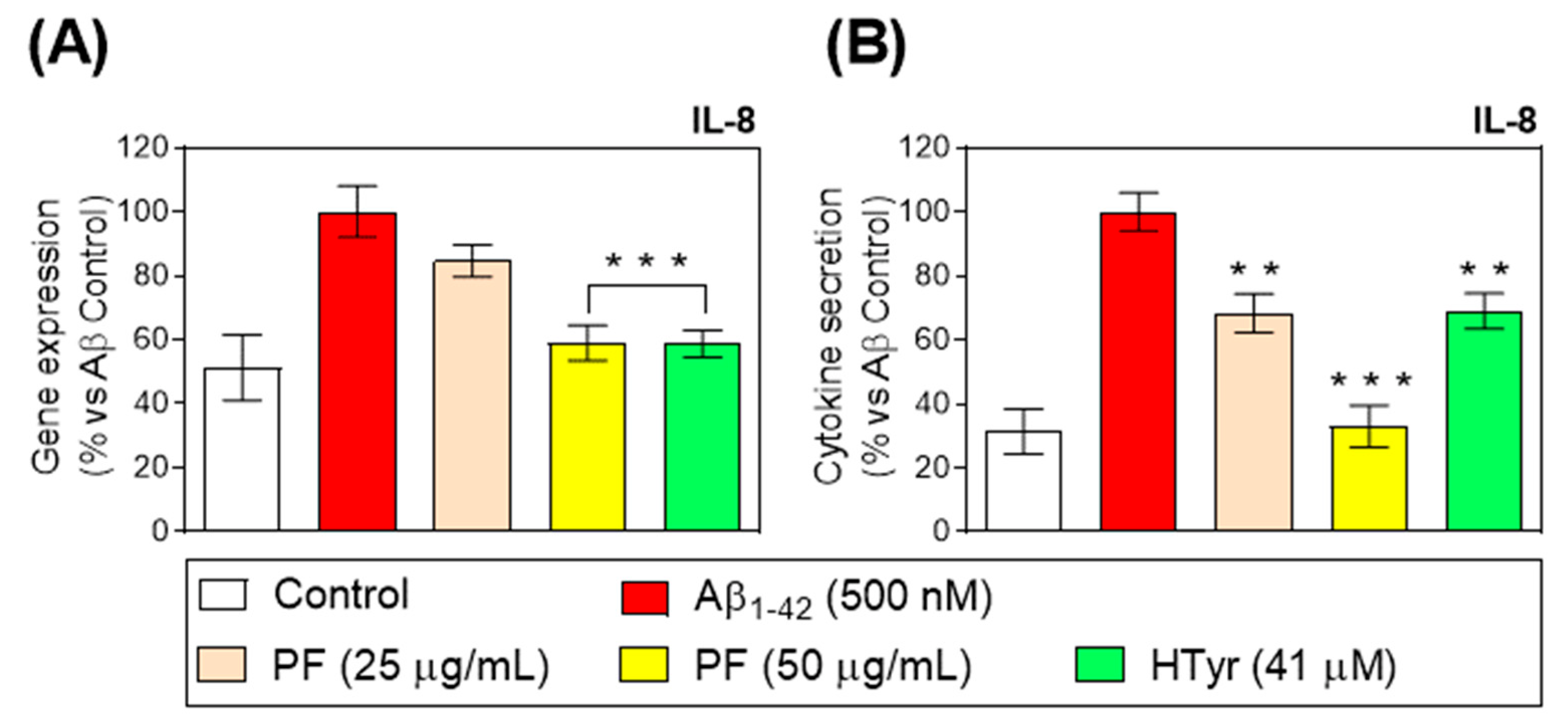
3.4. Down-Regulation of Toll-like Receptor 4 and Pro-Inflammatory Cytokine Gene Expression and Secretion
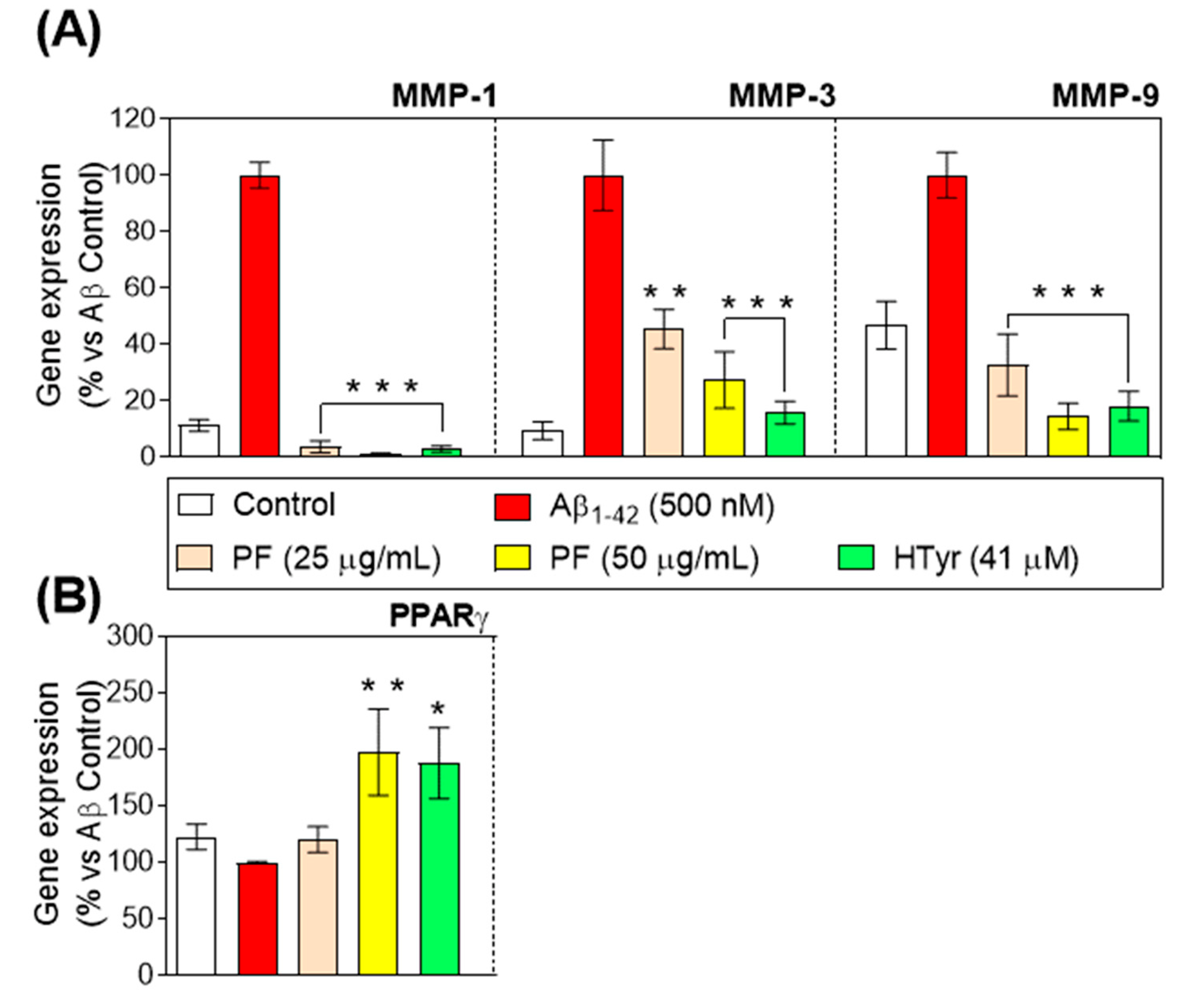
3.5. Regulation of Metalloproteinases and Peroxisome Proliferator-Activated Receptor-γ Gene Expression
3.6. Olive Oil Phenols Prevent Activation of Human Neutrophils
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Javier Basterra-Gortari, F.; Ruiz-Canela, M.; Martínez-González, M.A.; Babio, N.; Sorlí, J.V.; Fito, M.; Ros, E.; Gómez-Gracia, E.; Fiol, M.; Lapetra, J.; et al. Effects of a Mediterranean eating plan on the need for glucose-lowering medications in participants with type 2 diabetes: A subgroup analysis of the PREDIMED trial. Diabetes Care 2019, 42, 1390–1397. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean diet and health status: Active ingredients and pharmacological mechanisms. Br. J. Pharmacol. 2020, 177, 1241–1257. [Google Scholar] [CrossRef]
- Martin, M.E.; Millan-Linares, M.C.; Naranjo, M.C.; Toscano, R.; Abia, R.; Muriana, F.J.G.; Bermudez, B.; Montserrat-de la Paz, S. Minor compounds from virgin olive oil attenuate LPS-induced inflammation via visfatin-related gene modulation on primary human monocytes. J. Food Biochem. 2019, 43, e12941. [Google Scholar] [CrossRef]
- Cárdeno, A.; Aparicio-Soto, M.; Montserrat-de la Paz, S.; Bermudez, B.; Muriana, F.J.G.; Alarcón-de-la-Lastra, C. Squalene targets pro- and anti-inflammatory mediators and pathways to modulate over-activation of neutrophils, monocytes and macrophages. J. Funct. Foods 2015, 14, 779–790. [Google Scholar] [CrossRef]
- Reboredo-Rodríguez, P.; Varela-López, A.; Forbes-Hernández, T.Y.; Gasparrini, M.; Afrin, S.; Cianciosi, D.; Zhang, J.; Manna, P.P.; Bompadre, S.; Quiles, J.L.; et al. Phenolic compounds isolated from olive oil as nutraceutical tools for the prevention and management of cancer and cardiovascular diseases. Int. J. Mol. Sci. 2018, 19, 2305. [Google Scholar] [CrossRef] [PubMed]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Jiménez, J.; Berraquero-García, C.; Pérez-Gálvez, R.; García-Moreno, P.J.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Peptides and protein hydrolysates exhibiting anti- inflammatory activity: Sources, structural features and modulation mechanisms. Food Funct. 2022, 13, 12510–12540. [Google Scholar] [CrossRef]
- Qu, X.; Tang, Y.; Hua, S. Immunological approaches towards cancer and inflammation: A cross talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-de la Paz, S.; Carrillo-Berdasco, G.; Rivero-Pino, F.; Villanueva-Lazo, A.; Millan-Linares, M.C. Hemp Protein Hydrolysates Modulate Inflammasome-Related Genes in Microglial Cells. Biology 2023, 12, 49. [Google Scholar] [CrossRef]
- Schiller, M.; Ben-Shaanan, T.L.; Rolls, A. Neuronal regulation of immunity: Why, how and where? Nat. Rev. Immunol. 2021, 21, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Soto, M.; Sánchez-Hidalgo, M.; Rosillo, M.Á.; Castejón, M.L.; Alarcón-De-La-Lastra, C. Extra virgin olive oil: A key functional food for prevention of immune-inflammatory diseases. Food Funct. 2016, 7, 4492–4505. [Google Scholar] [CrossRef]
- Vazquez Roncero, A.; Janet del Valle, M.; Janet del Valle, L. Componentes fenolicos de la aceituna. Grasas Aceites 1976, 27. [Google Scholar]
- Cárdeno, A.; Sánchez-Hidalgo, M.; Aparicio-Soto, M.; Sánchez-Fidalgo, S.; Alarcón-De-La-Lastra, C. Extra virgin olive oil polyphenolic extracts downregulate inflammatory responses in LPS-activated murine peritoneal macrophages suppressing NFκB and MAPK signalling pathways. Food Funct. 2014, 5, 1270–1277. [Google Scholar] [CrossRef]
- Miho, H.; Moral, J.; Barranco, D.; Ledesma-Escobar, C.A.; Priego-Capote, F.; Díez, C.M. Influence of genetic and interannual factors on the phenolic profiles of virgin olive oils. Food Chem. 2021, 342, 128357. [Google Scholar] [CrossRef]
- Miho, H.; Moral, J.; López-González, M.A.; Díez, C.M.; Priego-Capote, F. The phenolic profile of virgin olive oil is influenced by malaxation conditions and determines the oxidative stability. Food Chem. 2020, 314, 126183. [Google Scholar] [CrossRef]
- Miho, H.; Díez, C.M.; Mena-Bravo, A.; Sánchez de Medina, V.; Moral, J.; Melliou, E.; Magiatis, P.; Rallo, L.; Barranco, D.; Priego-Capote, F. Cultivar influence on variability in olive oil phenolic profiles determined through an extensive germplasm survey. Food Chem. 2018, 266, 192–199. [Google Scholar] [CrossRef]
- Bucciantini, M.; Leri, M.; Nardiello, P.; Casamenti, F.; Stefani, M. Olive Polyphenols: Antioxidant and Anti-Inflammatory Properties. Antioxidants 2021, 10, 1044. [Google Scholar] [CrossRef] [PubMed]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef]
- Beauchamp, G.K.; Keast, R.S.J.; Morel, D.; Lin, J.; Pika, J.; Han, Q.; Lee, C.-H.; Smith, A.B.; Breslin, P.A.S. Phytochemistry: Ibuprofen-like activity in extra-virgin olive oil. Nature 2005, 437, 45–46. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Narumiya, S. Prostaglandin-cytokine crosstalk in chronic inflammation. Br. J. Pharmacol. 2019, 176, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Glennon-Alty, L.; Hackett, A.P.; Chapman, E.A.; Wright, H.L. Neutrophils and redox stress in the pathogenesis of autoimmune disease. Free Radic. Biol. Med. 2018, 125, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.J.; Kasus-Jacobi, A.; Pereira, H.A. The role of neutrophil granule proteins in neuroinflammation and Alzheimer’s disease. J. Neuroinflamm. 2018, 15, 1–15. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Kiss, A.K.; Naruszewicz, M. Inhibition of human neutrophils NEP activity, CD11b/CD18 expression and elastase release by 3,4-dihydroxyphenylethanol-elenolic acid dialdehyde, oleacein. Food Chem. 2014, 153, 1–8. [Google Scholar] [CrossRef]
- Montoya, T.; Sánchez-Hidalgo, M.; Castejón, M.L.; Rosillo, M.Á.; González-Benjumea, A.; Alarcón-De-la-lastra, C. Dietary oleocanthal supplementation prevents inflammation and oxidative stress in collagen-induced arthritis in mice. Antioxidants 2021, 10, 650. [Google Scholar] [CrossRef]
- Chen, L.; Yu, J. Modulation of Toll-Like Receptor Signaling in Innate Immunity by Natural Products. Int. Immunopharmacol. 2016, 37, 65–70. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Schorr, W. Role of interleukin-8 in neutophil signaling. Curr. Opin. Hematol. 2000, 7, 178–182. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Kobayashi, Y. The regulatory role of nitric oxide in proinflammatory cytokine expression during the induction and resolution of inflammation. J. Leukoc. Biol. 2010, 88, 1157–1162. [Google Scholar] [CrossRef]
- Alblihed, M.A. Hydroxytyrosol ameliorates oxidative challenge and inflammatory response associated with lipopolysaccharide-mediated sepsis in mice. Hum. Exp. Toxicol. 2021, 40, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Carluccio, M.A.; Martinelli, R.; Massaro, M.; Calabriso, N.; Scoditti, E.; Maffia, M.; Verri, T.; Gatta, V.; Caterina, R. De Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis. Nutrients 2021, 13, 3990. [Google Scholar] [CrossRef] [PubMed]
- Khokha, R.; Murthy, A.; Weiss, A. Metalloproteinases and their natural inhibitors in inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Youssef, J.; Badr, M. Role of peroxisome proliferator-activated receptors in inflammation control. J. Biomed. Biotechnol. 2004, 12, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.-C.; Zhou, M.; Shankavaram, U.; Peng, G.; Wahl, L.M. Differential Regulation of Lipopolysaccharide-Induced Monocyte Matrix Metalloproteinase (MMP)-1 and MMP-9 by p38 and Extracellular Signal-Regulated Kinase 1/2 Mitogen-Activated Protein Kinases. J. Immunol. 2003, 170, 6244–6249. [Google Scholar] [CrossRef] [PubMed]
- Löffek, S.; Schilling, O.; Franzke, C.W. Series “matrix metalloproteinases in lung health and disease” edited by J. Müller-Quernheim and O. Eickelberg number 1 in this series: Biological role of matrix metalloproteinases: A critical balance. Eur. Respir. J. 2011, 38, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Nissinen, L.; Kähäri, V.M. Matrix metalloproteinases in inflammation. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Scoditti, E.; Nestola, A.; Massaro, M.; Calabriso, N.; Storelli, C.; De Caterina, R.; Carluccio, M.A. Hydroxytyrosol suppresses MMP-9 and COX-2 activity and expression in activated human monocytes via PKCα and PKCβ1 inhibition. Atherosclerosis 2014, 232, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPAR γ and the Innate Immune System Mediate the Resolution of Inflammation. PPAR Res. 2015, 2015, e549691. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Selders, G.S.; Fetz, A.E.; Radic, M.Z.; Bowlin, G.L. An overview of the role of neutrophils in innate immunity, inflammation and host-biomaterial integration. Regen. Biomater. 2017, 4, 55–68. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Newton, R.; Bahaie, N.S.; Long, C.; Walcheck, B. ADAM17 cleaves CD16b (FcγRIIIb) in human neutrophils. Biochim. Biophys. Acta 2013, 1833, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ivetic, A. A head-to-tail view of L-selectin and its impact on neutrophil behaviour. Cell Tissue Res. 2018, 371, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Westra, J.; Rutgers, A.; Doornbos-Van der Meer, B.; Huitema, M.G.; Stegeman, C.A.; Abdulahad, W.H.; Satchell, S.C.; Mathieson, P.W.; Heeringa, P.; et al. Decreased CXCR1 and CXCR2 expression on neutrophils in anti-neutrophil cytoplasmic autoantibody-associated vasculitides potentially increases neutrophil adhesion and impairs migration. Arthritis Res. Ther. 2011, 13, R201. [Google Scholar] [CrossRef]
- Källquist, L.; Hansson, M.; Persson, A.M.; Janssen, H.; Calafat, J.; Tapper, H.; Olsson, I. The tetraspanin CD63 is involved in granule targeting of neutrophil elastase. Blood 2008, 112, 3444–3454. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.M.; Carruba, G.; Cicero, A.F.G.; Banach, M.; Nikolic, D.; Giglio, R.V.; Terranova, A.; Soresi, M.; Giannitrapani, L.; Montalto, G.; et al. Daily use of extra virgin olive oil with high oleocanthal concentration reduced body weight, waist circumference, alanine transaminase, inflammatory cytokines and hepatic steatosis in subjects with the metabolic syndrome: A 2-month intervention study. Metabolites 2020, 10, 392. [Google Scholar] [CrossRef]
- Noce, A.; Marrone, G.; Urciuoli, S.; Di Daniele, F.; Di Lauro, M.; Zaitseva, A.P.; Di Daniele, N.; Romani, A. Usefulness of extra virgin olive oil minor polar compounds in the management of chronic kidney disease patients. Nutrients 2021, 13, 581. [Google Scholar] [CrossRef] [PubMed]
- De Santis, S.; Clodoveo, M.L.; Corbo, F. Correlation between Chemical Characterization and Biological Activity: An Urgent Need for Human Studies Using Extra Virgin Olive Oil. Antioxidants 2022, 11, 258. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | GenBank Accession Number | Direction | Sequence (5′ → 3′) |
|---|---|---|---|
| TLR4 | NM_138554.4 | Forward Reverse | TGAGCAGTCGTGCTGGTATC CAGGGCTTTTCTGAGTCGTC |
| MPO | NM_000250 | Forward Reverse | CAGCCCAGATATACCCCTCA GACAACACAGGCATCACCAC |
| NE | NM_001972.4 | Forward Reverse | CATCGTGATTCTCCAGCTCA CTCACGAGAGTGCAGACGTT |
| COX-2 | NM_000963 | Forward Reverse | TCCCATGGGTTGTGTGTTTA AGATCATCTCTGCCTGAGTATCTT |
| IFNγ | NM_000619.2 | Forward Reverse | TCCCATGGGTTGTGTGTTTA AAGCACCAGGCATGAAATCT |
| IL-1β | NM_000576.2 | Forward Reverse | CTGTCCTGCGTGTTGAAAGA TTCTGCTTGAGAGGTGCTGA |
| TNF-α | NM_000594.3 | Forward Reverse | TCCTTCAGACACCCTCAACC AGGCCCCAGTTTGAATTCTT |
| IL-6 | NM_000600.4 | Forward Reverse | TACCCCCAGGAGAAGATTCC TTTTCTGCCAGTGCCTCTTT |
| IL-8 | NM_000584 | Forward Reverse | TAGCAAAATTGAGGCCAAGG AAACCAAGGCACAGTGGAAC |
| MMP-1 | NM_ 001145938 | Forward Reverse | CTGCTTGACCCTCAGAGACC ATGCTGAAACCCTGAAGGTG |
| MMP-3 | NM_002422 | Forward Reverse | GAGTGTCGGAGTCCAGCTTC GCAGTTTGCTCAGCCTATCC |
| MMP-9 | NM_004994 | Forward Reverse | CAGGGATCTCCCCTCCTTAG GTCTTGTGGAGGCTTTGAGC |
| PPARγ | NM_005037 | Forward Reverse | GCTGTGCAGGAGATCACAGA GGGCTCCATAAAGTCACCAA |
| GAPDH | NM_001289745.2 | Forward Reverse | ACAGTCAGCCGCATCTTCTT ACGACCAAATCCGTTGACTC |
| HPRT | NM_002046.6 | Forward Reverse | GAGTCAACGGATTTGGTCGT GACAAGCTTCCCGTTCTCAG |
| Phenol Composition | µM Phenol (50 µg PF/mL) |
|---|---|
| Hydroxytyrosol | 41.07 |
| Tyrosol | 43.09 |
| Vanillic acid | 5.09 |
| p-Coumaric acid | 3.42 |
| Decarboxymethyl oleuropein aglycone (dialdehyde) = oleacein | 10.27 |
| Tyrosol acetate | 4.97 |
| Decarboxymethyl ligstroside aglycone (dialdehyde) = oleocanthal | 11.33 |
| Pinoresinol | 6.21 |
| Cinnamic acid | 6.89 |
| Acetoxy-pinoresinol | 6.22 |
| Oleuropein aglycone, oxidized aldehyde form | 39.18 |
| Ligstroside aglycone, dialdehyde form | 26.20 |
| Luteolin | 4.18 |
| Apigenin | 0.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivero-Pino, F.; Grao-Cruces, E.; Lopez-Enriquez, S.; Alba, G.; Marquez-Paradas, E.; Claro-Cala, C.M.; Santa-Maria, C.; Montserrat-de la Paz, S. Modulation of Beta-Amyloid-Activated Primary Human Neutrophils by Dietary Phenols from Virgin Olive Oil. Nutrients 2023, 15, 941. https://doi.org/10.3390/nu15040941
Rivero-Pino F, Grao-Cruces E, Lopez-Enriquez S, Alba G, Marquez-Paradas E, Claro-Cala CM, Santa-Maria C, Montserrat-de la Paz S. Modulation of Beta-Amyloid-Activated Primary Human Neutrophils by Dietary Phenols from Virgin Olive Oil. Nutrients. 2023; 15(4):941. https://doi.org/10.3390/nu15040941
Chicago/Turabian StyleRivero-Pino, Fernando, Elena Grao-Cruces, Soledad Lopez-Enriquez, Gonzalo Alba, Elvira Marquez-Paradas, Carmen M. Claro-Cala, Consuelo Santa-Maria, and Sergio Montserrat-de la Paz. 2023. "Modulation of Beta-Amyloid-Activated Primary Human Neutrophils by Dietary Phenols from Virgin Olive Oil" Nutrients 15, no. 4: 941. https://doi.org/10.3390/nu15040941
APA StyleRivero-Pino, F., Grao-Cruces, E., Lopez-Enriquez, S., Alba, G., Marquez-Paradas, E., Claro-Cala, C. M., Santa-Maria, C., & Montserrat-de la Paz, S. (2023). Modulation of Beta-Amyloid-Activated Primary Human Neutrophils by Dietary Phenols from Virgin Olive Oil. Nutrients, 15(4), 941. https://doi.org/10.3390/nu15040941